The Neosartorya fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida auris Biofilms

 ,
, 
 ,
, 

Abstract
:1. Introduction
2. Results
2.1. In Vitro Susceptibility of Planktonic Cells
2.2. In Vitro Susceptibility of Sessile Biofilm Cells
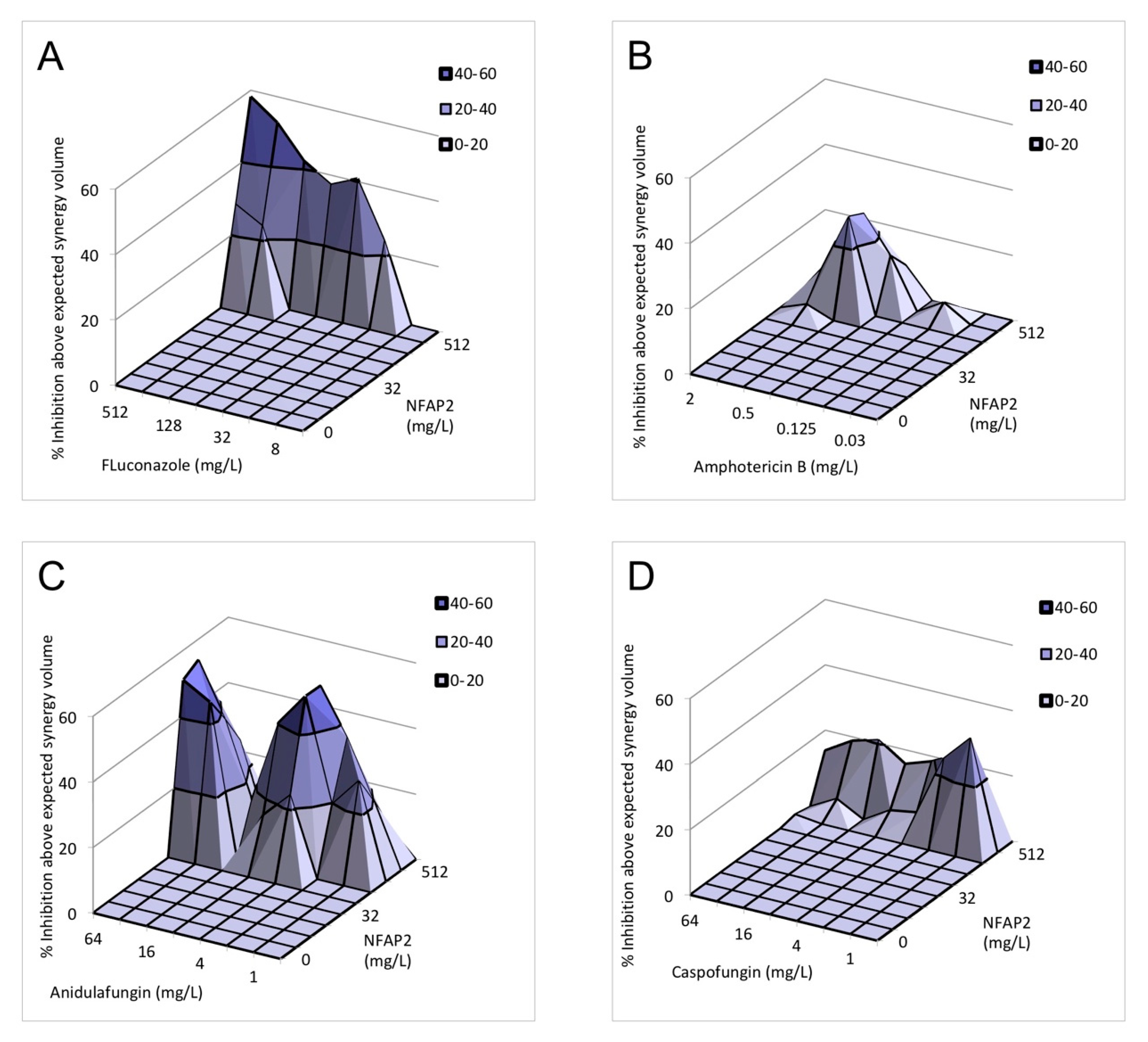
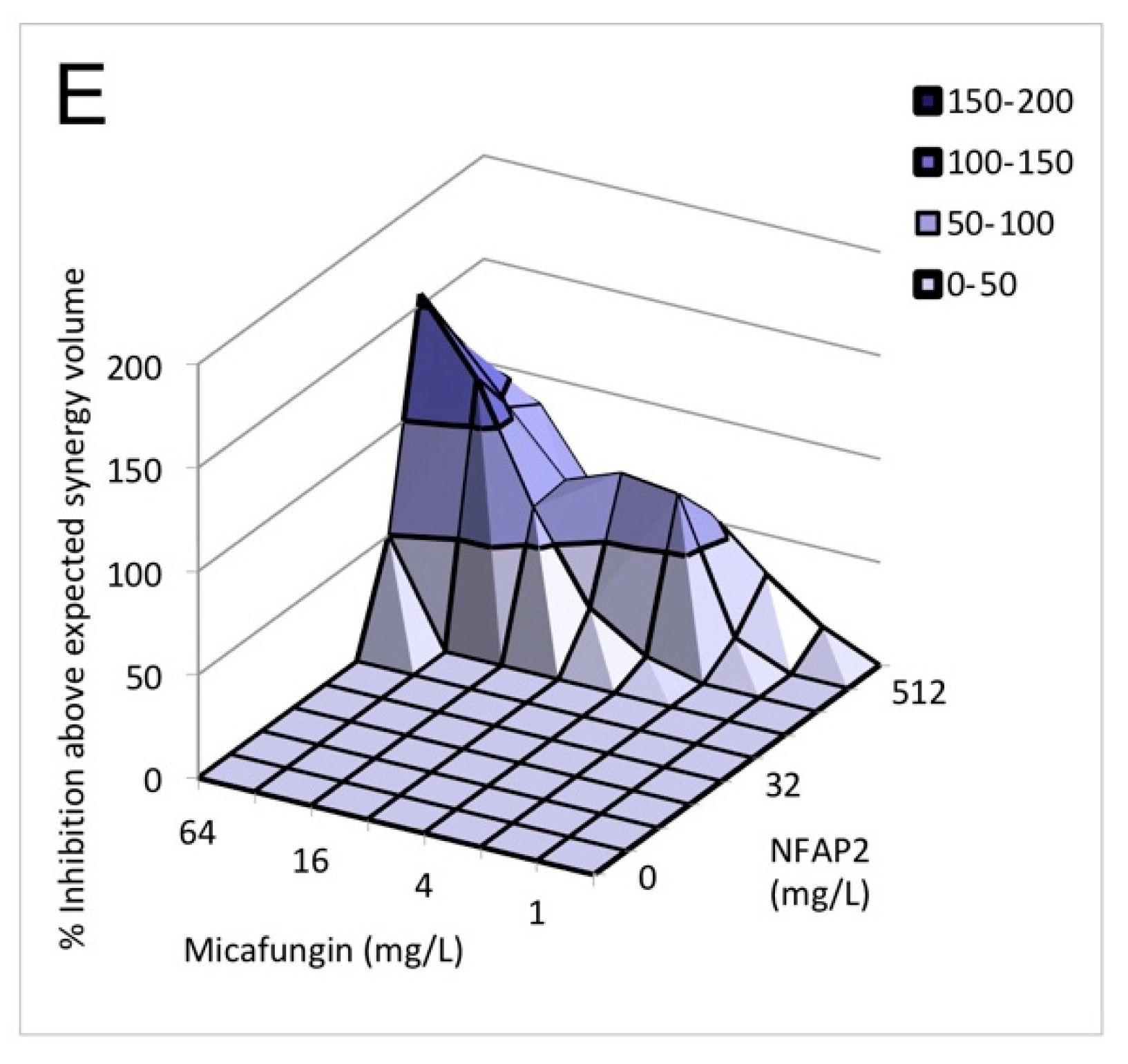
2.3. Nature of the NFAP2-Antifungal Drugs Interactions
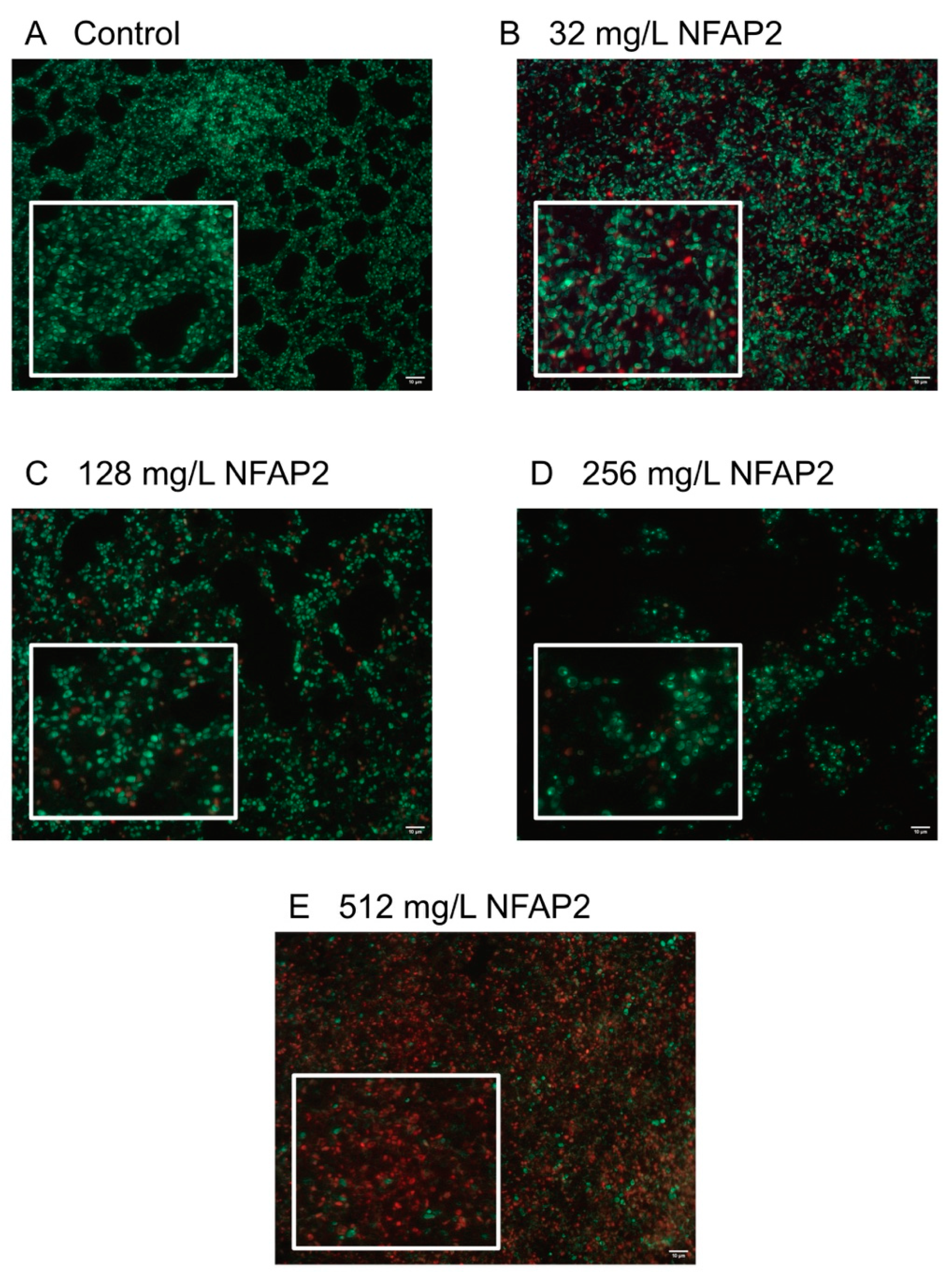
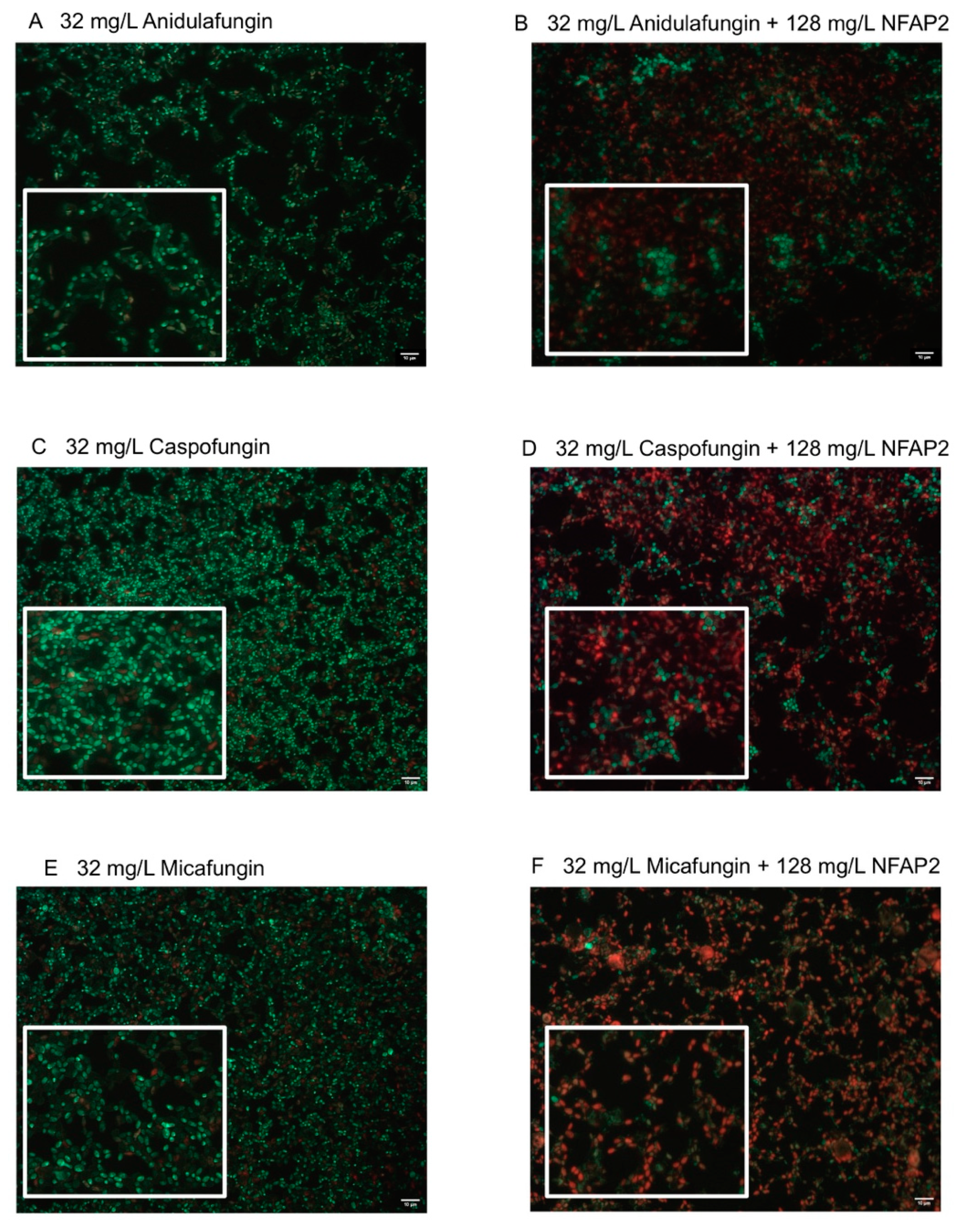
2.4. Fluorescence Viability Assay
3. Discussion
4. Materials and Methods
4.1. Isolates
4.2. Recombinant NFAP2 Production and Purification
4.3. In Vitro Susceptibility Testing of Planktonic Cells
4.4. Biofilm Development
4.5. Antifungal Susceptibility Testing of Biofilms
4.6. Evaluation of Interactions by Fractional Inhibitory Concentration Index (FICI) and Bliss Independence Model
4.7. Biofilm Viability Assay in the Presence or Absence of NFAP2
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, C.J.; Nguyen, M.H. Emergence of Candida auris: An International Call to Arms. Clin. Infect. Dis. 2017, 64, 141–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kean, R.; Brown, J.L.; Gülmez, D.; Ware, A.; Ramage, G. Candida auris: A Decade of Understanding of an Enigmatic Pathogenic Yeast. J. Fungi 2020, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.-C. First Three Reported Cases of Nosocomial Fungemia Caused by Candida auris. J. Clin. Microbiol. 2011, 49, 3139–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schelenz, S.; Hagen, F.; Rhodes, J.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First Hospital Outbreak of the Globally Emerging Candida auris in a European Hospital. Antimicrob. Resist. Infect. Control. 2016, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Taori, S.; Khonyongwa, K.; Hayden, I.; Athukorala, G.D.A.; Letters, A.; Fife, A.; Desai, N.; Borman, A.M. Candida auris Outbreak: Mortality, Interventions and Cost of Sustaining Control. J. Infect. 2019, 79, 601–611. [Google Scholar] [CrossRef]
- Sayeed, M.A.; Farooqi, J.; Jabeen, K.; Mahmood, S.F. Comparison of Risk Factors and Outcomes of Candida auris Candidemia with Non-Candida auris Candidemia: A Retrospective Study from Pakistan. Med. Mycol. 2020, 58, 721–729. [Google Scholar] [CrossRef]
