
miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2



Abstract
:1. Introduction
2. Results
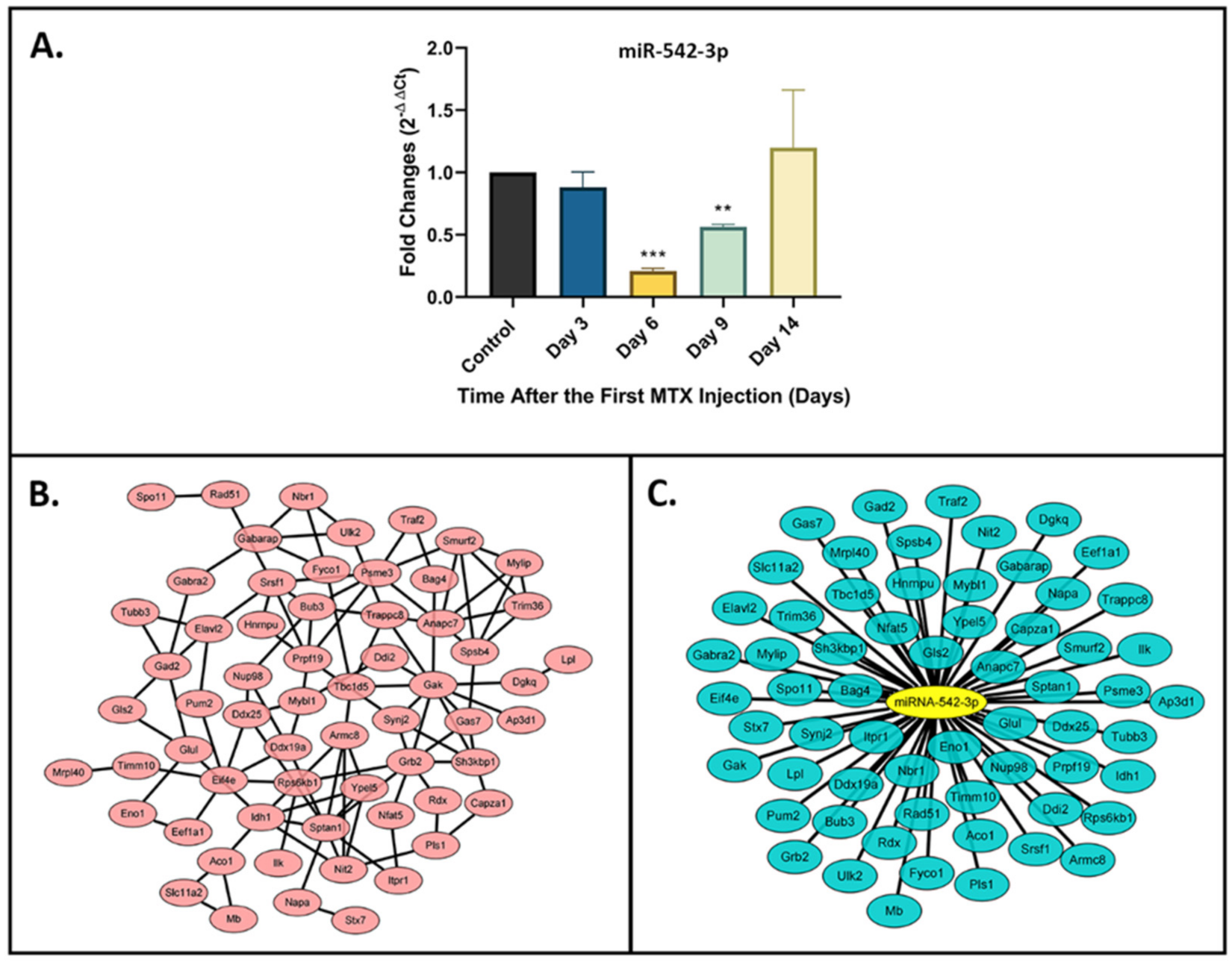
2.1. Downregulation of miR-542-3p in Bones of MTX-Treated Rats
2.2. Target Gene Prediction and Network Analyses for miR-542-3p
2.3. Functional Annotation and Enrichment Analyses of miR-542-3p
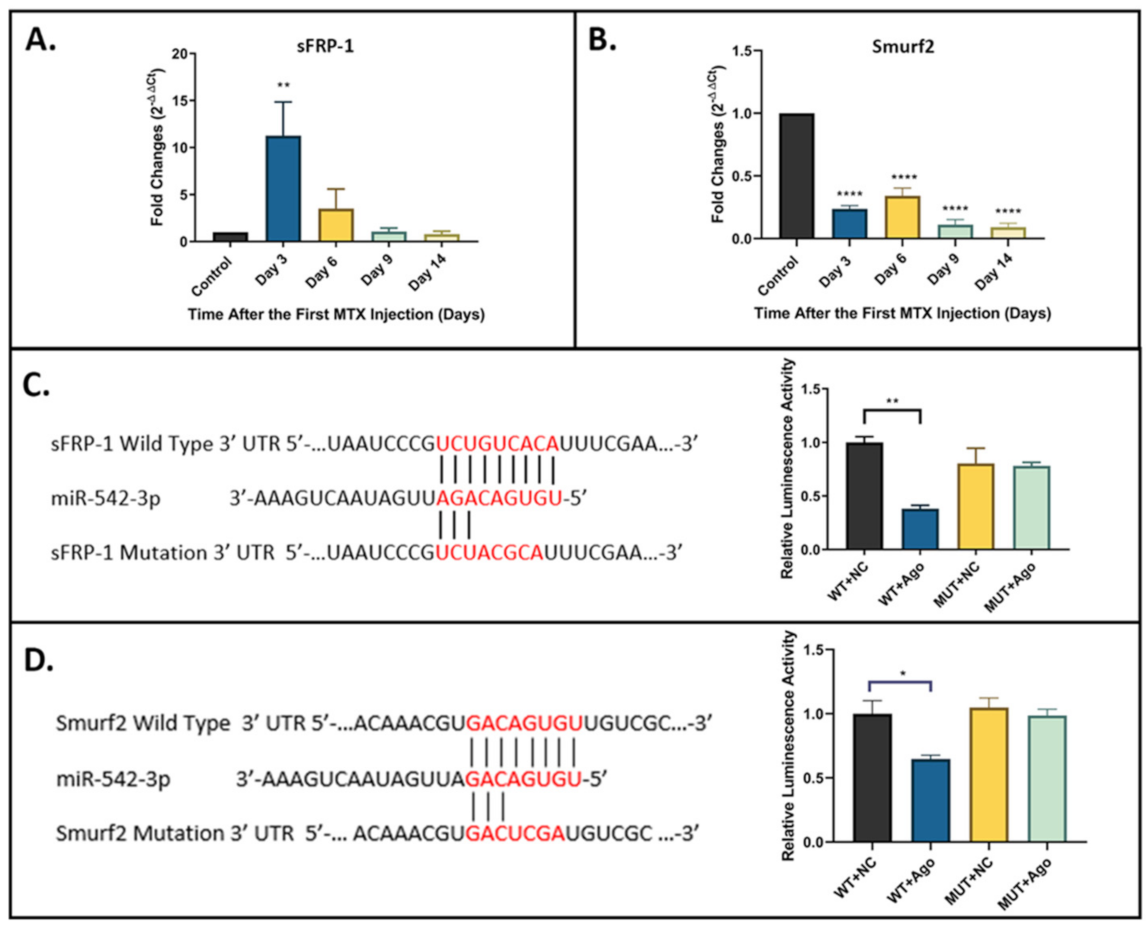
2.4. sFRP-1 and Smurf2 as Direct Targets of miR-542-3p
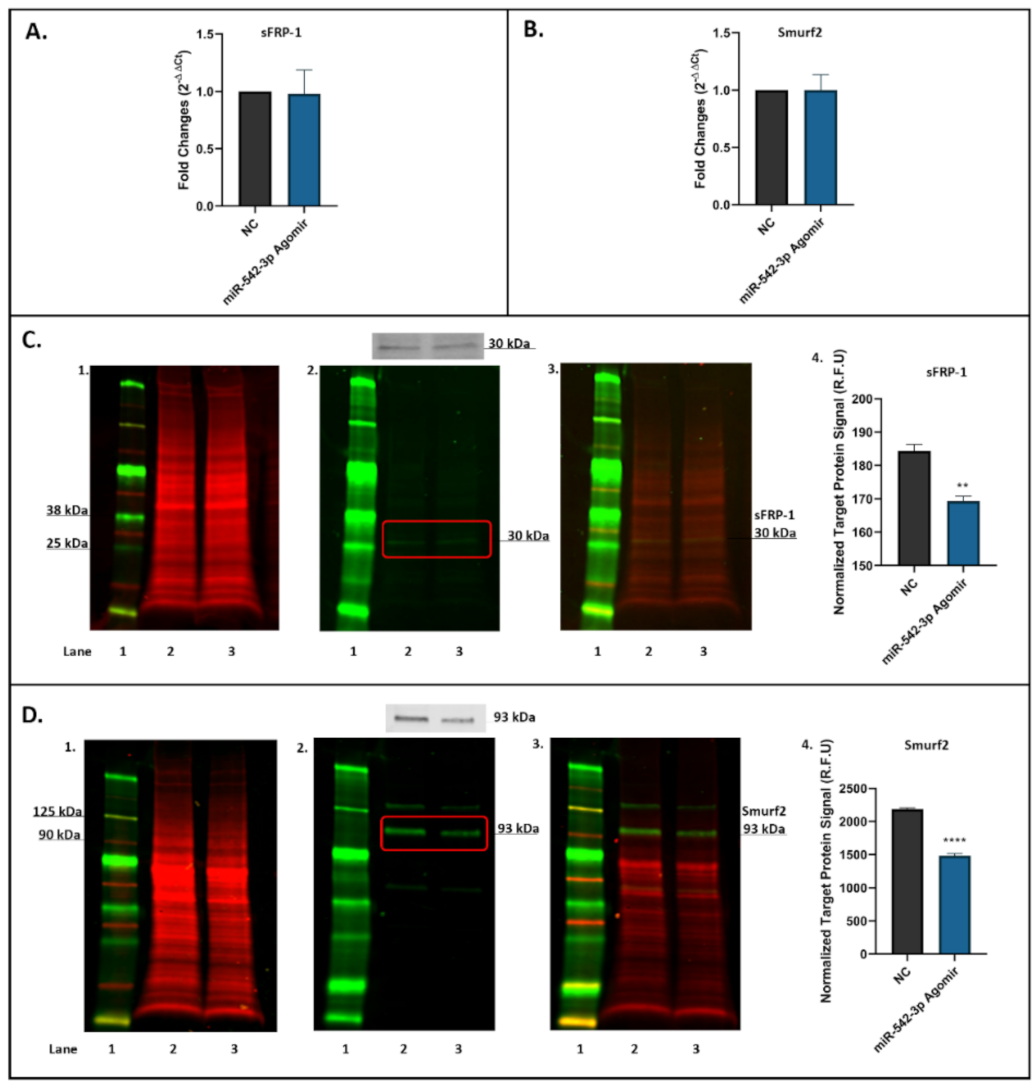
2.5. miR-542-3p Inhibits sFRP-1 and Smurf2 Expression in Osteoblastic Cells
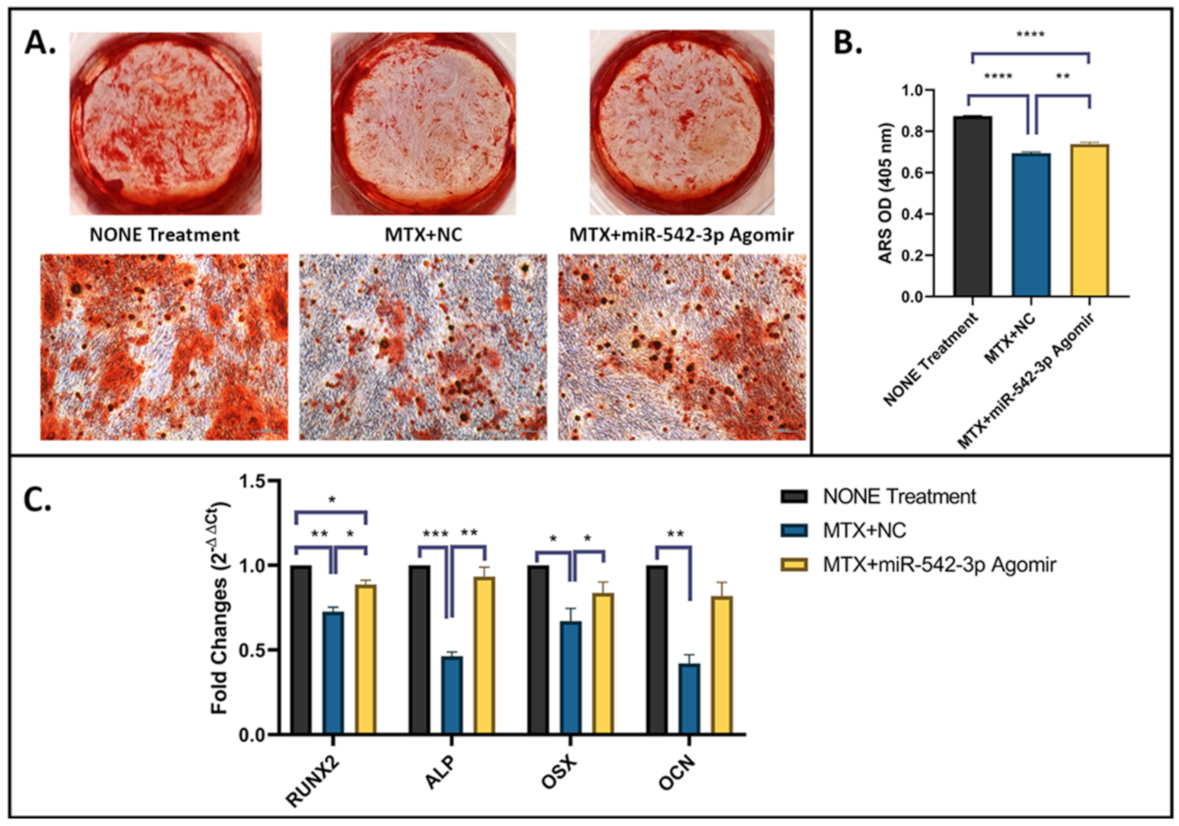
2.6. miR-542-3p Enhances Osteogenesis and Mineralization after MTX Treatment
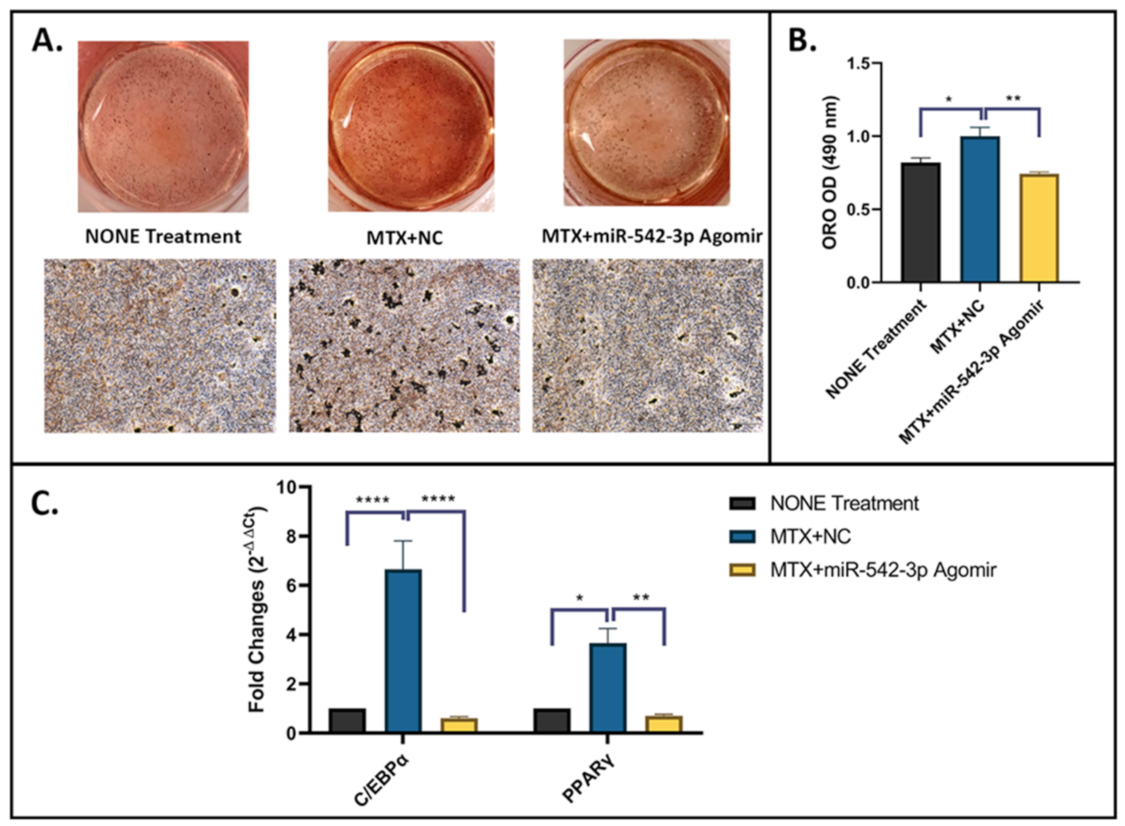
2.7. miR-542-3p Alleviates Increased Adipogenesis after MTX Treatment
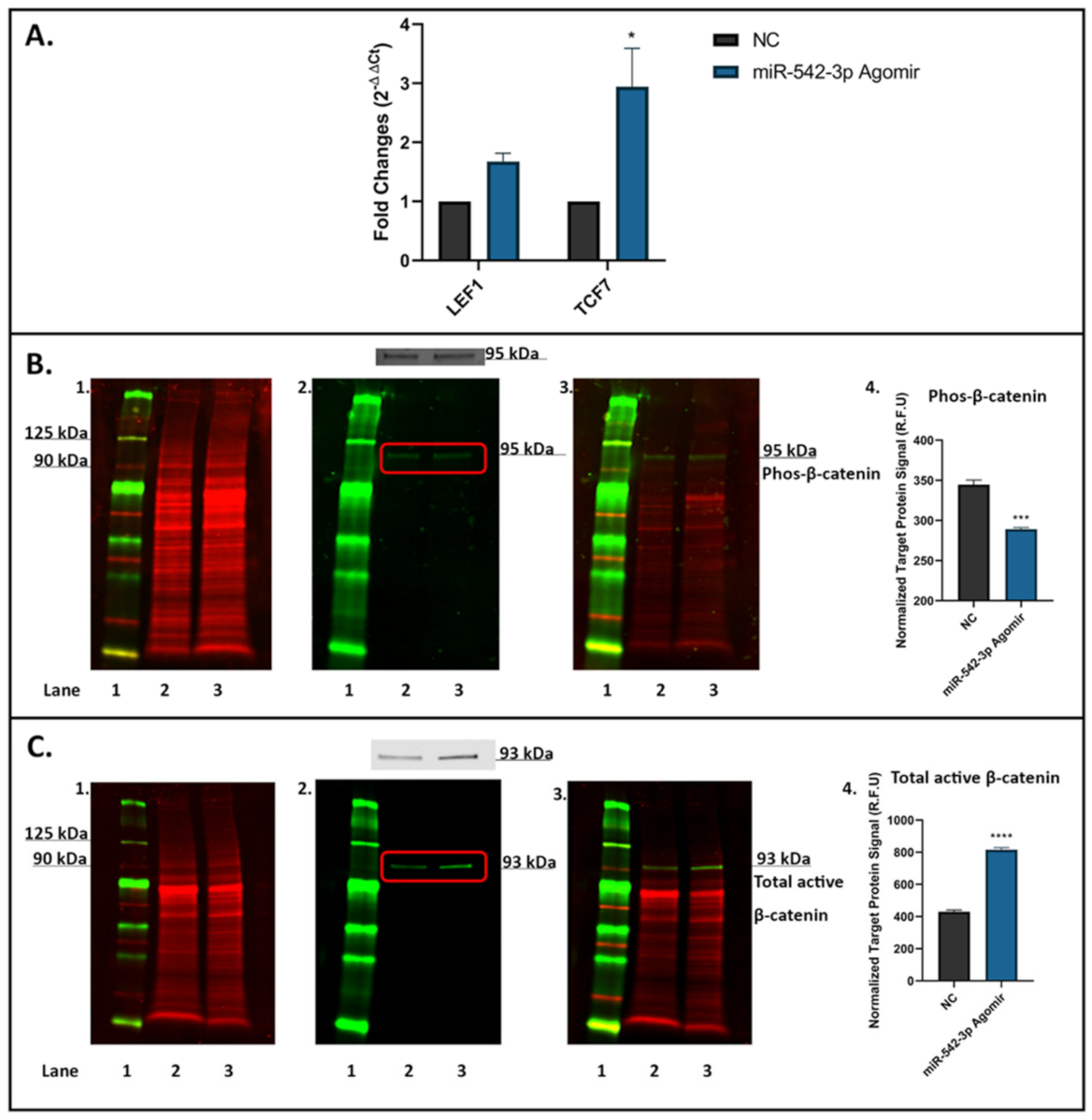
2.8. miR-542-3p Activates Wnt/β-Catenin Signalling in Osteoblastic Cells
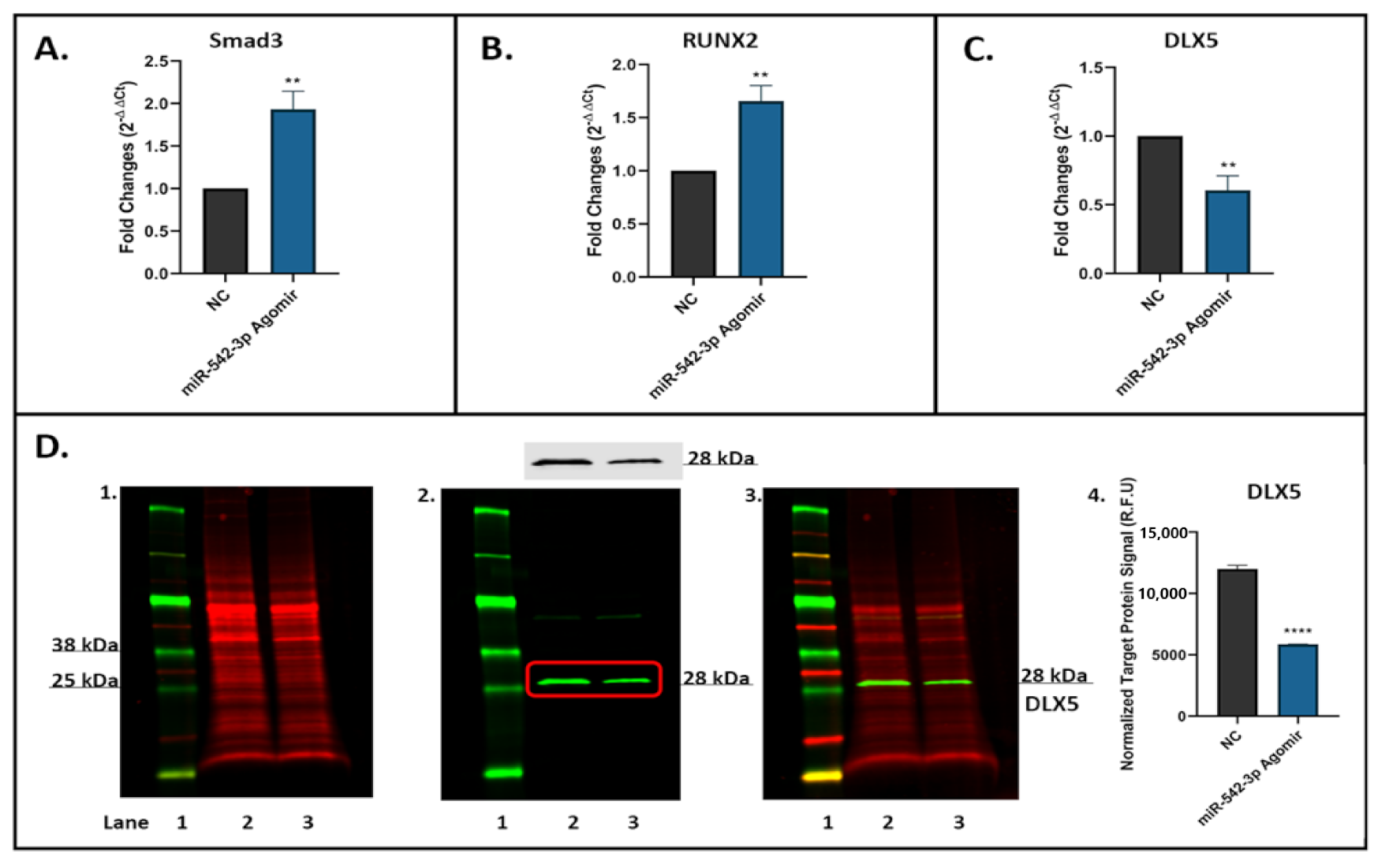
2.9. miR-542-3p Inhibits TGF-β Signalling in Osteoblastic Cells
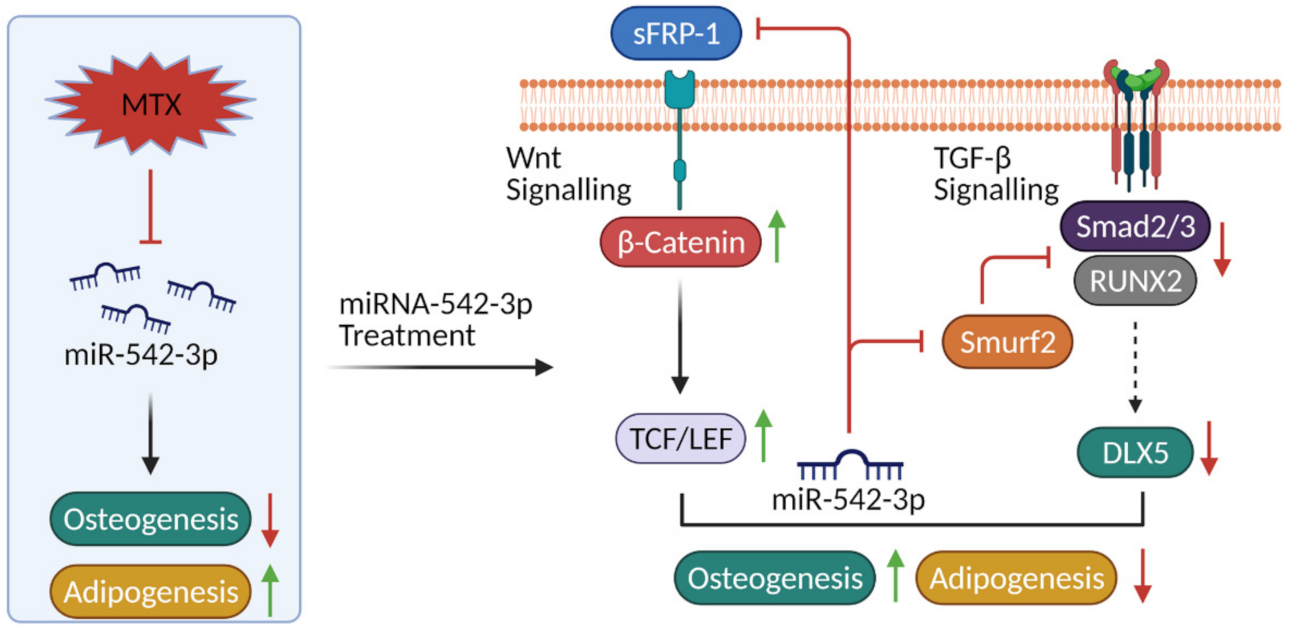
3. Discussion
3.1. miR-542-3p Is Downregulated in Bones of MTX-Treated Rats and sFRP-1 and Smurf2 Are Direct Targets of miR-542-3p
3.2. miR-542-3p Attenuates MTX-Induced Changes in Osteogenesis/Adipogenesis Associated with Alterations in Wnt/b-Catenin Signalling and TGF-b Signalling
4. Materials and Methods
4.1. MTX Treatment in Rats
4.2. RNA Isolation and RT-qPCR
4.3. Prediction of Target Genes for MiR-542-3p
4.4. Network Construction
4.5. Functional Annotation and Enrichment Analyses
4.6. Cell Culture and In Vitro Osteoblast Differentiation
4.7. Alizarin Red S (ARS) Staining for Mineralization
