Aryl Hydrocarbon Receptor and Dioxin-Related Health Hazards—Lessons from Yusho


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
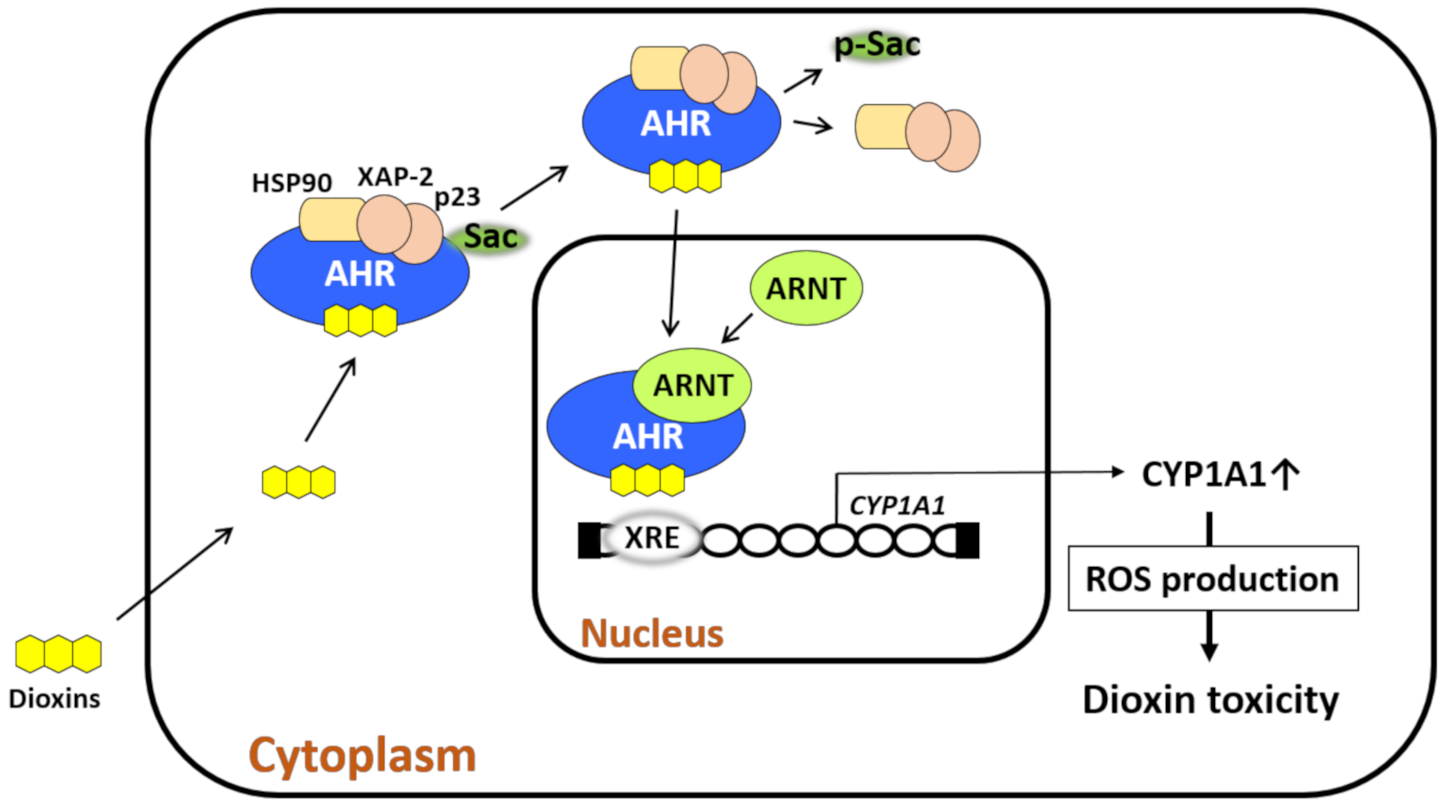
3. AHR and Its Biological Function
4. AHR and Clinical Symptoms in Yusho
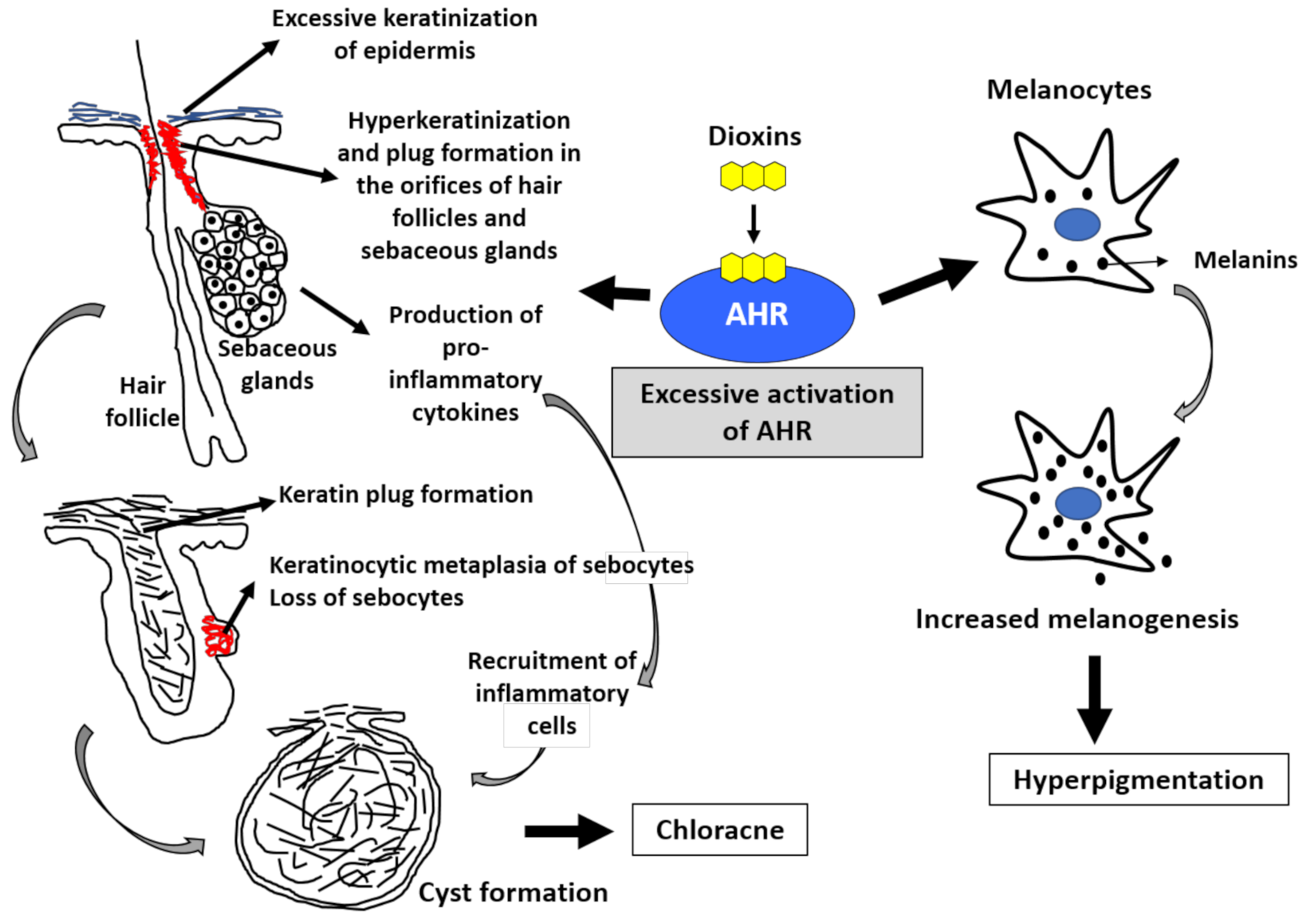
4.1. Chloracne and Hyperpigmentation
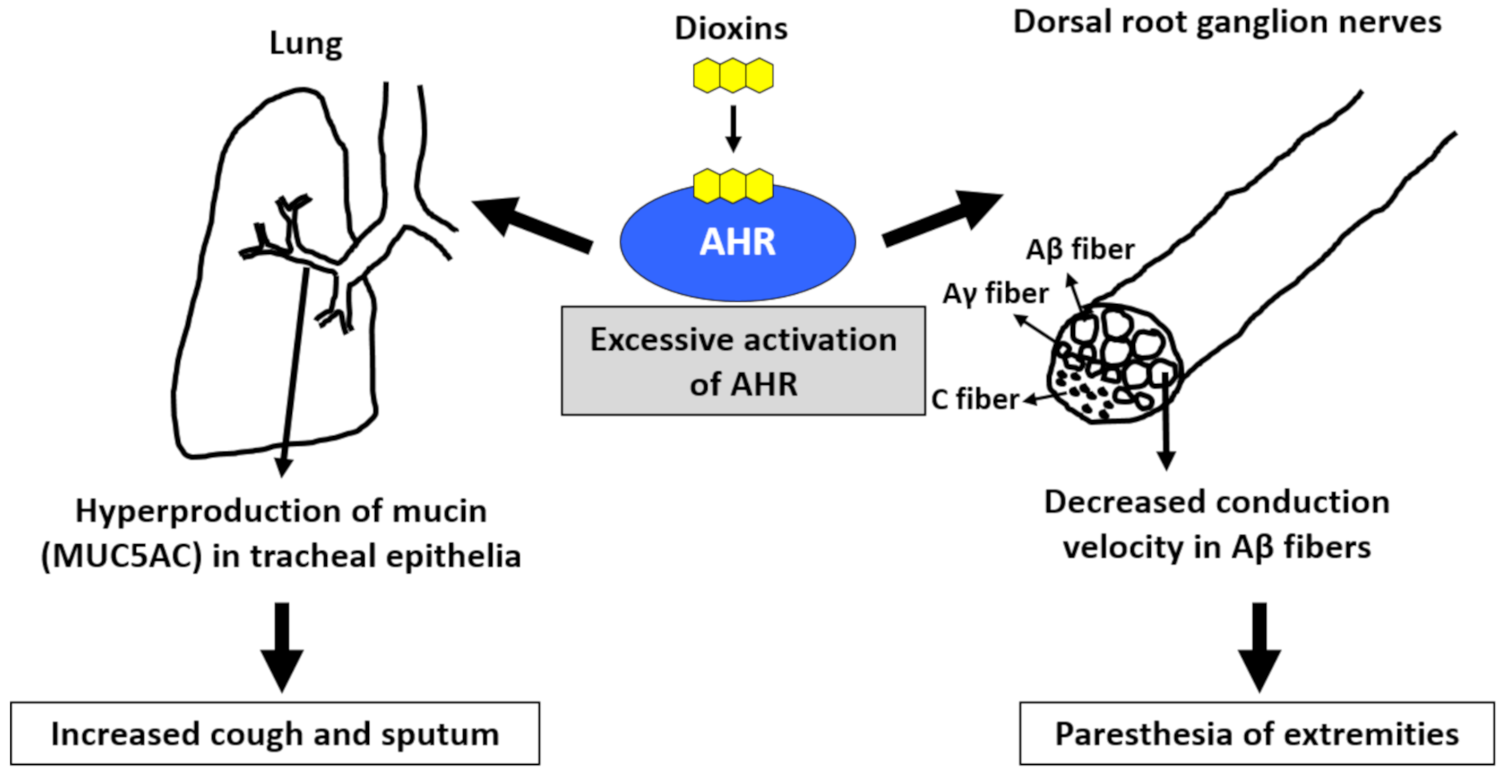
4.2. Cough and Sputum
4.3. Paresthesia, Pain, and Numbness of the Extremities
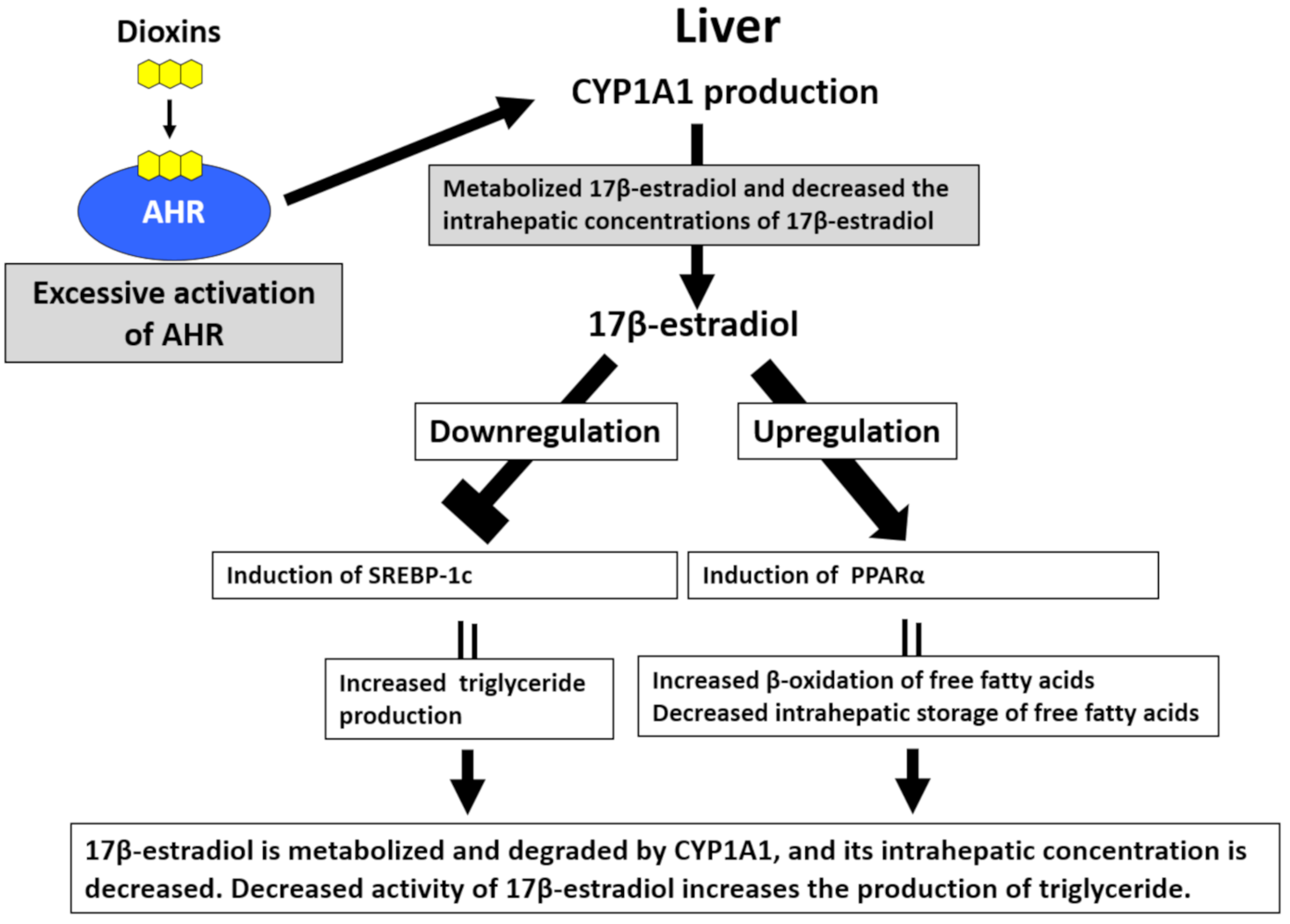
4.4. Blood Triglyceride Levels
4.5. Sleep Disturbance
4.6. Arthralgia and Brusitis
4.7. Immune Response
4.8. Mortality Rate and Carcinogenesis
4.9. Perinatal Abnormalities and the Impact in Offspring
4.10. Perinatal Abnormalities and Impact on Future Generations in Animal Models
5. High Blood Concentrations of Dioxins Are Sustained in Patients with Yusho
6. Therapeutic Approach in Yusho
6.1. Trials to Enhance Dioxin Excretion
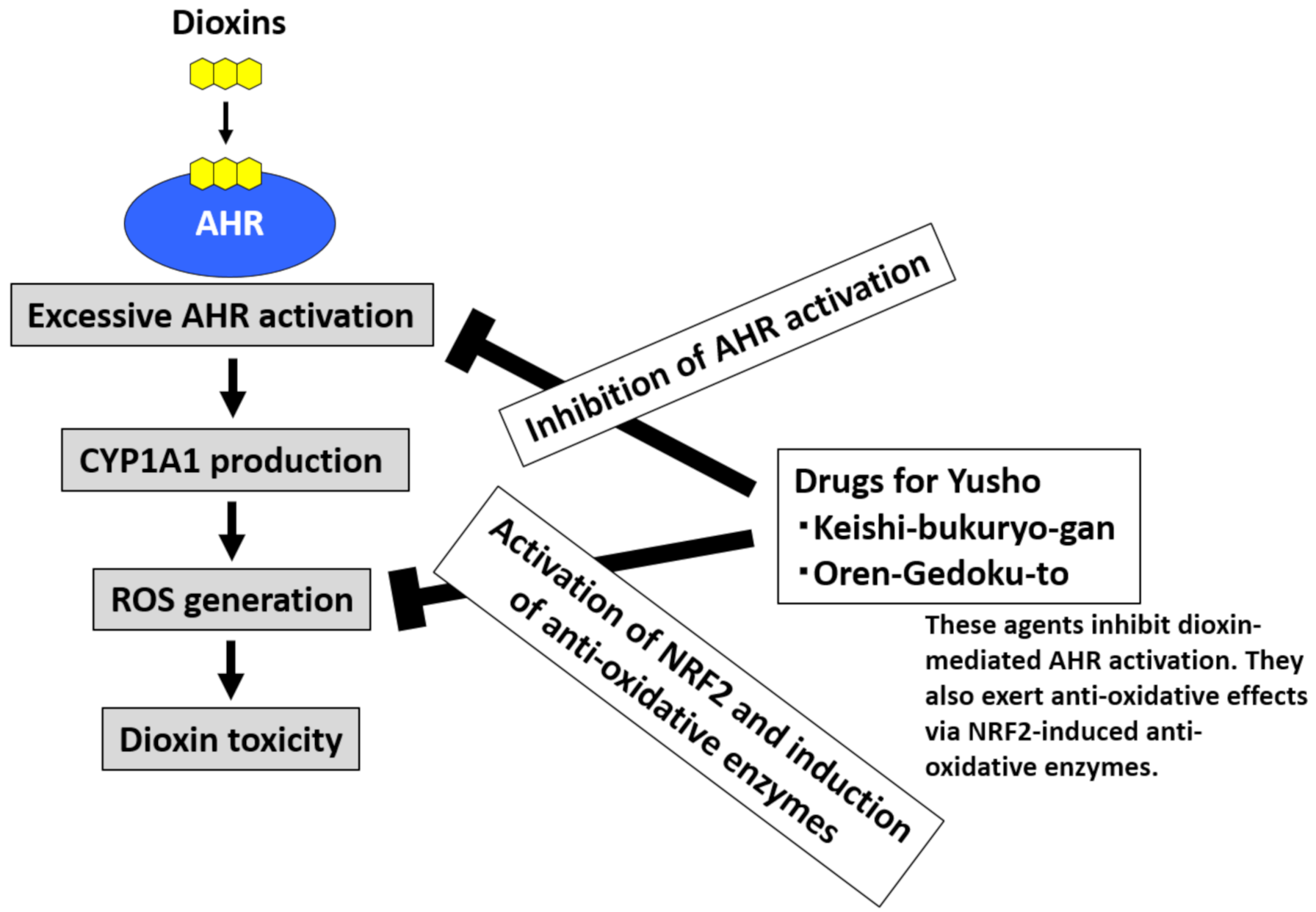
6.2. Agents to Minimize Dioxin Toxicity
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Masuda, Y.; Yoshimura, H. Chemical analysis and toxicity of polychlorinated biphenyls and dibenzofurans in relation to yusho. J. Toxicol. Sci. 1982, 7, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Yoshimura, H. Polychlorinated biphenyls and dibenzofurans in patients with yusho and their toxicological significance: A review. Am. J. Ind. Med. 1984, 5, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, T.; Sugihara, M.; Hirakawa, S.; Hori, T.; Kajiwara, J.; Katsuki, S.; Mitoma, C.; Furue, M. Compound-specific isotopic and congener-specific analyses of polychlorinated biphenyl in the heat medium and rice oil of the Yusho incident. Environ. Sci. Pollut. Res. Int. 2018, 25, 16464–16471. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs), and related compounds: Environmental and mechanistic considerations which support the development of toxic equivalency factors (TEFs). Crit. Rev. Toxicol. 1990, 21, 51–88. [Google Scholar] [CrossRef]