- Horton, M.V.; Nett, J.E. Candida auris Infection and Biofilm Formation: Going Beyond the Surface. Curr. Clin. Microbiol. Rep. 2020, 7, 51–56. [Google Scholar] [CrossRef]
- Arensman, K.; Miller, J.L.; Chiang, A.; Mai, N.; Levato, J.; Lachance, E.; Anderson, M.; Beganovic, M.; Pena, J.D. Clinical Outcomes of Patients Treated for Candida auris Infections in a Multisite Health System, Illinois, USA. Emerg. Infect. Dis. 2020, 26, 876–880. [Google Scholar] [CrossRef]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Imbert, C.; Rammaert, B. What Could Be the Role of Antifungal Lock-Solutions? From Bench to Bedside. Pathogens 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, J.; Long, L.; Isham, N.; Mukherjee, P.K.; Disciullo, G.; Appelt, K.; Ghannoum, M.A. In Vitro and In Vivo Activity of a Novel Catheter Lock Solution against Bacterial and Fungal Biofilms. Antimicrob. Agents Chemother. 2018, 62, e00722-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mermel, L.A.; Allon, M.; Bouza, E.; Craven, D.E.; Flynn, P.; O’Grady, N.P.; Raad, I.I.; Rijnders, B.J.; Sherertz, R.J.; Warren, D.K. Clinical Practice Guidelines for the Diagnosis and Management of Intravascular Catheter-Related Infection: 2009 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 49, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Romera, D.; Aguilera-Correa, J.J.; Gadea, I.; Viñuela-Sandoval, L.; García-Rodríguez, J.; Esteband, J. Candida auris: A Comparison between Planktonic and Biofilm Susceptibility to Antifungal Drugs. J. Med. Microbiol. 2019, 68, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Galgóczy, L. NFAP2, a Novel Cysteine-Rich Anti-Yeast Protein from Neosartorya fischeri NRRL 181: Isolation and Characterization. AMB Express 2016, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, L.; Váradi, G.; Borics, A.; Batta, G.; Kele, Z.; Vendrinszky, Á.; Tóth, R.; Ficze, H.; Tóth, G.K.; Vágvölgyi, C.; et al. Anti-Candidal Activity and Functional Mapping of Recombinant and Synthetic Neosartorya fischeri Antifungal Protein 2 (NFAP2). Front. Microbiol. