
IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach

 , ,
, ,  , , and
, , and 
Abstract
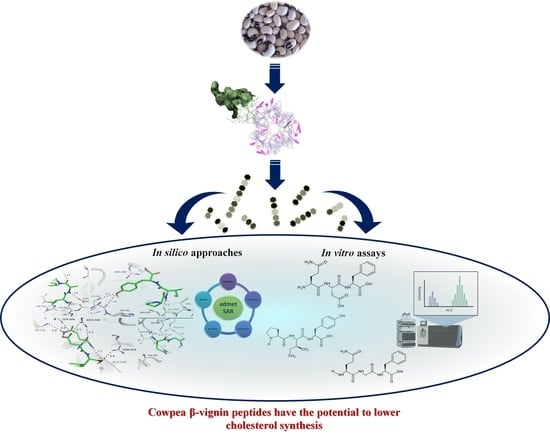
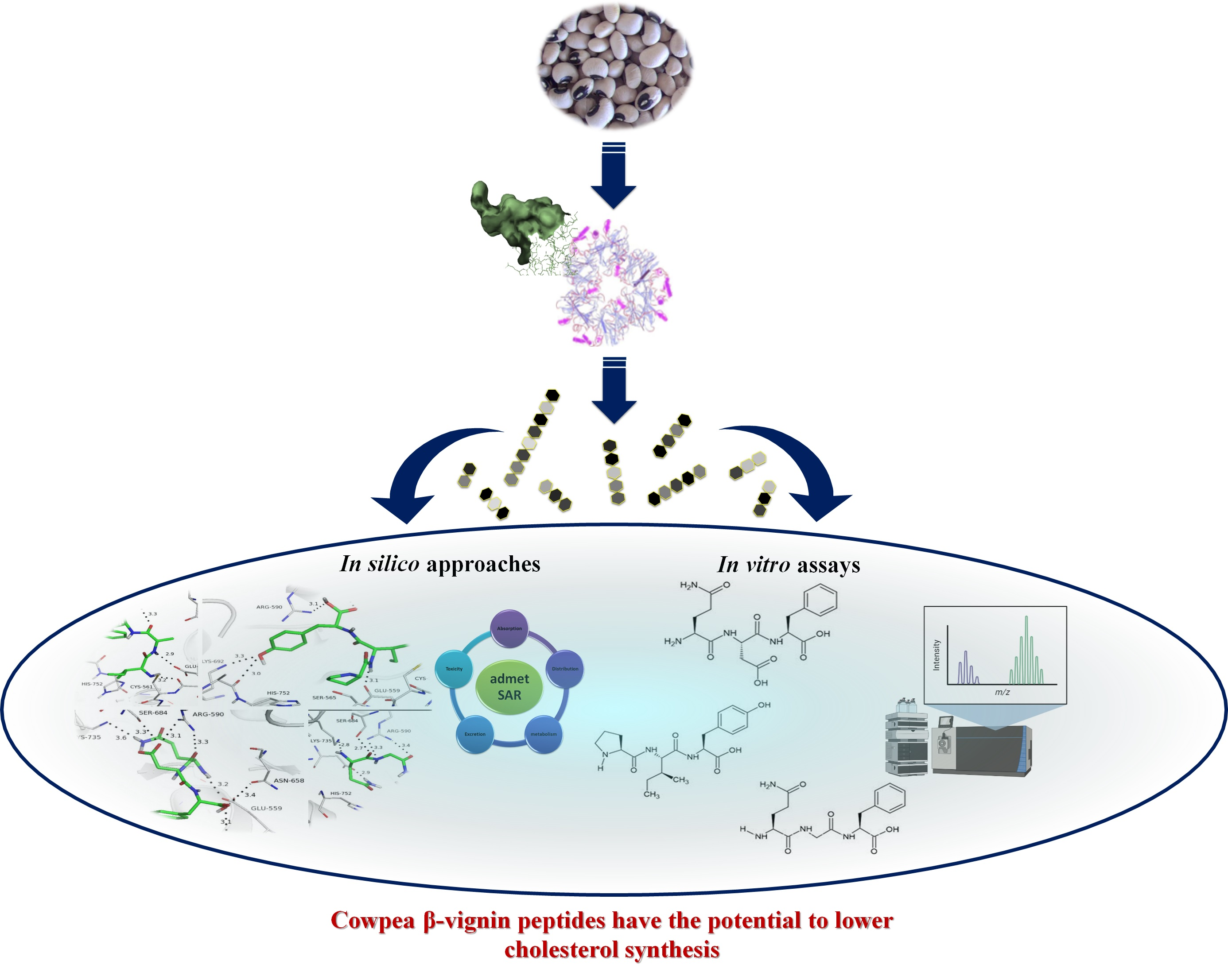
:
1. Introduction
2. Results and Discussion
2.1. Peptide-Binding Sites within HMG-CoA Reductase In Silico
2.2. ADMET Parameters
2.3. HMG-CoA Reductase Enzyme Inhibition In Vitro
3. Materials and Methods
3.1. In-Silico Studies
3.1.1. Hydrolysis of the Cowpea β-Vignin Primary Sequence
3.1.2. Molecular Features and Bioactivity Prediction
3.1.3. Prediction of Putative Peptide Binding Sites in HMG-CoA Reductase
3.1.4. Molecular Docking
3.1.5. Absorption, Distribution, Metabolism, Excretion, and Toxicity (ADMET) Parameters
3.2. In Vitro Experiments
3.2.1. Preparation of Peptide Fractions with Varying Molecular Sizes from Cowpea β-Vignin
3.2.2. Profiling of Peptide Fractions by RP-HPLC
3.2.3. Peptide Synthesis
3.2.4. HMG-CoA Reductase Enzyme Assay
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HMG-CoAR | 3-hydroxy-3-methylglutaryl-coenzyme A reductase enzyme |
| HMG-CoA | 3-hydroxy-3-methylglutaryl-coenzyme A Substrate |
| SRAEs | Statin-related adverse events |
| CK | Creatine kinase |
| A8YQH5_VIGUN | UniProtKB code of the primary sequence of cowpea β-vignin protein |
| PRS | PeptideRanker Score |
| PDB | Protein data bank |
| 1HW9 | PDB code of HMGCoA reductase in complex with simvastatin |
| 1HWL | PDB code of HMGCoA reductase in complex with atorvastatin |
| RMSD | Root mean square deviation |
| ADMET | Absorption, distribution, metabolism, excretion, and toxicity |
| PK | Pharmacokinetic |
| Fmoc | 9-fluorenylmethyloxycarbonyl |
| THE | Total hydrolyzed extract |
| SREBP2 | Sterol regulatory element-binding protein 2 |
| HepG2 | Human liver cancer cell line |
| LDLR | Low-density lipoprotein receptor |
| PepT1 | Peptide transporter 1 |
| OATP | Organic anion transporting polypeptides |
| CYP450 | Family of enzymes involved in the formation (synthesis) and breakdown (metabolism) |
References
- Mensah, G.A.; Roth, G.A.; Fuster, V. The global burden of cardiovascular diseases and risk factors: 2020 and beyond. J. Am. Coll. Cardiol. 2019, 74, 2529–2532. [Google Scholar] [CrossRef] [PubMed]
- Budreviciute, A.; Damiati, S.; Sabir, D.K.; Onder, K.; Schuller-Goetzburg, P.; Plakys, G.; Katileviciute, A.; Khoja, S.; Kodzius, R. Management and prevention strategies for non-communicable diseases (NCDs) and their risk factors. Front. Public Health 2020, 8, 788. [Google Scholar] [CrossRef]
- GBD 2015 Risk Factors Collaborators. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1659–1724. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.; Yi, J.-J.; Ohrr, H.; Yi, S.-W. Total cholesterol and mortality from ischemic heart disease and overall cardiovascular disease in Korean adults. Medicine 2019, 98, e17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istvan, E.S. Structural mechanism for statin inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase. Am. Heart J. 2002, 144, S27–S32. [Google Scholar] [CrossRef]
