
Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis B Virus Therapy: A Narrative Review

 ,
,
Abstract
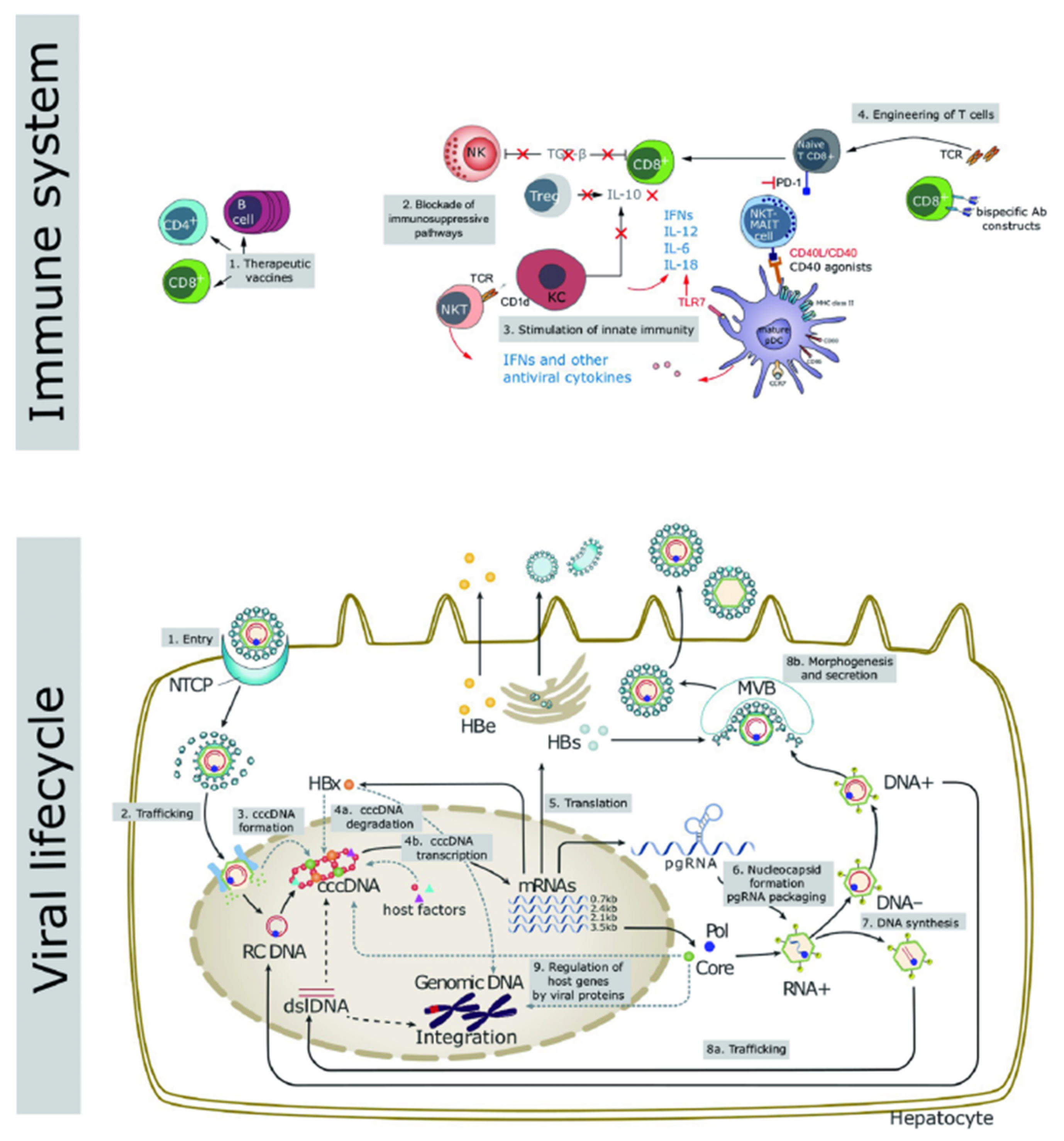
:1. Introduction
2. Methods
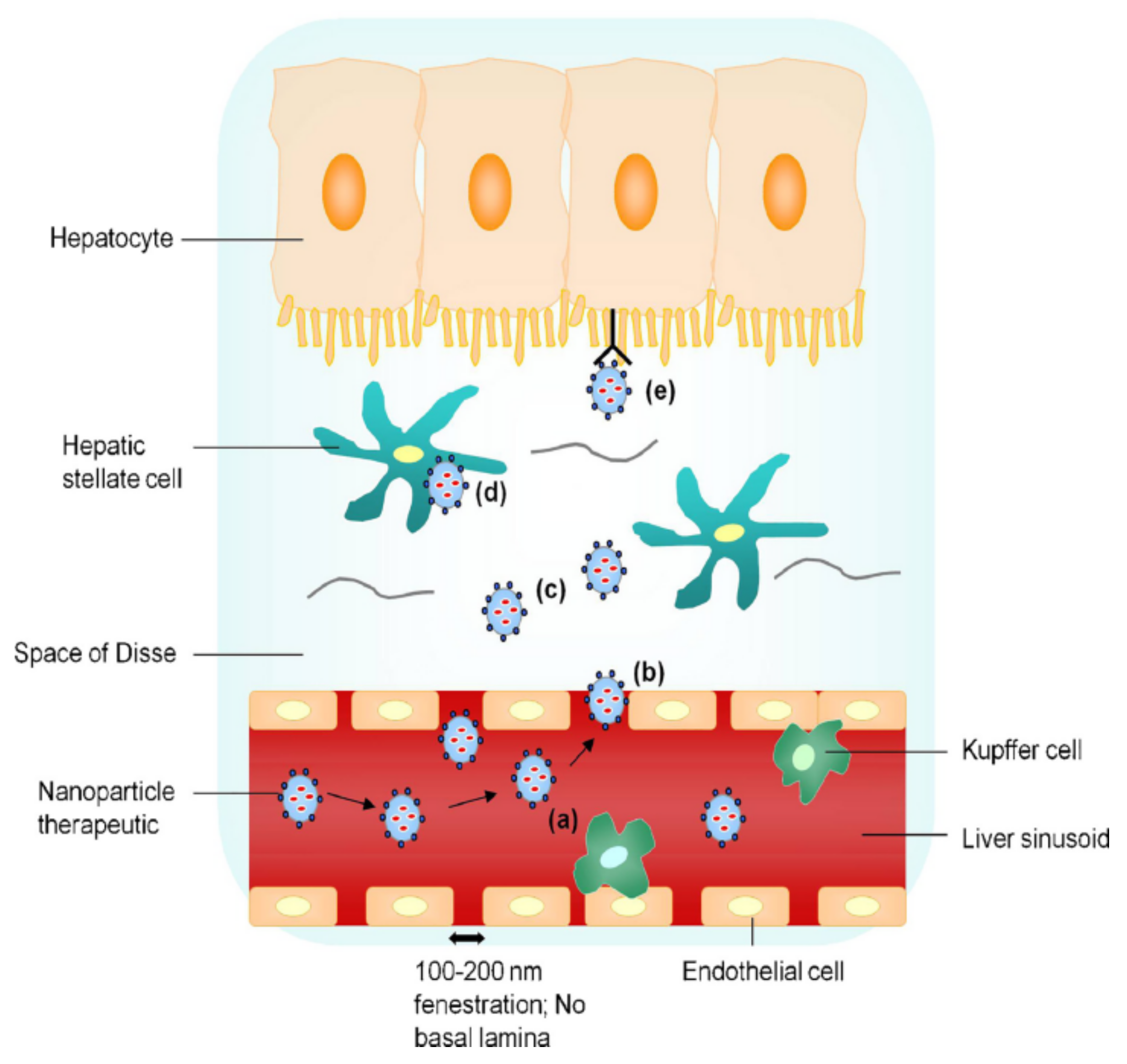
3. Characteristics of Nanoparticle Drug Delivery Systems for Anti-HBV Therapy
- (a)
- Controlling the particle size below 200 nm, which reduces phagocytosis by nonparenchymal cells such as Kupffer cells, and further controlling the particle size at 50–100 nm, which is conducive to uptake by the hepatocytes.