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovács, R.; Holzknecht, J.; Hargitai, Z.; Papp, C.; Farkas, A.; Borics, A.; Tóth, L.; Váradi, G.; Tóth, G.K.; Kovács, I.; et al. In Vivo Applicability of Neosartorya fischeri Antifungal Protein 2 (NFAP2) in Treatment of Vulvovaginal Candidiasis. Antimicrob. Agents Chemother. 2019, 63, e01777-18. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Antifungal Susceptibility Testing and Interpretation. 2020. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 29 May 2020).
- Ghannoum, M.A.; Isham, N.; Angulo, D.; Borroto-Esoda, K.; Barat, S.; Long, L. Efficacy of Ibrexafungerp (SCY-078) against Candida auris in an In Vivo Guinea Pig Cutaneous Infection Model. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef]
- Zhu, Y.; Kilburn, S.; Kapoor, M.; Chaturvedi, S.; Shaw, K.J.; Chaturvedi, V. In Vitro Activity of Manogepix against Multidrug-Resistant and Panresistant Candida auris from the New York Outbreak. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Wiederhold, N.P.; Lockhart, S.R.; Najvar, L.K.; Berkow, E.L.; Jaramillo, R.; Olivo, M.; Garvey, E.P.; Yates, C.M.; Schotzinger, R.J.; Catano, G.; et al. The Fungal Cyp51-Specific Inhibitor VT-1598 Demonstrates In Vitro and In Vivo Activity against Candida auris. Antimicrob. Agents Chemother. 2019, 63, e02233-18. [Google Scholar] [CrossRef] [Green Version]
- Tóth, Z.; Forgács, L.; Locke, J.B.; Kardos, G.; Nagy, F.; Kovács, R.; Szekely, A.; Borman, A.M.; Majoros, L. In Vitro Activity of Rezafungin against Common and Rare Candida Species and Saccharomyces cerevisiae. J. Antimicrob. Chemother. 2019, 74, 3505–3510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Cruz, N.; Reitzel, R.A.; Rosenblatt, J.; Chaftari, A.-M.; Dib, R.W.; Hachem, R.; Kontoyiannis, D.P.; Raad, I.I. Nitroglycerin-Citrate-Ethanol Catheter Lock Solution Is Highly Effective for In Vitro Eradication of Candida auris Biofilm. Antimicrob. Agents Chemother. 