Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study

Abstract
:1. Introduction
2. New Metabolites Isolated from Plant Endophytes
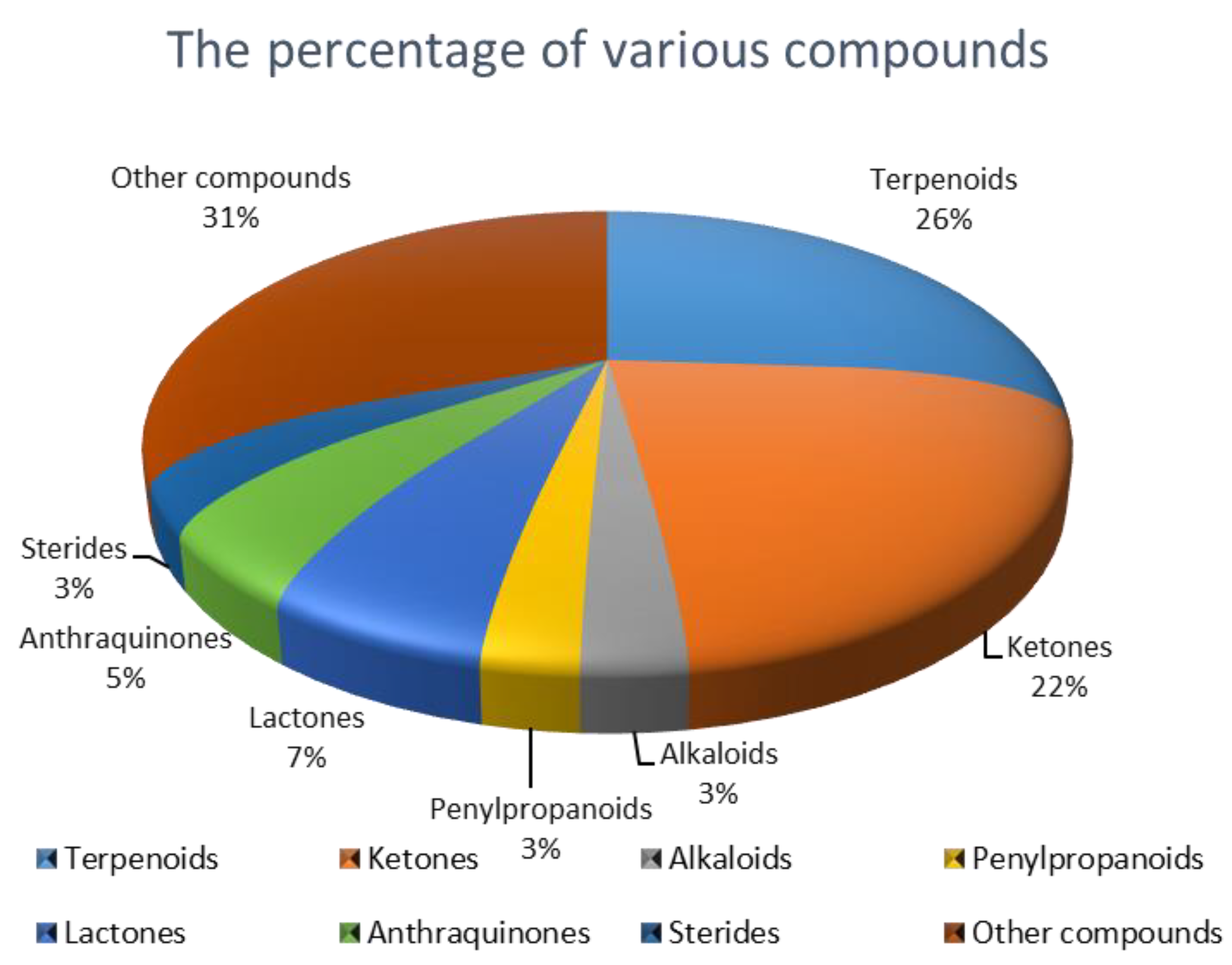
2.1. Terpenoids
2.1.1. Sesquiterpenoids and Their Derivatives
2.1.2. Diterpenoids
2.1.3. Other Terpenoids
2.2. Ketone Compounds
2.2.1. Polyketides
2.2.2. Other Ketones
2.3. Alkaloids and Their Derivatives
2.4. Penylpropanoids and Their Derivatives
2.5. Lactones
2.6. Anthraquinones
2.7. Sterides
2.8. Other Types of Compounds
3. Biological Activity
3.1. Antimicrobial Activity
3.1.1. Antifungal Activity
3.1.2. Antibacterial Activity
3.1.3. Antiviral Activity
3.2. Cytotoxic Activity or Anticancer
3.3. Other Activities
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Elshafie, H.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Camele, I.; Bufo, S.A. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res.—Bollettino della Società Italiana di Biologia Sperimentale 2018, 90, 96–103. [Google Scholar] [CrossRef]
- Camele, I.; Elshafie, H.S.; Caputo, L.; Sakr, S.H.; De Feo, V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res.—Bollettino della Società Italiana di Biologia Sperimentale 2019, 92, 39–45. [Google Scholar] [CrossRef]
- Kong, D.W.; Niu, R.C.; Mao, Y.Z.; Liu, L.L. Research progress on active metabolites of endophytes. Keshan Branch Heilongjiang Acad. Agric. Sci. 2019, 12, 151–154. [Google Scholar]
- Ding, W.J.; Wang, S.S.; Ren, J.Q.; Li, G.; Zhan, J.P. Progress on plant endophyte. Curr. Biotechnol. 2015, 5, 425–428. [Google Scholar]
- J in, J.; Zhao, Q.; Zhang, X.M.; Li, W.J. Research progress on bioactive products from endophytes. J. Microbiol. 2018, 38, 103–113. [Google Scholar]
- Elshafie, H.; Camele, I.; Sofo, A.; Mazzone, G.; Caivano, M.; Masi, S.; Caniani, D. Mycoremediation effect of Trichoderma harzianum strain T22 combined with ozonation in diesel-contaminated sand. Chemosphere 2020, 252, 126597. [Google Scholar] [CrossRef]
- Vogl, A. Mehlund die andiron mehlproduktdercerealien und leguminosen. Nahrunsm Unters Hug Waren 1898, 12, 25–29. [Google Scholar]
- Dar, R.A.; Shahnawaz, M.; Qazi, P.H. Fungal Endophytes: An Overview; LAP LAMBERT Academic Publishing: Beau Bassin, Mauritius, 2017; pp. 40–42. [Google Scholar]
- Yuan, C.; Ding, G.; Wang, H.-Y.; Guo, Y.-H.; Shang, H.; Ma, X.-J.; Zou, Z.-M. Polyketide-Terpene Hybrid Metabolites from an Endolichenic Fungus Pestalotiopsis sp. BioMed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Zhang, L.; Niaz, S.I.; Khan, D.; Wang, Z.; Zhu, Y.; Zhou, H.; Lin, Y.; Li, J.; Liu, L. Induction of Diverse Bioactive Secondary Metabolites from the Mangrove Endophytic Fungus Trichoderma sp. (Strain 307) by Co-Cultivation with Acinetobacter johnsonii (Strain B2). Mar. Drugs 2017, 15, 35. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Wu, Z.; Tan, D.; Yang, J.; Zhou, Q.; Zeng, F.; Zhang, M.; Bie, Q.; Chen, C.; Xue, Y.; et al. Atrichodermones A–C, three new secondary metabolites from the solid culture of an endophytic fungal strain, Trichoderma atroviride. Fitoterapia 2017, 123, 18–22. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Heering, C.; Janiak, C.; Lin, W.; Liu, Z.; Proksch, P. Inducing new secondary metabolites through co-cultivation of the fungus Pestalotiopsis sp. with the bacterium Bacillus subtilis. Tetrahedron Lett. 2017, 58, 257–261. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Kristiana, P.; Nurjanto, H.H.; Momma, H.; Kwon, E.; Ashitani, T.; Tawaraya, K.; Murayama, T.; Koseki, T.; Furuno, H.; et al. Nectrianolins A, B, and C, new metabolites produced by endophytic fungus Nectria pseudotrichia 120-1NP. Tetrahedron Lett. 2017, 58, 4082–4086. [Google Scholar] [CrossRef]
- Zhou, Q.-Y.; Yang, X.-Q.; Zhang, Z.-X.; Wang, B.-Y.; Hu, M.; Yang, Y.-B.; Zhou, H.; Ding, Z. New azaphilones and tremulane sesquiterpene from endophytic Nigrospora oryzae cocultured with Irpex lacteus. Fitoterapia 2018, 130, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.-J.; Zhang, S.-B.; Xian, P.-J.; Wu, X.; Yang, D.-F.; Fu, H.; Yang, X.-L. Emericellins A and B: Two sesquiterpenoids with an unprecedented tricyclo[4,4,2,1]hendecane scaffold from the liquid cultures of endophytic fungus Emericella sp. XL 029. Fitoterapia 2018, 131, 55–58. [Google Scholar] [CrossRef]
- Yang, H.-X.; Ai, H.-L.; Feng, T.; Wang, W.-X.; Wu, B.; Zheng, Y.; Sun, H.; He, J.; Li, Z.-H.; Liu, J.-K. Trichothecrotocins A–C, Antiphytopathogenic Agents from Potato Endophytic FungusTrichothecium crotocinigenum. Org. Lett. 2018, 20, 8069–8072. [Google Scholar] [CrossRef]
- Kong, Z.; Jing, R.; Wu, Y.; Guo, Y.; Geng, Y.; Ji, J.; Qin, L.-P.; Zheng, C.-J. Trichodermadiones A and B from the solid culture of Trichoderma atroviride S361, an endophytic fungus in Cephalotaxus fortunei. Fitoterapia 2018, 127, 362–366. [Google Scholar] [CrossRef]
- Wang, P.; Yu, J.-H.; Zhu, K.; Wang, Y.; Jiang, C.-S.; Jiang, C.-S.; Dai, J.-G.; Wu, J.; Zhang, H. Phenolic bisabolane sesquiterpenoids from a Thai mangrove endophytic fungus, Aspergillus sp. xy02. Fitoterapia 2018, 127, 322–327. [Google Scholar] [CrossRef]
- Xiao, J.; Lin, L.-B.; Hu, J.-Y.; Duan, D.-Z.; Shi, W.; Zhang, Q.; Han, W.-B.; Wang, L.; Wang, X.-L. Pestalustaines A and B, unprecedented sesquiterpene and coumarin derivatives from endophytic fungus Pestalotiopsis adusta. Tetrahedron Lett. 2018, 59, 1772–1775. [Google Scholar] [CrossRef]
- Jiang, C.-X.; Li, J.; Zhang, J.-M.; Jin, X.-J.; Yu, B.; Fang, J.; Wu, Q.-X. Isolation, Identification, and Activity Evaluation of Chemical Constituents from Soil Fungus Fusarium avenaceum SF-1502 and Endophytic Fungus Fusarium proliferatum AF-04. J. Agric. Food Chem. 2019, 67, 1839–1846. [Google Scholar] [CrossRef]
- Song, Y.-P.; Miao, F.-P.; Liu, X.-H.; Yin, X.-L.; Ji, N.-Y. Seven chromanoid norbisabolane derivatives from the marine-alga-endophytic fungus Trichoderma asperellum A-YMD-9-2. Fitoter 2019, 135, 107–113. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, X.; Yu, M.; Yu, Y.; Guo, Z.; Gong, T.; Niu, S.; Qin, J.; Zou, Z.; Ding, G. Sesquiterpenoids and mycotoxin swainsonine from the locoweed endophytic fungus Alternaria oxytropis. Phytochemistry 2019, 164, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Fan, W.; Guo, H.; Huang, C.; Yan, Z.; Long, Y. Two new secondary metabolites from the mangrove endophytic fungus Pleosporales sp. SK7. Nat. Prod. Res. 2020, 34, 2919–2925. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Qin, D.; Song, H.C.; Gao, T.; Zuo, S.; Yan, X.; Wang, J.-Q.; Ding, X.; Di, Y.-T.; Dong, J. Irpexlacte A-D, four new bioactive metabolites of endophytic fungus Irpex lacteus DR10-1 from the waterlogging tolerant plant Distylium chinense. Phytochemistry Lett. 2019, 32, 151–156. [Google Scholar] [CrossRef]
- Shi, X.-S.; Meng, L.-H.; Li, X.-M.; Li, X.; Wang, D.-J.; Li, H.-L.; Zhou, X.-W.; Wang, B.-G. Trichocadinins B–G: Antimicrobial Cadinane Sesquiterpenes from Trichoderma virens QA-8, an Endophytic Fungus Obtained from the Medicinal Plant Artemisia argyi. J. Nat. Prod. 2019, 82, 2470–2476. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hu, Y.-W.; Qu, W.; Chen, M.-H.; Zhou, L.-S.; Bi, Q.-R.; Luo, J.-G.; Liu, W.-Y.; Feng, F.; Zhang, J. Cytotoxic and neuroprotective activities of constituents from Alternaria alternate, a fungal endophyte of Psidium littorale. Bioorganic Chem. 2019, 90, 103046. [Google Scholar] [CrossRef]
- Li, H.T.; Tang, L.H.; Liu, T.; Yang, R.N.; Yang, Y.B.; Zhou, H.; Ding, Z.T. Protoilludane-type sesquiterpenoids from Armillaria sp. by co-culture with the endophytic fungus Epicoccum sp. associated with Gastrodia elata. Bioorg. Chem. 2019, 95, 103503. [Google Scholar] [CrossRef]
- Kemda, P.N.; Akone, S.H.; Tontsa, A.T.; Zhen, L.; Muller, W.E.; Proksch, P.; Nkengfack, A.E. Colletotrin: A sesquiterpene lactone from the endophytic fungus Colletotrichum gloeosporioides associated with Trichilia monadelpha. Zeitschrift für Naturforschung B 2017, 72, 697–703. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Xia, G.-Y.; Wang, L.-Y.; Ge, G.; Zhang, H.-W.; Zhang, J.-F.; Wu, Y.-Z.; Lin, S. Purpurolide A, 5/5/5 Spirocyclic Sesquiterpene Lactone in Nature from the Endophytic Fungus Penicillium purpurogenum. Org. Lett. 2018, 20, 7341–7344. [Google Scholar] [CrossRef]
- Chen, J.; Bai, X.; Hua, Y.; Zhang, H.; Wang, H. Fusariumins C and D, two novel antimicrobial agents from Fusarium oxysporum ZZP-R1 symbiotic on Rumex madaio Makino. Fitoterapia 2019, 134, 1–4. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Kristiana, P.; Aboshi, T.; Murayama, T.; Tawaraya, K.; Koseki, T.; Kurisawa, N.; Kimura, K.-I.; Shiono, Y. New isocoumarins, naphthoquinones, and a cleistanthane-type diterpene from Nectria pseudotrichia 120-1NP. Fitoterapia 2018, 127, 356–361. [Google Scholar] [CrossRef]
- Zhao, J.-C.; Wang, Y.-L.; Zhang, T.-Y.; Chen, Z.-J.; Yang, T.-M.; Wu, Y.-Y.; Sun, C.-P.; Ma, X.-C.; Zhang, Y.-X. Indole diterpenoids from the endophytic fungus Drechmeria sp. as natural antimicrobial agents. Phytochemistry 2018, 148, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-C.; Luan, Z.-L.; Liang, J.-H.; Cheng, Z.-B.; Sun, C.-P.; Wang, Y.-L.; Zhang, M.-Y.; Zhang, T.-Y.; Wang, Y.; Yang, T.-M.; et al. Drechmerin H, a novel 1(2), 2(18)-diseco indole diterpenoid from the fungus Drechmeria sp. as a natural agonist of human pregnane X receptor. Bioorganic Chem. 2018, 79, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Song, J.H.; Lee, D.; Lee, C.; Kim, S.; Kang, K.S.; Lee, J.; Shim, S.H. Neuroprotective Secondary Metabolite Produced by an Endophytic Fungus, Neosartorya fischeri JS0553, Isolated from Glehnia littoralis. J. Agric. Food Chem. 2019, 67, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Elsbaey, M.; Tanaka, C.; Miyamoto, T. New secondary metabolites from the mangrove endophytic fungus Aspergillus versicolor. Phytochem. Lett. 2019, 32, 70–76. [Google Scholar] [CrossRef]
- Bao, S.-S.; Liu, H.-H.; Zhang, X.-Q.; Liu, C.-X.; Li, X.-C.; Guo, Z.-Y. Xylaroisopimaranin A, a New Isopimarane Derivative from an Endophytic Fungus Xylaralyce sp. Nat. Prod. Sci. 2019, 25, 228–232. [Google Scholar] [CrossRef]