- (b)
- Modification of the nanoparticle delivery system to achieve targeted uptake by hepatocytes [26].
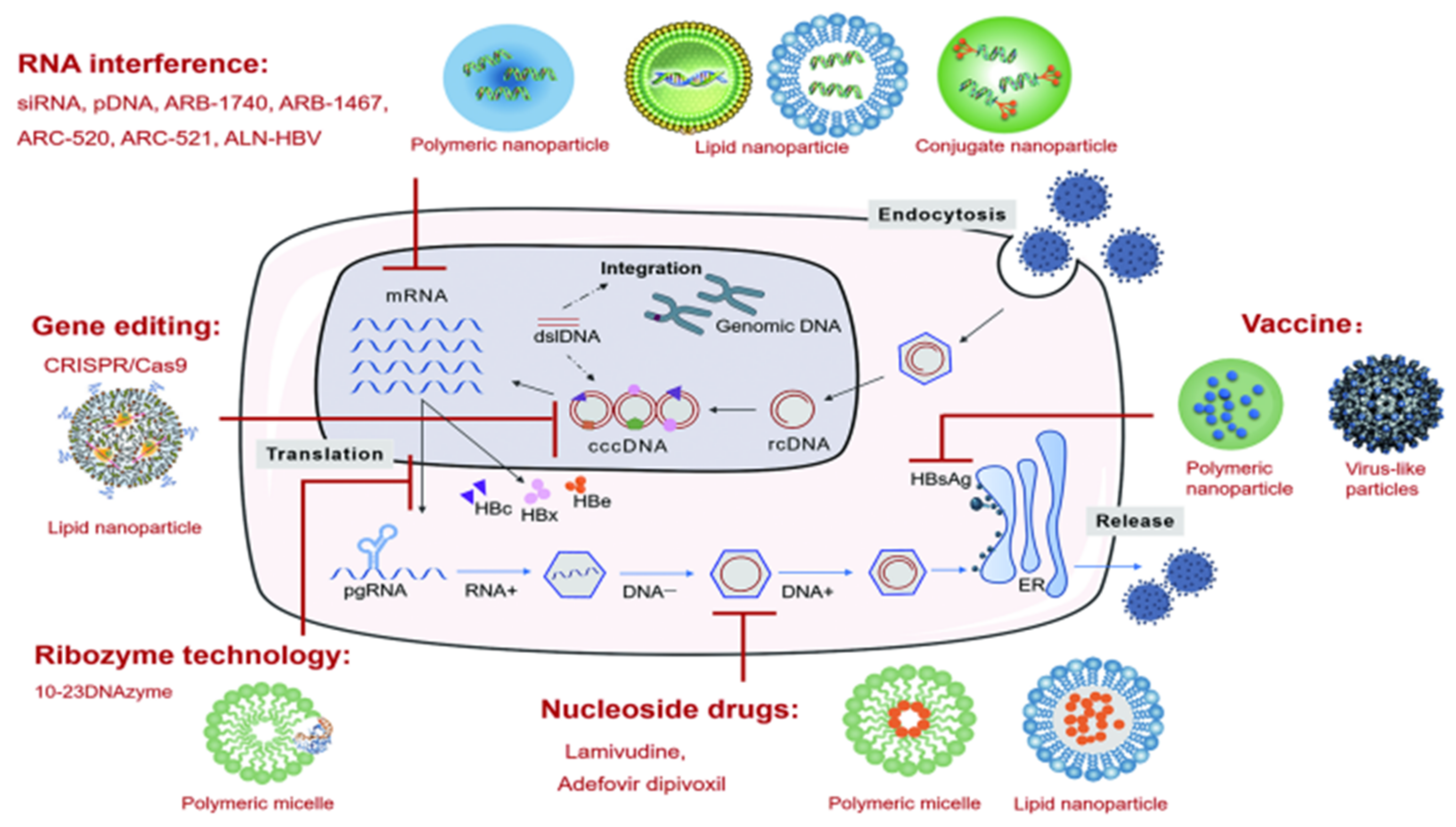
4. Nanoparticle Delivery Systems for Anti-HBV Nucleoside Drugs
4.1. Polymeric Micelles
4.2. Lipid Nanoparticles
5. Nanoparticle Delivery Systems for Anti-HBV Gene Therapy
5.1. Ribozyme Technology
5.2. RNA Interference Technology
5.2.1. Lipid Nanoparticles (LNPs)
5.2.2. Polymeric Nanoparticles
5.2.3. Conjugate Nanoparticles
5.3. Gene Editing Technology
6. Nanoparticle Delivery Systems for Vaccines
6.1. Polymeric Nanoparticles
6.2. Virus-like Particles
7. Discussion
- (1)
- Further research on nanoparticle drug delivery systems is required for their effective distribution in vivo, efficient cell targeting, drug enzyme protection, safe and stable delivery, and rapid drug release, among other aspects.