- Istvan, E.S.; Palnitkar, M.; Buchanan, S.K.; Deisenhofer, J. Crystal structure of the catalytic portion of human HMG-CoA reductase: Insights into regulation of activity and catalysis. EMBO J. 2000, 19, 819–830. [Google Scholar] [CrossRef]
- Istvan, E.S.; Deisenhofer, J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science 2001, 292, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.-Y.; Li, H.; Tang, J.-J.; Wang, J.; Luo, J.; Liu, B.; Wang, J.-K.; Shi, X.-J.; Cui, H.-W.; Tang, J.; et al. Discovery of a potent HMG-CoA reductase degrader that eliminates statin-induced reductase accumulation and lowers cholesterol. Nat. Commun. 2018, 9, 5138. [Google Scholar] [CrossRef] [Green Version]
- Golomb, B.A.; Evans, M.A. Statin adverse effects: A review of the literature and evidence for a mitochondrial mechanism. Am. J. Cardiovasc. Drugs 2008, 8, 373–418. [Google Scholar] [CrossRef]
- Du Souich, P.; Roederer, G.; Dufour, R. Myotoxicity of statins: Mechanism of action. Pharmacol. Ther. 2017, 175, 1–16. [Google Scholar] [CrossRef]
- Collins, R.; Reith, C.; Emberson, J.; Armitage, J.; Baigent, C.; Blackwell, L.; Blumenthal, R.; Danesh, J.; Smith, G.D.; DeMets, D.; et al. Interpretation of the evidence for the efficacy and safety of statin therapy. Lancet 2016, 388, 2532–2561. [Google Scholar] [CrossRef] [Green Version]
- Bellosta, S.; Corsini, A. Statin drug interactions and related adverse reactions: An update. Expert Opin. Drug Saf. 2018, 17, 25–37. [Google Scholar] [CrossRef]
- Ferreira, E.S.; Amaral, A.L.S.; Demonte, A.; Zanelli, C.F.; Capraro, J.; Duranti, M.; Neves, V.A. Hypocholesterolaemic effect of rat-administered oral doses of the isolated 7S globulins from cowpeas and adzuki beans. J. Nutr. Sci. 2015, 4, e7. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, E.S.; Silva, M.A.; Demonte, A.; Neves, V.A. β-conglycinin combined with fenofibrate or rosuvastatin have exerted distinct hypocholesterolemic effects in rats. Lipids Health Dis. 2012, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lammi, C.; Zanoni, C.; Arnoldi, A. Three peptides from soy glycinin modulate glucose metabolism in human hepatic HepG2 cells. Int. J. Mol. Sci. 2015, 16, 27362–27370. [Google Scholar] [CrossRef] [Green Version]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Two peptides from soy β-conglycinin induce a hypocholesterolemic effect in HepG2 cells by a statin-like mechanism: Comparative in vitro and in silico modeling studies. J. Agric. Food Chem. 2015, 63, 7945–7951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pak, V.V.; Koo, M.; Lee, N.; Oh, S.K.; Kim, M.S.; Lee, J.-S.; Kwon, D.Y. Hypocholesterolemic soybean peptide (IAVP) inhibits HMG-CoA reductase in a competitive manner. Food Sci. Biotechnol. 2005, 14, 727–731. [Google Scholar]