- Wang, H.; Umeokoli, B.O.; Eze, P.M.; Heering, C.; Janiak, C.; Müller, W.E.; Orfali, R.S.; Hartmann, R.; Dai, H.; Lin, W.; et al. Secondary metabolites of the lichen-associated fungus Apiospora montagnei. Tetrahedron Lett. 2017, 58, 1702–1705. [Google Scholar] [CrossRef]
- Cai, R.; Jiang, H.; Mo, Y.; Guo, H.; Li, C.; Long, Y.; Zang, Z.; She, Z. Ophiobolin-Type Sesterterpenoids from the Mangrove Endophytic Fungus Aspergillus sp. ZJ-68. J. Nat. Prod. 2019, 82, 2268–2278. [Google Scholar] [CrossRef]
- Qin, D.; Shen, W.; Wang, J.-Q.; Han, M.; Chai, F.; Duan, X.; Yan, X.; Guo, J.; Gao, T.; Zuo, S.; et al. Enhanced production of unusual triterpenoids from Kadsura angustifolia fermented by a symbiont endophytic fungus, Penicillium sp. SWUKD4.1850. Phytochemistry 2019, 158, 56–66. [Google Scholar] [CrossRef]
- Ma, K.-L.; Wei, W.-J.; Li, H.-Y.; Song, Q.-Y.; Dong, S.-H.; Gao, K. Meroterpenoids with diverse ring systems and dioxolanone-type secondary metabolites from Phyllosticta capitalensis and their phytotoxic activity. Tetrahedron 2019, 75, 4611–4619. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.; Li, H.; Li, S.; Tan, H.; Liu, Z.; Li, D.; Liu, H.; Zhang, W. Four new metabolites from the endophytic fungus Diaporthe lithocarpus A740. Fitoterapia 2019, 137, 104260. [Google Scholar] [CrossRef]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic Modulation of Endophytic Eupenicillium sp. LG41 by a Histone Deacetylase Inhibitor for Production of Decalin-Containing Compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-W.; Wang, W.-G.; Li, A.; Yan, B.-C.; Chen, R.; Li, X.-N.; Du, X.; Sun, H.-D.; Pu, J.-X. Polyketides from the endophytic fungus Phomopsis sp. sh917 by using the one strain/many compounds strategy. Tetrahedron 2017, 73, 3577–3584. [Google Scholar] [CrossRef]
- Wang, J.; Liang, R.; Liao, S.-R.; Yang, B.; Tu, Z.-C.; Lin, X.-P.; Wang, B.-G.; Liu, Y. Vaccinols J–S, ten new salicyloid derivatives from the marine mangrove-derived endophytic fungus Pestalotiopsis vaccinii. Fitoterapia 2017, 120, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Liu, X.; Mo, T.; Li, S.-S.; Wang, J.; Shi, X.-P.; Wang, X.-H.; Zhu, Z.-X.; Zhao, Y.-F.; Jin, H.-W.; et al. Nitric oxide inhibitory polyketides from Penicillium chrysogenum MT-12, an endophytic fungus isolated from Huperzia serrata. Fitoterapia 2017, 123, 35–43. [Google Scholar] [CrossRef]
- Kamdem, R.S.; Pascal, W.; Rehberg, N.; Van Geelen, L.; Höfert, S.-P.; Knedel, T.; Janiak, C.; Sureechatchaiyan, P.; Kassack, M.U.; Lin, W.; et al. Metabolites from the endophytic fungus Cylindrocarpon sp. isolated from tropical plant Sapium ellipticum. Fitoterapia 2018, 128, 175–179. [Google Scholar] [CrossRef]
- Hu, H.-B.; Luo, Y.-F.; Wang, P.; Wang, W.-J.; Wu, J. Xanthone-derived polyketides from the Thai mangrove endophytic fungus Phomopsis sp. xy21. Fitoterapia 2018, 131, 265–271. [Google Scholar] [CrossRef]
- Tawfike, A.F.; Romli, M.; Clements, C.; Abbott, G.; Young, L.; Schumacher, M.; Diederich, M.; Farag, M.; Edrada-Ebel, R. Isolation of anticancer and anti-trypanosome secondary metabolites from the endophytic fungus Aspergillus flocculus via bioactivity guided isolation and MS based metabolomics. J. Chromatogr. B 2019, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-P.; Zheng, C.-J.; Chen, G.-Y.; Song, X.-P.; Wang, Z. Three new polyketides from a mangrove-derived fungus Colletotrichum gloeosporioides. J. Antibiot. 2019, 72, 513–517. [Google Scholar] [CrossRef]
- Suzuki, T.; Ariefta, N.R.; Koseki, T.; Furuno, H.; Kwon, E.; Momma, H.; Harneti, D.; Maharani, R.; Supratman, U.; Kimura, K.-I.; et al. New polyketides, paralactonic acids A–E produced by Paraconiothyrium sp. SW-B-1, an endophytic fungus associated with a seaweed, Chondrus ocellatus Holmes. Fitoterapia 2019, 132, 75–81. [Google Scholar] [CrossRef]
- Yang, H.; Qi, B.; Ding, N.; Jiang, F.; Jia, F.; Luo, Y.; Xu, X.; Wang, L.; Zhu, Z.; Liu, X.; et al. Polyketides from Alternaria alternata MT-47, an endophytic fungus isolated from Huperzia serrata. Fitoterapia 2019, 137, 104282. [Google Scholar] [CrossRef]
- Narmani, A.; Teponno, R.B.; Helaly, S.E.; Arzanlou, M.; Stadler, M. Cytotoxic, anti-biofilm and antimicrobial polyketides from the plant associated fungus Chaetosphaeronema achilleae. Fitoterapia 2019, 139, 104390. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, G.F.; Adpressa, D.A.; Bruhn, T.; Loesgen, S. Polyketides from Marine-Derived Aspergillus porosus: Challenges and Opportunities for Determining Absolute Configuration. J. Nat. Prod. 2019, 82, 2780–2789. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, W.; Zou, G.; Chen, S.; Pang, J.; She, Z. Bioactive polyketides from the mangrove endophytic fungi Phoma sp. SYSU-SK-7. Fitoterapia 2019, 139, 104369. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Höfert, S.-P.; Moussa, M.; Janiak, C.; Müller, W.E.; Umeokoli, B.O.; Dai, H.; Liu, Z.; Proksch, P. Polyketides and nitrogenous metabolites from the endophytic fungus Phomopsis sp. D15a2a. Tetrahedron Lett. 2019, 60, 151325. [Google Scholar] [CrossRef]
- Xia, G.Y.; Wang, L.Y.; Xia, H.; Wu, Y.Z.; Wang, Y.N.; Lin, P.C.; Lin, S. Three new polyketides from the endophytic fungus Penicillium purpurogenum. J. Asian Nat. Prod. Res. 2019, 22, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sarotti, A.M.; Yang, B.; Turkson, J.; Cao, S. A New N-methoxypyridone from the Co-Cultivation of Hawaiian Endophytic Fungi Camporesia sambuci FT1061 and Epicoccum sorghinum FT1062. Molecules 2017, 22, 1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMullin, D.R.; Green, B.D.; Prince, N.C.; Tanney, J.B.; Miller, J.D. Natural Products ofPiceaEndophytes from the Acadian Forest. J. Nat. Prod. 2017, 80, 1475–1483. [Google Scholar] [CrossRef]
- Song, H.-C.; Qin, D.; Han, M.-J.; Wang, L.; Zhang, K.; Dong, J.-Y. Bioactive 2-pyrone metabolites from an endophytic Phomopsis asparagi SWUKJ5.2020 of Kadsura angustifolia. Phytochem. Lett. 2017, 22, 235–240. [Google Scholar] [CrossRef]
- Teponno, R.B.; Noumeur, S.R.; Helaly, S.E.; Hüttel, S.; Harzallah, D.; Stadler, M. Furanones and Anthranilic Acid Derivatives from the Endophytic Fungus Dendrothyrium variisporum. Molecules 2017, 22, 1674. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, H.; Chen, Y.; Sun, Z.-H.; Li, H.-H.; Li, S.; Yan, M.; Zhang, W. Two New Metabolites from the Endophytic Fungus Alternaria sp. A744 Derived from Morinda officinalis. Molecules 2017, 22, 765. [Google Scholar] [CrossRef] [Green Version]
- Zin, W.W.M.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inácio, Â.S.; Da Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.S.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Saetang, P.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J.; Borwornpinyo, S.; Seemakhan, S.; Muanprasat, C. Depsidones and an α-pyrone derivative from Simpilcillium sp. PSU-H41, an endophytic fungus from Hevea brasiliensis leaf. Phytochemistry 2017, 143, 115–123. [Google Scholar] [CrossRef]
- Liu, H.; Tan, H.; Liu, Y.; Chen, Y.; Li, S.; Sun, Z.-H.; Li, H.; Qiu, S.-X.; Zhang, W.-M. Three new highly-oxygenated metabolites from the endophytic fungus Cytospora rhizophorae A761. Fitoterapia 2017, 117, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.-J.; Chen, H.-Q.; Wang, H.; Cai, C.; Mei, W.-L.; Dai, H.-F. New secondary metabolites from the endophytic fungus Fusarium sp. HP-2 isolated from “Qi-Nan” agarwood. Fitoterapia 2018, 130, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.-J.; Liao, H.-X.; Mei, R.-Q.; Huang, G.-L.; Yang, L.-J.; Zhou, X.-M.; Shao, T.-M.; Chen, G.-Y.; Wang, C.-Y. Two new benzophenones and one new natural amide alkaloid isolated from a mangrove-derived Fungus Penicillium citrinum. Nat. Prod. Res. 2018, 33, 1127–1134. [Google Scholar] [CrossRef]
- Kim, J.W.; Choi, H.G.; Song, J.H.; Kang, K.S.; Shim, S.H. Bioactive secondary metabolites from an endophytic fungus Phoma sp. PF2 derived from Artemisia princeps Pamp. J. Antibiot. 2019, 72, 174–177. [Google Scholar] [CrossRef]
- Kamel, R.A.; Abdel-Razek, A.S.; Hamed, A.; Ibrahim, R.R.; Stammler, H.-G.; Frese, M.; Sewald, N.; Shaaban, M. Isoshamixanthone: A new pyrano xanthone from endophytic Aspergillus sp. ASCLA and absolute configuration of epiisoshamixanthone. Nat. Prod. Res. 2019, 34, 1080–1090. [Google Scholar] [CrossRef]
- Yan, W.; Cao, L.-L.; Zhang, Y.-Y.; Zhao, R.; Zhao, S.-S.; Khan, B.; Ye, Y. New Metabolites from Endophytic Fungus Chaetomium globosum CDW7. Molecules 2018, 23, 2873. [Google Scholar] [CrossRef] [Green Version]
- Kamdem, R.S.; Wang, H.; Wafo, P.; Ebrahim, W.; Özkaya, F.C.; Makhloufi, G.; Janiak, C.; Sureechatchaiyan, P.; Kassack, M.U.; Lin, W.; et al. Induction of new metabolites from the endophytic fungus Bionectria sp. through bacterial co-culture. Fitoterapia 2018, 124, 132–136. [Google Scholar] [CrossRef]
- Ebada, S.S.; El-Neketi, M.; Ebrahim, W.; Mándi, A.; Kurtán, T.; Kalscheuer, R.; Müller, W.E.G.; Proksch, P. Cytotoxic secondary metabolites from the endophytic fungus Aspergillus versicolor KU258497. Phytochem. Lett. 2018, 24, 88–93. [Google Scholar] [CrossRef]
- Rao, L.; You, Y.-X.; Su, Y.; Liu, Y.; He, Q.; Fan, Y.; Hu, F.; Xu, Y.-K.; Zhang, C.-R. Two spiroketal derivatives with an unprecedented amino group and their cytotoxicity evaluation from the endophytic fungus Pestalotiopsis flavidula. Fitoterapia 2019, 135, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.-M.; Xu, Y.-L.; Yu, H.-F.; Zhang, C.-X.; Mao, W.; Tong, C.-P.; Zhang, Z.-D.; Tang, Q.-Y.; Zhang, Y.; Shan, W.-G.; et al. Biotransformation of Huperzine A by Irpex lacteus-A fungal endophyte of Huperzia serrata. Fitoterapia 2019, 138, 104341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yuan, X.-L.; Du, Y.-M.; Zhang, H.-B.; Shen, G.-M.; Zhang, Z.-F.; Liang, Y.-J.; Zhao, D.-L.; Xu, K. Angularly Prenylated Indole Alkaloids with Antimicrobial and Insecticidal Activities from an Endophytic Fungus Fusarium sambucinum TE-6L. J. Agric. Food Chem. 2019, 67, 11994–12001. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, X.; Li, G.; Feng, Z.; Xu, J. Pestalotiopisorin B, a new isocoumarin derivative from the mangrove endophytic fungus Pestalotiopsis sp. HHL101. Nat. Prod. Res. 2019, 34, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Tao, T.; Wang, L.; Zhao, Y.; Huang, H.; Zhang, D.; Liu, M.; Wang, Z.; Han, J. Bioprospecting of Novel and Bioactive Metabolites from Endophytic Fungi Isolated from Rubber Tree Ficus elastica Leaves. J. Microbiol. Biotechnol. 2019, 29, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Li, X.-M.; Li, X.; Li, H.-L.; Meng, L.-H.; Wang, B.-G. New lactone and isocoumarin derivatives from the marine mangrove-derived endophytic fungus Penicillium coffeae MA-314. Phytochem. Lett. 2019, 32, 1–5. [Google Scholar] [CrossRef]