- (2)
- Future anti-HBV strategies should consider combined treatment methods, such as NAs, immune activators, and therapeutic vaccines, in order to significantly improve the quality of life of patients with CHB and achieve the ideal goal of a functional and complete cure.
- (3)
- Researchers should pay attention to the latest progress in the basic research on HBV and anti-HBV treatment, as well as identify and evaluate the factors that affect anti-HBV nanoparticle delivery in vivo, thereby improving its efficiency.
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- European Association for the Study of the Liver. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [Green Version]
- Mutimer, D.J.; Lok, A. Management of HBV- and HCV-induced end stage liver disease. Gut 2012, 61 (Suppl. S1), i59–i67. [Google Scholar] [CrossRef]
- Terrault, N.A.; Lok, A.; McMahon, B.J.; Chang, K.M.; Hwang, J.P.; Jonas, M.M.; Brown, R.J.; Bzowej, N.H.; Wong, J.B. Update on prevention, diagnosis, and treatment of chronic hepatitis B: AASLD 2018 hepatitis B guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef] [PubMed]
- Trepo, C.; Chan, H.L.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Prevention, Care and Treatment of Persons with Chronic Hepatitis B Infection; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Chevaliez, S.; Hezode, C.; Bahrami, S.; Grare, M.; Pawlotsky, J.M. Long-term hepatitis B surface antigen (HBsAg) kinetics during nucleoside/nucleotide analogue therapy: Finite treatment duration unlikely. J. Hepatol. 2013, 58, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Will, H.; Cattaneo, R.; Koch, H.G.; Darai, G.; Schaller, H.; Schellekens, H.; van Eerd, P.M.; Deinhardt, F. Cloned HBV DNA causes hepatitis in chimpanzees. Nature 1982, 299, 740–742. [Google Scholar] [CrossRef]
- Seto, W.K.; Lo, Y.R.; Pawlotsky, J.M.; Yuen, M.F. Chronic hepatitis B virus infection. Lancet 2018, 392, 2313–2324. [Google Scholar] [CrossRef]
- Dandri, M. Epigenetic modulation in chronic hepatitis B virus infection. Semin. Immunopathol. 2020, 42, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Nassal, M. HBV cccDNA: Viral persistence reservoir and key obstacle for a cure of chronic hepatitis B. Gut 2015, 64, 1972–1984. [Google Scholar] [CrossRef] [Green Version]
- Hong, X.; Kim, E.S.; Guo, H. Epigenetic regulation of hepatitis B virus covalently closed circular DNA: Implications for epigenetic therapy against chronic hepatitis B. Hepatology 2017, 66, 2066–2077. [Google Scholar] [CrossRef]
- Prescott, N.A.; Bram, Y.; Schwartz, R.E.; David, Y. Targeting Hepatitis B Virus Covalently Closed Circular DNA and Hepatitis B Virus X Protein: Recent Advances and New Approaches. ACS Infect. Dis. 2019, 5, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Gounder, P.P.; Bulkow, L.R.; Snowball, M.; Negus, S.; Spradling, P.R.; Simons, B.C.; McMahon, B.J. Nested case-control study: Hepatocellular carcinoma risk after hepatitis B surface antigen seroclearance. Aliment. Pharm. Ther. 2016, 43, 1197–1207. [Google Scholar] [CrossRef] [Green Version]