2019, 63, e00299-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, G.; Chaturvedi, A.K.; Wormley, F.L., Jr.; Wiederhold, N.P.; Patterson, H.P.; Patterson, T.F.; Lopez-Ribot, J.L. Screening a Repurposing Library for Inhibitors of Multidrug-Resistant Candida auris Identifies Ebselen as a Repositionable Candidate for Antifungal Drug Development. Antimicrob. Agents Chemother. 2018, 62, e01084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, T.L.; Rossato, L.; De Freitas, A.L.D.; Meis, J.F.; Lopes, L.B.; Colombo, A.L.; Ishida, K. Miltefosine as an Alternative Strategy in the Treatment of the Emerging Fungus Candida auris. Int. J. Antimicrob. Agents 2020, 56, 106049. [Google Scholar] [CrossRef] [PubMed]
- Nagy, F.; Tóth, Z.; Daróczi, L.; Székely, A.; Borman, A.M.; Majoros, L.; Kovács, R. Farnesol Increases the Activity of Echinocandins against Candida auris Biofilms. Med. Mycol. 2019, 58, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Muñoz, R.; Lopez, F.D.; Lopez-Ribot, J.L. Silver Nanoantibiotics Display Strong Antifungal Activity against the Emergent Multidrug-Resistant Yeast Candida auris under Both Planktonic and Biofilm Growing Conditions. Front. Microbiol. 2020, 11, 1673. [Google Scholar] [CrossRef]
- Vazquez-Muñoz, R.; Lopez, F.D.; Lopez-Ribot, J.L. Bismuth Nanoantibiotics Display Anticandidal Activity and Disrupt the Biofilm and Cell Morphology of the Emergent Pathogenic Yeast Candida auris. Antibiotics 2020, 9, 461. [Google Scholar] [CrossRef]
- Reitzel, R.A.; Rosenblatt, J.; Gerges, B.Z.; Vargas-Cruz, N.; Raad, I.I. Minocycline-EDTA-Ethanol Antimicrobial Catheter Lock Solution Is Highly Effective In Vitro for Eradication of Candida auris Biofilms. Antimicrob. Agents Chemother. 2020, 64, e02146-19. [Google Scholar] [CrossRef]
- Galgóczy, L.; Yap, A.; Marx, F. Cysteine-Rich Antifungal Proteins from Filamentous Fungi Are Promising Bioactive Natural Compounds in Anti-Candida Therapy. Isr. J. Chem. 2019, 59, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Mas, C.D.; Rossato, L.; Shimizu, T.; Oliveira, E.B.; Da Silva, J.P.I.; Meis, J.F.; Colombo, A.L.; Hayashi, M.A.F. Effects of the Natural Peptide Crotamine from a South American Rattlesnake on Candida auris, an Emergent Multidrug Antifungal Resistant Human Pathogen. Biomolecules 2019, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Van Eijk, M.; Boerefijn, S.; Cen, L.; Rosa, M.; Morren, M.J.H.; Van Der Ent, C.K.; Kraak, B.; Dijksterhuis, J.; Valdes, I.D.; Haagsman, H.P.; et al. Cathelicidin-Inspired Antimicrobial Peptides as Novel Antifungal Compounds. Med. Mycol. 