- Shiuan, D.; Lin, S.-H.; Hwang, K.-J.; Weng, C.-F. Exploration of natural product ingredients as inhibitors of human HMG-CoA reductase through structure-based virtual screening. Drug Des. Devel. Ther. 2015, 9, 3313. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sharma, P.; Bairagya, H.R.; Sharma, S.; Singh, T.P.; Tiku, P.K. Inhibition of human 3-hydroxy-3-methylglutaryl CoA reductase by peptides leading to cholesterol homeostasis through SREBP2 pathway in HepG2 cells. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Pak, V.V.; Koo, M.; Kwon, D.Y.; Yun, L. Design of a highly potent inhibitory peptide acting as a competitive inhibitor of HMG-CoA reductase. Amino Acids 2012, 43, 2015–2025. [Google Scholar] [CrossRef]
- Silva, M.B.d.C.e.; Souza, C.A.daC.; Philadelpho, B.O.; Cunha, M.M.N.d.; Batista, F.P.R.; Silva, J.R.d.; Druzian, J.I.; Castilho, M.S.; Cilli, E.M.; Ferreira, E.S. In vitro and in silico studies of 3-hydroxy-3-methyl-glutaryl coenzyme A reductase inhibitory activity of the cowpea Gln-Asp-Phe peptide. Food Chem. 2018, 259, 270–277. [Google Scholar] [CrossRef]
- Arnoldi, A.; Boschin, G.; Zanoni, C.; Lammi, C. The health benefits of sweet lupin seed flours and isolated proteins. J. Funct. Foods 2015, 18, 550–563. [Google Scholar] [CrossRef]
- Pak, V.V.; Koo, M.; Kim, M.J.; Yang, H.J.; Yun, L.; Kwon, D.Y. Modeling an active conformation for linear peptides and design of a competitive inhibitor for HMG-CoA reductase. J. Mol. Recognit. 2008, 21, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.R.; Soares Freitas, R.A.M.; Corrêa Carlos, A.C.; Siguemoto, É.S.; Fontanari, G.G.; Arêas, J.A.G. Peptides from cowpea present antioxidant activity, inhibit cholesterol synthesis and its solubilisation into micelles. Food Chem. 2015, 168, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H. Molecular and integrative physiology of intestinal peptide transport. Annu. Rev. Physiol. 2004, 66, 361–384. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A. IAVPGEVA, IAVPTGVA, and LPYP, three peptides from soy glycinin, modulate cholesterol metabolism in HepG2 cells through the activation of the LDLR-SREBP2 pathway. J. Funct. Foods 2015, 14, 469–478. [Google Scholar] [CrossRef]
- Freeman, H.J. Clinical relevance of intestinal peptide uptake. World J. Gastrointest. Pharmacol. Ther. 2015, 6, 22–27. [Google Scholar] [CrossRef]
- Marczak, M.; Okoniewska, K.; Grabowski, T. Classification model of amino acid sequences prone to aggregation of therapeutic proteins. In Silico Pharmacol. 