4.8. Cell Culture and In Vitro Adipocyte Differentiation
4.9. Oil Red O (ORO) Staining for Adipogenesis
4.10. Dual-Luciferase Assays
4.11. Western Blot Analyses
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Braun, J. Methotrexate: Optimizing the efficacy in rheumatoid arthritis. Ther. Adv. Musculoskelet Dis. 2011, 3, 151–158. [Google Scholar] [CrossRef]
- Fotoohi, A.K.; Albertioni, F. Mechanisms of antifolate resistance and methotrexate efficacy in leukemia cells. Leuk. Lymphoma 2008, 49, 410–426. [Google Scholar] [CrossRef] [PubMed]
- Hani, N.; Casper, C.; Groth, W.; Krieg, T.; Hunzelmann, N. Stevens-Johnson syndrome-like exanthema secondary to methotrexate histologically simulating acute graft-versus-host disease. Eur. J. Dermatol. 2000, 10, 548–550. [Google Scholar] [PubMed]
- Wojtuszkiewicz, A.; Peters, G.J.; van Woerden, N.L.; Dubbelman, B.; Escherich, G.; Schmiegelow, K.; Sonneveld, E.; Pieters, R.; van de Ven, P.M.; Jansen, G.; et al. Methotrexate resistance in relation to treatment outcome in childhood acute lymphoblastic leukemia. J. Hematol. Oncol. 2015, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Patel, G.; Johnson, D.; Ogunremi, L.; Barron, J. Effect of low dose weekly methotrexate on bone mineral density and bone turnover. Ann. Rheum. Dis. 2003, 62, 186–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westhovens, R.; Dequeker, J. Rheumatoid arthritis and osteoporosis. Zeitschrift für Rheumatologie 2000, 59 (Suppl. 1), 33–38. [Google Scholar] [CrossRef]