- van den Berg, M.; Birnbaum, L.S.; Denison, M.; De Vito, M.; Farland, W.; Feeley, M.; Fiedler, H.; Hakansson, H.; Hanberg, A.; Haws, L.; et al. The 2005 World Health Organization reevaluation of human and mammalian toxic equivalencyfactors for dioxins and dioxin-like compounds. Toxicol. Sci. 2006, 93, 223–241. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Takasuga, T.; Masunaga, S.; Nakanishi, J. Detailed study on the levels of polychlorinated dibenzo-p-dioxins, polychlorinated dibenzofurans and polychlorinated biphenyls in Yusho rice oil. Chemosphere 2002, 46, 1461–1469. [Google Scholar] [CrossRef]
- Iida, T.; Todaka, T. Measurement of dioxins in human blood: Improvement of analytical method. Ind. Health 2003, 41, 197–204. [Google Scholar] [CrossRef]
- Kuratsune, M.; Yoshimura, T.; Matsuzaka, J.; Yamaguchi, A. Yusho, a poisoning caused by rice oil contaminated with polychlorinated biphenyls. HSMHA Health Rep. 1971, 86, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Uenotsuchi, T.; Urabe, K.; Ishikawa, T.; Kuwabata, M. Overview of Yusho. J. Dermatol. Sci. 2005, 1, S3–S10. [Google Scholar] [CrossRef]
- Mitoma, C.; Mine, Y.; Utani, A.; Imafuku, S.; Muto, M.; Akimoto, T.; Kanekura, T.; Furue, M.; Uchi, H. Current skin symptoms of Yusho patients exposed to high levels of 2,3,4,7,8-pentachlorinated dibenzofuran and polychlorinated biphenyls in 1968. Chemosphere 2015, 137, 45–51. [Google Scholar] [CrossRef]
- Imamura, T.; Matsumoto, S.; Kanagawa, Y.; Tajima, B.; Matsuya, S.; Furue, M.; Oyama, H. A technique for identifying three diagnostic findings using association analysis. Med. Biol. Eng. Comput. 2007, 45, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Kanagawa, Y.; Koike, S.; Akahane, M.; Uchi, H.; Shibata, S.; Furue, M.; Imamura, T. Twenty-year changes of penta-chlorodibenzofuran (PeCDF) level and symptoms in Yusho patients, using association analysis. BMC Res. Notes 2010, 3, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, Y.; Kataoka, K.; Tokunaga, S.; Hirohata, T.; Shinohara, S.; Tokiwa, H. Association between blood polychlorinated biphenyl concentration and serum triglyceride level in chronic “Yusho” (polychlorinated biphenyl poisoning) patients. Int. Arch. Occup. Environ. Health 1993, 65, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Kyushu University, Graduate School of Medical Sciences, Department of Dermatology. Available online: https://www.kyudai-derm.org/part/yusho/index.html (accessed on 30 November 2020).