2016, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Ghersi, D.; Sanchez, R. Improving accuracy and efficiency of blind protein-ligand docking by focusing on predicted binding sites. Proteins 2009, 74, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetényi, C.; van der Spoel, D. Efficient docking of peptides to proteins without prior knowledge of the binding site. Protein Sci. 2009, 11, 1729–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, N.M.; Alhossary, A.A.; Mu, Y.; Kwoh, C.K. Protein-ligand blind docking using quickvina-w with inter-process spatio-temporal integration. Sci. Rep. 2017, 7, 15451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.N. Morphological similarity: A 3D molecular similarity method correlated with protein-ligand recognition. J. Comput. Aided Mol. Des. 2000, 14, 199–213. [Google Scholar] [CrossRef]
- Pagadala, N.S.; Syed, K.; Tuszynski, J. Software for molecular docking: A review. Biophys. Rev. 2017, 9, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Toldrá, F.; Nasri, M. In silico analysis and molecular docking study of angiotensin I-converting enzyme inhibitory peptides from smooth-hound viscera protein hydrolysates fractionated by ultrafiltration. Food Chem. 2018, 239, 453–463. [Google Scholar] [CrossRef]
- Ferreira, L.L.G.; Andricopulo, A.D. ADMET modeling approaches in drug discovery. Drug Discov. Today 2019, 24, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Kazi, D.S.; Penko, J.M.; Bibbins-Domingo, K. Statins for primary prevention of cardiovascular disease: Review of evidence and recommendations for clinical practice. Med. Clin. N. Am. 2017, 101, 689–699. [Google Scholar] [CrossRef]
- Spanier, B.; Rohm, F. Proton coupled oligopeptide transporter 1 (PepT1) function, regulation, and influence on the intestinal homeostasis. Compr. Physiol. 2018, 8, 843–869. [Google Scholar] [CrossRef]
- Maeda, K. Organic anion transporting polypeptide (OATP)1B1 and OATP1B3 as important regulators of the pharmacokinetics of substrate drugs. Biol. Pharm. Bull. 2015, 38, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, S.; Mazzanti, R.; Beijnen, J.H.; Schellens, J.H.M. Concise review: Clinical relevance of drug drug and herb drug interactions mediated by the ABC transporter ABCB1 (MDR1, P-glycoprotein). Oncologist 2007, 12, 927–941. [Google Scholar] [CrossRef] [Green Version]
- Matheny, C.J.; Lamb, M.W.; Brouwer, K.R.; Pollack, G.M. Pharmacokinetic and pharmacodynamic implications of P-glycoprotein modulation. Pharmacotherapy 2001, 21, 778–796. [Google Scholar] [CrossRef]
- Rowland Yeo, K.; Rostami-Hodjegan, A.; Tucker, G.T. Abundance of cytochromes P450 in human liver: A meta-analysis. Br. J. Clin. Pharmacol. 2004, 57, 687–688. [Google Scholar]
- Björnsson, E.; Jacobsen, E.I.; Kalaitzakis, E. Hepatotoxicity associated with statins: Reports of idiosyncratic liver injury post-marketing. J. Hepatol. 2012, 56, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Archetti, F.; Lanzeni, S.; Messina, E.; Vanneschi, L. Genetic programming and other machine learning approaches to predict median oral lethal dose (LD50) and plasma protein binding levels (%PPB) of drugs. In Proceedings of the 5th European Conference on Evolutionary Computation, Machine Learning and Data Mining in Bioinformatics (EvoBIO’07), Valencia, Spain, 11–13 April 2007; Marchiori, E., Moore, J.H., Rajapakse, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 11–23. [Google Scholar]