- Georgiou, K.R.; King, T.J.; Scherer, M.A.; Zhou, H.; Foster, B.K.; Xian, C.J. Attenuated Wnt/beta-catenin signalling mediates methotrexate chemotherapy-induced bone loss and marrow adiposity in rats. Bone 2012, 50, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, K.R.; Hui, S.K.; Xian, C.J. Regulatory pathways associated with bone loss and bone marrow adiposity caused by aging, chemotherapy, glucocorticoid therapy and radiotherapy. Am. J. Stem Cells 2012, 1, 205–224. [Google Scholar]
- Lai, E.C. Micro RNAs are complementary to 3′ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- Tornesello, M.L.; Faraonio, R.; Buonaguro, L.; Annunziata, C.; Starita, N.; Cerasuolo, A.; Pezzuto, F.; Tornesello, A.L.; Buonaguro, F.M. The Role of microRNAs, Long Non-coding RNAs, and Circular RNAs in Cervical Cancer. Front. Oncol. 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.G.; Wang, X.B.; Zhao, H.; Zhou, C.N. MicroRNA-9-5p promotes osteoporosis development through inhibiting osteogenesis and promoting adipogenesis via targeting Wnt3a. Eur. Rev. Med. Pharm. Sci. 2019, 23, 456–463. [Google Scholar] [CrossRef]
- Li, H.; Fan, J.; Fan, L.; Li, T.; Yang, Y.; Xu, H.; Deng, L.; Li, J.; Li, T.; Weng, X.; et al. MiRNA-10b Reciprocally Stimulates Osteogenesis and Inhibits Adipogenesis Partly through the TGF-beta/SMAD2 Signaling Pathway. Aging Dis. 2018, 9, 1058–1073. [Google Scholar] [CrossRef] [Green Version]
- Seenprachawong, K.; Tawornsawutruk, T.; Nantasenamat, C.; Nuchnoi, P.; Hongeng, S.; Supokawej, A. miR-130a and miR-27b Enhance Osteogenesis in Human Bone Marrow Mesenchymal Stem Cells via Specific Down-Regulation of Peroxisome Proliferator-Activated Receptor gamma. Front. Genet. 2018, 9, 543. [Google Scholar] [CrossRef] [PubMed]