- Masuda, Y. Toxic effects of PCB/PCDF to human observed in Yusho and other poisonings. Fukuoka Igaku Zasshi. 2009, 100, 141–155. [Google Scholar] [PubMed]
- Uchi, H. Current status of Yusho and treatment of chronic dioxin poisoning. Nishinihon J. Dermatol. 2014, 76, 535–539. [Google Scholar] [CrossRef]
- Mitoma, C.; Uchi, H.; Tsuji, G.; Koga, S.; Furue, M. Latest findings of Yusho and its therapeutic development. Fukuoka Igaku Zasshi. 2015, 106, 184–194. [Google Scholar] [PubMed]
- Mitoma, C.; Uchi, H.; Tsukimori, K.; Yamada, H.; Akahane, M.; Imamura, T.; Utani, A.; Furue, M. Yusho and its latest findings-A review in studies conducted by the Yusho Group. Environ. Int. 2015, 82, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, C.; Uchi, H.; Tsukimori, K.; Todaka, T.; Kajiwara, J.; Shimose, T.; Akahane, M.; Imamura, T.; Furue, M. Current state of yusho and prospects for therapeutic strategies. Environ. Sci. Pollut. Res. Int. 2018, 25, 16472–16480. [Google Scholar] [CrossRef]
- Furue, M.; Takahara, M.; Nakahara, T.; Uchi, H. Role of AhR/ARNT systemin skin homeostasis. Arch. Dermatol. Res. 2014, 306, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, M.; Uchi, H.; Mitoma, C.; Hashimoto-Hachiya, A.; Chiba, T.; Ito, T.; Nakahara, T.; Tsuji, G. Antioxidants for Healthy Skin: The emerging role of aryl hydrocarbon receptors and nuclear factor-erythroid 2-related factor-2. Nutrients 2017, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Sugihara, K.; Watanabe, Y.; Yamaji, S.; Kitamura, S.; Ohta, S. Aryl hydrocarbon receptor ligand activity of extracts from 62 herbal medicines and effect on cytochrome P450 activity. Yakugaku Zasshi 2015, 135, 1185–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enan, E.; Matsumura, F. Identification of c-Src as the integral component of the cytosolic Ah receptor complex, transducing the signal of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) through the protein phosphorylation pathway. Biochem. Pharmacol. 1996, 52, 1599–1612. [Google Scholar] [CrossRef]
- Mimura, J.; Fujii-Kuriyama, Y. Functional role of AhR in the expression of toxic effects by TCDD. Biochim. Biophys. Acta. 2003, 1619, 263–268. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ito, T.; Tsuji, G.; Furue, M. Baicalein inhibits benzo[a]pyrene-induced toxic response by downregulating Src phosphorylation and by upregulating NRF2-HMOX1 system. Antioxidants (Basel) 2020, 9, 507. [Google Scholar] [CrossRef]
- Warner, J.; Osuch, J.R.; Karmaus, W.; Landgraf, J.R.; Taffe, B.; O’Keefe, M.; Mikucki, D.; Haan, P. Common classification schemes for PCB congeners and the gene expression of CYP17, CYP19, ESR1 and ESR2. Sci. Total Environ. 2012, 414, 81–89. [Google Scholar] [CrossRef]
- Kennedy, L.H.; Sutter, C.H.; Leon Carrion, S.; Tran, Q.T.; Bodreddigari, S.; Kensicki, E.; Mohney, R.P.; Sutter, T.R. 2,3,7,8-Tetrachlorodibenzo-p-dioxin-mediated production of reactive oxygen species is an essential step in the mechanism of action to accelerate human keratinocyte differentiation. Toxicol. Sci. 2013, 132, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Stockinger, B.; Di Meglio, P.; Gialitakis, M.; Duarte, J.H. The aryl hydrocarbon receptor: Multitasking in the immune system. Annu. Rev. Immunol. 2014, 32, 403–432. [Google Scholar] [CrossRef]
- Fritsche, E.; Schäfer, C.; Calles, C.; Bernsmann, T.; Bernshausen, T.; Wurm, M.; Hübenthal, U.; Cline, J.E.; Hajimiragha, H.; Schroeder, P.; et al. Lightening up the UV response by identification of the arylhydrocarbon receptor as a cytoplasmatic target for ultraviolet B radiation. Proc. Natl. Acad. Sci. USA 2007, 104, 8851–8856. [Google Scholar] [CrossRef] [Green Version]
- Nagayoshi, H.; Murayama, N.; Kakimoto, K.; Tsujino, M.; Takenaka, S.; Katahira, J.; Lim, Y.R.; Kim, D.; Yamazaki, H.; Komori, M.; et al. Oxidation of flavone, 5-hydroxyflavone, and 5,7-dihydroxyflavone to mono-, di-, and tri-hydroxyflavones by human cytochrome P450 enzymes. Chem. Res. Toxicol. 2019, 32, 1268–1280. [Google Scholar] [CrossRef]
- Androutsopoulos, V.P.; Papakyriakou, A.; Vourloumis, D.; Spandidos, D.A. Comparative CYP1A1 and CYP1B1 substrate and inhibitor profile of dietary flavonoids. Bioorg. Med. Chem. 2011, 19, 2842–2849. [Google Scholar] [CrossRef]
- Esser, C.; Bargen, I.; Weighardt, H.; Haarmann-Stemmann, T.; Krutmann, J. Functions of the aryl hydrocarbon receptor in the skin. Semin. Immunopathol. 2013, 35, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ogawa, F.; Thiele, J.J.; Lee, J.B.; Bae, S.; Sato, S. Increased levels of urinary nitrite and nitrotyrosine in Yusho victims 40 years after accidental poisoning with polychlorinated biphenyls in Nagasaki, Japan. J. Appl. Toxicol. 2008, 28, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ogawa, F.; Thiele, J.J.; Bae, S.; Sato, S. Lipid peroxidation is enhanced in Yusho victims 35 years after accidental poisoning with polychlorinated biphenyls in Nagasaki, Japan. J. Appl. Toxicol. 2007, 27, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Takahara, M.; Uchi, H.; Takeuchi, S.; Mitoma, C.; Moroi, Y.; Furue, M. An environmental contaminant, benzo (a) pyrene, induces oxidative stress-mediated interleukin-8 production in human keratinocytes via the aryl hydrocarbon receptor signaling pathway. J. Dermatol. Sci. 2011, 62, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Uchi, H.; Hashimoto-Hachiya, A.; Furue, M. Tryptophan photoproduct FICZ upregulates IL1A, IL1B, and IL6 expression via oxidative stress in keratinocytes. Oxid. Med. Cell Longev. 2018, 2018, 9298052. [Google Scholar] [CrossRef]
- Kopf, P.G.; Walker, M.K. 2,3,7,8-tetrachlorodibenzo-p-dioxin increases reactive oxygen species production in human endothelial cells via induction of cytochrome P4501A1. Toxicol. Appl. Pharmacol. 2010, 245, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Takao, T.; Tsunematsu, R.; Morokuma, S.; Fukushima, K.; Kobayashi, H.; Saito, T.; Furue, M.; Wake, N.; Asanoma, K. Inhibition of AHR transcription by NF1C is affected by a single-nucleotide polymorphism, and is involved in suppression of human uterine endometrial cancer. Oncogene 2013, 32, 4950–4959. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; He, X.; Szklarz, G.D.; Bi, Y.; Rojanasakul, Y.; Ma, Q. The aryl hydrocarbon receptor interacts with nuclear factor erythroid 2-related factor 2 to mediate induction of NAD(P)H:quinoneoxidoreductase 1 by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Arch. Biochem. Biophys. 2013, 537, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Cui, W.; Klaassen, C.D. Nrf2 protects against 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced oxidative injury and steatohepatitis. Toxicol. Appl. Pharmacol. 2011, 256, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Mimura, J.; Yamashita, K.; Nakamura, K.; Morita, M.; Takagi, T.N.; Nakao, K.; Ema, M.; Sogawa, K.; Yasuda, M.; Katsuki, M.; et al. Loss of teratogenic response to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in mice lacking the Ah (dioxin) receptor. Genes Cells 1997, 2, 645–654. [Google Scholar] [CrossRef]
- Shimizu, Y.; Nakatsuru, Y.; Ichinose, M.; Takahashi, Y.; Kume, H.; Mimura, J.; Fujii-Kuriyama, Y.; Ishikawa, T. Benzo[a]pyrene carcinogenicity is lost in mice lacking the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 779–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.H.; Chang, K.T.; Lu, Y.D. Polychlorinated biphenyls and polychlorinated dibenzofurans in the toxic rice-bran oil that caused PCB poisoning in Taichung. Bull. Environ. Contam. Toxicol. 1981, 26, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Caramaschi, F.; del Corno, G.; Favaretti, C.; Giambelluca, S.E.; Montesarchio, E.; Fara, G.M. Chloracne following environmental contamination by TCDD in Seveso, Italy. Int. J. Epidemiol. 1981, 10, 135–143. [Google Scholar] [CrossRef]
- Barbieri, S.; Pirovano, C.; Scarlato, G.; Tarchini, P.; Zappa, A.; Maranzana, M. Long-term effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on the peripheral nervous system. Clinical and neurophysiological controlled study on subjects with chloracne from the Seveso area. Neuroepidemiology 1988, 7, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, L.S. Dioxin and the AH Receptor: Synergy of Discovery. Curr. Opin. Toxicol. 2017, 2, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Suskind, R.R. Chloracne, “the hallmark of dioxin intoxication”. Scand. J. Work. Environ. Health 1985, 11, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Fuyuno, Y.; Mitoma, C.; Uchi, H.; Tsuji, G. Therapeutic agents with AHR inhibiting and NRF2 activating activity for managing chloracne. Antioxidants (Basel) 2018, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Tsuji, G. Chloracne and hyperpigmentation caused by exposure to hazardous aryl hydrocarbon receptor ligands. Int. J. Environ. Res. Public Health 2019, 16, 4864. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Tsuji, G.; Mitoma, C.; Nakahara, T.; Chiba, T.; Morino-Koga, S.; Uchi, H. Gene regulation of filaggrin and other skin barrier proteins via aryl hydrocarbon receptor. J. Dermatol. Sci. 2015, 80, 83–88. [Google Scholar] [CrossRef]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Uchi, H.; Tahakara, M.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T.; Furue, M. Antioxidant soybean tar Glyteer rescues T-helper-mediated downregulation of filaggrin expression via aryl hydrocarbon receptor. J. Dermatol. 2015, 42, 171–180. [Google Scholar] [CrossRef]