2020, 58, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubiczek, D.; Raber, H.; García, M.G.; Vicente, F.E.M.; Ständker, L.; Otero-González, A.; Rosenau, F. Derivates of the Antifungal Peptide Cm-p5 Inhibit Development of Candida auris Biofilms In Vitro. Antibiotics 2020, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Comparative Pathogenicity of United Kingdom Isolates of the Emerging Pathogen Candida auris and Other Key Pathogenic Candida Species. mSphere 2016, 1, e00189-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Isolates of the Emerging Pathogen Candida auris Present in the UK Have Several Geographic Origins. Med. Mycol. 2017, 55, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Approved Standard, 3rd ed.; M27-A3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Kovács, R.; Bozó, A.; Gesztelyi, R.; Domán, M.; Kardos, G.; Nagy, F.; Tóth, Z.; Majoros, L. Effect of Caspofungin and Micafungin in Combination with Farnesol against Candida parapsilosis Biofilms. Int. J. Antimicrob. Agents 2016, 47, 304–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, F.; Vitális, E.; Jakab, Á.; Borman, A.M.; Forgács, L.; Tóth, Z.; Majoros, L.; Kovács, R. In Vitro and In Vivo Effect of Exogenous Farnesol Exposure against Candida auris. Front. Microbiol. 2020, 11, 957. [Google Scholar] [CrossRef]
- Hawser, S. Adhesion of Different Candida spp. to Plastic: XTT Formazan Determinations. J. Med. Vet. Mycol. 1996, 34, 407–410. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Verweij, P.E.; Dorsthorst, D.T.A.T.; Meis, J.F.; Mouton, J.W. Assessing In Vitro Combinations of Antifungal Drugs against Yeasts and Filamentous Fungi: Comparison of Different Drug Interaction Models. Med. Mycol. 2005, 43, 133–152. [Google Scholar] [CrossRef] [Green Version]
- Prichard, M.N.; Shipman, C.J. A Three-Dimensional Model to Analyze Drug-Drug Interactions. Antivir. Res. 1990, 14, 181–205. [Google Scholar] [CrossRef] [Green Version]