- Karbiener, M.; Fischer, C.; Nowitsch, S.; Opriessnig, P.; Papak, C.; Ailhaud, G.; Dani, C.; Amri, E.Z.; Scheideler, M. microRNA miR-27b impairs human adipocyte differentiation and targets PPARgamma. Biochem. Biophys. Res. Commun. 2009, 390, 247–251. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Liu, L.; Peymanfar, Y.; Anderson, P.; Xian, C.J. Roles of MicroRNAs in Osteogenesis or Adipogenesis Differentiation of Bone Marrow Stromal Progenitor Cells. Int. J. Mol. Sci. 2021, 22, 7210. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-P.; Yao, J.; Guan, J.; Zhou, Z.-Q.; Zhang, Z.-Y.; Yang, J. MicroRNA-542-3p functions as a tumor suppressor via directly targeting survivin in hepatocellular carcinoma. Biomed. Pharmacother. 2018, 99, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, M.-H.; Zhou, J.-D.; Chi, Q. Upregulation of miR-542-3p inhibits the growth and invasion of human colon cancer cells through PI3K/AKT/survivin signaling. Oncol. Rep. 2017, 38, 3545–3553. [Google Scholar] [CrossRef] [Green Version]
- Kureel, J.; Dixit, M.; Tyagi, A.M.; Mansoori, M.N.; Srivastava, K.; Raghuvanshi, A.; Maurya, R.; Trivedi, R.; Goel, A.; Singh, D. miR-542-3p suppresses osteoblast cell proliferation and differentiation, targets BMP-7 signaling and inhibits bone formation. Cell Death Dis. 2014, 5, e1050. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Y.; Zhang, C.; Liu, J.; Sun, T.; Li, D.; Na, Q.; Xian, C.J.; Wang, L.; Teng, Z. miR-542-3p prevents ovariectomy-induced osteoporosis in rats via targeting SFRP1. J. Cell. Physiol. 2018, 233, 6798–6806. [Google Scholar] [CrossRef] [Green Version]