- Doi, K.; Mitoma, C.; Nakahara, T.; Uchi, H.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, M.; Furue, M. Antioxidant Houttuynia cordata extract upregulates filaggrin expression in an aryl hydrocarbon-dependent manner. Fukuoka Igaku Zasshi. 2014, 105, 205–213. [Google Scholar] [PubMed]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, T.; Furue, M. Galactomyces fermentation filtrate prevents T helper 2-mediated reduction of filaggrin in an aryl hydrocarbon receptor-dependent manner. Clin. Exp. Dermatol. 2015, 40, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, T.; Mitoma, C.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Uchi, H.; Yan, X.; Hachisuka, J.; Chiba, T.; Esaki, H.; et al. Antioxidant Opuntia ficus-indica extract activates AHR-NRF2 signaling and upregulates filaggrin and loricrin expression in human keratinocytes. J. Med. Food 2015, 18, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, A.; Goto, M.; Mitsui, T.; Hashimoto-Hachiya, A.; Tsuji, G.; Furue, M. Antioxidant Artemisia princeps extract enhances the expression of filaggrin and loricrin via the AHR/OVOL1 pathway. Int. J. Mol. Sci. 2017, 18, 1948. [Google Scholar] [CrossRef]
- Hashimoto-Hachiya, A.; Tsuji, G.; Murai, M.; Yan, X.; Furue, M. Upregulation of FLG, LOR, and IVL expression by Rhodiola crenulata root extract via aryl hydrocarbon receptor: Differential involvement of OVOL1. Int. J. Mol. Sci. 2018, 19, 1654. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Uchi, H.; Mitoma, C.; Hashimoto-Hachiya, A.; Tanaka, Y.; Ito, T.; Tsuji, G. Implications of tryptophan photoproduct FICZ in oxidative stress and terminal differentiation of keratinocytes. G. Ital. Dermatol. Venereol. 2019, 154, 37–41. [Google Scholar] [CrossRef]
- Loertscher, J.A.; Sattler, C.A.; Allen-Hoffmann, B.L. 2,3,7,8-Tetrachlorodibenzo-p-dioxin alters the differentiation pattern of human keratinocytes in organotypic culture. Toxicol. Appl. Pharmacol. 2001, 175, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Loertscher, J.A.; Lin, T.M.; Peterson, R.E.; Allen-Hoffmann, B.L. In utero exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin causes accelerated terminal differentiation in fetal mouse skin. Toxicol. Sci. 2002, 68, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Muenyi, C.S.; Carrion, S.L.; Jones, L.A.; Kennedy, L.H.; Slominski, A.T.; Sutter, C.H.; Sutter, T.R. Effects of in utero exposure of C57BL/6J mice to 2,3,7,8-tetrachlorodibenzo- p-dioxin on epidermal permeability barrier development and function. Environ. Health Perspect. 2014, 122, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Ju, Q.; Fimmel, S.; Hinz, N.; Stahlmann, R.; Xia, L.; Zouboulis, C.C. 2,3,7,8-Tetrachlorodibenzo-p-dioxin alters sebaceous gland cell differentiation in vitro. Exp. Dermatol. 2011, 20, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Forrester, A.R.; Elias, M.S.; Woodward, E.L.; Graham, M.; Williams, F.M.; Reynolds, N.J. Induction of a chloracne phenotype in an epidermal equivalent model by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) is dependent on aryl hydrocarbon receptor activation and is not reproduced by aryl hydrocarbon receptor knock down. J. Dermatol. Sci. 2014, 73, 10–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, S.; Ozawa, N.; Yoshimura, H.; Masuda, Y.; Yamaryo, T.; Kuroki, H.; Murai, K.; Aakagi, K.; Yamanaka, M.; Omae, T.; et al. Preliminary studies on the experimental PCB poisoning in Rhesus Monkeys. Fukuoka Igaku Zasshi 1979, 70, 135–172. [Google Scholar] [PubMed]
- Mitamura, Y.; Nunomura, S.; Nanri, Y.; Ogawa, M.; Yoshihara, T.; Masuoka, M.; Tsuji, G.; Nakahara, T.; Hashimoto-Hachiya, A.; Conway, S.J.; et al. The IL-13/periostin/IL-24 pathway causes epidermal barrier dysfunction in allergic skin inflammation. Allergy 2018, 73, 1881–1891. [Google Scholar] [CrossRef]
- Liu, G.; Asanoma, K.; Takao, T.; Tsukimori, K.; Uchi, H.; Furue, M.; Kato, K.; Wake, N. Aryl hydrocarbon receptor SNP -130 C/T associates with dioxins susceptibility through regulating its receptor activity and downstream effectors including interleukin. Toxicol. Lett. 2015, 232, 384–392. [Google Scholar] [CrossRef]
- Vu, Y.H.; Hashimoto-Hachiya, A.; Takemura, M.; Yumine, A.; Mitamura, Y.; Nakahara, T.; Furue, M.; Tsuji, G. IL-24 negatively regulates keratinocyte differentiation induced by aryl hydrocarbon receptor activation: Implication in the treatment of atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 9412. [Google Scholar] [CrossRef]
- Peng, F.; Tsuji, G.; Zhang, J.Z.; Chen, Z.; Furue, M. Potential role of PM2.5 in melanogenesis. Environ. Int. 2019, 132, 105063. [Google Scholar] [CrossRef]
- Schikowski, T.; Krutmann, J. Air pollution (particulate matter and nitrogen dioxide) and skin aging. Hautarzt 2019, 70, 158–162. [Google Scholar] [CrossRef]
- Imamura, T.; Kanagawa, Y.; Matsumoto, S.; Tajima, B.; Uenotsuchi, T.; Shibata, S.; Furue, M. Relationship between clinical features and blood levels of pentachlorodibenzofuran in patients with Yusho. Environ. Toxicol. 2007, 22, 124–131. [Google Scholar] [CrossRef]
- Kanagawa, Y.; Matsumoto, S.; Koike, S.; Tajima, B.; Fukiwake, N.; Shibata, S.; Uchi, H.; Furue, M.; Imamura, T. Association of clinical findings in Yusho patients with serum concentrations of polychlorinated biphenyls, polychlorinated quarterphenyls and 2,3,4,7,8-pentachlorodibenzofuran more than 30 years after the poisoning event. Environ. Health 2008, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Imamura, T.; Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Koike, S.; Tajima, B.; Matsuya, S.; Uchi, H.; Shibata, S.; Furue, M. Cutaneous symptoms such as acneform eruption and pigmentation are closely associated with blood levels of 2,3,4,7,8-penta-chlorodibenzofurans in Yusho patients, using data mining analysis. BMC Res. Notes 2009, 2, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akahane, M.; Matsumoto, S.; Kanagawa, Y.; Mitoma, C.; Uchi, H.; Yoshimura, T.; Furue, M.; Imamura, T. Long-term health effects of PCBs and related compounds: A comparative analysis of patients suffering from Yusho and the general population. Arch. Environ. Contam. Toxicol. 2018, 74, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, J.; Todaka, T.; Hirakawa, H.; Hori, T.; Kajiwara, J.; Yoshimura, T.; Furue, M. Polychlorinated dibenzofurans as a causal agent of fetal Yusho. Chemosphere 2010, 80, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ueda, Y.; Hayashi, M.; Kato, H.; Furuhashi, T.; Morita, A. Tobacco smoke- induced skin pigmentation is mediated by the aryl hydrocarbon receptor. Exp. Dermatol. 2013, 22, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Nishida, E.; Morita, A. Action spectrum of melanoblast maturation and involvement of the aryl hydrocarbon receptor. Exp. Dermatol. 2016, 25 (Suppl. 3), 41–44. [Google Scholar] [CrossRef]
- Peng, F.; Xue, C.H.; Hwang, S.K.; Li, W.H.; Chen, Z.; Zhang, J.Z. Exposure to fine particulate matter associated with senile lentigo in Chinese women: A cross-sectional study. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Uchi, H.; Tsuji, G.; Gondo, H.; Moroi, Y.; Furue, M. Arylhydrocarbon receptor (AhR) activation in airway epithelial cells induces MUC5AC via reactive oxygen species (ROS) production. Pulm. Pharmacol. Ther. 2011, 24, 133–140. [Google Scholar] [CrossRef]
- Chiba, T.; Uchi, H.; Yasukawa, F.; Furue, M. Role of the arylhydrocarbon receptor in lung disease. Int. Arch. Allergy Immunol. 2011, 155 (Suppl. 1), 129–134. [Google Scholar] [CrossRef]
- Chiba, T.; Chihara, J.; Furue, M. Role of the Arylhydrocarbon Receptor (AhR) in the Pathology of Asthma and COPD. J. Allergy (Cairo) 2012, 2012, 372384. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, M.; He, J.; Shi, T.; Zhang, S.; Tang, N.; Chen, W. Polychlorinated dibenzo-dioxins and polychlorinated dibenzo-furans exposure and altered lung function: The mediating role of oxidative stress. Environ. Int. 2020, 137, 105521. [Google Scholar] [CrossRef]
- Guerrina, N.; Traboulsi, H.; Eidelman, D.H.; Baglole, C.J. The aryl hydrocarbon receptor and the maintenance of lung Health. Int. J. Mol. Sci. 2018, 19, 3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.C.; Yoshimura, M. Effects of benzo(a)pyrene on conduction velocity of the rat dorsal root and synaptic transmission in Substantia Gelatinosaneurons of the rat spinal dorsal horn. Fukuoka Igaku Zasshi 2017, 108, 75–82. [Google Scholar]
- Kanaya, K.; Yamashita, T.; Kawaguchi, S.; Takebayshi, T.; Katahira, G. The current perception threshold in lumbar disc herniation. Rinsho Seikei Geka 2003, 38, 301–305. [Google Scholar]
- Shackleford, G.; Sampathkumar, N.K.; Hichor, M.; Weill, L.; Meffre, D.; Juricek, L.; Laurendeau, I.; Chevallier, A.; Ortonne, N.; Larousserie, F.; et al. Involvement of aryl hydrocarbon receptor in myelination and in human nerve sheath tumorigenesis. Proc. Natl. Acad. Sci. USA 2018, 115, E1319–E1328. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xie, H.Q.; Sha, R.; Xu, T.; Zhang, S.; Fu, H.; Xia, Y.; Liu, Y.; Xu, L.; Zhao, B. 2,3,7,8-Tetrachlorodibenzo-p-dioxin and up-regulation of neurofilament expression in neuronal cells: Evaluation of AhR and MAPK pathways. Environ. Int. 2020, 134, 105193. [Google Scholar] [CrossRef]
- Okumura, M. Past and current medical states of yusho patients. Am. J. Ind. Med. 1984, 5, 13–18. [Google Scholar] [CrossRef]
- Tokunaga, S.; Kataoka, K. A longitudinal analysis on the association of serum lipids and lipoproteins concentrations with blood polychlorinated biphenyls level in chronic “Yusho” patients. Fukuoka Igaku Zasshi 2003, 94, 110–117. [Google Scholar]
- Ohashi, H.; Nishioka, K.; Nakajima, S.; Kim, S.; Suzuki, R.; Aizaki, H.; Fukasawa, M.; Kamisuki, S.; Sugawara, F.; Ohtani, N.; et al. The aryl hydrocarbon receptor-cytochrome P450 1A1 pathway controls lipid accumulation and enhances the permissiveness for hepatitis C virus assembly. J. Biol. Chem. 2018, 293, 19559–19571. [Google Scholar] [CrossRef] [Green Version]