- Guo, L.; Niu, S.; Chen, S.; Liu, L. Diaporone A, a new antibacterial secondary metabolite from the plant endophytic fungus Diaporthe sp. J. Antibiot. 2019, 73, 116–119. [Google Scholar] [CrossRef]
- Orfali, R.S.; Ebrahim, W.; El-Shafae, A.M. Secondary Metabolites from Alternaria sp., a Fungal Endophyte Isolated from the Seeds of Ziziphus jujuba. Chem. Nat. Compd. 2017, 53, 1031–1034. [Google Scholar] [CrossRef]
- Silva, G.H.; Zeraik, M.L.; De Oliveira, C.M.; Teles, H.L.; Trevisan, H.C.; Pfenning, L.H.; Nicolli, C.P.; Young, M.C.M.; Mascarenhas, Y.P.; Abreu, L.M.; et al. Lactone Derivatives Produced by a Phaeoacremonium sp., an Endophytic Fungus from Senna spectabilis. J. Nat. Prod. 2017, 80, 1674–1678. [Google Scholar] [CrossRef] [Green Version]
- Tchoukoua, A.; Ota, T.; Akanuma, R.; Ju, Y.-M.; Supratman, U.; Murayama, T.; Koseki, T.; Shiono, Y. A phytotoxic bicyclic lactone and other compounds from endophyte Xylaria curta. Nat. Prod. Res. 2017, 31, 2113–2118. [Google Scholar] [CrossRef]
- Yuan, W.-H.; Teng, M.-T.; Sun, S.-S.; Ma, L.; Yuan, B.; Ren, Q.; Zhang, P. Active Metabolites from Endolichenic FungusTalaromycessp. Chem. Biodivers. 2018, 15, e1800371. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-H.; Yang, D.-S.; Li, G.-H.; Liu, R.; Huang, X.; Zhang, K.-Q.; Zhao, P.-J. New secondary metabolites from an engineering mutant of endophytic Streptomyces sp. CS. Fitoterapia 2018, 130, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Wu, P.; Xue, J.; Liu, L.; Li, H.; Wei, X. Asperimides A–D, anti-inflammatory aromatic butenolides from a tropical endophytic fungus Aspergillus terreus. Fitoterapia 2018, 131, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Gao, W.; Wang, J.; Liu, M.; Zhang, Y.; Chen, C.; Hu, Z.; Xue, Y.; Li, D.; Zhang, Q.; et al. Terrusnolides A-D, new butenolides with anti-inflammatory activities from an endophytic Aspergillus from Tripterygium wilfordii. Fitoterapia 2018, 130, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Basnet, B.B.; Chen, B.; Suleimen, Y.M.; Ma, K.; Guo, S.; Bao, L.; Huang, Y.; Liu, H. Cytotoxic Secondary Metabolites from the Endolichenic Fungus Hypoxylon fuscum. Planta Medica 2019, 85, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Y.; Wang, X.-J.; Liu, Z.; Meng, F.-X.; Sun, S.-F.; Ye, F.; Liu, Y.-B. Nonadride and Spirocyclic Anhydride Derivatives from the Plant Endophytic Fungus Talaromyces purpurogenus. J. Nat. Prod. 2019, 82, 2953–2962. [Google Scholar] [CrossRef]
- Yang, H.-G.; Li, J.-J.; Chen, S.-M.; Mou, L.-M.; Zou, J.; Wang, C.-X.; Chen, G.; Qin, S.-Y.; Yao, X.-S.; Gao, H. Phenylisotertronic acids from the TCM endophytic fungus Phyllosticta sp. Fitoterapia 2018, 124, 86–91. [Google Scholar] [CrossRef]
- Rathnayake, G.R.N.; Kumar, N.S.; Jayasinghe, L.; Araya, H.; Fujimoto, Y. Secondary Metabolites Produced by an Endophytic Fungus Pestalotiopsis microspora. Nat. Prod. Bioprospecting 2019, 9, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, J.; Liu, D.; Guo, J.; Liu, T.; Xin, Z. Pencitrin and pencitrinol, two new citrinin derivatives from an endophytic fungus Penicillium citrinum salicorn 46. Phytochem. Lett. 2017, 22, 229–234. [Google Scholar] [CrossRef]
- Tan, X.-M.; Li, L.-Y.; Sun, L.-Y.; Sun, B.-D.; Niu, S.-B.; Wang, M.-H.; Zhang, X.-Y.; Sun, W.-S.; Zhang, G.-S.; Deng, H.; et al. Spiciferone analogs from an endophytic fungus Phoma betae collected from desert plants in West China. J. Antibiot. 2018, 71, 613–617. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, H.; Liu, Y.; Gu, Q.; Xu, J.; Huang, X.; She, Z. Ethylnaphthoquinone derivatives as inhibitors of indoleamine-2, 3-dioxygenase from the mangrove endophytic fungus Neofusicoccum austral SYSU-SKS024. Fitoterapia 2018, 125, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Liao, Z.; Zhou, M.; Wang, G.; Wu, Y.; Gao, S.; Qiu, D.; Liu, X.; Lin, T.-; Chen, H. Cytoskyrin C, an unusual asymmetric bisanthraquinone with cage-like skeleton from the endophytic fungus Diaporthe sp. Fitoterapia 2018, 128, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, J.-H.; Zhao, J.-C.; Wang, Y.-L.; Dong, P.-P.; Liu, X.-G.; Zhang, T.-Y.; Wu, Y.-Y.; Shang, D.; Zhang, Y.-X.; et al. Xylarianins A-D from the endophytic fungus Xylaria sp. SYPF 8246 as natural inhibitors of human carboxylesterase 2. Bioorganic Chem. 2018, 81, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.; Masi, M.; Cimmino, A.; Tuzi, A.; Jena, S.; Tayung, K.; Evidente, A. Funiculosone, a substituted dihydroxanthene-1,9-dione with two of its analogues produced by an endolichenic fungus Talaromyces funiculosus and their antimicrobial activity. Phytochemistry 2019, 157, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Chinpha, S.; Saetang, P.; Phongpaichit, S.; Jungsuttiwong, S.; Hadsadee, S.; Sakayaroj, J.; Preedanon, S.; Temkitthawon, P.; Ingkaninan, K. Depsidones and a dihydroxanthenone from the endophytic fungi Simplicillium lanosoniveum (J.F.H. Beyma) Zare & W. Gams PSU-H168 and PSU-H261. Fitoterapia 2019, 138, 104286. [Google Scholar] [CrossRef]
- Supratman, U.; Hirai, N.; Sato, S.; Watanabe, K.; Malik, A.; Annas, S.; Harneti, D.; Maharani, R.; Koseki, T.; Shiono, Y. New naphthoquinone derivatives from Fusarium napiforme of a mangrove plant. Nat. Prod. Res. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, X.-Q.; Yang, Y.-B.; Zhao, L.-X.; Zhou, Q.-Y.; Zhang, Z.-X.; Zhou, H.; Hu, M.; Ruan, B.-H.; Ding, Z.-T. A Novel Steroid Derivative and a New Steroidal Saponin from Endophytic Fungus Xylaria sp. Nat. Prod. Commun. 2017, 12, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.-X.; Li, Z.; Chen, Y.; Yang, Y.-H.; Li, G.-H.; Zhao, P.-J. Four new steroids from the endophytic fungus Chaetomium sp. M453 derived of Chinese herbal medicine Huperzia serrata. Fitoterapia 2017, 117, 41–46. [Google Scholar] [CrossRef]
- Huang, R.-L.; Zheng, C.-J.; Zhou, X.-M.; Song, X.-M.; Wu, P.-P.; Zhao, Y.-F.; Chen, G.-Y.; Song, X.-P.; Han, C.-R. Three new methylated Δ8-pregnene steroids from the Polyalthia laui-derived fungus Stemphylium sp. AZGP4-2. Bioorganic Chem. 2020, 95, 102927. [Google Scholar] [CrossRef]
- Khayat, M.T.; Ibrahim, S.R.M.; Mohamed, G.A.A.; Abdallah, H.M. Anti-inflammatory metabolites from endophytic fungus Fusarium sp. Phytochem. Lett. 2019, 29, 104–109. [Google Scholar] [CrossRef]
- Wang, Z.-R.; Li, G.; Ji, L.-X.; Wang, H.-H.; Gao, H.; Peng, X.-P.; Lou, H.-X. Induced production of steroids by co-cultivation of two endophytes from Mahonia fortunei. Steroids 2019, 145, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhu, Y.-X.; Peng, C.; Li, J. Two new sterol derivatives isolated from the endophytic fungus Aspergillus tubingensis YP-2. Nat. Prod. Res. 2019, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Chen, S.; Long, Y.; Li, C.; Huang, X.; She, Z. Depsidones from Talaromyces stipitatus SK-4, an endophytic fungus of the mangrove plant Acanthus ilicifolius. Phytochem. Lett. 2017, 20, 196–199. [Google Scholar] [CrossRef]
- Chang, H.-S.; Cheng, M.-J.; Cheng, M.-J.; Chan, H.-Y.; Hsieh, S.-Y.; Lin, C.-H.; Yech, Y.-J.; Chen, I.-S. Secondary metabolites produced by an endophytic fungus Cordyceps ninchukispora from the seeds of Beilschmiedia erythrophloia Hayata. Phytochem. Lett. 2017, 22, 179–184. [Google Scholar] [CrossRef]
- Deng, Z.; Li, C.; Luo, D.; Teng, P.; Guo, Z.; Tu, X.; Zou, K.; Gong, D. A new cinnamic acid derivative from plant-derived endophytic fungus Pyronema sp. Nat. Prod. Res. 2017, 31, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Wijeratne, E.M.K.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A.L. An epigenetic modifier induces production of (10′S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorganic Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [Green Version]
- Kongprapan, T.; Xu, X.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J.; Chen, J.; Shen, X. Cytosporone derivatives from the endophytic fungus Phomopsis sp. PSU-H188. Phytochem. Lett. 2017, 22, 219–223. [Google Scholar] [CrossRef]
- Kyekyeku, J.O.; Kusari, S.; Adosraku, R.K.; Bullach, A.; Golz, C.; Strohmann, C.; Spiteller, M. Antibacterial secondary metabolites from an endophytic fungus, Fusarium solani JK10. Fitoterapia 2017, 119, 108–114. [Google Scholar] [CrossRef]
- Li, X.-B.; Chen, G.-Y.; Liu, R.-J.; Zheng, C.; Song, X.-M.; Han, C.-R. A new biphenyl derivative from the mangrove endophytic fungus Phomopsis longicolla HL-2232. Nat. Prod. Res. 2017, 31, 2264–2267. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, W.; Wen, L.; Zhou, L.; Kang, X.; Chen, G. A New Hexanedioic Acid Analogue from the Endophytic Fungus Penicillium sp. OC-4 of Orchidantha chinensis. Chem. Nat. Compd. 2017, 53, 834–838. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Farthouse, J.; Islam, M.T.; Schüffler, A.; Laatsch, H. Metabolites from the Endophytic FungusCurvulariasp. M12 Act as Motility Inhibitors againstPhytophthora capsiciZoospores. J. Nat. Prod. 2017, 80, 347–355. [Google Scholar] [CrossRef]