- Xian, C.J.; Cool, J.C.; Scherer, M.A.; Macsai, C.E.; Fan, C.; Covino, M.; Foster, B.K. Cellular mechanisms for methotrexate chemotherapy-induced bone growth defects. Bone 2007, 41, 842–850. [Google Scholar] [CrossRef]
- Pala, E.; Denkçeken, T. Differentially expressed circulating miRNAs in postmenopausal osteoporosis: A meta-analysis. Biosci. Rep. 2019, 39, BSR20190667. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Macsai, C.E.; Georgiou, K.R.; Foster, B.K.; Zannettino, A.C.; Xian, C.J. Microarray expression analysis of genes and pathways involved in growth plate cartilage injury responses and bony repair. Bone 2012, 50, 1081–1091. [Google Scholar] [CrossRef]
- Zimmerli, D.; Hausmann, G.; Cantu, C.; Basler, K. Pharmacological interventions in the Wnt pathway: Inhibition of Wnt secretion versus disrupting the protein-protein interfaces of nuclear factors. Br. J. Pharm. 2017, 174, 4600–4610. [Google Scholar] [CrossRef]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H.; et al. BMP signaling in mesenchymal stem cell differentiation and bone formation. J. Biomed. Sci. Eng. 2013, 6, 32–52. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Kim, Y.-J.; Jeong, H.M.; Jin, Y.-H.; Yeo, C.-Y.; Lee, K.Y. Akt enhances Runx2 protein stability by regulating Smurf2 function during osteoblast differentiation. FEBS J. 2014, 281, 3656–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.W.; Chen, K.M.; Hassanshahi, M.; Tang, Q.; Howe, P.R.; Xian, C.J. Childhood cancer chemotherapy-induced bone damage: Pathobiology and protective effects of resveratrol and other nutraceuticals. Ann. N. Y. Acad. Sci. 2017, 1403, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, H.; Yang, S.; Qian, D. Downregulation of miR-542-3p promotes osteogenic transition of vascular smooth muscle cells in the aging rat by targeting BMP7. Hum. Genom. 