- Rhoden, E.; Ng, T.F.F.; Campagnoli, R.; Nix, W.A.; Konopka-Anstadt, J.; Selvarangan, R.; Briesach, L.; Oberste, M.S.; Weldon, W.C. Antifungal Triazole Posaconazole Targets an Early Stage of the Parechovirus A3 Life Cycle. Antimicrob. Agents Chemother. 2020, 64, e02372-19. [Google Scholar] [CrossRef] [Green Version]
- Basas, J.; Morer, A.; Ratia, C.; Martín, M.T.; Del Pozo, J.L.; Gomis, X.; Rojo-Molinero, E.; Torrents, E.; Almirante, B.; Gavaldà, J. Efficacy of Anidulafungin in the Treatment of Experimental Candida parapsilosis Catheter Infection Using an Antifungal-Lock Technique. J. Antimicrob. Chemother. 2016, 71, 2895–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, L.; Agrappi, S.; Bortolin, M.; Toscano, M.; Romanò, C.L.; De Vecchi, E. How to Study Biofilms after Microbial Colonization of Materials Used in Orthopaedic Implants. Int. J. Mol. Sci. 2016, 17, 293. [Google Scholar] [CrossRef] [PubMed]
- Kagan, S.; Jabbour, A.; Sionov, E.; Alquntar, A.A.; Steinberg, D.; Srebnik, M.; Nir-Paz, R.; Weiss, A.; Polacheck, I. Anti-Candida albicans biofilm effect of novel heterocyclic compounds. J. Antimicrob. Chemother. 2014, 69, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Isolate | Median MIC (Range) of Drug Used (50% OD492 Reduction in Metabolic Activity) | |||
|---|---|---|---|---|---|
| Alone | In Combination | ||||
| Drug (mg/L) | NFAP2 (mg/L) | Drug (mg/L) | NFAP2 (mg/L) | ||
| FLU | 10 | >512 a | >512 a | 32 | 256 (256–512) |
| 12 | >512 a | >512 a | 16 (16–32) | 256 | |
| 20 | >512 a | >512 a | 32 (32–64) | 256 (256–512) | |
| 27 | >512 a | >512 a | 8 (8–16) | 512 (256–512) | |
| 82 | >512 a | >512 a | 32 (32–128) | 512 | |
| AMB | 10 | 1 | 512 (256–512) | 0.25 | 128 |
| 12 | 1 | >512 a | 0.03 (0.03–0.06) | 64 (64–128) | |
| 20 | 1 | >512 a | 0.03 (0.03–0.06) | 16 (16–64) | |
| 27 | 1 | 512 | 0.25 | 64 | |
| 82 | 2 | 512 | 0.03 (0.03–0.06) | 4 (4–16) | |
| ANI | 10 | 16 (16–32) | >512 a | 1 | 128 (128–256) |
| 12 | >64 b | >512 a | 1 | 32 | |
| 20 | >64 b | >512 a | 1 | 128 (128–256) | |
| 27 | 16 | 512 | 1 | 4 (4–8) | |
| 82 | >64 b | 512 | 1 | 64 (64–128) | |