- Siridechakorn, I.; Yue, Z.; Mittraphab, Y.; Lei, X.; Pudhom, K. Identification of spirobisnaphthalene derivatives with anti-tumor activities from the endophytic fungus Rhytidhysteron rufulum AS21B. Bioorg. Med. Chem. 2017, 25, 2878–2882. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; She, Z.; Li, C. Penochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semi-synthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Maha, A.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Tyrosine and hydantoin derivatives from the fungus Phoma herbarum PSU-H256 isolated from Hevea brasiliensis. Tetrahedron 2017, 73, 4597–4601. [Google Scholar] [CrossRef]
- Elkhayat, E.S.; Goda, A.M. Antifungal and cytotoxic constituents from the endophytic fungus Penicillium sp. Bull. Fac. Pharmacy Cairo Univ. 2017, 55, 85–89. [Google Scholar] [CrossRef]
- Sharma, N.; Kushwaha, M.; Arora, D.; Jain, S.; Singamaneni, V.; Sharma, S.; Shankar, R.; Bhushan, S.; Gupta, P.; Jaglan, S. New cytochalasin from Rosellinia sanctae-cruciana, an endophytic fungus of Albizia lebbeck. J. Appl. Microbiol. 2018, 125, 111–120. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.; Yuan, X.-L.; Du, Y.; Wang, B.-G.; Zhang, Y.-M.D.A.Z.-F. Antifungal Prenylated Diphenyl Ethers from Arthrinium arundinis, an Endophytic Fungus Isolated from the Leaves of Tobacco (Nicotiana tabacum L.). Molecules 2018, 23, 3179. [Google Scholar] [CrossRef] [Green Version]
- Akhter, N.; Pan, C.; Liu, Y.; Shi, Y.; Wu, B. Isolation and structure determination of a new indene derivative from endophytic fungus Aspergillus flavipes Y-62. Nat. Prod. Res. 2018, 33, 2939–2944. [Google Scholar] [CrossRef]
- De Oliveira, D.M.; Pereira, C.B.; Mendes, G.; Junker, J.; Kolloff, M.; Rosa, L.H.; Rosa, C.A.; Alves, T.M.; Zani, C.L.; Johann, S.; et al. Two new usnic acid derivatives from the endophytic fungus Mycosphaerella sp. Zeitschrift für Naturforschung C 2018, 73, 449–455. [Google Scholar] [CrossRef]
- Mafezoli, J.; Xu, Y.-M.; Hilário, F.; Freidhof, B.; Espinosa-Artiles, P.; Dos Santos, L.C.; De Oliveira, M.C.; Gunatilaka, A.A.L. Modulation of polyketide biosynthetic pathway of the endophytic fungus, Anteaglonium sp. FL0768, by copper (II) and anacardic acid. Phytochem. Lett. 2018, 28, 157–163. [Google Scholar] [CrossRef]
- Xie, J.; Wu, Y.-Y.; Zhang, T.-Y.; Zhang, M.-Y.; Peng, F.-; Lin, B.; Zhang, Y.-X. New antimicrobial compounds produced by endophytic Penicillium janthinellum isolated from Panax notoginseng as potential inhibitors of FtsZ. Fitoterapia 2018, 131, 35–43. [Google Scholar] [CrossRef]
- Savi, D.C.; Shaaban, K.A.; Gos, F.M.W.R.; Ponomareva, L.V.; Thorson, J.S.; Glienke, C.; Rohr, J. Phaeophleospora vochysiae Savi & Glienke sp. nov. Isolated from Vochysia divergens Found in the Pantanal, Brazil, Produces Bioactive Secondary Metabolites. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Wu, Y.-Z.; Zhang, H.-W.; Sun, Z.-H.; Dai, J.-G.; Hu, Y.-C.; Li, R.; Lin, P.-C.; Xia, G.; Wang, L.; Qiu, B.-L.; et al. Bysspectin A, an unusual octaketide dimer and the precursor derivatives from the endophytic fungus Byssochlamys spectabilis IMM0002 and their biological activities. Eur. J. Med. Chem. 2018, 145, 717–725. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Elkhayat, E.S.; Ross, S.A. Fusarithioamide B, a new benzamide derivative from the endophytic fungus Fusarium chlamydosporium with potent cytotoxic and antimicrobial activities. Bioorganic Med. Chem. 2018, 26, 786–790. [Google Scholar] [CrossRef]
- Maciel, O.M.C.; Tavares, R.S.N.; Caluz, D.R.E.; Gaspar, L.R.; Debonsi, H.M. Photoprotective potential of metabolites isolated from algae-associated fungi Annulohypoxylon stygium. J. Photochem. Photobiol. B Biol. 2018, 178, 316–322. [Google Scholar] [CrossRef]
- Wu, X.; Wang, S.; Liu, C.; Zhang, C.; Guo, J.; Shang, X. A new 2H-benzindazole compound from Alternaria alternata Shm-1, an endophytic fungus isolated from the fresh wild fruit of Phellinus igniarius. J. Nat. Med. 2019, 73, 620–626. [Google Scholar] [CrossRef]
- Bai, M.; Zheng, C.-J.; Tang, D.-Q.; Zhang, F.; Wang, H.-Y.; Chen, G.-Y. Two new secondary metabolites from a mangrove-derived fungus Cladosporium sp. JS1-2. J. Antibiot. 2019, 72, 779–782. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Ponomareva, L.V.; Rodrigues, R.; Rohr, J.; Thorson, J.S.; Glienke, C.; Shaaban, K.A. Vochysiamides A and B: Two new bioactive carboxamides produced by the new species Diaporthe vochysiae. Fitoterapia 2019, 138, 104273. [Google Scholar] [CrossRef]
- Wang, W.-X.; Li, Z.-H.; He, J.; Feng, T.; Li, J.; Liu, J.-K. Cytotoxic cytochalasans from fungus Xylaria longipes. Fitoterapia 2019, 137, 104278. [Google Scholar] [CrossRef]
- Wickramarachchi, S.R.; Samanthi, U.; Wijeratne, K.; Paranagama, P.A. A new antioxidant active compound from the endolichenic fungus, Penicillium citrinum inhabiting the lichen, Parmotrema sp. Int. J. Pharm. Sci. Res. 2019, 10, 3412–3420. [Google Scholar] [CrossRef]
- Inose, K.; Tanaka, K.; Koshino, H.; Hashimoto, M. Cyclopericodiol and new chlorinated melleins isolated from Periconia macrospinosa KT3863. Tetrahedron 2019, 75, 130470. [Google Scholar] [CrossRef]
- Kumar, S.; Pagar, A.D.; Ahmad, F.; Dwibedi, V.; Wani, A.; Bharatam, P.V.; Chhibber, M.; Saxena, S.; Singh, I.P. Xanthine oxidase inhibitors from an endophytic fungus Lasiodiplodia pseudotheobromae. Bioorganic Chem. 2019, 87, 851–856. [Google Scholar] [CrossRef]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; El-Neketi, M.; Ebrahim, W.; Singab, A.B. New secondary metabolites from the mangrove-derived fungus Aspergillus sp. AV-2. Phytochem. Lett. 2019, 29, 1–5. [Google Scholar] [CrossRef]
- Yu, G.; Sun, Z.; Peng, J.; Zhu, M.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Secondary Metabolites Produced by Combined Culture of Penicillium crustosum and a Xylaria sp. J. Nat. Prod. 2019, 82, 2013–2017. [Google Scholar] [CrossRef]
- Li, H.-L.; Li, X.-M.; Yang, S.-Q.; Cao, J.; Li, Y.-H.; Wang, B.-G. Induced terreins production from marine red algal-derived endophytic fungus Aspergillus terreus EN-539 co-cultured with symbiotic fungus Paecilomyces lilacinus EN-531. J. Antibiot. 2020, 73, 108–111. [Google Scholar] [CrossRef]
- Riga, R.; Happyana, N.; Quentmeier, A.; Zammarelli, C.; Kayser, O.; Hakim, E.H. Secondary metabolites from Diaporthe lithocarpus isolated from Artocarpus heterophyllus. Nat. Prod. Res. 2019, 1–5. [Google Scholar] [CrossRef]
- Ma, H.; Wang, F.; Jin, X.; Jiang, J.; Hu, S.; Cheng, L.; Zhang, G. A new diketopiperazine from an endophytic fungus Aspergillus aculeatus F027. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Nikmawahda, H.T.; Aboshi, T.; Murayama, T.; Tawaraya, K.; Koseki, T.; Katagi, G.; Kakihara, Y.; Shiono, Y. Fusaspirols A-D, novel oxaspirol derivatives isolated from Fusarium solani B-18. Tetrahedron 2019, 75, 1371–1377. [Google Scholar] [CrossRef]
- Supratman, U.; Suzuki, T.; Nakamura, T.; Yokoyama, Y.; Harneti, D.; Maharani, R.; Salam, S.; Abdullah, F.F.; Koseki, T.; Shiono, Y. New metabolites produced by endophyte Clonostachys rosea B5-2. Nat. Prod. Res. 2019, 1–7. [Google Scholar] [CrossRef]
- Choi, H.G.; Kim, J.W.; Choi, H.; Kang, K.S.; Shim, S.H. New hydroxydecanoic acid derivatives produced by an endophytic yeast Aureobasidium pullulans AJF1 from flowers of Aconitum carmichaeli. Molecules 2019, 24, 4051. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Li, W.; Bang, S.; Lee, S.J.; Kang, N.-Y.; Kim, S.; Kim, T.I.; Go, Y.; Shim, S.H. Secondary Metabolites of The Endophytic Fungus Alternaria alternata JS0515 Isolated from Vitex rotundifolia and Their Effects on Pyruvate Dehydrogenase activity. Molecules 2019, 24, 4450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
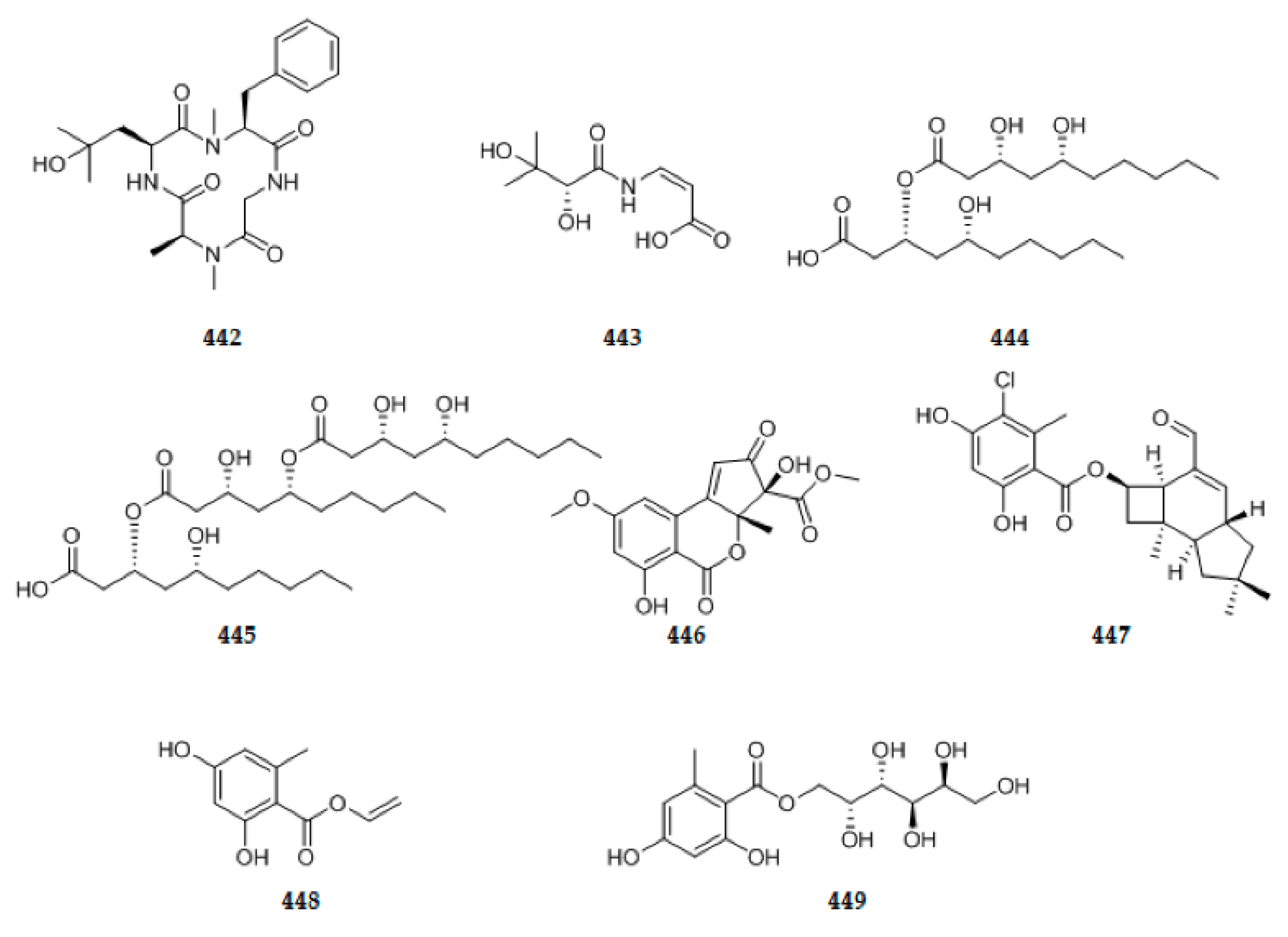{kind=link}
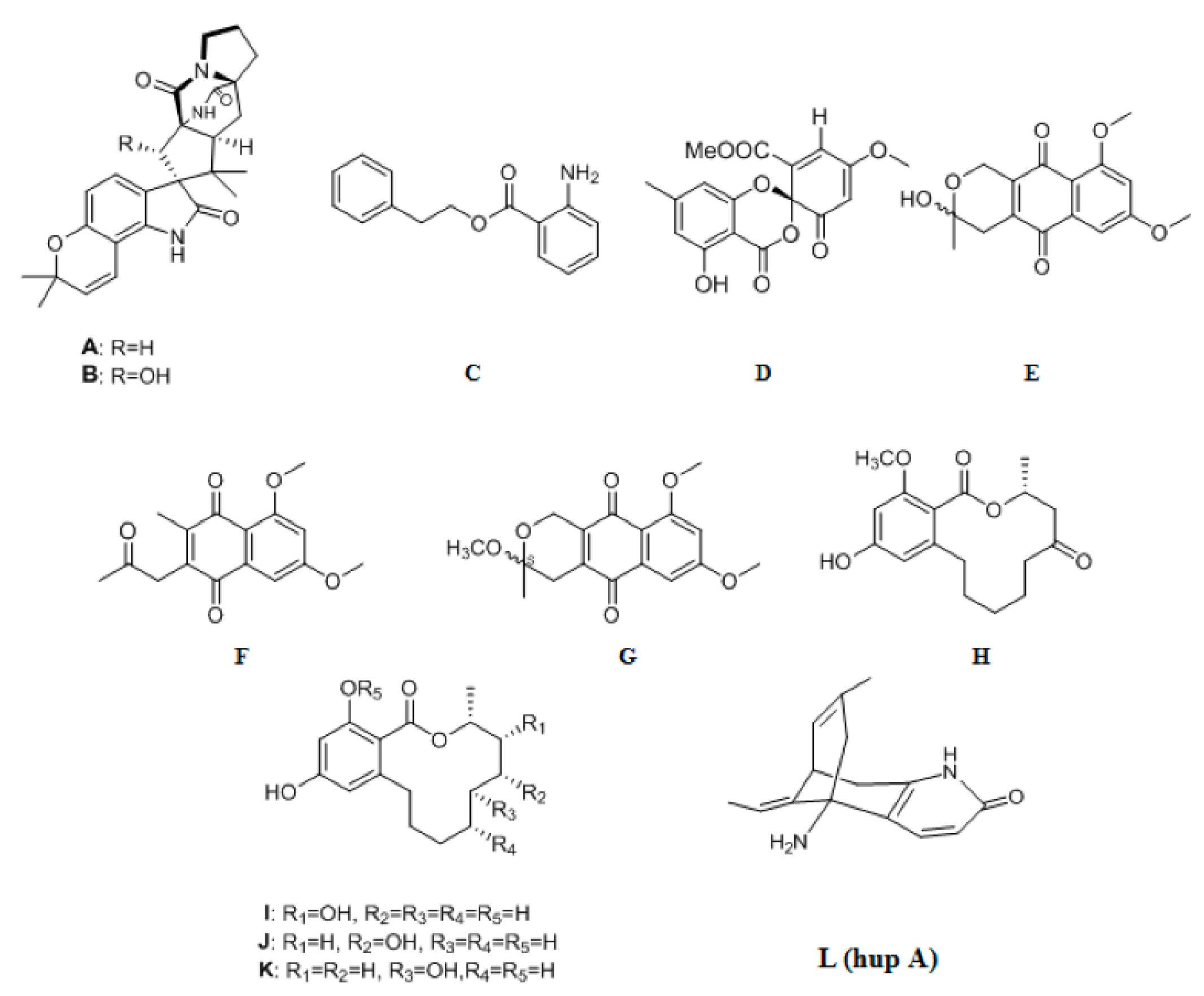{kind=link}
| Compound | Molecular Formula | Degree of Unsaturation | Color and Morphology | Endophytic Fungus | Host Plant | Site and Nation | Biological Activity | Ref. |
|---|---|---|---|---|---|---|---|---|
| Terpenoids Sesquiterpenoids and derivatives | ||||||||
| 1 | C19H26O7 | 7 | brown oil | Pestalotiopsissp.T | lichen Cetraria islandica (L.) Ach. | Yunnan Province, China | Inhibit the growth of plant pathogenic fungus (1,5) | [9] |
| 2 | C21H28O8 | 8 | ||||||
| 3 | C21H30O8 | 7 | ||||||
| 4 | C21H28O8 | 8 | ||||||
| 5 | C19H26O7 | 7 | ||||||
| 6 | C15H20O4 | 6 | white powder | Co-culture Strain 307: Trichoderma sp. the stem bark of Clerodendrum inerme Bacterium B2: Acinetobacter johnsonii From an aquaculture pond | Guangdong Province, China | Show moderate inhibitory activity against α-glucosidase (7) | [10] | |
| 7 | C15H20O4 | 6 | ||||||
| 8 | C15H24O2 | 4 | colorless gum | Trichoderma atroviride | bulb of Lycoris radiata. | Hubei Province China | Inactive | [11] |
| 9 | C15H26O3 | 3 | white amorphous powder | Co-culture Pestalotiopsis sp. fruits of Drepanocarpus lunatus (Fabaceae) Bacillus subtilis | Weak antibacterial activities (9) | [12] | ||
| 10 | C15H24O3 | 4 | colorless oi | |||||
| 11 | C22H32O5 | 7 | colorless crystal | Nectria pseudotrichia 120-1NP | Inner tissue of Gliricidia sepium healthy stem | Cytotoxicity (11–13) | [13] | |
| 12 | C26H38O7 | 8 | yellow oil | |||||
| 13 | C15H26O | 3 | yellow oil | |||||
| 14 | C15H26O3 | 3 | Co-culture Nigrospora oryzae Irpex lacteus | seeds of Dendrobium officinale | Yunnan Province, China | Anti-AChE activity | [14] | |
| 15 | C15H26O2 | 3 | white powder | Emericella sp. XL 029 | leaves of Panax notoginseng | Hebei province, China | Antifungal activity Antibacterial activity (15,16) | [15] |
| 16 | C15H26O3 | 3 | colorless oil | |||||
| 17 | C19H24O4 | 8 | colorless oil | Trichothecium crotocinigenum | Antiphytopathogenic activity (17–20) | [16] | ||
| 18 | C19H25ClO5 | 7 | colorless crystals | |||||
| 19 | C22H28O5 | 9 | colorless crystals | |||||
| 20 | ||||||||
| 21 | C15H22O4 | 5 | colorless oil | Trichoderma atroviride S361 | Bark of Cephalotaxus fortunei | Zhejiang province, China | Inactive | [17] |
| 22 | C15H20O4 | 6 | white amorphous powder | Aspergillus sp. xy02 | leaves of mangrove Xylocarpus moluccensis | Trang Province, Thailand | Antibacterial activity (23–24,26,28) | [18] |
| 23 | ||||||||
| 24 | ||||||||
| 25 | ||||||||
| 26 | colorless oil | |||||||
| 27 | ||||||||
| 28 | ||||||||