2019, 13, 67. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yao, J.N.; Huang, W.T.; He, R.Q.; Ma, J.; Chen, G.; Wei, Q.J. Expression of miR-542-3p in osteosarcoma with miRNA microarray data, and its potential signaling pathways. Mol. Med. Rep. 2019, 19, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; You, J.; Li, F.; Wang, F.; Wang, Y. MicroRNA-542-3p suppresses tumor cell proliferation via targeting Smad2 inhuman osteosarcoma. Oncol. Lett. 2018, 15, 6895–6902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Kaplan, J.B.; Chae, Y.K.; Giles, F.J. Wnt/beta-catenin pathway: Modulating anticancer immune response. J. Hematol. Oncol. 2017, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Greenblatt, M.B.; Yan, G.; Feng, H.; Sun, J.; Lotinun, S.; Brady, N.; Baron, R.; Glimcher, L.H.; Zou, W. SMURF2 regulates bone homeostasis by disrupting SMAD3 interaction with vitamin D receptor in osteoblasts. Nat. Commun. 2017, 8, 14570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, B.; Cai, J.-H.; Lam, A.K.-Y.; He, B.-X. MicroRNA-542-3p inhibits oral squamous cell carcinoma progression by inhibiting ILK/TGF-β1/Smad2/3 signaling. Oncotarget 2017, 8, 70761–70776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiou, K.R.; Scherer, M.A.; Fan, C.-M.; Cool, J.C.; King, T.J.; Foster, B.K.; Xian, C.J. Methotrexate chemotherapy reduces osteogenesis but increases adipogenic potential in the bone marrow. J. Cell. Physiol. 2012, 227, 909–918. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Vander Griend, R.A.; Wronski, T.J.; Miller, G.J.; Keith, E.E.; Graves, J.E. The short- and long-term effects of methotrexate on the rat skeleton. Bone 1995, 16, 215–221. [Google Scholar] [CrossRef]