| CAS | 10 | >64 b | 512 (128–512) | 32 | 32 (32–64) |
| 12 | >64 b | >512 a (512–>512) | 1 (1–2) | 64 | |
| 20 | >64 b | 512 (256–512) | 1 | 32 (32–64) | |
| 27 | >64 b | 512 (128–512) | 32 | 32 (32–64) | |
| 82 | >64 b | 512 (256–512) | 1 (1–4) | 32 | |
| MICA | 10 | >64 b | >512 a | 2 (2–4) | 256 |
| 12 | >64 b | >512 a | 1 | 4 | |
| 20 | >64 b | >512 a | 1 | 256 (128–256) | |
| 27 | >64 b | >512 a | 1 | 4 | |
| 82 | >64 b | >512 a | 1 | 32 (32–64) | |
| Drug | Isolate | FICI | MacSynergy II Analysis | ||||
|---|---|---|---|---|---|---|---|
| Median (Range) of FICI | Interaction | Individual Log Volume of Isolates, (Synergy/Antagonism μM2%) | Interaction Based on Individual Log Volume | Cumulative Log Volume of Five Isolates, (Synergy/Antagonism μM2%) | Interaction Based on Cumulative Log Volume | ||
| FLU | 10 | 0.5 (0.5–0.53) | Synergy | 72.43/−126.35 | Antagonism for most combinations | 17.54/0 | Synergy |
| 12 | 0.312 | Synergy | 12.31/0 | Synergy | |||
| 20 | 0.312 (0.312–0.375) | Synergy | 210.17/−3.67 | Synergy for most combinations | |||
| 27 | 0.375 (0.375–0.625) | Synergy | 70/−78.26 | Antagonism for most combinations | |||
| 82 | 0.375 (0.375–0.625) | Synergy | 119.31/−11.47 | Synergy for most combinations | |||
| AMB | 10 | 0.5 (0.5–0.75) | Synergy | 53.9/−42.3 | Synergy for most combinations | 2.16/0 | Synergy |
| 12 | 0.312 (0.312–0.375) | Synergy | 196.99/−31.39 | Synergy for most combinations | |||
| 20 | 0.155 (0.155–0.185) | Synergy | 126.38/−4.52 | Synergy for most combinations | |||
| 27 | 0.375 | Synergy | 46.51/−27.26 | Synergy for most combinations | |||
| 82 | 0.25 | Synergy | 75.39/−12.25 | Synergy for most combinations | |||
| ANI | 10 | 0.375 (0.25–0.5) | Synergy | 127.59/−18.52 | Synergy for most combinations | 33.31/0 | Synergy |
| 12 | 0.037 | Synergy | 371.84/−2.43 | Synergy for most combinations | |||
| 20 | 0.185 (0.185–0.312) | Synergy | 154.33/−40.86 | Synergy for most combinations | |||
| 27 | 0.069 (0.069–0.09) | Synergy | 71.52/−10.83 | Synergy for most combinations | |||
| 82 | 0.312 (0.185–0.312) | Synergy | 148.15/−7.3 | Synergy for most combinations | |||