- Ferreira, E.D.S.; Capraro, J.; Sessa, F.; Magni, C.; Demonte, A.; Consonni, A.; Neves, V.A.; Cilli, E.M.; Duranti, M.; Scarafoni, A. New molecular features of cowpea bean (Vigna unguiculata, L. Walp) β-vignin. Biosci. Biotechnol. Biochem. 2018, 82, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amigo-Benavent, M.; Clemente, A.; Caira, S.; Stiuso, P.; Ferranti, P.; del Castillo, M.D. Use of phytochemomics to evaluate the bioavailability and bioactivity of antioxidant peptides of soybean β-conglycinin. Electrophoresis 2014, 35, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Fassini, P.G.; de Souza Ferreira, E.; da Silva, M.A.; Neves, V.A.; Demonte, A. Soybean glycinin (11S) increases HDL-cholesterol in hypercholesterolemic rats. Nutr. Food Sci. 2012, 42, 102–110. [Google Scholar] [CrossRef]
- Amaral, A.L.; Ferreira, E.S.; Silva, M.A.; Neves, V.A.; Demonte, A. The Vicilin protein (Vigna radiata L.) of mung bean as a functional food: Evidence of “in vitro” hypocholesterolemic activity. Nutr. Food Sci. 2017, 47, 907–916. [Google Scholar] [CrossRef] [Green Version]
- De Ferreira, E.S.; Silva, M.A.; Demonte, A.; Neves, V.A. β-Conglycinin (7S) and glycinin (11S) exert a hypocholesterolemic effect comparable to that of fenofibrate in rats fed a high-cholesterol diet. J. Funct. Foods 2010, 2, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Akeson, W.R.; Stahmann, M.A. A pepsin pancreatin digest index of protein quality evaluation. J. Nutr. 1964, 83, 257–261. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Merrifield, R.B. Solid phase peptide synthesis. I. The synthesis of a tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Sequence a | N° aa a | Localisation a | MW b | Isoelectric Point b | Hydrophobicity (Kcal/mol) b | Net Charge c | Ring c | PRS d |
|---|---|---|---|---|---|---|---|---|---|
| 1 | VPL | 3 | 1–3 | 327.42 | 5.49 | 2.1333 | 0 | 1 | 0.37 |
| 2 | GVL | 3 | 7–9 | 287.36 | 5.52 | 2.5333 | 0 | 0 | 0.31 |
| 3 | ASL | 3 | 12–14 | 289.33 | 5.57 | 1.6000 | 0 | 0 | 0.25 |
| 4 | SVSF | 4 | 15–18 | 438.48 | 5.24 | 1.3500 | 0 | 1 | 0.41 |
| 5 | GIVHR | 5 | 19–23 | 580.69 | 9.76 | 0.1200 | +1 | 1 | 0.20 |
| 6 | GHQESQEESEPR | 12 | 24–35 | 1412.39 | 4.48 | −2.6916 | −3 | 2 | 0.09 |
| 7 | GQNNPF | 6 | 36–41 | 675.70 | 5.52 | −1.6166 | 0 | 2 | 0.80 |
| 8 | DSDR | 4 | 44–47 | 491.46 | 4.21 | −3.0749 | −1 | 0 | 0.20 |
| 9 | HTL | 3 | 50–52 | 369.42 | 6.74 | −0.0332 | 0 | 1 | 0.14 |
| 10 | NQY | 3 | 55–57 | 423.43 | 5.52 | −2.7666 | 0 | 1 | 0.16 |
| 11 | GHL | 3 | 58–60 | 325.37 | 6.74 | 0.0667 | 0 | 1 | 0.59 |
| 12 | VL | 2 | 62–63; 202–203 | 230.31 | 5.49 | 4.0000 | 0 | 0 | 0.13 |