- Pelker, R.R.; Friedlaender, G.E.; Panjabi, M.M.; Markham, T.; Hausman, M.; Doganis, A.C.; McKay, J. Chemotherapy-induced alterations in the biomechanics of rat bone. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 1985, 3, 91–95. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Pillman, K.A.; Hayball, J.; Su, Y.-W.; Xian, C.J. Differentially expressed miRNAs in bone after methotrexate treatment. J. Cell. Physiol. 2021, 1, 1–18. [Google Scholar] [CrossRef]
- Zou, B.; Li, J.; Xu, K.; Liu, J.L.; Yuan, D.Y.; Meng, Z.; Zhang, B. Identification of key candidate genes and pathways in oral squamous cell carcinoma by integrated Bioinformatics analysis. Exp. Ther. Med. 2019, 17, 4089–4099. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | RT Primer | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| rSmurf2 | Random Primer (Bio-Rad) | AACACCCGGTTAAAGCACTG | AAACAAGTGTGGGCTTTTGG |
| mSmurf2 | Random Primer (Bio-Rad) | GTGAAGAGCTCGGTCCTTTG | TCGCTTGTATCTTGGCACTG |
| rsFRP-1 | Random Primer (Bio-Rad) | CCCGAGATGCTCAAATGTGAC | AGATGTTCGATGATGGCCTCC |
| msFRP-1 | Random Primer (Bio-Rad) | TGCGAGCCGGTCATGCAGTT | ACACGGTTGTACCTTGGGGCT |
| mSmad3 | Random Primer (Bio-Rad) | CTGGGCCTACTGTCCAATGT | GCAGCAAATTCCTGGTTGTT |
| mRUNX2 | Random Primer (Bio-Rad) | CCCAGCCACCTTTACCTACA | TATGGAGTGCTGCTGGTCTG |
| mALP | Random Primer (Bio-Rad) | GCTGATCATTCCCACGTTTT | CTGGGCCTGGTAGTTGTTGT |
| mOSX | Random Primer (Bio-Rad) | ACTCATCCCTATGGCTCGTG | GGTAGGGAGCTGGGTTAAGG |