| CAS | 10 | 0.375 (0.375–0.75) | Synergy | 8.64/−33.23 | Antagonism for most combinations | 10.72/0 | Synergy |
| 12 | 0.067 (0.067–0.14) | Synergy | 235.24/0 | Synergy | |||
| 20 | 0.075 (0.075–0.257) | Synergy | 22.23/−17.48 | Synergy for most combinations | |||
| 27 | 0.375 (0.375–0.75) | Synergy | 14.76/−44.63 | Antagonism for most combinations | |||
| 82 | 0.122 | Synergy | 20.2/−11.59 | Synergy for most combinations | |||
| MICA | 10 | 0.375 | Synergy | 164.37/−99.58 | Synergy for most combinations | 111.19/0 | Synergy |
| 12 | 0.064 | Synergy | 277.54/0 | Synergy | |||
| 20 | 0.375 | Synergy | 378.15/−17.51 | Synergy for most combinations | |||
| 27 | 0.253 | Synergy | 100.94/−41.11 | Synergy for most combinations | |||
| 82 | 0.132 | Synergy | 212.19/−18.37 | Synergy for most combinations | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, R.; Nagy, F.; Tóth, Z.; Forgács, L.; Tóth, L.; Váradi, G.; Tóth, G.K.; Vadászi, K.; Borman, A.M.; Majoros, L.; et al. The Neosartorya fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida auris Biofilms. Int. J. Mol. Sci. 2021, 22, 771. https://doi.org/10.3390/ijms22020771
Kovács R, Nagy F, Tóth Z, Forgács L, Tóth L, Váradi G, Tóth GK, Vadászi K, Borman AM, Majoros L, et al. The Neosartorya fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida auris Biofilms. International Journal of Molecular Sciences. 2021; 22(2):771. https://doi.org/10.3390/ijms22020771
Chicago/Turabian StyleKovács, Renátó, Fruzsina Nagy, Zoltán Tóth, Lajos Forgács, Liliána Tóth, Györgyi Váradi, Gábor K. Tóth, Karina Vadászi, Andrew M. Borman, László Majoros, and et al. 2021. "The Neosartorya fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida auris Biofilms" International Journal of Molecular Sciences 22, no. 2: 771. https://doi.org/10.3390/ijms22020771
APA StyleKovács, R., Nagy, F., Tóth, Z., Forgács, L., Tóth, L., Váradi, G., Tóth, G. K., Vadászi, K., Borman, A. M., Majoros, L., & Galgóczy, L. (2021). The Neosartorya fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida auris Biofilms. International Journal of Molecular Sciences, 22(2), 771. https://doi.org/10.3390/ijms22020771