| 13 | QR | 2 | 64–65 | 302.33 | 9.75 | −3.9999 | +1 | 0 | 0.27 |
| 14 | DQR | 3 | 67–69 | 417.42 | 5.84 | −3.8332 | 0 | 0 | 0.19 |
| 15 | SK | 2 | 70–71; 184–185 | 233.27 | 8.47 | −2.3499 | +1 | 0 | 0.07 |
| 16 | QIQNL | 5 | 72–76 | 614.70 | 5.52 | −0.4399 | 0 | 0 | 0.23 |
| 17 | ENY | 3 | 77–79 | 424.41 | 4.00 | −2.7666 | −1 | 1 | 0.07 |
| 18 | VVEF | 4 | 81–84 | 492.57 | 4.00 | 1.9250 | −1 | 1 | 0.13 |
| 19 | QSK | 3 | 85–87 | 361.40 | 8.75 | −2.7332 | +1 | 0 | 0.07 |
| 20 | PNTL | 4 | 88–91 | 443.50 | 5.96 | −0.4999 | 0 | 1 | 0.32 |
| 21 | PHHADADF | 8 | 94–101 | 908.93 | 5.05 | −1.0749 | −2 | 4 | 0.51 |
| 22 | VVL | 3 | 104–106 | 329.44 | 5.49 | 4.0667 | 0 | 0 | 0.06 |
| 23 | NGR | 3 | 107–109 | 345.36 | 9.75 | −2.7999 | +1 | 0 | 0.46 |
| 24 | AIL | 3 | 110–112 | 315.41 | 5.57 | 3.3667 | 0 | 0 | 0.33 |
| 25 | TL | 2 | 113–114 | 232.28 | 5.19 | 1.5500 | 0 | 0 | 0.14 |
| 26 | VNPDGR | 6 | 115–120 | 656.70 | 5.81 | −1.5499 | 0 | 1 | 0.39 |
| 27 | DSY | 3 | 121–123 | 383.36 | 3.80 | −1.8666 | −1 | 1 | 0.18 |
| 28 | IL | 2 | 124–125 | 244.33 | 5.52 | 4.1500 | 0 | 0 | 0.39 |
| 29 | EQGHAQK | 6 | 126–132 | 796.84 | 6.85 | −2.3142 | 0 | 1 | 0.08 |
| 30 | TPAGTTF | 7 | 133–139 | 693.75 | 5.19 | 0.0714 | 0 | 2 | 0.33 |
| 31 | VNHDDNENL | 9 | 142–150 | 1069.05 | 4.02 | −1.7999 | −3 | 1 | 0.15 |
| 32 | IVK | 3 | 152–154 | 358.48 | 8.75 | 1.6000 | +1 | 0 | 0.05 |
| 33 | AVPVNNPHR | 9 | 156–164 | 1003.13 | 9.80 | −0.8555 | +1 | 3 | 0.47 |
| 34 | QDF | 3 | 166–168 | 408.41 | 3.80 | −1.3999 | −1 | 1 | 0.74 |
| 35 | SSTEAQQSY | 9 | 171–179 | 999.99 | 4.00 | −1.4555 | −1 | 1 | 0.08 |
| 36 | QGF | 3 | 181–183 | 350.37 | 5.52 | −0.3666 | 0 | 1 | 0.94 |
| 37 | NIL | 3 | 186–188 | 358.44 | 5.52 | 1.6000 | 0 | 0 | 0.26 |
| 38 | EASF | 4 | 189–192 | 452.46 | 4.00 | 0.0750 | −1 | 1 | 0.30 |
| 39 | DSDF | 4 | 193–196 | 482.45 | 3.56 | −1.2499 | −2 | 1 | 0.60 |
| 40 | EINR | 4 | 198–201 | 530.58 | 6.10 | −1.7499 | 0 | 0 | 0.08 |
| 41 | GEEEQK | 6 | 205–210 | 718.72 | 4.25 | −3.0499 | −2 | 0 | 0.04 |
| 42 | QQDEESQQEGVIVQL | 12 | 211–225 | 1729.82 | 3.50 | −1.0666 | −4 | 0 | 0.11 |
| 43 | EQIR | 4 | 228–231 | 544.61 | 6.10 | −1.7499 | 0 | 0 | 0.09 |
| 44 | EL | 2 | 232–233 | 260.29 | 4.00 | 0.1500 | −1 | 0 | 0.07 |
| 45 | MK | 2 | 234–235 | 277.38 | 8.50 | −0.9999 | +1 | 0 | 0.45 |
| 46 | HAK | 3 | 236–238 | 354.41 | 8.76 | −1.7666 | +1 | 1 | 0.11 |
| 47 | STSK | 4 | 239–242 | 421.45 | 8.47 | −1.5499 | +1 | 0 | 0.06 |
| 48 | SL | 2 | 244–245 | 218.25 | 5.24 | 1.5000 | 0 | 0 | 0.33 |
| 49 | STQNEPF | 7 | 246–252 | 821.84 | 4.00 | −1.5428 | −1 | 2 | 0.35 |
| 50 | NL | 2 | 253–254 | 245.28 | 5.52 | 0.1500 | 0 | 0 | 0.29 |
| 51 | SQK | 3 | 256–258 | 361.40 | 8.47 | −2.7332 | +1 | 0 | 0.08 |
| 52 | PIY | 3 | 259–261 | 391.47 | 5.95 | 0.5333 | 0 | 2 | 0.60 |
| 53 | SNK | 3 | 262–264 | 347.37 | 8.47 | −2.7332 | +1 | 0 | 0.09 |
| 54 | GR | 2 | 266–267 | 231.25 | 9.75 | −2.4499 | +1 | 0 | 0.77 |