| 29 | C15H24O3 | 4 | colorless oil | Pestalotiopsis adusta | stem bark of medicinal plant Sinopodophyllum hexandrum (Royle) Ying | Qinling Mountains China | Weak to moderate cytotoxic activity | [19] |
| 30 | C15H26O2 | 3 | colorless oil | F. proliferatum AF-04 | green Chinese onion | Lanzhou, China | [20] | |
| 31 | C14H24O3 | 3 | colorless crystals | Trichoderma asperellum A-YMD-9–2 | marine Red alga Gracilaria verrucosa | Yangma Island, Yantai, China | Potent inhibition of several marine phytoplankton species 31–37 | [21] |
| 32 | C14H20O2 | 5 | colorless oil | |||||
| 33 | ||||||||
| 34 | ||||||||
| 35 | C22H37NO7 | 5 | colorless oil | |||||
| 36 | ||||||||
| 37 | ||||||||
| 38 | C15H24O4 | 4 | crystal powder | Alternaria oxytropis | desert plant locoweed Oxytropis glabra | Inner Mongolia, China | Displayed an effect on the root growth in Arabidopsis thaliana (38) | [22] |
| 39 | colourless oil | |||||||
| 40 | C15H22O4 | 5 | colourless oil | |||||
| 41 | C15H24O5 | 4 | crystal powder | |||||
| 42 | C15H22O3 | 5 | colourless oil | |||||
| 43 | C15H24O5 | 4 | ||||||
| 44 | C15H26O4 | 3 | ||||||
| 45 | ||||||||
| 46 | C15H26O3 | 3 | ||||||
| 47 | ||||||||
| 48 | C15H22O3 | 5 | colorless crystal | Pleosporales sp. SK7 | mangrove plant Kandelia candel | Guangxi Province, China | [23] | |
| 49 | C15H22O4 | 5 | yellowish needle crystals | Irpex lacteus DR10-1 | waterlogging tolerant plant D. chinense | Chongqing China | Antioxidant activity Antibacterial activity | [24] |
| 50 | C15H16O3 | 8 | colorless crystals | Trichoderma virens QA-8 | fresh inner tissue of the medicinal plant Artemisia argyi | Hubei Province, China | Antibacterial (50–52,55) Antifungal activity (50–55) | [25] |
| 51 | C15H16O4 | 8 | colorless oil | |||||
| 52 | C15H22O2 | 5 | amorphous powder | |||||
| 53 | C15H22O3 | 5 | amorphous powder | |||||
| 54 | C15H24O3 | 4 | colorless oil | |||||
| 55 | C14H16O4 | 7 | amorphous powder | |||||
| 56 | C15H26O2 | 3 | colorless needle | Alternaria alternate | leaves of Psidium littorale Raddi | Fujian Province, China | [26] | |
| 57 | C15H22O3 | 5 | colorless oil | Epicoccum sp. YUD17002 & Armillaria sp. | rhizomes of the underground portion of Gastrodia elata | Yunnan Province, China | [27] | |
| 58 | C15H24O4 | 4 | ||||||
| 59 | C15H22O2 | 5 | ||||||
| 60 | C15H22O3 | 5 | ||||||
| 61 | C15H24O4 | 4 | white amorphous powder | |||||
| 62 | C29H42O9 | 9 | sticky and optically active oi | Colletotrichum gloeosporioides | Cameroonian medicinal plant Trichilia monadelpha (Meliaceae) | Yaounde, Central region, Cameroon | [28] | |
| 63 | C17H22O7 | 7 | white powder | Penicillium purpurogenum IMM003 | leaf tissue of the medicinal plant Edgeworthia chrysantha. | China | Show significant inhibitory activity against pancreatic lipase | [29] |
| 64 | C17H20O7 | 8 | colorless crystals | |||||
| 65 | C16H20O6 | 7 | ||||||
| 66 | C16H24O3 | 4 | yellow oil | Fusarium oxysporum ZZP-R1 | coastal plant Rumex madaio Makino | Putuo Island (Zhoushan, China) | Moderate antibacterial effect | [30] |
| Terpenoids Diterpenoids | ||||||||
| 67 | C20H30O6 | 6 | colorless oil | Nectria pseudotrichia 120-1NP | healthy stem of Gliricidia sepium | Yogyakarta, Indonesia | [31] | |
| 68 | C28H39NO3 | 10 | amorphous powder | Drechmeria sp. | root of Panax notoginseng | Yunnan, China | Display inhibitory effect (69) Weak antimicrobial effects. (68,70,74) | [32] |
| 69 | C28H37NO5 | 11 | ||||||
| 70 | C33H45NO5 | 12 | ||||||
| 71 | C32H43NO7 | 12 | ||||||
| 72 | C32H43NO7 | 12 | ||||||
| 73 | C33H45NO7 | 12 | ||||||
| 74 | C27H33NO5 | 12 | ||||||
| 75 | C32H33NO9 | 17 | amorphous powder | Drechmeria sp. | root of Panax notoginseng | Yunnan province, China | Display the significant agonistic effect on pregnane X receptor (PXR) (76) | [33] |
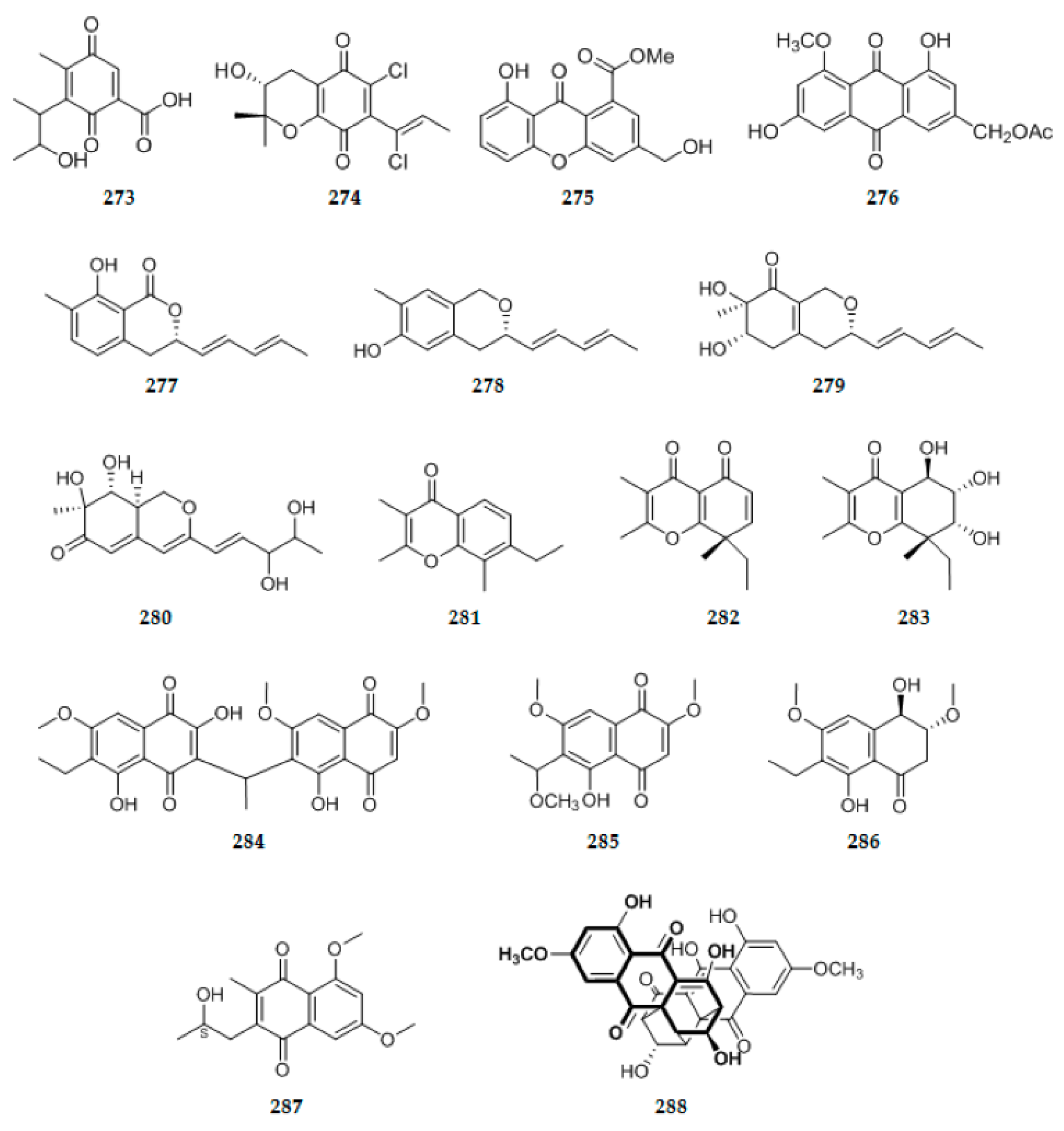
| 76 | C32H41NO6 | 13 | ||||||
| 77 | C26H40O5 | 7 | colorless oil | Neosartorya fifischeri JS0553 | Plant G. littoralis | Suncheon, Korea | [34] | |
| 78 | C28H39NO3 | 10 | Pale yellow oil | Aspergillus versicolor | fruits of the mangrove Avicennia marina | Red Sea, Egypt | Weak cytotoxic activity (79) | [35] |
| 79 | ||||||||
| 80 | C20H26O4 | 8 | colorless crystals | Xylaralyce sp. | healthy leaves of Distylium chinense | China | Display brine shrimp inhibiting activity | [36] |
| 81 | C20H26O5 | 8 | colorless crystals | Apiospora montagnei | lichen Cladonia sp. | [37] | ||
| Terpenoids Other terpenoids | ||||||||
| 82 | C26H37NO3 | 9 | colorless oil | Aspergillus sp. ZJ-68 | fresh leaves of the mangrove plant Kandelia candel | Guangdong Province, China. | Exhibit inhibitory effects on lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophage cells (89–91) Show comparable inhibition of Mycobacterium tuberculosis protein tyrosine phosphatase B (89) | [38] |
| 83 | C25H35NO3 | 9 | ||||||
| 84 | C25H35NO2 | 9 | ||||||
| 85 | C26H39NO3 | 8 | ||||||
| 86 | C25H34O3 | 9 | ||||||
| 87 | C25H36O4 | 8 | ||||||
| 88 | C25H36O4 | 8 | ||||||
| 89 | C25H36O4 | 8 | ||||||
| 90 | C25H34O3 | 9 | ||||||
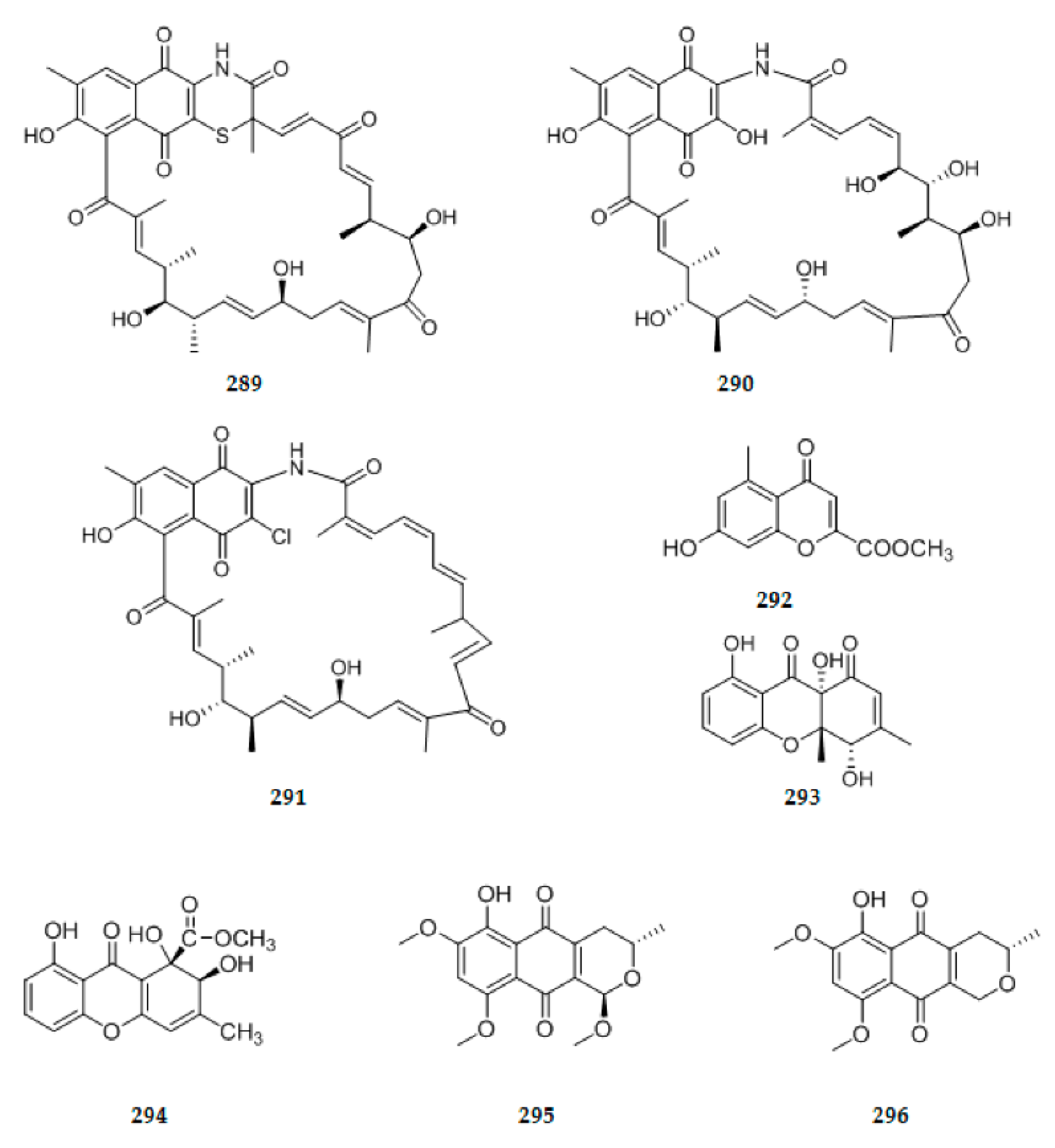
| 91 | C25H38O5 | 7 | ||||||
| 92 | C25H38O3 | 7 | ||||||
| 93 | C30H40O6 | 11 | yellowish needle crystals | Kadsura angustifolia & Penicillium sp. SWUKD4.1850 | fresh healthy branches of K. angustifolia | China | Moderate cytotoxic activity (93–100) | [39] |
| 94 | C30H40O6 | 11 | white needle crystals | |||||
| 95 | C30H40O6 | 11 | white amorphous solid | |||||
| 96 | C30H40O6 | 11 | ||||||
| 97 | C32H44O7 | 11 | white amorphous powder | |||||
| 98 | C30H42O6 | 10 | white powder | |||||
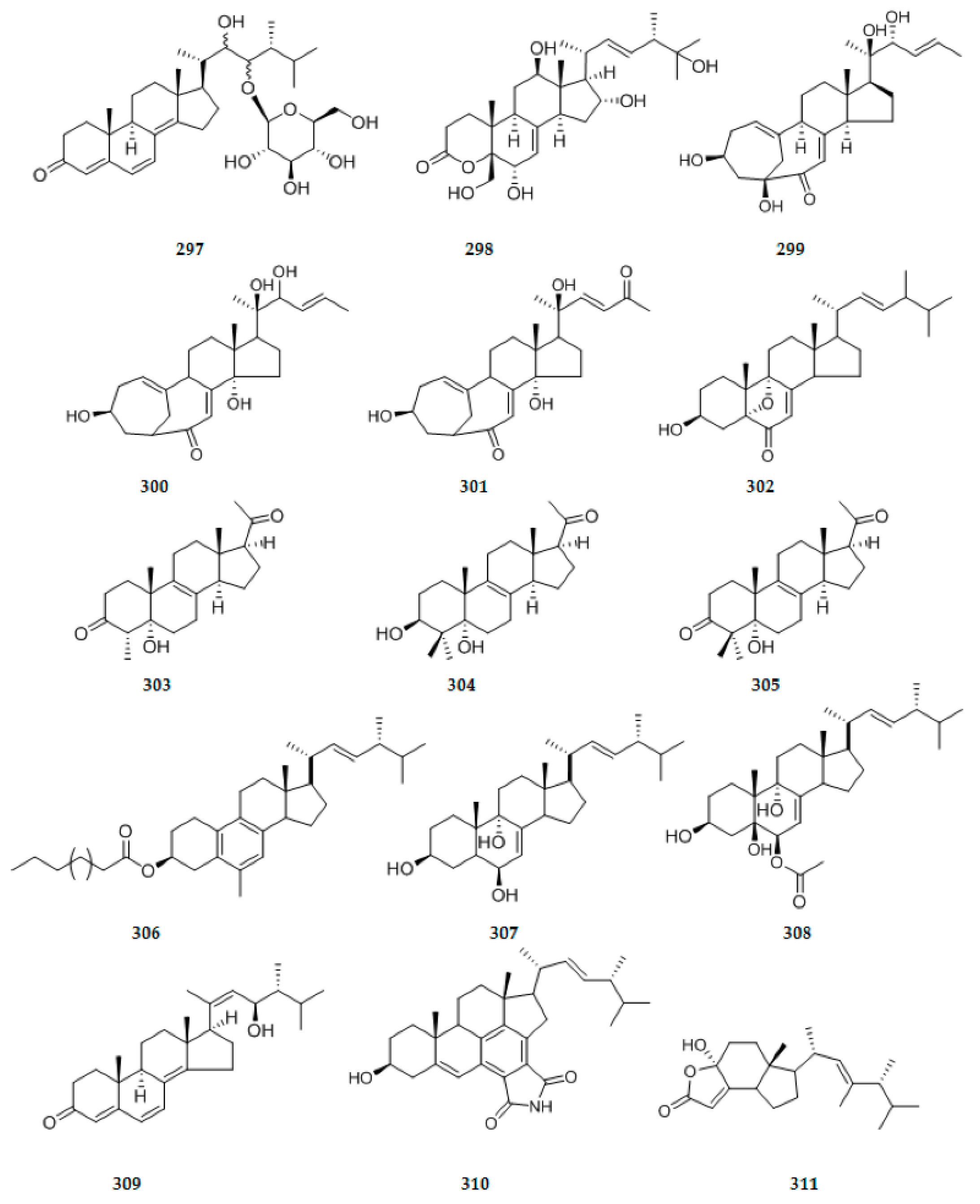
| 99 | C34H46O8 | 12 | yellow amorphous solid | |||||
| 100 | C31H44O6 | 10 | yellow amorphous solid | |||||
| 101 | C30H46O6 | 8 | white amorphous powder | |||||
| 102 | C17H26O5 | 5 | colorless oil | Phyllosticta capitalensis | leaves of Cephalotaxus fortunei Hook | Shanxi Province, China | [40] | |
| 103 | C17H24O5 | 6 | ||||||
| 104 | C17H22O5 | 7 | ||||||
| 105 | C22H32O6 | 7 | ||||||
| 106 | C17H26O5 | 5 | ||||||
| 107 | C15H20O5 | 6 | Aspergillus versicolor | Show weak cytotoxic activities against Hela cells. (113–114) | [35] | |||
| 108 | C15H20O5 | 6 | ||||||
| 109 | C15H20O5 | 6 | rose-colored oil | |||||
| 110 | C17H22O6 | 7 | ||||||
| 111 | C15H20O5 | 6 | ||||||
| 112 | C15H20O4 | 6 | colorless oil | |||||
| 113 | C17H22O5 | 7 | ||||||
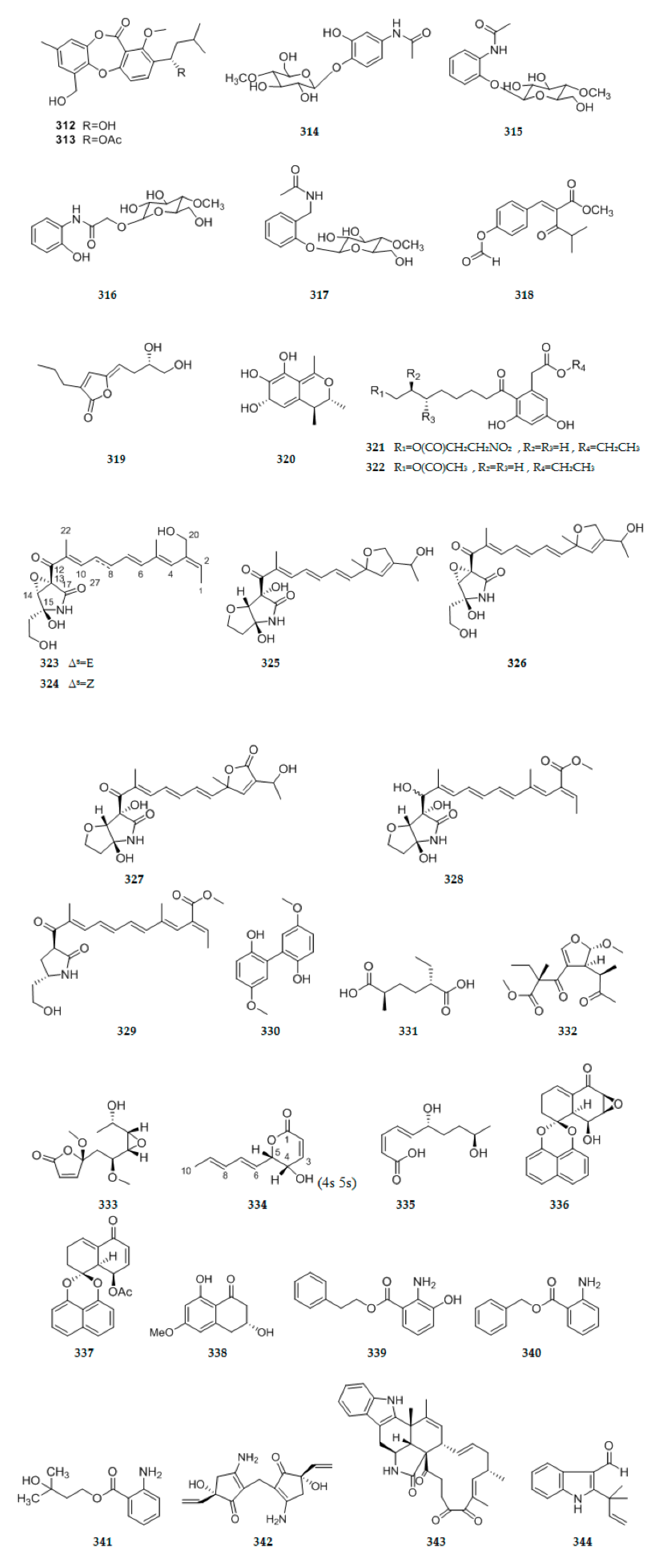
| 114 | C17H22O5 | 7 | ||||||
| 115 | C16H24O5 | 5 | ||||||
| 116 | C21H32O3 | 6 | yellow oil | Fusarium oxysporum ZZP-R1 | coastal plant Rumex madaio Makino | Putuo Island (Zhoushan, China) | Antimicrobial activity | [30] |
| 117 | C12H20O4 | 3 | yellow oil | Diaporthe lithocarpus A740 | from the twigs of medicinal plant Morinda officinalis | Guangdong province China | [41] | |
| Ketones | ||||||||
| 118 | C20H32O4N2 | 6 | white powder | Eupenicillium sp. LG41 | Chinese medicinal plant Xanthium sibiricum | China | Cytotoxic activity Antimicrobial activity (Antibacterial) | [42] |
| 119 | C38H59O6N | 10 | ||||||
| 120 | C12H18O5 | 4 | colorless crystals | Phomopsis sp. sh917 | fresh stems of I. eriocalyx var. laxiflflora | Kunming, China | [43] | |
| 121 | C12H18O5 | 4 | colorless powders | |||||
| 122 | C11H12O5 | 6 | Brown needles | |||||
| 123 | C20H26O10 | 8 | colorless needles | |||||
| 124 | C13H14O5 | 7 | brown solids | |||||
| 125 | C17H20O3 | 8 | white amorphous powder | Pestalotiopsis vaccinii (cgmcc3.9199) | branch of mangrove plant Kandelia candel (L.) Druce (Rhizophoraceae) | coastal and estuarine areas of southern China | Anti-enterovirus 7l (EV71) | [44] |
| 126 | C14H18O4 | 6 | colorless oil | |||||
| 127 | C12H16O4 | 5 | ||||||
| 128 | C12H18O4 | 4 | ||||||
| 129 | C12H14O4 | 6 | ||||||
| 130 | C17H22O4 | 7 | ||||||
| 131 | C17H24O5 | 6 | ||||||
| 132 | C18H28O6 | 5 | ||||||
| 133 | C17H22O5 | 7 | ||||||
| 134 | C17H22O5 | 7 | ||||||
| 135 | C11H12O5 | 6 | pale yellow powder | Penicillium chrysogenum MT-12 | Huperzia serrata (Thunb. ex Murray) Trev. | Fujian Province, China | Exhibit inhibition of nitric oxide production in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophage cells (138,139,142,143,145,146) | [45] |
| 136 | C9H8O5 | 6 | ||||||
| 137 | C10H10O5 | 6 | yellow powder | |||||
| 138 | C15H20O6 | 6 | ||||||
| 139 | C14H16O6 | 7 | ||||||
| 140 | C15H16O6 | 8 | pale yellow powder | |||||
| 141 | C15H18O6 | 7 | ||||||
| 142 | C15H20O6 | 6 | ||||||
| 143 | C16H22O7 | 6 | yellow powder | |||||
| 144 | C15H20O5 | 6 | ||||||
| 145 | C15H20O5 | 6 | ||||||
| 146 | C15H20O5 | 6 | ||||||
| 147 | C14H12O6 | 9 | yellow powder | Cylindrocarpon sp. | fresh roots of Sapium ellipticum | Haut Plateaux region, Cameroon | [46] | |
| 148 | C14H12O7 | 9 | ||||||
| 149 | C13H10O7 | 9 | ||||||
| 150 | C14H12O6 | 9 | yellow crystals | Phomopsis sp. xy21 | leaves of the Thai mangrove Xylocarpus granatum | Trang Province, Thailand | Weak anti-HIV activity (150) | [47] |
| 151 | C15H16O7 | 8 | colorless crystals | |||||
| 152 | C15H16O7 | 8 | White amorphous solid | |||||
| 153 | C15H16O5 | 8 | ||||||
| 154 | C15H12O6 | 10 | ||||||
| 155 | C15H10O7 | 11 | ||||||
| 156 | C17H28O3 | 4 | white powder | Aspergillus flocculus | stem of the medicinal plant Markhamia platycalyx | [48] | ||
| 157 | C10H10O4 | 6 | colorless crystals | Colletotrichum gloeosporioides | mangrove Ceriops tagal | Hainan Province China | Show potent antibacterial activity (157,159) | [49] |
| 158 | C10H14O4 | 4 | brown oil | |||||
| 159 | C10H12O3 | 5 | white powder | |||||
| 160 | C14H18O4 | 6 | amorphous powder | Paraconiothyrium sp. SW-B-1 | the seaweed, Chondrus ocellatus Holmes | Yamagata Prefecture, Japan | Show moderate antibacterial activity (164) | [50] |
| 161 | C14H18O5 | 6 | ||||||
| 162 | C14H16O6 | 7 | ||||||
| 163 | C12H16O6 | 5 | ||||||
| 164 | C22H20O4 | 13 | ||||||
| 165 | C14H16O6 | 7 | pale brown, amorphous powder | Alternaria alternata MT-47 | medicinal plant of Huperzia serrata | Fujian Province, China | Exhibit inhibitory activity on the ATP release of thrombin-activated platelets (168) | [51] |