| mOCN | Random Primer (Bio-Rad) | AAGCAGGAGGGCAATAAGGT | TTTGTAGGCGGTCTTCAAGC |
| mC/EBPα | Random Primer (Bio-Rad) | TGGACAAGAACAGCAACGAG | CCTTGACCAAGGAGCTCTCA |
| mPPARγ | Random Primer (Bio-Rad) | TTTTCAAGGGTGCCAGTTTC | AATCCTTGGCCCTCTGAGAT |
| mLEF1 | Random Primer (Bio-Rad) | TATGAACAGCGACCCGTACA | ACATCTGACGGGATGTGTGA |
| mTCF7 | Random Primer (Bio-Rad) | GCCAGAAGCAAGGAGTTCAC | TACACCAGATCCCAGCATCA |
| mDLX5 | Random Primer (Bio-Rad) | CCACCAGCCAGCCAGAGAAA | GGGGCATCTCCCCGTTTTT |
| rmCycA | Random Primer (Bio-Rad) | CGTTGGATGGCAAGCATGTG | TGCTGGTCTTGCCATTCCTG |
| miR-542-3p | GAAAGAAGGCGAGGAGCAGATCGAGGAAGAAGACGGAAGAATGTGCGTCTCGCCTTCTTTCTTCAGTTA | TGTGACAGATTGATAACTGA | GAGGAAGAAGACGGAAGAAT |
| U6 | CGCTTCACGAATTTGCGTG | GCTTCGGCAGCACATATAC | CGCTTCACGAATTTGCGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-L.; Liu, L.; Su, Y.-W.; Xian, C.J. miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2. Int. J. Mol. Sci. 2021, 22, 10988. https://doi.org/10.3390/ijms222010988
Zhang Y-L, Liu L, Su Y-W, Xian CJ. miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2. International Journal of Molecular Sciences. 2021; 22(20):10988. https://doi.org/10.3390/ijms222010988
Chicago/Turabian StyleZhang, Ya-Li, Liang Liu, Yu-Wen Su, and Cory J. Xian. 2021. "miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2" International Journal of Molecular Sciences 22, no. 20: 10988. https://doi.org/10.3390/ijms222010988
APA StyleZhang, Y. -L., Liu, L., Su, Y. -W., & Xian, C. J. (2021). miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2. International Journal of Molecular Sciences, 22(20), 10988. https://doi.org/10.3390/ijms222010988