| 55 | HEITPEK | 7 | 269–275 | 852.94 | 5.40 | −1.6999 | −1 | 2 | 0.09 |
| 56 | NPQL | 4 | 276–279 | 470.53 | 5.52 | −1.1999 | 0 | 1 | 0.47 |
| 57 | DL | 2 | 281–282 | 246.26 | 3.80 | 0.1500 | −1 | 0 | 0.33 |
| 58 | DVF | 3 | 283–285 | 379.41 | 3.80 | 1.1667 | −1 | 1 | 0.55 |
| 59 | TSVDIK | 6 | 287–292 | 661.75 | 5.50 | −0.0332 | 0 | 0 | 0.08 |
| 60 | EGGL | 4 | 293–296 | 374.39 | 4.00 | −0.1249 | −1 | 0 | 0.37 |
| 61 | MPNY | 4 | 298–301 | 523.60 | 5.27 | −1.1249 | 0 | 2 | 0.74 |
| 62 | NSK | 3 | 302–304 | 347.37 | 8.75 | −2.7332 | +1 | 0 | 0.07 |
| 63 | AIVIL | 5 | 305–309 | 527.70 | 5.57 | 3.7600 | 0 | 0 | 0.24 |
| 64 | VVNK | 4 | 310–313 | 458.56 | 8.72 | 0.2500 | +1 | 0 | 0.03 |
| 65 | GEANIEL | 7 | 314–320 | 744.80 | 3.79 | −0.1142 | −2 | 0 | 0.14 |
| 66 | VGQR | 4 | 321–324 | 458.52 | 9.72 | −1.0499 | +1 | 0 | 0.17 |
| 67 | EQQQQQQEESW | 11 | 325–335 | 1447.44 | 3.67 | −3.0181 | −3 | 2 | 0.09 |
| 68 | EVQR | 4 | 336–339 | 530.58 | 6.10 | −1.8249 | 0 | 0 | 0.04 |
| 69 | AEVSDDDVF | 9 | 342–350 | 996.00 | 3.37 | −0.1999 | −4 | 1 | 0.19 |
| 70 | VIPASY | 6 | 351–356 | 648.76 | 5.49 | 1.1333 | 0 | 2 | 0.17 |
| 71 | PVAITATSNL | 10 | 357–366 | 986.13 | 5.96 | 0.8800 | 0 | 1 | 0.14 |
| 72 | NF | 2 | 367–368;381–382 | 279.30 | 5.52 | −0.3499 | 0 | 1 | 0.94 |
| 73 | IAF | 3 | 369–371 | 349.43 | 5.52 | 3.0333 | 0 | 1 | 0.82 |
| 74 | GINAENNQR | 9 | 372–380 | 1015.05 | 6.00 | −1.7888 | 0 | 0 | 0.12 |
| 75 | AGEEDNVMSEIPTEVL | 16 | 384–399 | 1732.88 | 3.45 | −0.2562 | −5 | 1 | 0.16 |
| 76 | DVTF | 4 | 400–403 | 480.52 | 3.80 | 0.7000 | −1 | 1 | 0.32 |
| 77 | PASGEK | 6 | 404–409 | 587.63 | 6.43 | −1.3999 | 0 | 1 | 0.17 |
| 78 | VEK | 3 | 410–412 | 374.44 | 5.97 | −1.0666 | 0 | 0 | 0.02 |
| 79 | INK | 3 | 414–416 | 373.45 | 8.75 | −0.9666 | +1 | 0 | 0.10 |
| 80 | QSDSHF | 6 | 417–422 | 719.71 | 5.08 | −1.4999 | −1 | 2 | 0.52 |
| 81 | TDHSSK | 6 | 423–428 | 673.68 | 6.41 | −2.1499 | 0 | 1 | 0.06 |
| 82 | EER | 3 | 430–432 | 432.43 | 4.53 | −3.8332 | −1 | 0 | 0.04 |
| N° | Sequence a | PRS b | Lipophilicity (LogP) | SURFLEX-SIM c | |||
|---|---|---|---|---|---|---|---|
| NADH | SIM | ATO | ROS | ||||
| 1 | QGF | 0.94 | −0.478 | 4.28 | 6.18 * | 5.39 * | 5.42 * |
| 2 | NF | 0.94 | 0.094 | 3.33 | 5.42 | 4.44 | 4.24 |
| 3 | IAF | 0.82 | −0.555 | 3.83 | 6.01 | 5.57 * | 4.91 * |
| 4 | GQNNPF | 0.80 | 0.533 | 3.52 | 4.28 | 4.70 | 4.06 |
| 5 | GR | 0.77 | 0.689 | 3.53 | 5.37 | 3.95 | 4.41 |
| 6 | QDF | 0.74 | −1.089 | 4.30 | 6.09 * | 5.61 * | 4.36 |
| 7 | MPNY | 0.74 | 1.897 | 3.97 | 4.86 | 4.32 | 4.29 |
| 8 | PIY | 0.60 | −1.859 | 4.05 | 6.23 * | 4.95 | 5.08 * |
| 9 | DSDF | 0.60 | 1.223 | 3.96 | 5.27 | 5.27 | 5.07 * |
| 10 | GHL | 0.59 | 1.056 | 4.58 | 5.75 | 4.81 | 4.50 |
| 11 | DVF | 0.55 | 0.793 | 3.58 | 6.09 * | 5.52 * | 4.67 |
| 12 | QSDSHF | 0.52 | 2.159 | 3.45 | 4.15 | 4.70 | 3.63 |
| 13 | PHHADADF | 0.51 | −1.990 | 3.64 | 3.00 | 2.94 | 2.58 |