| 166 | C15H12O8 | 10 | pale yellow amorphous powder | |||||
| 167 | C18H18O9 | 10 | white amorphous powder | |||||
| 168 | C18H20O9 | 9 | ||||||
| 169 | C10H11NO4 | 6 | white gum | Chaetosphaeronema achilleae | shoots | English Yew (Taxus baccata), Iran | Weak antifungal activity and antibacterial activity (170) Cytotoxicity (169,170) Biofilm formation (169) | [52] |
| 170 | C10H10O5 | 6 | ||||||
| 171 | C27H38O6 | 9 | colorless oil | Aspergillus porosus | algal | [53] | ||
| 172 | C27H38O6 | 9 | ||||||
| 173 | C26H36O6 | 9 | ||||||
| 174 | C26H36O6 | 9 | ||||||
| 175 | C25H38O3 | 7 | colorless oil | Alternaria alternate | leaves of Psidium littorale Raddi | Fujian Province, China | [26] | |
| 176 | C29H30O10 | 15 | amorphous powder | Phoma sp. SYSU-SK-7 | healthy branch of the marine Kandelia candel | Guangxi Province, China | Show strong antibacterial activity (176) Exhibit significant antifungal and antibacterial activity (177) Show significant α-glucosidase inhibitory activity (176–178) Cytotoxicity (176) Exhibit radical scavenging activity against DPPH (179) | [54] |
| 177 | C11H14O4 | 5 | white solid | |||||
| 178 | C21H24O7 | 10 | ||||||
| 179 | C13H12O5 | 8 | ||||||
| 180 | C11H16O3 | 4 | colourless oil | |||||
| 181 | C10H14O3 | 4 | Phomopsis sp. D15a2a | leaves of Alternanthera bettzickiana (Amaranthaceae) | Anambra state of Nigeria | [55] | ||
| 182 | C11H16O4 | 4 | ||||||
| 183 | C11H16O4 | 4 | ||||||
| 184 | C23H26O7 | 11 | Penicillium purpurogenum IMM003 | fresh healthy leaves of Edgeworthia chrysantha | Zhejiang Province, China | [56] | ||
| 185 | C22H26O6 | 10 | ||||||
| 186 | C10H8O5 | 7 | ||||||
| 187 | C18H27NO4 | 6 | colorless gum | Camporesia sambuci FT1061 & Epicoccum sorghinum FT1062 | healthy fruit of the plant Rhodomyrtus tomentosa | the Big Island in Hawaii | [57] | |
| 188 | C14H20O6 | 5 | light yellow solid | Rhytismataceae sp. DAOMC 251461 | healthy P. mariana needles | New Brunswick, Canada. | Exhibit moderate antifungal activity (189) | [58] |
| 189 | C15H22O6 | 5 | ||||||
| 190 | C9H12O6 | 4 | colorless plate | Phomopsis asparagi SWUKJ5.2020 | fresh, healthy branches of medicinal plant Kadsura angustifolia | Yunnan province China | Exhibit notable cytotoxicity (192–194) | [59] |
| 191 | C9H12O5 | 4 | ||||||
| 192 | C9H10O6 | 5 | colorless crystals | |||||
| 193 | C11H14O4 | 5 | colorless plates | |||||
| 194 | C11H14O5 | 5 | ||||||
| 195 | C14H18O4 | 6 | colorless oil | Dendrothyrium variisporum | roots of the Algerian plant Globularia alypum | Ain Touta, Batna 05000 (Algeria) | [60] | |
| 196 | C18H24O5 | 7 | ||||||
| 197 | C11H10O5 | 7 | colorless oil | Alternaria sp. | twigs of Morinda offificinalis | Guangdong province China | [61] | |
| 198 | C12H11O5 | 8 | yellow oil | |||||
| 199 | C8H12O3 | 3 | colorless gum | Trichoderma atroviride | bulb of Lycoris radiata | Hubei Province China | [11] | |
| 200 | C13H14O5 | 7 | yellow viscous liquid | Eurotium chevalieri KUFA 0006 | healthy twig of Rhizophora mucronata Poir | Chanthaburi Province, Eastern Thailand | Prevent biofilm formation | [62] |
| 201 | C9H14O4 | 3 | colorless gum | Simplicillium sp. PSU-H41 | leaf of Hevea brasiliensis | Songkhla Province Thailand | [63] | |
| 202 | C15H12O8 | 10 | yellowish crystal | Cytospora rhizophorae | Morinda offificinalis | Guangdong province China | Exhibit weak growth inhibitory activity against the tumor cell lines (202) | [64] |
| 203 | C14H10O6 | 10 | brown gum | |||||
| 204 | C14H8O7 | 11 | yellowish green powder | |||||
| 205 | C13H16O5 | 6 | yellow gum | Fusarium sp. HP-2 | Chinese agarwood “Qi-Nan” | Hainan Province China | Show weak acetylcholinesterase inhibitory activity (207) | [65] |
| 206 | C14H14O4 | 8 | red crystals | |||||
| 207 | C16H18O6 | 8 | red solid | |||||
| 208 | C18H17ClO7 | 10 | yellowish powder | Penicillium citrinum HL-5126 | mangrove Bruguiera sexangula var. rhynchopetala | South China Sea | Display cytotoxic activity (209) Show weak antibacterial activity (208) | [66] |
| 209 | C18H18O7 | 10 | ||||||
| 210 | C13H16O5 | 6 | amorphous white powder | Phoma sp. PF2 | Artemisia princeps | Show moderate inhibitory activities on nitric oxide levels (210–211) | [67] | |
| 211 | C14H18O5 | 6 | ||||||
| 212 | C25H26O5 | 13 | polar yellow solid | Aspergillus sp. ASCLA | healthy leaf tissue of the medicinal plant Callistemon subulatus | Exert moderate-high activities against Staphylococcus aureus | [68] | |
| 213 | C12H18O3 | 4 | white powder | Cylindrocarpon sp. | fresh roots of Sapium ellipticum | Haut Plateaux region, Cameroon | [46] | |
| 214 | C25H28O6 | 12 | yellow oil | Diaporthe lithocarpus A740 | twigs of medicinal plant Morinda officinalis. | Guangdong province China | [41] | |
| Alkaloids and their derivatives | ||||||||
| 215 | C26H33O8N | 11 | Apiospora montagnei | lichen Cladonia sp. | [37] | |||
| 216 | C16H19NO3 | 8 | colorless amorphous solid | Chaetomium globosum CDW7 | [69] | |||
| 217 | C17H24N2O3 | 7 | colorless crystals | Penicillium citrinum HL-5126 | mangrove Bruguiera sexangula var. rhynchopetala | South China Sea | [66] | |
| 218 | C13H15NO2 | 7 | colorless powder | Bionectria sp. | seeds of the tropical plant Raphia taedigera | Haut Plateaux region, Cameroon | [70] | |
| 219 | C16H15NO5 | 10 | yellow powder | Cylindrocarpon sp. | fresh roots of Sapium ellipticum | Haut Plateaux region, Cameroon | [46] | |
| 220 | C14H21NO5 | 5 | white powder | |||||
| 221 | C26H29NO6 | 13 | pale yellow amorphous solid | Aspergillus versicolor | leaves of the Egyptian water hyacinth Eichhornia crassipes | Egypt | Exhibit moderate antiproliferative activity | [71] |
| 222 | C17H15NO8 | 11 | white amorphous solid | Pestalotiopsis flavidula | branches of Cinnamomum camphora | Yunnan province china | Moderate cytotoxicity (222–223) | [72] |
| 223 | ||||||||
| 224 | C19H24N2O2 | 9 | white amorphous powder | Irpex lacteus-A | medicinal plant Huperzia serrata | Fujian Province China | Show moderate neuroprotective activity (224–225) | [73] |
| 225 | C19H24N2O2 | 9 | ||||||
| 226 | C14H17NO3 | 7 | colorless solid | Alternaria alternate | leaves of Psidium littorale Raddi | Fujian Province, China | [26] | |
| 227 | C27H31N3O5 | 14 | brilliant yellowish oil | Fusarium sambucinum TE-6L | fresh leaves of cultivated tobacco (N. tabacum L.). N. tabacum L. | Hubei province China | Show potent inhibitory effects (227–228) Exhibit remarkable larvicidal activity (228) | [74] |
| 228 | C27H31N3O5 | 14 | white solid | |||||
| Penylpropanoids and their derivatives | ||||||||
| 229 | C10H14O5 | 4 | clear solid | Mycosphaerellaceae sp. DAOMC 250863 | healthy needles from Picea rubens (red spruce) and P. mariana (black spruce) | Eastern Canada | Show modest antibiotic activity to E. coli | [58] |
| 230 | C16H18O4 | 8 | light-yellow powder | C. globosum CDW7 | Ginkgo biloba | China | Show moderate antifungal activity | [69] |
| 231 | C12H14O4 | 6 | colorless amorphous solid | Pestalotiopsis sp. HHL-101 | fresh twigs of the mangrove plant Rhizophora stylosa | Hainan Island, China | Exhibit moderate antibacterial activity | [75] |
| 232 | C12H12O4 | 7 | white amorphous powder | Nectria pseudotrichia 120–1NP | healthy stem of Gliricidia sepium | Yogyakarta, Indonesia | [31] | |
| 233 | C13H14O4 | 7 | ||||||
| 234 | C21H12O12 | 16 | off-white amorphous solid | Aspergillus versicolor | leaves of the Egyptian water hyacinth Eichhornia crassipes | Egypt | [71] | |
| 235 | C22H14O12 | 16 | yellowish amorphous powder | |||||
| 236 | C21H22O6 | 11 | colorless crystals | Pestalotiopsis adusta | stem bark of wild rare medicinal plant Sinopodophyllum hexandrum (Royle) Ying | Qinling Mountains China | Show weak to moderate cytotoxic activity | [19] |
| 237 | C13H14O7 | 7 | white solid powder | T. harzianum Fes1712 | Rubber Tree Ficus elastica Leaves | China | Exhibit inhibitory activity against Gram-negative bacteria (237–238) | [76] |
| 238 | ||||||||
| 239 | C11H12O6 | 6 | white amorphous powder | Penicillium coffeae MA-314 | fresh inner tissue of the leaf of marine mangrove plant Laguncularia racemosa | Hainan island, China | [77] | |
| 240 | ||||||||
| 241 | C18H22O3 | 8 | yellow oil | Diaporthe sp. | branches of Pteroceltis tatarinowii Maxim | Nanjing province, China | Show modest antibacterial activity Weak cytotoxicity | [78] |
| Lactones | ||||||||
| 242 | C11H10O5 | 7 | yellowish brown solid | Alternaria sp. | seeds of the plant Ziziphus jujuba | Uzbekistan | [79] | |
| 243 | C16H26O6 | 4 | white, amorphous powder | Phaeoacremonium sp. | leaves of Senna spectabilis | Araraquara Cerrado area, Sao Paulo state, Brazil. | Exhibit antifungal activity (244–245) Cytotoxicity (244) | [80] |
| 244 | C16H26O5 | 4 | ||||||
| 245 | ||||||||
| 246 | C9H12O2 | 4 | amorphous powder | Xylaria curta 92092022 | barks | Taiwan China | Show moderate antibacterial and phytotoxic activities | [81] |
| 247 | C16H22O5 | 6 | white powder | Trichoderma sp. 307 & Acinetobacter johnsonii B2 | Strain 307, stem bark of Clerodendrum inerme | Guangdong Province, China | Exhibit potent α-glucosidase inhibitory activity (247–248) show moderate inhibitory activity against α-glucosidase (249) | [10] |
| 248 | C16H20O5 | 7 | ||||||
| 249 | colorless needles | |||||||
| 250 | C10H16O3 | 3 | colorless oil | Pestalotiopsis sp. | fruits of Drepanocarpus lunatus (Fabaceae) | [12] | ||
| 251 | C13H18O5 | 5 | ||||||
| 252 | C11H14O5 | 5 | colorless crystals | Talaromyces sp. | Xanthoparmelia angustiphylla | Stockholm, Sweden | Exhibit selective cytotoxicities | [82] |
| 253 | C32H50O7 | 8 | yellow powder | Mutant CS/asm21-4 | Maytenus hookeri | China | Exhibit antibacterial activity | [83] |
| 254 | C22H21NO4 | 13 | light yellow gum | Aspergillus terreus | Yongxing Island fresh, healthy leaves of S. maritima L. | South China Sea, China | Show strong inhibitory effects on the production of NO (256–257) | [84] |
| 255 | C22H19O4 | 14 | ||||||
| 256 | C22H21NO5 | 13 | ||||||
| 257 | C22H21NO5 | 13 | ||||||