- Biljes, D.; Hammerschmidt-Kamper, C.; Kadow, S.; Diel, P.; Weigt, C.; Burkart, V.; Esser, C. Impaired glucose and lipid metabolism in ageing aryl hydrocarbon receptor deficient mice. EXCLI J. 2015, 14, 1153–1163. [Google Scholar]
- Zhu, X.Y.; Xia, H.G.; Wang, Z.H.; Li, B.; Jiang, H.Y.; Li, D.L.; Jin, R.; Jin, Y. In vitro and in vivo approaches for identifying the role of aryl hydrocarbon receptor in the development of nonalcoholic fatty liver disease. Toxicol. Lett. 2020, 319, 85–94. [Google Scholar] [CrossRef]
- das Garcas Abeles, E.; Cordeiro, L.M.; Martins, A.; Pesquero, J.L.; Reis, A.M.; Andrade, S.P.; Botion, L.M. Estrogen therapy attenuates adiposity markers in spontaneously hypertensive rats. Metabolism 2012, 61, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Huang, F.; Wang, J.; Du, Y. Effects of co-exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin and polychlorinated biphenyls on nonalcoholic fatty liver disease in mice. Environ. Toxicol. 2015, 30, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Shearman, L.P.; Weaver, D.R.; Zylka, M.J.; de Vries, G.J.; Reppert, S.M. A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell 1999, 96, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, U. Timing to perfection: The biology of central and peripheral circadian clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, C.; Tischkau, S.A. Role of aryl hydrocarbon receptor in circadian clock disruption and metabolic dysfunction. Environ. Health Insights 2016, 10, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Mukai, M.; Tischkau, S.A. Effects of tryptophan photoproducts in the circadian timing system: Searching for a physiological role for aryl hydrocarbon receptor. Toxicol. Sci. 2007, 95, 172–181. [Google Scholar] [CrossRef]
- Kondo, H.; Tanio, K.; Nagaura, Y.; Nagayoshi, M.; Mitoma, C.; Furue, M.; Maeda, T. Sleep disorders among Yusho patients highly intoxicated with dioxin-related compounds: A 140-case series. Environ. Res. 2018, 166, 261–268. [Google Scholar] [CrossRef]
- Guo, Y.L.; Yu, M.L.; Hsu, C.C.; Rogan, W.J. Chloracne, goiter, arthritis, and anemia after polychlorinated biphenyl poisoning: 14-year follow-up of the Taiwan Yucheng cohort. Environ. Health Perspect. 1999, 107, 715–719. [Google Scholar] [CrossRef]
- Lee, H.G.; Yang, J.H. PKC-δ mediates TCDD-induced apoptosis of chondrocyte in ROS-dependent manner. Chemosphere 2010, 81, 1039–1044. [Google Scholar] [CrossRef]
- Yang, J.H.; Lee, H.G. 2,3,7,8-Tetrachlorodibenzo-p-dioxin induces apoptosis of articular chondrocytes in culture. Chemosphere 2010, 79, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Fukushi, J.I.; Tsushima, H.; Matsumoto, Y.; Mitoma, C.; Furue, M.; Miyahara, H.; Nakashima, Y. Influence of dioxin-related compounds on physical function in Yusho incident victims. Heliyon 2019, 5, e02702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukushi, J.; Tokunaga, S.; Nakashima, Y.; Motomura, G.; Mitoma, C.; Uchi, H.; Furue, M.; Iwamoto, Y. Effects of dioxin-related compounds on bone mineral density in patients affected by the Yusho incident. Chemosphere 2016, 145, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, M.; Hashimoto-Hachiya, A.; Tsuji, G. Aryl hydrocarbon receptor in atopic dermatitis and psoriasis. Int. J. Mol. Sci. 2019, 20, 5424. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.C.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef]
- Schiering, C.; Vonk, A.; Das, S.; Stockinger, B.; Wincent, E. Cytochrome P4501-inhibiting chemicals amplify aryl hydrocarbon receptor activation and IL-22 production in T helper 17 cells. Biochem. Pharmacol. 2018, 151, 47–58. [Google Scholar] [CrossRef]
- Funatake, C.J.; Marshall, N.B.; Steppan, L.B.; Mourich, D.V.; Kerkvliet, N.I. Cutting edge: Activation of the aryl hydrocarbon receptor by 2,3,7,8-tetrachlorodibenzo-p-dioxin generates a population of CD4+ CD25+ cells with characteristics of regulatory T cells. J. Immunol. 2005, 175, 4184–4188. [Google Scholar] [CrossRef] [Green Version]
- Kamio, Y.; Gunge, Y.; Koike, Y.; Kuwatsuka, Y.; Tsuruta, K.; Yanagihara, K.; Furue, M.; Murota, H. Insight into innate immune response in “Yusho”: The impact of natural killer cell and regulatory T cell on inflammatory prone diathesis of Yusho patients. Environ. Res. 2020, 185, 109415. [Google Scholar] [CrossRef]
- Kuwatsuka, Y.; Shimizu, K.; Akiyama, Y.; Koike, Y.; Ogawa, F.; Furue, M.; Utani, A. Yusho patients show increased serum IL-17, IL-23, IL-1β, and TNFα levels more than 40 years after accidental polychlorinated biphenyl poisoning. J. Immunotoxicol. 2014, 11, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Morino-Koga, S.; Uchi, H.; Tsuji, G.; Takahara, M.; Kajiwara, J.; Hirata, T.; Furue, M. Reduction of CC-chemokine ligand 5 by aryl hydrocarbon receptor ligands. J. Dermatol. Sci. 2013, 72, 9–15. [Google Scholar] [CrossRef]
- Ito, T.; Inouye, K.; Nohara, K.; Tohyama, C.; Fujimaki, H. TCDD exposure exacerbates atopic dermatitis-related inflammation in NC/Nga mice. Toxicol. Lett. 2008, 177, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, T.; Ogawa, E.; Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Fujimura, T.; Aiba, S.; Nakayama, K.; Okuyama, R.; et al. The aryl hydrocarbon receptor AhR links atopic dermatitis and air pollution via induction of the neurotrophic factor artemin. Nat. Immunol. 2017, 18, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Uchi, H.; Furue, M. Prevalence of atopic dermatitis and serum IgE of Yusho patients. Fukuoka Igaku Zasshi 2009, 100, 127–130. [Google Scholar] [PubMed]
- Takeuchi, S.; Esaki, H.; Furue, M. Epidemiology of atopic dermatitis in Japan. J. Dermatol. 2014, 41, 200–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Warner, M.; Mocarelli, P.; Brambilla, P.; Eskenazi, B. Prenatal exposure to TCDD and atopic conditions in the Seveso second generation: A prospective cohort study. Environ. Health 2018, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Kransler, K.M.; McGarrigle, B.P.; Olson, J.R. Comparative developmental toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in the hamster, rat and guinea pig. Toxicology 2007, 229, 214–225. [Google Scholar] [CrossRef]
- Flaveny, C.A.; Perdew, G.H. Transgenic Humanized AHR Mouse Reveals Differences between Human and Mouse AHR Ligand Selectivity. Mol. Cell Pharmacol. 2009, 1, 119–123. [Google Scholar] [CrossRef]
- Sun, Y.V.; Boverhof, D.R.; Burgoon, L.D.; Fielden, M.R.; Zacharewski, T.R. Comparative analysis of dioxin response elements in human, mouse and rat genomic sequences. Nucleic Acids Res. 2004, 32, 4512–4523. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.W.; Ohrr, H. Agent Orange exposure and cancer incidence in Korean Vietnam veterans: A prospective cohort study. Cancer 2014, 120, 3699–3706. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.W.; Ryu, S.Y.; Ohrr, H.; Hong, J.S. Agent Orange exposure and risk of death in Korean Vietnam veterans: Korean Veterans Health Study. Int. J. Epidemiol. 2014, 43, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Onozuka, D.; Yoshimura, T.; Kaneko, S.; Furue, M. Mortality after exposure to polychlorinated biphenyls and polychlorinated dibenzofurans: A 40-year follow-up study of Yusho patients. Am. J. Epidemiol. 2009, 169, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.C.; Tsai, P.C.; Chen, P.C.; Hsieh, C.J.; Guo, Y.L.; Rogan, W.J. Mortality after exposure to polychlorinated biphenyls and dibenzofurans: 30 years after the “Yucheng accident”. Environ. Res. 2013, 120, 71–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.C.; Chen, P.C.; Tsai, P.C.; Furue, M.; Onozuka, D.; Hagihara, A.; Uchi, H.; Yoshimura, T.; Guo, Y.L. Mortality after exposure to polychlorinated biphenyls and polychlorinated dibenzofurans: A meta-analysis of two highly exposed cohorts. Int. J. Cancer 2015, 137, 1427–1432. [Google Scholar] [CrossRef] [Green Version]
- Onozuka, D.; Nakamura, Y.; Tsuji, G.; Furue, M. Mortality in Yusho patients exposed to polychlorinated biphenyls and polychlorinated dibenzofurans: A 50-year retrospective cohort study. Environ. Health 2020, 19, 119. [Google Scholar] [CrossRef] [PubMed]
- Onozuka, D.; Hirata, T.; Furue, M. Relative survival after exposure to polychlorinated biphenyls and dioxins: A follow-up of Japanese patients affected in the Yusho incident. Sci. Total Environ. 2011, 409, 2361–2365. [Google Scholar] [CrossRef] [PubMed]
- Onozuka, D.; Hirata, T.; Furue, M. Net survival after exposure to polychlorinated biphenyls and dioxins: The Yusho study. Environ. Int. 2014, 73, 28–32. [Google Scholar] [CrossRef]
- Tsay, J.J.; Tchou-Wong, K.M.; Greenberg, A.K.; Pass, H.; Rom, W.N. Aryl hydrocarbon receptor and lung cancer. Anticancer Res. 2013, 33, 1247–1256. [Google Scholar]
- Lu, P.; Cai, X.; Guo, Y.; Xu, M.; Tian, J.; Locker, J.; Xie, W. Constitutive activation of the human aryl hydrocarbon receptor in mice promotes hepatocarcinogenesis independent of its coactivator Gadd45b. Toxicol. Sci. 2019, 167, 581–592. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hankinson, O. 2,3,7,8-tetrachlorodibenzo-p-dioxin suppresses the growth of human colorectal cancer cells in vitro: Implication of the aryl hydrocarbon receptor signaling. Int. J. Oncol. 2019, 54, 1422–1432. [Google Scholar] [CrossRef]
- Huerta-Yepez, S.; Tirado-Rodriguez, A.; Montecillo-Aguado, M.R.; Yang, J.; Hammock, B.D.; Hankinson, O. Aryl Hydrocarbon Receptor-Dependent inductions of omega-3 and omega-6 polyunsaturated fatty acid metabolism act inversely on tumor progression. Sci. Rep. 2020, 10, 7843. [Google Scholar] [CrossRef]