- Lau, K.; Burak, K.W.; Coffin, C.S. Impact of Hepatitis B Virus Genetic Variation, Integration, and Lymphotropism in Antiviral Treatment and Oncogenesis. Microorganisms 2020, 8, 1470. [Google Scholar] [CrossRef]
- Testoni, B.; Durantel, D.; Zoulim, F. Novel targets for hepatitis B virus therapy. Liver Int. 2017, 37 (Suppl. S1), 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, V.; Barreiro, P.; Benitez, L.; Pena, J.M.; de Mendoza, C. New antivirals for the treatment of chronic hepatitis B. Expert Opin. Investig. Drugs 2017, 26, 843–851. [Google Scholar] [CrossRef]
- Bertoletti, A.; Le Bert, N. Immunotherapy for Chronic Hepatitis B Virus Infection. Gut Liver 2018, 12, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, A.; Vasilieva, L.; Karayiannis, P. New Approaches to the Treatment of Chronic Hepatitis B. J. Clin. Med. 2020, 9, 3187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Miao, J.; Li, M.W.; Jiang, S.P.; Hu, F.Q.; Du, Y.Z. Solid lipid nanoparticles loading adefovir dipivoxil for antiviral therapy. J. Zhejiang Univ. Sci. B 2008, 9, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Du, Y.Z.; Yuan, H.; Zhang, X.G.; Miao, J.; Cui, F.D.; Hu, F.Q. Synthesis of lamivudine stearate and antiviral activity of stearic acid-g-chitosan oligosaccharide polymeric micelles delivery system. Eur. J. Pharm. Sci. 2010, 41, 498–507. [Google Scholar] [CrossRef]
- Mishra, D.; Jain, N.; Rajoriya, V.; Jain, A.K. Glycyrrhizin conjugated chitosan nanoparticles for hepatocyte-targeted delivery of lamivudine. J. Pharm. Pharm. 2014, 66, 1082–1093. [Google Scholar] [CrossRef]
- Cavalcanti, S.; Nunes, C.; Lima, S.; Soares-Sobrinho, J.L.; Reis, S. Multiple Lipid Nanoparticles (MLN), a New Generation of Lipid Nanoparticles for Drug Delivery Systems: Lamivudine-MLN Experimental Design. Pharm. Res. 2017, 34, 1204–1216. [Google Scholar] [CrossRef] [PubMed]
- Gudima, S.; He, Y.; Meier, A.; Chang, J.; Chen, R.; Jarnik, M.; Nicolas, E.; Bruss, V.; Taylor, J. Assembly of hepatitis delta virus: Particle characterization, including the ability to infect primary human hepatocytes. J. Virol. 2007, 81, 3608–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wang, H.; Ong, Z.Y.; Xu, K.; Ee, P.L.R.; Zheng, S.; Hedrick, J.L.; Yang, Y. Polymer- and lipid-based nanoparticle therapeutics for the treatment of liver diseases. Nano Today 2010, 5, 296–312. [Google Scholar] [CrossRef]
- Sato, Y.; Matsui, H.; Yamamoto, N.; Sato, R.; Munakata, T.; Kohara, M.; Harashima, H. Highly specific delivery of siRNA to hepatocytes circumvents endothelial cell-mediated lipid nanoparticle-associated toxicity leading to the safe and efficacious decrease in the hepatitis B virus. J. Control. Release 2017, 266, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Cuestas, M.L.; Mathet, V.L.; Oubina, J.R.; Sosnik, A. Drug delivery systems and liver targeting for the improved pharmacotherapy of the hepatitis B virus (HBV) infection. Pharm. Res. 2010, 27, 1184–1202. [Google Scholar] [CrossRef]