| 258 | C20H22O3 | 10 | yellow oil | Aspergillus sp. | root of Tripterygium wilfordii | Wuhan, China | Exhibited weak AchE and BACE1 inhibitory activity (260–261) Showed excellent inhibitory effects on the production of IL-1β, TNF-α, and NO (258–261) | [85] |
| 259 | C24H26O6 | 12 | yellow oil | |||||
| 260 | C24H26O6 | 12 | colorless oil | |||||
| 261 | C23H26O6 | 11 | ||||||
| 262 | C22H36O8 | 5 | oil | H. fuscum | lichen Usnea sp. | Yunnan, China | Exhibit moderate cytotoxicity | [86] |
| 263 | C26H34O12 | 10 | white powder | Talaromyces purpurogenus | fresh leaves of the toxic medicinal plant Tylophora ovata | China | [87] | |
| 264 | C28H36O12 | 11 | ||||||
| 265 | C26H40O9 | 7 | ||||||
| 266 | C11H18O3 | 3 | yellow oil | Penicillium coffeae MA-314 | fresh inner tissue of the leaf of marine mangrove plant Laguncularia racemosa | Hainan island, China | Exhibit potent antifungal activity | [77] |
| 267 | C12H12O5 | 7 | brown solids | Phomopsis sp. | stems of Isodon eriocalyx var. laxiflflora | Kunming, China | [43] | |
| 268 | C17H14O3 | 11 | white amorphous powder | Phyllosticta sp. J13-2-12Y | leaves of Acorus tatarinowii | Guangxi Province, China | [88] | |
| 269 | ||||||||
| 270 | C19H16O5 | 12 | colorless oil | |||||
| 271 | C16H12O3 | 11 | colorless crystal | |||||
| 272 | C22H26O6 | 10 | luminous yellow oil | Pestalotiopsis microspora | fruits of Manilkara zapota | Kandy, Sri Lanka | [89] | |
| Anthraquinones | ||||||||
| 273 | C12H14O5 | 6 | yellow amorphous powder. | Penicillium citrinum Salicorn 46 | Salicornia herbacea Torr. | China | [90] | |
| 274 | C14H14O4Cl2 | 7 | yellow oil | Lachnum cf. pygmaeum DAOMC 250335 | dead P. rubens twig | NB, Canada | Inhibit the growth of M. violaceum, | [58] |
| 275 | C16H12O6 | 11 | Apiospora montagnei | lichen Cladonia sp. | [37] | |||
| 276 | C18H14O7 | 12 | yellow crystal | Eurotium chevalieri KUFA 0006 | healthy twig of Rhizophora mucronata Poir. | Chanthaburi Province, Eastern Thailand | Cause a significant reduction in biofilm production | [62] |
| 277 | C15H16O3 | 8 | Nigrospora oryzae co-cultured with Irpex lacteus | seeds of Dendrobium offifficinale | Yunnan Province China | [14] | ||
| 278 | C15H18O2 | 7 | ||||||
| 279 | C15H20O4 | 6 | ||||||
| 280 | C15H20O6 | 6 | ||||||
| 281 | C14H16O2 | 7 | Phoma betae | Kalidium foliatum (Pall.) | China | Cytotoxic activities (281) | [91] | |
| 282 | C14H16O3 | 7 | ||||||
| 283 | C14H20O5 | 5 | ||||||
| 284 | C27H24O10 | 16 | red powder | Neofusicoccum austral SYSU-SKS024 | branches of the mangrove plant Kandelia candel | Guangxi province, China | Show inhibitory effects against Indoleamine 2,3-dioxygenase (IDO) | [92] |
| 285 | C15H16O6 | 8 | yellow powder | |||||
| 286 | C14H18O5 | 6 | white powder | |||||
| 287 | C16H18O5 | 8 | yellow amorphous powder | Nectria pseudotrichia 120-1NP | healthy stem of Gliricidia sepium | Yogyakarta, Indonesia | Exhibit antibacterial activity Exhibit cytotoxicity | [31] |
| 288 | C30H22O12 | 20 | yellow powder | ARL-09 (Diaporthe sp.) | Anoectochilus roxburghii | China | Cytotoxicity Effects on NF-κB signaling pathway | [93] |
| 289 | C40H45NO10S | 19 | red powder | CS/asm21-4 | callus of Chinese medicinal plant Maytenus hookeri | China | Show moderate antimicrobial activities (antibacterial activities and antifungal activity) (289–291) | [83] |
| 290 | C40H49NO12 | 17 | yellow powder | |||||
| 291 | C40H44NO8Cl | 19 | ||||||
| 292 | C12H18O6 | 4 | colorless oil | Xylaria sp. SYPF 8246 | root of Panax notoginseng | Yunnan, China | [94] | |
| 293 | C15H14O6 | 9 | Talaromyces funiculosus | lichen thallus of Diorygma hieroglyphicum | India | Display antimicrobial activity | [95] | |
| 294 | C16H14O7 | 10 | yellow gum | Simplicillium lanosoniveum Zare & W. Gams PSU-H168 and PSU-H261 | leaves of Hevea brasiliensis | Songkhla Province, Thailand | Display antifungal activity | [96] |
| 295 | C17H18O7 | 9 | red amorphous powder | Fusarium napiforme | mangrove plant, Rhizophora mucronata | Makassar, Indonesia | Exhibit moderate antibacterial activity (295–296) Phytotoxic (295–296) | [97] |
| 296 | C16H16O6 | 9 | orange amorphous powder | |||||
| Sterides | ||||||||
| 297 | C34H52O8 | 9 | faint yellow oil | Xylaria sp. | leaves of Panax notoginseng | Yunnan province China | Show cytotoxicity (297) | [98] |
| 298 | C28H44O7 | 7 | semitransparent oil | |||||
| 299 | C25H36O5 | 8 | colorless needle | Chaetomium sp. M453 | Chinese herbal medicine Huperzia serrata | Yunnan Province, China | Show weak acetylcholinesterase inhibitory activity (302) | [99] |
| 300 | C25H36O5 | 8 | colorless amorphism | |||||
| 301 | C25H34O5 | 9 | ||||||
| 302 | C28H42O3 | 8 | yellow oil | |||||
| 303 | C22H32O3 | 7 | colorless crystals | Stemphylium sp. AZGP4–2 | root of Polyalthia laui | Hainan Province China | Show antibacterial activity against Escherichia coli (303) Exhibit antibacterial activity (304) | [100] |
| 304 | C23H36O3 | 6 | ||||||
| 305 | C23H34O3 | 7 | colorless needle crystals | |||||
| 306 | C44H72O2 | 9 | white amorphous powder | Fusarium sp. | Mentha longifolia L. (Labiatae) roots | Saudi Arabia | Possessed 5-LOX inhibitory potential (306–308) | [101] |
| 307 | C28H46O3 | 6 | ||||||
| 308 | C30H48O5 | 7 | ||||||
| 309 | C28H40O2 | 9 | colorless powder | Pleosporales sp. F46 and Bacillus wiedmannii. Com1 | medicinal plant Mahonia fortunei | Qingdao, China. | Exhibit moderate antibacterial efficacy | [102] |
| 310 | C32H41NO3 | 13 | white power | Aspergillustubingensis YP-2 | bark of Taxus yunnanensis | Yunnan Province, China | Show weak cytotoxicities (311) | [103] |
| 311 | C22H34O3 | 6 | ||||||
| Other types of compounds | ||||||||
| 312 | C21H24O6 | 10 | colorless oil | Talaromyces stipitatus SK-4 | leaves of a mangrove plant Acanthus ilicifolius | Guangxi Province, China | Show antibacterial activity and inhibitory against α-glucosidase (313) | [104] |
| 313 | C23H26O7 | 11 | ||||||
| 314 | C15H21NO8 | 6 | whitish needles | C. ninchukispora BCRC 31900 | seeds of medicinal plant Beilschmiedia erythrophloia Hayata | Taiwan China | Show anti-inflammatory effects through inhibition of NO production (317,314–315) | [105] |
| 315 | C15H21NO7 | 6 | ||||||
| 316 | C16H23NO7 | 6 | ||||||
| 317 | C15H21NO8 | 6 | yellowish solid | |||||
| 318 | C15H16O5 | 8 | white amorphous powder | Pyronema sp. (A2-1 & D1-2) | Taxus mairei | Hubei province, China | Exhibit moderate antibiotic activity | [106] |
| 319 | C11H16O4 | 4 | yellow oil | Phoma sp. nov. LG0217 | branches of Parkinsonia microphylla | Tucson, Arizona | [107] | |
| 320 | C12H16O4 | 5 | colorless amorphous powder | Penicillium citrinum Salicorn 46 | Salicornia herbacea Torr | China | Exhibit potent cytotoxic activity | [90] |
| 321 | C21H29NO9 | 8 | colorless gum | Phomopsis sp. PSU-H188 | midrib of Hevea brasiliensis | Trang Province, Thailand | [108] | |
| 322 | C20H28O7 | 7 | ||||||
| 323 | C21H27O6N | 9 | yellow amorphous solid | Fusarium solani JK10 | root of the Ghanaian medicinal plant Chlorophora regia | Eastern Region of Ghana | Exhibit antibacterial efficacies (325–326,328) | [109] |
| 324 | ||||||||
| 325 | C21H27O7N | 9 | ||||||
| 326 | ||||||||
| 327 | C21H25O8N | 10 | ||||||
| 328 | C22H29O7N | 9 | pale yellow amorphous solid | |||||
| 329 | C22H29O5N | 9 | yellow amorphous solid | |||||
| 330 | C14H14O4 | 8 | colourless oil | Phomopsis longicolla HL-2232 | fresh healthy leaf of Brguiera sexangula var. rhynchopetala | South China Sea | Show moderate antibacterial activities | [110] |
| 331 | C9H16O4 | 2 | white needles | Penicillium sp. OC-4 | leaves of Orchidantha chinensis | Guangdong Province, China | Display strong antioxidant activity | [111] |
| 332 | C16H24O6 | 5 | colorless, amorphous solid | Curvularia sp. | leaf of the medicinal plant Murraya koenigii | Bangladesh | Exhibit zoospore motility impairment activity (333–334) | [112] |
| 333 | C12H18O6 | 4 | ||||||
| 334 | C10H12O3 | 5 | colorless crystals | |||||
| 335 | C10H16O4 | 3 | colorless oil | |||||
| 336 | C20H16O5 | 13 | yellow viscous oil | Rhytidhysteron rufulum AS21B | leaves of Azima armentosa | Samutsakhon province, Thailand | Display the most promising anti-tumor activity (337) | [113] |
| 337 | C22H18O5 | 14 | pale yellow gum | |||||
| 338 | C11H12O4 | 6 | brown solids | Phomopsis sp. sh917 | stems of Isodon eriocalyx var. laxiflora | Kunming, China | [43] | |
| 339 | C15H15NO3 | 9 | brown gum | Dendrothyrium variisporum | roots of the Algerian plant Globularia alypum | Algeria | Show the strongest activity against Bacillus subtilis and Micrococcus luteus (339) | [60] |
| 340 | C14H13NO2 | 9 | ||||||
| 341 | C12H17NO3 | 5 | ||||||
| 342 | C15H18N2O4 | 8 | light yellow gum | Trichoderma atroviride | bulb of Lycoris radiata | china | [11] | |
| 343 | C32H34N2O4 | 17 | yellow crystal. | Penicillium chrysogenum V11 | vein of Myoporum bontioides A. Gray | Leizhou Peninsula, China | Display significant antifungal activity and remarkable cytotoxicities | [114] |
| 344 | C14H15NO | 8 | yellow crystal | Eurotium chevalieri KUFA 0006 | healthy twig of Rhizophora mucronata Poir. | Chanthaburi Province, Eastern Thailand | Show inhibition of biofilm production (344–345) | [62] |
| 345 | C14H15NO | 8 | yellowish viscous liquid | |||||
| 346 | C13H15NO3 | 7 | ||||||
| 347 | C18H18O6 | 10 | colorless solid | Simplicillium sp. PSU-H41 | leaf of Hevea brasiliensis (Euphorbiaceae) | Songkhla, Thailand | Display weak antibacterial against Staphylococcus aureus (347) Exhibit weak antifungal activity against Cryptococcus neoformans (349) | [63] |
| 348 | C19H20O6 | 10 | pale yellow solid | |||||
| 349 | C20H20O6 | 11 | ||||||
| 350 | C25H24O7 | 14 | ||||||
| 351 | ||||||||
| 352 | C25H22O8 | 15 | yellow gum | |||||
| 353 | C24H26O7 | 12 | pale yellow gum | |||||
| 354 | C34H30O11 | 20 | colorless solid | |||||
| 355 | C31H28O8 | 18 | pale yellow gum | |||||
| 356 | C17H24N2O6 | 7 | colorless viscous oil | Phoma herbarum PSU-H256 | leaf of Hevea brasiliensis | Songkhla, Thailand | [115] | |
| 357 | C12H13NO6 | 7 | ||||||
| 358 | C16H19NO7 | 8 | ||||||
| 359 | C15H17NO5 | 8 | ||||||