- Kajiwara, J.; Todaka, T.; Hirakawa, H.; Hori, T.; Onozuka, D.; Takao, Y.; Hirata, T.; Iida, T.; Uchi, H.; Furue, M. The difference between male and female dioxin concentrations in the blood of Yusho patients. Fukuoka Igaku Zasshi 2011, 102, 140–144. [Google Scholar] [PubMed]
- Wakx, A.; Nedder, M.; Tomkiewicz-Raulet, C.; Dalmasso, J.; Chissey, A.; Boland, S.; Vibert, F.; Degrelle, S.A.; Fournier, T.; Coumoul, X.; et al. Expression, localization, and activity of the aryl hydrocarbon receptor in the human placenta. Int. J. Mol. Sci. 2018, 19, 3762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukimori, K.; Uchi, H.; Mitoma, C.; Yasukawa, F.; Fukushima, K.; Todaka, T.; Kajiwara, J.; Yoshimura, T.; Hirata, T.; Wake, N.; et al. Comparison of the concentrations of polychlorinated biphenyls and dioxins in mothers affected by the Yusho incident and their children. Chemosphere 2011, 84, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Tsukimori, K.; Uchi, H.; Tokunaga, S.; Yasukawa, F.; Chiba, T.; Kajiwara, J.; Hirata, T.; Furue, M. Blood levels of PCDDs, PCDFs, and coplanar PCBs in Yusho mothers and their descendants: Association with fetal Yusho disease. Chemosphere 2013, 90, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Morokuma, S.; Tsukimori, K.; Hori, T.; Kato, K.; Furue, M. The Vernix caseosa is the main site of dioxin excretion in the human foetus. Sci. Rep. 2017, 7, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsueda, T.; Iida, T.; Hirakawa, H.; Fukamachi, K.; Tokiwa, H. Concentration of PCDDs, PCDFs and Coplanar PCBs in breast milk of Yusho patients and normal subjects. Fukuoka Igaku Zasshi 1993, 84, 263–272. [Google Scholar]
- Tsukimori, K.; Tokunaga, S.; Shibata, S.; Uchi, H.; Nakayama, D.; Ishimaru, T.; Nakano, H.; Wake, N.; Yoshimura, T.; Furue, M. Long-term effects of polychlorinated biphenyls and dioxins on pregnancy outcomes in women affected by the Yusho incident. Environ. Health Perspect. 2008, 116, 626–630. [Google Scholar] [CrossRef] [Green Version]
- Tsukimori, K.; Uchi, H.; Mitoma, C.; Yasukawa, F.; Chiba, T.; Todaka, T.; Kajiwara, J.; Yoshimura, T.; Hirata, T.; Fukushima, K.; et al. Maternal exposure to high levels of dioxins in relation to birth weight in women affected by Yusho disease. Environ. Int. 2012, 38, 79–86. [Google Scholar] [CrossRef]
- Wesselink, A.; Warner, M.; Samuels, S.; Parigi, A.; Brambilla, P.; Mocarelli, P.; Eskenazi, B. Maternal dioxin exposure and pregnancy outcomes over 30 years of follow-up in Seveso. Environ. Int. 2014, 63, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Ames, J.; Warner, M.; Mocarelli, P.; Brambilla, P.; Signorini, S.; Siracusa, C.; Huen, K.; Holland, N.; Eskenazi, B. AHR gene-dioxin interactions and birthweight in the Seveso Second Generation Health Study. Int. J. Epidemiol. 2018, 47, 1992–2004. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.L.; Guo, Y.L.; Hsu, C.C.; Rogan, W.J. Menstruation and reproduction in women with polychlorinated biphenyl (PCB) poisoning: Long-term follow-up interviews of the women from the Taiwan Yucheng cohort. Int. J. Epidemiol. 2000, 29, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskenazi, B.; Warner, M.; Marks, A.R.; Samuels, S.; Needham, L.; Brambilla, P.; Mocarelli, P. Serum dioxin concentrations and time to pregnancy. Epidemiology 2010, 21, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Warner, M.; Brambilla, P.; Signorini, S.; Ames, J.; Mocarelli, P. The Seveso accident: A look at 40 years of health research and beyond. Environ. Int. 2018, 121, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Tamura, M.; Yamashita, J.; Suzuki, C.; Tomita, T. Repeated in utero and lactational 2,3,7,8-tetrachlorodibenzo-p-dioxin exposure affects male gonads in offspring, leading to sex ratio changes in F2 progeny. Toxicol. Appl. Pharmacol. 2005, 206, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Hertz-Picciotto, I.; Jusko, T.A.; Willman, E.J.; Baker, R.J.; Keller, J.A.; Teplin, S.W.; Charles, M.J. A cohort study of in utero polychlorinated biphenyl (PCB) exposures in relation to secondary sex ratio. Environ. Health 2008, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocarelli, P.; Brambilla, P.; Gerthoux, P.M.; Patterson, D.G., Jr.; Needham, L.L. Change in sex ratio with exposure to dioxin. Lancet 1996, 348, 409. [Google Scholar] [CrossRef]
- Rogan, W.J.; Gladen, B.C.; Guo, Y.L.; Hsu, C.C. Sex ratio after exposure to dioxin-like chemicals in Taiwan. Lancet 1999, 353, 206–207. [Google Scholar] [CrossRef]
- del Rio Gomez, I.; Marshall, T.; Tsai, P.; Shao, Y.S.; Guo, Y.L. Number of boys born to men exposed to polychlorinated byphenyls. Lancet 2002, 360, 143–144. [Google Scholar] [CrossRef]
- Rowlands, J.C.; Budinsky, R.A.; Aylward, L.L.; Faqi, A.S.; Carney, E.W. Sex ratio of the offspring of Sprague-Dawley rats exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in utero and lactationally in a three-generation study. Toxicol. Appl. Pharmacol. 2006, 216, 29–33. [Google Scholar] [CrossRef]
- Bircsak, K.M.; Copes, L.T.; King, S.; Prantner, A.M.; Hwang, W.T.; Gerton, G.L. The aryl hydrocarbon receptor mediates sex ratio distortion in the embryos sired by TCDD-exposed male mice. Reprod. Toxicol. 2020, 94, 75–83. [Google Scholar] [CrossRef]
- Tsukimori, K.; Yasukawa, F.; Uchi, H.; Furue, M.; Morokuma, S. Sex ratio in two generations of the Yusho cohort. Epidemiology 2012, 23, 349–350. [Google Scholar] [CrossRef]
- Viluksela, M.; Pohjanvirta, R. Multigenerational and transgenerational effects of dioxins. Int. J. Mol. Sci. 2019, 20, 2947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, M.; Eskenazi, B.; Olive, D.L.; Samuels, S.; Quick-Miles, S.; Vercellini, P.; Gerthoux, P.M.; Needham, L.; Patterson, D.G.; Mocarelli, P. Serum dioxin concentrations and quality of ovarian function in women of Seveso. Environ. Health Perspect. 2007, 115, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Mocarelli, P.; Gerthoux, P.M.; Needham, L.L.; Patterson, D.G., Jr.; Limonta, G.; Falbo, R.; Signorini, S.; Bertona, M.; Crespi, C.; Sarto, C.; et al. Perinatal exposure to low doses of dioxin can permanently impair human semen quality. Environ. Health Perspect. 2011, 119, 713–718. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.C.; Huang, W.; Yao, W.J.; Wu, M.H.; Guo, Y.L.; Lambert, G.H. Sperm changes in men exposed to polychlorinated biphenyls and dibenzofurans. JAMA 2003, 289, 2943–2944. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Li, M.C.; Lee, Y.C.; Kuo, P.L.; Guo, Y.L. Polychlorinated biphenyls and dibenzofurans increased abnormal sperm morphology without alterations in aneuploidy: The Yucheng study. Chemosphere 2016, 165, 294–297. [Google Scholar] [CrossRef]
- Johnson, K.J.; Passage, J.; Lin, H.; Sriram, S.; Budinsky, R.A. Dioxin male rat reproductive toxicity mode of action and relative potency of 2,3,7,8-tetrachlorodibenzo-p-dioxin and 2,3,7,8-tetrachlorodibenzofuran characterized by fetal pituitary and testis transcriptome profiling. Reprod. Toxicol. 2020, 93, 146–162. [Google Scholar] [CrossRef]
- Ames, J.; Warner, M.; Brambilla, P.; Mocarelli, P.; Satariano, W.A.; Eskenazi, B. Neurocognitive and physical functioning in the Seveso Women’s Health Study. Environ. Res. 2018, 162, 55–62. [Google Scholar] [CrossRef]
- Slama, N.; Warner, M.; Mocarelli, P.; Brambilla, P.; Eskenazi, B. The 2nd to 4th digit length ratio (2D:4D) among children of Seveso women exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Early Hum. Dev. 2019, 131, 45–50. [Google Scholar] [CrossRef]
- Ames, J.; Warner, M.; Siracusa, C.; Signorini, S.; Brambilla, P.; Mocarelli, P.; Eskenazi, B. Prenatal dioxin exposure and neuropsychological functioning in the Seveso Second Generation Health Study. Int. J. Hyg. Environ. Health 2019, 222, 425–433. [Google Scholar] [CrossRef]
- Warner, M.; Rauch, S.; Ames, J.; Mocarelli, P.; Brambilla, P.; Signorini, S.; Eskenazi, B. In utero dioxin exposure and cardiometabolic risk in the Seveso Second Generation Study. Int. J. Obes. (Lond.) 2019, 43, 2233–2243. [Google Scholar] [CrossRef] [PubMed]
- Warner, M.; Rauch, S.; Ames, J.; Mocarelli, P.; Brambilla, P.; Signorini, S.; Eskenazi, B. Prenatal dioxin exposure and thyroid hormone levels in the Seveso second generation study. Environ. Res. 2020, 183, 109280. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.P.; Wu, S.W.; Huang, Y.J.; Chiang, M.C.; Hsieh, S.T.; Guo, Y.L. Neuroimaging signatures of brain plasticity in adults with prenatal exposure to polychlorinated biphenyls: Altered functional connectivity on functional MRI. Environ. Pollut. 2019, 250, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Li, M.C.; Wu, H.P.; Yang, C.Y.; Chen, P.C.; Lambert, G.H.; Guo, Y.L. Gestational exposure to polychlorinated biphenyls and dibenzofurans induced asymmetric hearing loss: Yucheng children study. Environ. Res. 2015, 137, 65–71. [Google Scholar] [CrossRef]
- Poland, A.; Knutson, J.C. 2,3,7,8-tetrachlorodibenzo-p-dioxin and related halogenated aromatic hydrocarbons: Examination of the mechanism of toxicity. Annu. Rev. Pharmacol. Toxicol. 1982, 22, 517–554. [Google Scholar] [CrossRef]
- Peterson, R.E.; Theobald, H.M.; Kimmel, G.L. Developmental and reproductive toxicity of dioxins and related compounds: Cross-species comparisons. Crit. Rev. Toxicol. 1993, 23, 283–335. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.S. In utero 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) alters reproductive morphology and function in female rat offspring. Toxicol. Appl. Pharmacol. 1995, 133, 285–294. [Google Scholar] [CrossRef]