| ADMET Parameters | IAF | QDF | SIM | ATOR |
|---|---|---|---|---|
| Absorption | Probability | Probability | Probability | Probability |
| Human oral bioavailability | 0.5429 | 0.5857 | 0.9571 | 0.9196 |
| Caco-2 | 0.8014 | 0.9394 | 0.5503 | 0.8264 |
| Human Intestinal Absorption | 0.7591 | 0.5081 | 0.9767 | 0.9669 |
| OATP2B1 inhibitor | 1.0000 | 1.0000 | 1.0000 | 0.6023 |
| OATP1B1 inhibitor | 0.9011 | 0.9287 | 0.7740 | 0.8473 |
| OATP1B3 inhibitor | 0.9466 | 0.9505 | 0.9480 | 0.9237 |
| Distribution | Probability | Probability | Probability | Probability |
| P-glycoprotein inhibitor | 0.7977 | 0.6699 | 0.9198 | 0.7590 |
| P-glycoprotein substrate | 0.5570 | 0.7014 | 0.9344 | 0.6434 |
| Blood–Brain Barrier (BBB) | 0.9164 | 0.9524 | 0.9822 | 0.5857 |
| Mitochondrial subcellular distribution | 0.5377 | 0.6796 | 0.7384 | 0.7153 |
| Metabolism | Probability | Probability | Probability | Probability |
| CYP3A4 substrate | 0.5726 | 0.5225 | 0.7410 | 0.6733 |
| CYP2C9 substrate | 0.5943 | 0.5648 | 1.0000 | 0.7914 |
| CYP2D6 substrate | 0.8036 | 0.7860 | 0.8893 | 0.7542 |
| CYP inhibitory activity | 0.9732 | 0.9845 | 0.8682 | 0.7663 |
| Toxicity | Probability | Probability | Probability | Probability |
| Carcinogenicity (binary) | 0.6571 | 0.7143 | 0.9286 | 0.8143 |
| Carcinogenicity (trinary) | 0.6963 | 0.7645 | 0.7060 | 0.4690 |
| Hepatotoxicity | 0.5750 | 0.6500 | 0.9000 | 0.6500 |
| ADMET predicted profile Regressions | Unit | Unit | Unit | Unit |
| Water solubility (logS) | −2.319 | −1.964 | −5.483 | −3.813 |
| Plasma protein binding (%) | 0.770 | 0.595 | 0.918 | 0.878 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.; Philadelpho, B.; Santos, J.; Souza, V.; Souza, C.; Santiago, V.; Silva, J.; Souza, C.; Azeredo, F.; Castilho, M.; et al. IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach. Int. J. Mol. Sci. 2021, 22, 11067. https://doi.org/10.3390/ijms222011067
Silva M, Philadelpho B, Santos J, Souza V, Souza C, Santiago V, Silva J, Souza C, Azeredo F, Castilho M, et al. IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach. International Journal of Molecular Sciences. 2021; 22(20):11067. https://doi.org/10.3390/ijms222011067
Chicago/Turabian StyleSilva, Mariana, Biane Philadelpho, Johnnie Santos, Victória Souza, Caio Souza, Victória Santiago, Jaff Silva, Carolina Souza, Francine Azeredo, Marcelo Castilho, and et al. 2021. "IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach" International Journal of Molecular Sciences 22, no. 20: 11067. https://doi.org/10.3390/ijms222011067
APA StyleSilva, M., Philadelpho, B., Santos, J., Souza, V., Souza, C., Santiago, V., Silva, J., Souza, C., Azeredo, F., Castilho, M., Cilli, E., & Ferreira, E. (2021). IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach. International Journal of Molecular Sciences, 22(20), 11067. https://doi.org/10.3390/ijms222011067