| 360 | C7H12N2O3 | 3 | ||||||
| 361 | C14H14N2O5 | 9 | ||||||
| 362 | C11H12O3 | 6 | white amorphous solid. | Penicillium sp. | leaf of Senecio flavus (Asteraceae) | Al-Azhar University Egypt | Show antifungal activity and cytotoxic activity | [116] |
| 363 | C30H37NO7 | 13 | white amorphous powder | R. sanctae-cruciana | leaves of the medicinal plant A. lebbeck. | India | Show considerable cytotoxic potential | [117] |
| 364 | C24H30O4 | 10 | yellowish oil | Arthrinium arundinis TE-3 | fresh leaves of cultivated tobacco | Hubei Province China | Show selective antifungal activity (364–365) Display moderate in vitro cytotoxicity (365) | [118] |
| 365 | C20H24O4 | 9 | ||||||
| 366 | C20H24O3 | 9 | ||||||
| 367 | C23H24O5 | 12 | brown powder | Aspergillus flavipes Y-62 | stems of plant Suaeda glauca (Bunge) Bunge | Zhejiang province, East China | Show weak antimicrobial activity | [119] |
| 368 | C16H14O6 | 9 | colorless crystals | Mycosphaerella sp. (UFMGCB2032) | healthy leaves of Eugenia bimarginata | Atlanta, GA, USA | Exhibit moderate antifungal activities | [120] |
| 369 | C17H18O9 | 9 | colorless solid | |||||
| 370 | C20H16O5 | 13 | off-white gum | Anteaglonium sp. FL0768 | Living photosynthetic tissue of sand spikemoss (Selaginella arenicola; Selaginellaceae) | [121] | ||
| 371 | C28H26N2O5 | 17 | amorphous light yellow powder | Penicillium janthinellum SYPF 7899 | three-year-old healthy P. notoginseng | Yunnan province, China | Exhibit significant inhibitory activities (371–373) | [122] |
| 372 | ||||||||
| 373 | C15H19NO6 | 7 | brown oil | |||||
| 374 | C14H24O4 | 3 | colorless oil | Phaeophleospora vochysiae sp. nov | Vochysia divergens | wetland in Brazil | Show considerable antimicrobial activity | [123] |
| 375 | C12H17NO6 | 5 | colorless oil | Bionectria sp. | fresh seeds of R. teadigera | Haut Plateaux region, Cameroon | [70] | |
| 376 | C18H14N2O6 | 13 | white powder | |||||
| 377 | C13H19NO4 | 5 | yellowish oil | Trichoderma atroviride S361 | bark of Cephalotaxus fortunei | Zhejiang province, China | [17] | |
| 378 | ||||||||
| 379 | C18H20O7 | 9 | amorphous powder | Xylaria sp. SYPF 8246 | root of Panax notoginseng | Wenshan, Yunnan, China | Display significant inhibitory activities against human carboxylesterase 2 (hCE 2) (379,383–385) | [94] |
| 380 | C12H10O5 | 8 | colorless oil | |||||
| 381 | C12H18O6 | 4 | ||||||
| 382 | C12H20O5 | 3 | ||||||
| 383 | C19H22O7 | 9 | ||||||
| 384 | C19H21O7Cl | 9 | ||||||
| 385 | C18H19O7Cl | 9 | ||||||
| 386 | C32H42O4 | 12 | brown oil | Byssochlamys spectabilis | leaf tissue of the medicinal plant Edgeworthia chrysantha | Zhejiang Province, China | weakly active against Escherichia coli and Staphyloccocus aureus (388) Display selective inhibitory effects toward hCE2-mediated FD hydrolysis (386) | [124] |
| 387 | C16H22O3 | 7 | yellow oil | |||||
| 388 | C16H26O2 | 5 | ||||||
| 389 | C20H29N5O6 | 9 | white amorphous powder | Fusarium chlamydosporium | Anvillea garcinii (Burm.f.) DC. leaves | Egypt | Exhibit selective antifungal activity and cytotoxic effect possess high antibacterial potential | [125] |
| 390 | C15H16N2O2 | 9 | Annulohypoxylon stygium | red seaweed Bostrychia radicans | Ubatuba city, São Paulo State, Brazil | [126] | ||
| 391 | C23H16O2N2 | 17 | purple-red powder | Alternaria alternata Shm-1 | fresh wild body of Phellinus igniarius | Shanxi Province, China | [127] | |
| 392 | C10H12O6 | 5 | colorless crystals | Cladosporium sp. JS1–2 | mangrove Ceriops tagal | Hainan Province in China | Show moderate antibacterial activities (392–393) Showed growth inhibition activities against newly hatched larvae of H. armigera Hubner (392–393) | [128] |
| 393 | C10H14N2O2 | 5 | yellow powder | |||||
| 394 | C8H13NO4 | 3 | white solid | Diaporthe vochysiae sp. nov. (LGMF1583) | medicinal plant Vochysia divergens | Display considerable antibacterial activity (395) Show low to moderate cytotoxic activity (394–395) | [129] | |
| 395 | C11H17NO4 | 4 | white solid | |||||
| 396 | C28H40O6 | 9 | yellow oil | Diaporthe lithocarpus A740 | Twigs of medicinal plant Morinda officinalis | Guangdong province , China | Show weak cytotoxic activity (396–397) | [41] |
| 397 | C28H40O6 | 9 | ||||||
| 398 | C30H37O7N | 13 | colorless powder | Xylaria longipes | Ailao Moutain | [130] | ||
| 399 | C30H39O9N | 12 | ||||||
| 400 | C32H41O8N | 13 | ||||||
| 401 | ||||||||
| 402 | C30H37NO7 | 13 | ||||||
| 403 | C18H18O7 | 10 | Penicillium citrinum | Parmotrema sp. | Hakgala montane forest in Sri Lanka | Show moderate antioxidant activity | [131] | |
| 404 | C11H11ClO5 | 6 | Periconia macrospinosa KT3863 | a terrestrial herbaceous plant | Kanagawa prefecture, Japan | [132] | ||
| 405 | C12H13ClO4 | 6 | ||||||
| 406 | C7H12O3 | 2 | light yellow liquid | Lasiosdiplodia pseudotheobromae | Exhibite XO inhibition (407) oxidized form of 406 show high XO inhibition | [133] | ||
| 407 | C13H22O3 | 3 | ||||||
| 408 | C17H16O8 | 10 | pale-yellow needles | Pleosporales sp. SK7 | leaves of the mangrove plant Kandelia candel | Guangxi Province, China | [23] | |
| 409 | C15H19N2O2 | 8 | faint yellow oil | Aspergillus sp. AV-2 | inner healthy leaves of mangrove plant Avicennia marina | Hurghada, Egypt | [134] | |
| 410 | C19H22O5 | 9 | yellow powder | |||||
| 411 | C10H14O3 | 4 | yellowish oil | Irpex lacteus DR10-1 | Roots of waterlogging tolerant plant Distylium chinense | Chongqing in the TGR area, China | Exhibit strong antioxidant activity (413) Show moderate antibacterial activity (411–413) | [24] |
| 412 | C10H14O3 | 4 | ||||||
| 413 | C12H16O4 | 5 | brown flaky solid | |||||
| 414 | C33H50O6 | 9 | pale yellow oil | Penicillium crustosum PRB-2 & Xylaria sp. HDN13-249 | Xylaria sp. HDN13-249: root of Sonneratia caseolaris | Hainan province, China | Show antibacterial activity (415–416) Show promising activity against M. phlei (416) | [135] |
| 415 | C33H50O9S | 9 | ||||||
| 416 | C24H40O5 | 5 | pale yellow oils | Xylaria sp. HDN13-249 | ||||
| 417 | C24H40O8S | 5 | ||||||
| 418 | C9H14O2 | 3 | colorless oil | Aspergillus terreus EN-539 & Paecilomyces lilacinus EN-531 | inner tissues of the marine red alga Laurencia okamurai | China | Exhibit inhibitory activity against bacteria and fungi | [136] |
| 419 | C23H20O5 | 14 | white powder | Diaporthe lithocarpus | leaves of Artocarpus heterophyllus | Dortmund, Germany | [137] | |
| 420 | C16H20N2O4 | 8 | colourless oil | Aspergillus aculeatus F027 | fresh leaves of Ophiopogon japonicus (Linn. f.) Ker-Gawl | Hubei province of China | [138] | |
| 421 | C17H20O6 | 8 | reddish oil | Fusarium solani B-18 | inner tissue of the unidentifified forest litters | Mount Merapi area Sleman, Yogyakarta, Indonesia. | Activat a signaling pathway in osteoclastic differentiation of murine macrophage (421) | [139] |
| 422 | C17H20O6 | 8 | yellow oil | |||||
| 423 | C15H18O5 | 7 | reddish oil | |||||
| 424 | C15H18O5 | 7 | pale-yellow oil | |||||
| 425 | C16H20O5 | 7 | amorphous powder | Hypoxylon fuscu | lichen Usnea sp. | Lilong Snow Mountain in Lijiang, Yunnan, China | Exhibit moderate cytotoxicity (426–427) | [86] |
| 426 | C21H36O6 | 4 | white solid | |||||
| 427 | C18H30O7 | 4 | white powder | |||||
| 428 | C18H28O6 | 5 | ||||||
| 429 | C25H24O6 | 14 | colorless gum | Simplicillium lanosoniveum (J.F.H. Beyma) Zare & W. Gams PSU-H168 and PSU-H261 | leaves of Hevea brasiliensis | Songkhla Province, Thailand | Exhibit antibacterial activity (430) Display antifungal activity (430–431) | [96] |
| 430 | C32H34O8 | 16 | ||||||
| 431 | C16H14O7 | 10 | yellow gum | |||||
| 432 | C14H20O4 | 5 | white amorphous powder | Clonostachys rosea B5-2 | mangrove plants | Garut, Indonesia | Exhibit phytotoxicity against lettuce seedlings (432) | [140] |
| 433 | C7H10O3 | 3 | colourless oil | |||||
| 434 | C9H12O3 | 4 | white amorphous powder | |||||
| 435 | C9H14O4 | 3 | ||||||
| 436 | C26H32O12 | 11 | white powder | Talaromyces purpurogenus | fresh leaves of the toxic medicinal plant Tylophora ovata | Guangxi Province, China | Show significant inhibitory activity against NO production in LPS-induced RAW264.7 cells (436) Show moderate inhibitory activities toward XOD and PTP1b (437,441) | [87] |
| 437 | C26H38O11 | 8 | white powder | |||||
| 438 | C27H28O8 | 14 | white powders | |||||
| 439 | C29H40O9 | 10 | ||||||
| 440 | C27H40O7 | 8 | ||||||
| 441 | C26H34O7 | 10 | ||||||
| 442 | C22H32N4O5 | 9 | white powder | Phomopsis sp. D15a2a | leaves of Alternanthera bettzickiana (Amaranthaceae) | Anambra state of Nigeria | [55] | |
| 443 | C8H13NO5 | 3 | ||||||
| 444 | C20H38O7 | 2 | colorless oil | Aureobasidium pullulans AJF1 | flower of Aconitum carmichaeli, | Jangbaek Mountain, Gangwon-do, Korea | [141] | |
| 445 | C30H56O10 | 3 | ||||||
| 446 | C16H14O8 | 10 | yellow amorphous powder | Alternaria alternata JS0515 | Vitex rotundifolia (beach vitex) | Suncheon, Korea | [142] | |
| 447 | C23H27O5Cl | 10 | colorless oil | Armillaria sp. & Epicoccum sp. YUD17002 | YUD17002: rhizomes of the underground portion of Gastrodia elata | Yunnan Province, China | Exhibit moderate in vitro cytotoxic activities (447) Show weak acetylcholinesterase Inhibitory activity (447) | [27] |
| 448 | C10H10O4 | 6 | white amorphous powder | |||||
| 449 | C14H20O9 | 5 | light-yellow oil | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study. Int. J. Mol. Sci. 2021, 22, 959. https://doi.org/10.3390/ijms22020959
Zheng R, Li S, Zhang X, Zhao C. Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study. International Journal of Molecular Sciences. 2021; 22(2):959. https://doi.org/10.3390/ijms22020959
Chicago/Turabian StyleZheng, Ruihong, Shoujie Li, Xuan Zhang, and Changqi Zhao. 2021. "Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study" International Journal of Molecular Sciences 22, no. 2: 959. https://doi.org/10.3390/ijms22020959
APA StyleZheng, R., Li, S., Zhang, X., & Zhao, C. (2021). Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study. International Journal of Molecular Sciences, 22(2), 959. https://doi.org/10.3390/ijms22020959