- Bjerke, D.L.; Peterson, R.E. Reproductive toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in male rats: Different effects of in utero versus lactational exposure. Toxicol. Appl. Pharmacol. 1994, 127, 241–249. [Google Scholar] [CrossRef]
- McCarthy, M.M. How it’s made: Organisational effects of hormones on the developing brain. J. Neuroendocrinol. 2010, 22, 736–742. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Takeda, T.; Koga, T.; Ishii, Y. Role of critical period in sex and brain differentiation: Learning from dioxin-induced disorders in next generations. Yakugaku Zasshi 2014, 134, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T. Molecular mechanism wherby maternal exposure to dioxin suppresses sexual maturation of the offspring after growing up. Yakugaku Zasshi 2017, 137, 1373–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, T.; Ishida, T.; Takeda, T.; Ishii, Y.; Uchi, H.; Tsukimori, K.; Yamamoto, M.; Himeno, M.; Furue, M.; Yamada, H. Restoration of dioxin-induced damage to fetal steroidogenesis and gonadotropin formation by maternal co-treatment with α-lipoic acid. PLoS ONE 2012, 7, e40322. [Google Scholar] [CrossRef]
- Mutoh, J.; Taketoh, J.; Okamura, K.; Kagawa, T.; Ishida, T.; Ishii, Y.; Yamada, H. Fetal pituitary gonadotropin as an initial target of dioxin in its impairment of cholesterol transportation and steroidogenesis in rats. Endocrinology 2006, 147, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, T.; Matsumoto, Y.; Koga, T.; Mutoh, J.; Nishimura, Y.; Shimazoe, T.; Ishii, Y.; Ishida, T.; Yamada, H. Maternal exposure to dioxin disrupts gonadotropin production in fetal rats and imprints defects in sexual behavior. J. Pharmacol. Exp. Ther. 2009, 329, 1091–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taura, J.; Takeda, T.; Fujii, M.; Hattori, Y.; Ishii, Y.; Kuroki, H.; Tsukimori, K.; Uchi, H.; Furue, M.; Yamada, H. 2,3,4,7,8-Pentachlorodibenzofuran is far less potent than 2,3,7,8-tetrachlorodibenzo-p-dioxin in disrupting the pituitary-gonad axis of the rat fetus. Toxicol. Appl. Pharmacol. 2014, 281, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Fujii, M.; Taura, J.; Ishii, Y.; Yamada, H. Dioxin silences gonadotropin expression in perinatal pups by inducing histone deacethylases: A new insight into the mechanism for the imprinting of sexual immaturity by dioxin. J. Biol. Chem. 2012, 287, 18440–18450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, T.; Fujii, M.; Hattori, Y.; Yamamoto, M.; Shimazoe, T.; Ishii, Y.; Himeno, M.; Yamada, H. Maternal exposure to dioxin imprints sexual immaturity of the pups through fixing the status of the reduced expression of hypothalamic gonadotropin-releasing hormone. Mol. Pharmacol. 2014, 85, 74–82. [Google Scholar] [CrossRef]
- Takeda, T.; Taura, J.; Hattori, Y.; Ishii, Y.; Yamada, H. Dioxin-induced retardation of development through a reduction in the expression of pituitary hormones and possible involvement of an aryl hydrocarbon receptor in this defect: A comparative study using two strains of mice with different sensitivities to dioxin. Toxicol. Appl. Pharmacol. 2014, 278, 220–229. [Google Scholar]
- Kuroki, J.; Koga, N.; Yoshimura, H. High affinity of 2,3,4,7,8-pentachlorodibenzofuran to cytochrome P-450 in the hepatic microsome of rats. Chemosphere 1986, 15, 731–738. [Google Scholar] [CrossRef]
- Chen, C.; Hamm, J.T.; Hass, J.R.; Birnbaum, L.S. Disposition of polychlorinated dibenzo-p-dioxins, dibenzofurans, and non-ortho polychlorinated biphenyls in pregnant Long Evans rats and the transfer to offspring. Toxicol. Appl. Pharmacol. 2001, 173, 65–88. [Google Scholar] [CrossRef]
- Diliberto, J.J.; Burgin, D.; Birnbaum, L.S. Role of CYP1A2 in hepatic sequestration of dioxin: Studies using CYP1A2 knock-out mice. Biochem. Biophys. Res. Commun. 1997, 236, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Tsukimori, K.; Morokuma, S.; Hori, T.; Takahashi, K.; Hirata, T.; Otera, Y.; Fukushima, K.; Kawamoto, T.; Wake, N. Characterization of placental transfer of polychlorinated dibenzo-p-dioxins, dibenzofurans and polychlorinated biphenyls in normal pregnancy. J. Obstet. Gynaecol. Res. 2013, 39, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Komiya, Y.; Koga, T.; Ishida, T.; Ishii, Y.; Kikuta, Y.; Nakaya, M.; Kurose, H.; Yokomizo, T.; Shimizu, T.; et al. Dioxin-induced increase in leukotriene B4 biosynthesis through the aryl hydrocarbon receptor and its relevance to hepatotoxicity owing to neutrophil infiltration. J. Biol. Chem. 2017, 292, 10586–10599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, Y.; Takeda, T.; Nakamura, A.; Nishida, K.; Shioji, Y.; Fukumitsu, H.; Yamada, H.; Ishii, Y. The aryl hydrocarbon receptor is indispensable for dioxin-induced defects in sexually-dimorphic behaviors due to the reduction in fetal steroidogenesis of the pituitary-gonadal axis in rats. Biochem. Pharmacol. 2018, 154, 213–221. [Google Scholar] [CrossRef]
- Thiel, R.; Koch, E.; Ulbrich, B.; Chahoud, I. Peri and postnatal exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin: Effects on physiological development, reflexes, locomotor activity and learning behaviour in Wistar rats. Arch. Toxicol. 1994, 69, 79–86. [Google Scholar] [CrossRef]
- Myllymäki, S.A.; Haavisto, T.E.; Brokken, L.J.; Viluksela, M.; Toppari, J.; Paranko, J. In utero and lactational exposure to TCDD; steroidogenic outcomes differ in male and female rat pups. Toxicol. Sci. 2005, 88, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Nishijo, M.; Kuriwaki, J.; Hori, E.; Tawara, K.; Nakagawa, H.; Nishijo, H. Effects of maternal exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin on fetal brain growth and motor and behavioral development in offspring rats. Toxicol. Lett. 2007, 30, 41–47. [Google Scholar] [CrossRef]
- Hojo, R.; Kakeyama, M.; Kurokawa, Y.; Aoki, Y.; Yonemoto, J.; Tohyama, C. Learning behavior in rat offspring after in utero and lactational exposure to either TCDD or PCB126. Environ. Health Prev. Med. 2008, 13, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Negishi, T.; Shimomura, H.; Koyama, T.; Kawasaki, K.; Ishii, Y.; Kyuwa, S.; Yasuda, M.; Kuroda, Y.; Yoshikawa, Y. Gestational and lactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin affects social behaviors between developing rhesus monkeys (Macaca mulatta). Toxicol. Lett. 2006, 160, 233–244. [Google Scholar] [CrossRef]
- Chen, Y.C.; Guo, Y.L.; Hsu, C.C.; Rogan, W.J. Cognitive development of Yu-Cheng (“oil disease”) children prenatally exposed to heat-degraded PCBs. JAMA 1992, 268, 3213–3218. [Google Scholar] [CrossRef]
- Hattori, Y.; Takeda, T.; Fujii, M.; Taura, J.; Ishii, Y.; Yamada, H. Dioxin-induced fetal growth retardation: The role of a preceding attenuation in the circulating level of glucocorticoid. Endocrine 2014, 47, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Takeda, T.; Taura, J.; Kuroki, H.; Ishii, Y.; Yamada, H. The effect of maternal exposure to dioxins on the level of thyroid hormone in developing pups. Fukuoka Igaku Zasshi 2015, 106, 127–134. [Google Scholar]
- Takeda, T.; Fujii, M.; Izumoto, W.; Hattori, Y.; Matsushita, T.; Yamada, H.; Ishii, Y. Gestational dioxin exposure suppresses prolactin-stimulated nursing in lactating dam rats to impair development of postnatal offspring. Biochem. Pharmacol. 2020, 178, 114106. [Google Scholar] [CrossRef] [PubMed]
- Ema, M.; Ohe, N.; Suzuki, M.; Mimura, J.; Sogawa, K.; Ikawa, S.; Fujii-Kuriyama, Y. Dioxin biding activities of polymorphic form of mouse and human arylhydrocarbon receptors. J. Biol. Chem. 1994, 269, 27337–27343. [Google Scholar] [CrossRef]
- Grandjean, P.; Grønlund, C.; Kjær, I.M.; Jensen, T.K.; Sørensen, N.; Andersson, A.M.; Juul, A.; Skakkebæk, N.E.; Budtz-Jørgensen, E.; Weihe, P. Reproductive hormone profile and pubertal development in 14-year-old boys prenatally exposed to polychlorinated biphenyls. Reprod. Toxicol. 2012, 34, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winneke, G.; Ranft, U.; Wittsiepe, J.; Kasper-Sonnenberg, M.; Fürst, P.; Krämer, U.; Seitner, G.; Wilhelm, M. Behavioral sexual dimorphism in school-age children and early developmental exposure to dioxins and PCBs: A follow-up study of the Duisburg Cohort. Environ. Health Perspect. 2014, 122, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Nowack, N.; Wittsiepe, J.; Kasper-Sonnenberg, M.; Wilhelm, M.; Schölmerich, A. Influence of Low-Level Prenatal Exposure to PCDD/Fs and PCBs on Empathizing, Systemizing and Autistic Traits: Results from the Duisburg Birth Cohort Study. PLoS ONE 2015, 10, e0129906. [Google Scholar] [CrossRef]
- Su, K.Y.; Li, M.C.; Lee, N.W.; Ho, B.C.; Cheng, C.L.; Chuang, Y.C.; Yu, S.L.; Guo, Y.L. Perinatal polychlorinated biphenyls and polychlorinated dibenzofurans exposure are associated with DNA methylation changes lasting to early adulthood: Findings from Yucheng second generation. Environ. Res. 2019, 170, 481–486. [Google Scholar] [CrossRef]
- Sorg, O.; Zennegg, M.; Schmid, P.; Fedosyuk, R.; Valikhnovskyi, R.; Gaide, O.; Kniazevych, V.; Saurat, J.H. 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) poisoning in Victor Yushchenko: Identification and measurement of TCDD metabolites. Lancet 2009, 374, 1179–1185. [Google Scholar] [CrossRef]
- Saurat, J.H.; Kaya, G.; Saxer-Sekulic, N.; Pardo, B.; Becker, M.; Fontao, L.; Mottu, F.; Carraux, P.; Pham, X.C.; Barde, C.; et al. The cutaneous lesions of dioxin exposure: Lessons from the poisoning of Victor Yushchenko. Toxicol. Sci. 2012, 125, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, S.; Miyawaki, T.; Hori, T.; Kajiwara, J.; Katsuki, S.; Hirano, M.; Yoshinouchi, Y.; Iwata, H.; Mitoma, C.; Furue, M. Accumulation properties of polychlorinated biphenyl congeners in Yusho patients and prediction of their cytochrome P450-dependent metabolism by in silico analysis. Environ. Sci. Pollut. Res. Int. 2018, 25, 16455–16463. [Google Scholar] [CrossRef] [PubMed]