- D’Souza, A.A.; Devarajan, P.V. Asialoglycoprotein receptor mediated hepatocyte targeting—Strategies and applications. J. Control. Release 2015, 203, 126–139. [Google Scholar] [CrossRef]
- Lin, A.; Liu, Y.; Huang, Y.; Sun, J.; Wu, Z.; Zhang, X.; Ping, Q. Glycyrrhizin surface-modified chitosan nanoparticles for hepatocyte-targeted delivery. Int. J. Pharm. 2008, 359, 247–253. [Google Scholar] [CrossRef]
- Wang, T.; Zhen, Y.; Ma, X.; Wei, B.; Li, S.; Wang, N. Mannosylated and lipid A-incorporating cationic liposomes constituting microneedle arrays as an effective oral mucosal HBV vaccine applicable in the controlled temperature chain. Colloids Surf. B Biointerfaces 2015, 126, 520–530. [Google Scholar] [CrossRef]
- Biessen, E.A.; Valentijn, A.R.; De Vrueh, R.L.; Van De Bilt, E.; Sliedregt, L.A.; Prince, P.; Bijsterbosch, M.K.; Van Boom, J.H.; Van Der Marel, G.A.; Abrahams, P.J.; et al. Novel hepatotrophic prodrugs of the antiviral nucleoside 9-(2-phosphonylmethoxyethyl)adenine with improved pharmacokinetics and antiviral activity. FASEB J. 2000, 14, 1784–1792. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, D.T.; Yan, K.C.; Sedgwick, A.C.; Chen, G.R.; He, X.P.; James, T.D.; Ye, B.; Hu, X.L.; Chen, D. A glycoconjugate-based gold nanoparticle approach for the targeted treatment of Pseudomonas aeruginosa biofilms. Nanoscale 2020, 12, 23234–23240. [Google Scholar] [CrossRef]
- Kato, Y.; Onishi, H.; Machida, Y. Biological characteristics of lactosaminated N-succinyl-chitosan as a liver-specific drug carrier in mice. J. Control. Release 2001, 70, 295–307. [Google Scholar] [CrossRef]
- Wang, H.X.; Xiong, M.H.; Wang, Y.C.; Zhu, J.; Wang, J. N-acetylgalactosamine functionalized mixed micellar nanoparticles for targeted delivery of siRNA to liver. J. Control. Release 2013, 166, 106–114. [Google Scholar] [CrossRef]
- Detampel, P.; Witzigmann, D.; Krahenbuhl, S.; Huwyler, J. Hepatocyte targeting using pegylated asialofetuin-conjugated liposomes. J. Drug Target. 2014, 22, 232–241. [Google Scholar] [CrossRef]
- Nishiyama, N.; Kato, Y.; Sugiyama, Y.; Kataoka, K. Cisplatin-loaded polymer-metal complex micelle with time-modulated decaying property as a novel drug delivery system. Pharm. Res. 2001, 18, 1035–1041. [Google Scholar] [CrossRef]
- Kovacevic, M.; Balaz, I.; Marson, D.; Laurini, E.; Jovic, B. Mixed-monolayer functionalized gold nanoparticles for cancer treatment: Atomistic molecular dynamics simulations study. Biosystems 2021, 202, 104354. [Google Scholar] [CrossRef] [PubMed]
- Amarnath, P.R.; Sumathra, M.; Sam, E.R.; Vignesh, S.; Shakila, H.; Rajan, M. Fabrication of bioactive rifampicin loaded kappa-Car-MA-INH/Nano hydroxyapatite composite for tuberculosis osteomyelitis infected tissue regeneration. Int. J. Pharm. 2019, 565, 543–556. [Google Scholar] [CrossRef]
- Constantin, M.; Bucatariu, S.; Harabagiu, V.; Popescu, I.; Ascenzi, P.; Fundueanu, G. Poly(N-isopropylacrylamide-co-methacrylic acid) pH/thermo-responsive porous hydrogels as self-regulated drug delivery system. Eur. J. Pharm. Sci. 2014, 62, 86–95. [Google Scholar] [CrossRef]