- Todaka, T.; Hirakawa, H.; Hori, T.; Tobiishi, K.; Iida, T.; Furue, M. Concentrations of polychlorinated dibenzo-p-dioxins, polychlorinated dibenzofurans, and non-ortho and mono-ortho polychlorinated biphenyls in blood of Yusho patients. Chemosphere 2007, 66, 1983–1989. [Google Scholar] [CrossRef] [PubMed]
- Todaka, T.; Hori, T.; Hirakawa, H.; Kajiwara, J.; Yasutake, D.; Onozuka, D.; Iida, T.; Furue, M. Concentrations of polychlorinated biphenyls in blood of Yusho patients over 35 years after the incident. Chemosphere 2009, 74, 902–909. [Google Scholar] [CrossRef]
- Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Koike, S.; Yoshimura, T.; Mitoma, C.; Shibata, S.; Uchi, H.; Furue, M.; Imamura, T. Variation in half-life of penta-chlorodibenzofuran (PeCDF) blood level among Yusho patients. Chemosphere 2009, 77, 658–662. [Google Scholar] [CrossRef]
- Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Kajiwara, J.; Mitoma, C.; Uchi, H.; Furue, M.; Imamura, T. Unexpectedly long half-lives of blood 2,3,4,7,8-pentachlorodibenzofuran (PeCDF) levels in Yusho patients. Environ. Health 2015, 14, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Kajiwara, J.; Mitoma, C.; Uchi, H.; Furue, M.; Imamura, T. Change in decay rates of dioxin-like compounds in Yusho patients. Environ. Health 2016, 15, 95. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Akahane, M.; Kanagawa, Y.; Kajiwara, J.; Todaka, T.; Yasukawa, F.; Uchi, H.; Furue, M.; Imamura, T. Individuals’ half-lives for 2,3,4,7,8-penta-chlorodibenzofuran (PeCDF) in blood: Correlation with clinical manifestations and laboratory results in subjects with Yusho. Chemosphere 2013, 92, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Kuroki, H.; Masuda, Y.; Koga, N.; Yoshimura, H. Accumulation of PCDFs and enzyme induction in the liver of monkeys which were poisoned by PCBs. Fukuoka Igaku Zasshi 1983, 74, 234–239. [Google Scholar] [PubMed]
- Kuroki, H.; Masuda, Y.; Yoshihara, S.; Yoshimura, H. Accumulation of polychlorinated dibenzofurans in the livers of monkeys and rats. Food Cosmet. Toxicol. 1980, 18, 387–392. [Google Scholar] [CrossRef]
- Yoshimura, H.; Kuroki, J.; Koga, N.; Kuroki, H.; Masuda, Y.; Fukasaku, N.; Hasegawa, M. High accumulation of 2,3,4,7,8-pentachlorodibenzofuran to hepatic microsomes of rats. J. Pharmacobiodyn. 1984, 7, 414–419. [Google Scholar] [CrossRef]
- Masuda, Y.; Kuroki, H.; Haraguchi, K.; Nagayama, J. PCB and PCDF congeners in the blood and tissues of yusho and yu-cheng patients. Environ. Health Perspect. 1985, 59, 53–58. [Google Scholar] [PubMed] [Green Version]
- Brewster, D.W.; Birnbaum, L.S. Disposition and excretion of 2,3,4,7,8-pentachlorodibenzofuran in the rat. Toxicol. Appl. Pharmacol. 1987, 90, 243–252. [Google Scholar] [CrossRef]
- Okumura, M.; Yamanaka, M.; Nakamuta, S. Ten year follow-up study on serum triglyceride levels in 24 patients with PCB poisoning. Fukuoka Igaku Zasshi 1979, 70, 208–210. [Google Scholar] [PubMed]
- Yoshimura, H.; Yamamoto, H. A novel route of excretion of 2, 4, 3′,4′-tetrachlorobiphenyl in rats. Bull. Environ. Contam. Toxicol. 1975, 13, 681–688. [Google Scholar] [CrossRef]
- Yoshimura, H.; Kamimura, H.; Oguri, K.; Saeki, S. Stimulating effect of squalene on fecal excretion of high toxic 2,3,4,7,8-pentachlorodibenzofuran (PenCDF) in rats. Fukuoka Igaku Zasshi 1985, 76, 184–189. [Google Scholar]
- Kamimura, H.; Yoshimura, H. Stimulation of fecal excretion of etiological compounds of Yusho in rats. Fukuoka Igaku Zasshi 1987, 78, 266–280. [Google Scholar]
- Richter, E.; Lay, J.P.; Klein, W.; Korte, F. Paraffin-stimulated excretion of 2,4,6,2′4′-pentachlorobi[1 4C]phenyl by rats. Toxicol. Appl. Pharmacol. 1979, 50, 17–23. [Google Scholar] [CrossRef]
- Richter, E.; Schäfer, S.G.; Fichtl, B. Stimulation of the faecal excretion of 2,4,5,2′,4′,5′-hexachlorobiphenyl in rats by squalane. Xenobiotica 1983, 13, 337–343. [Google Scholar] [CrossRef]
- Sakurai, K.; Fukata, H.; Todaka, E.; Saito, Y.; Bujo, H.; Mori, C. Colestimide reduces blood polychlorinated biphenyl (PCB) levels. Intern. Med. 2006, 45, 327–328. [Google Scholar] [CrossRef] [Green Version]
- Geusau, A.; Tschachler, E.; Meixner, M.; Sandermann, S.; Päpke, O.; Wolf, C.; Valic, E.; Stingl, G.; McLachlan, M. Olestra increases faecal excretion of 2,3,7,8-tetrachlorodibenzo-p-dioxin. Lancet 1999, 354, 1266–1267. [Google Scholar] [CrossRef]
- Geusau, A.; Schmaldienst, S.; Derfler, K.; Päpke, O.; Abraham, K. Severe 2,3,7,8-tetrachlorodibenzo- p-dioxin (TCDD) intoxication: Kinetics and trials to enhance elimination in two patients. Arch. Toxicol. 2002, 76, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Jandacek, R.J.; Heubi, J.E.; Buckley, D.D.; Khoury, J.C.; Turner, W.E.; Sjödin, A.; Olson, J.R.; Shelton, C.; Helms, K.; Bailey, T.D.; et al. Reduction of the body burden of PCBs and DDE by dietary intervention in a randomized trial. J. Nutr. Biochem. 2014, 25, 483–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, T.; Hirakawa, H.; Matsueda, T.; Nakagawa, R.; Takenaka, S.; Morita, K.; Narazaki, Y.; Fukamachi, K.; Tokiwa, H.; Takahashi, K.; et al. Therapeutic trials for promotion of fecal excretion of PCDFs and PCBs by the administration of cholestyramine in Yusho patients. Fukuoka Igaku Zasshi 1991, 82, 317–325. [Google Scholar] [PubMed]
- Takenaka, S.; Morita, K.; Takahashi, K. Stimulation of the fecal excretion of polychlorinated biphenyls (KC-600) by diets containing rice bran fiber and cholestyramine. Fukuoka Igaku Zasshi 1991, 82, 310–316. [Google Scholar]
- Morita, K.; Hirakawa, H.; Mtaueda, T.; Iida, T.; Tokiwa, H. Stimulating effect of dietary fiber on fecal excretion of polychlorinated dibenzofurans (PCDF) and polychlorinated dibenxo-p-dioxins (PCDD) in rats. Fukuoka Igaku Zasshi 1993, 84, 273–281. [Google Scholar]
- Iida, T.; Hiraka, H.; Matsueda, T.; Nakagawa, R.; Morita, K.; Tokiwa, H.; Tsuji, H.; Nakayama, J.; Hori, Y. Therapeutic trials for promotion of fecal excretion of PCDFs by the administration of rice bran fiber and cholestyramine in Yusho patients. Fukuoka Igaku Zasshi 1993, 84, 257–262. [Google Scholar]
- Tsuji, H.; Ikeda, K.; Nomiyama, K.; Fujishima, M. Effects of treatment with rice bran fiber and cholestyramine on clinical and laboratory findings in Yusho patients. Fukuoka Igaku Zasshi 1993, 84, 282–286. [Google Scholar]
- Iida, T.; Nakagawa, R.; Hirakawa, H.; Matsueda, T.; Morita, K.; Hamamura, K.; Nakayama, J.; Hori, Y.; Guo, Y.L.; Chang, F.M.; et al. Clinical trial of a combination of rice bran fiber and cholestyramine for promotion of fecal excretion of retained polychlorinated dibenzofuran and polychlorinated biphenyl in Yu-Cheng patients. Fukuoka Igaku Zasshi 1995, 86, 226–233. [Google Scholar]
- Todaka, T.; Honda, A.; Imaji, M.; Takao, Y.; Mitoma, C.; Furue, M. Effect of colestimide on the concentrations of polychlorinated dibenzo-p-dioxins, polychlorinated dizenzofurans, and polychlorinated biphenyls in blood of Yusho patients. Environ. Health 2016, 15, 63. [Google Scholar] [CrossRef] [Green Version]
- Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Mitoma, C.; Furue, M. Inhibition of aryl hydrocarbon receptor signaling and induction of NRF2-mediated antioxidant activity by cinnamaldehyde in human keratinocytes. J. Dermatol. Sci. 2017, 85, 36–43. [Google Scholar] [CrossRef]
- Uchi, H.; Tokunaga, S.; Mitoma, C.; Shibata, S.; Hamada, N.; Nakanishi, Y.; Kajiwara, J.; Yoshimura, T.; Furue, M. A clinical trial of kampo formulae for the treatment of symptoms of yusho, a poisoning caused by dioxins and related organochlorine compounds. Evid. Based Complement Alternat. Med. 2011, 2011, 589724. [Google Scholar] [CrossRef] [PubMed]
- Fuyuno, Y.; Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Tanaka, Y.; Mitoma, C.; Furue, M. Perillaldehyde inhibits AHR signaling and activates NRF2 antioxidant pathway in human keratinocytes. Oxid. Med. Cell Longev. 2018, 2018, 9524657. [Google Scholar] [CrossRef]
- Jung, D.; Konietzko, J.; Reill-Konietzko, G.; Muttray, A.; Zimmermann-Hölz, H.J.; Doss, M.; Beck, H.; Edler, L.; Kopp-Schneider, A. Porphyrin studies in TCDD-exposed workers. Arch. Toxicol. 1994, 68, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.M. Toxic effects of 2,3,7,8 tetrachlorodibenzo 1,4 dioxin in laboratory workers. Br. J. Ind. Med. 1975, 32, 49–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geusau, A.; Jurecka, W.; Nahavandi, H.; Schmidt, J.B.; Stingl, G.; Tschachler, E. Punctate keratoderma-like lesions on the palms and soles in a patient with chloracne: A new clinical manifestation of dioxin intoxication? Br. J. Dermatol. 2000, 143, 1067–1071. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furue, M.; Ishii, Y.; Tsukimori, K.; Tsuji, G. Aryl Hydrocarbon Receptor and Dioxin-Related Health Hazards—Lessons from Yusho. Int. J. Mol. Sci. 2021, 22, 708. https://doi.org/10.3390/ijms22020708
Furue M, Ishii Y, Tsukimori K, Tsuji G. Aryl Hydrocarbon Receptor and Dioxin-Related Health Hazards—Lessons from Yusho. International Journal of Molecular Sciences. 2021; 22(2):708. https://doi.org/10.3390/ijms22020708
Chicago/Turabian StyleFurue, Masutaka, Yuji Ishii, Kiyomi Tsukimori, and Gaku Tsuji. 2021. "Aryl Hydrocarbon Receptor and Dioxin-Related Health Hazards—Lessons from Yusho" International Journal of Molecular Sciences 22, no. 2: 708. https://doi.org/10.3390/ijms22020708
APA StyleFurue, M., Ishii, Y., Tsukimori, K., & Tsuji, G. (2021). Aryl Hydrocarbon Receptor and Dioxin-Related Health Hazards—Lessons from Yusho. International Journal of Molecular Sciences, 22(2), 708. https://doi.org/10.3390/ijms22020708