- Mohanta, V.; Madras, G.; Patil, S. Albumin-mediated incorporation of water-insoluble therapeutics in layer-by-layer assembled thin films and microcapsules. J. Mater. Chem. B 2013, 1, 4819–4827. [Google Scholar] [CrossRef] [PubMed]
- Muppalla, R.; Jewrajka, S.K.; Prasad, K. Self-assembly of model graft copolymers of agarose and weak polyelectrolyte-based amphiphilic diblock copolymers: Controlled drug release and degradation. J. Biomed. Mater. Res. Part A 2013, 101, 1637–1650. [Google Scholar] [CrossRef]
- Battaglia, L.; Gallarate, M. Lipid nanoparticles: State of the art, new preparation methods and challenges in drug delivery. Expert Opin. Drug Deliv. 2012, 9, 497–508. [Google Scholar] [CrossRef]
- Zhao, G.; Hu, C.; Sun, R.; Ni, S.; Li, Q.; Xia, Q. Development of novel composite antioxidant multiple lipid particles from combination of W/O/W multiple emulsions and solid lipid nanoparticles. Eur. J. Lipid Sci. Tech. 2015, 117, 1056–1065. [Google Scholar] [CrossRef]
- Schinazi, R.F.; Ehteshami, M.; Bassit, L.; Asselah, T. Towards HBV curative therapies. Liver Int. 2018, 38 (Suppl. S1), 102–114. [Google Scholar] [CrossRef] [Green Version]
- Fanning, G.C.; Zoulim, F.; Hou, J.; Bertoletti, A. Therapeutic strategies for hepatitis B virus infection: Towards a cure. Nat. Rev. Drug Discov. 2019, 18, 827–844. [Google Scholar] [CrossRef]
- Gane, E.J. Future anti-HBV strategies. Liver Int. 2017, 37 (Suppl. S1), 40–44. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.E.; Ehrhardt, A.; Kay, M.A. Progress and problems with the use of viral vectors for gene therapy. Nat. Rev. Genet. 2003, 4, 346–358. [Google Scholar] [CrossRef]
- Miao, J.; Zhang, X.; Hong, Y.; Rao, Y.; Li, Q.; Xie, X.; Wo, J.; Li, M. Inhibition on hepatitis B virus e-gene expression of 10–23 DNAzyme delivered by novel chitosan oligosaccharide–stearic acid micelles. Carbohyd. Polym. 2012, 87, 1342–1347. [Google Scholar] [CrossRef]
- Miao, J.; Yang, X.Q.; Gao, Z.; Li, Q.; Meng, T.T.; Wu, J.Y.; Yuan, H.; Hu, F.Q. Redox-responsive chitosan oligosaccharide-SS-Octadecylamine polymeric carrier for efficient anti-Hepatitis B Virus gene therapy. Carbohydr. Polym. 2019, 212, 215–221. [Google Scholar] [CrossRef]
- Carmona, S.; Jorgensen, M.R.; Kolli, S.; Crowther, C.; Salazar, F.H.; Marion, P.L.; Fujino, M.; Natori, Y.; Thanou, M.; Arbuthnot, P.; et al. Controlling HBV replication in vivo by intravenous administration of triggered PEGylated siRNA-nanoparticles. Mol. Pharm. 2009, 6, 706–717. [Google Scholar] [CrossRef]
- Thi, E.P.; Dhillon, A.P.; Ardzinski, A.; Bidirici-Ertekin, L.; Cobarrubias, K.D.; Cuconati, A.; Kondratowicz, A.S.; Kwak, K.; Li, A.; Miller, A.; et al. ARB-1740, a RNA Interference Therapeutic for Chronic Hepatitis B Infection. ACS Infect. Dis. 2019, 5, 725–737. [Google Scholar] [CrossRef]
- Ye, X.; Tateno, C.; Thi, E.P.; Kakuni, M.; Snead, N.M.; Ishida, Y.; Barnard, T.R.; Sofia, M.J.; Shimada, T.; Lee, A. Hepatitis B Virus Therapeutic Agent ARB-1740 Has Inhibitory Effect on Hepatitis Delta Virus in a New Dually-Infected Humanized Mouse Model. ACS Infect. Dis. 2019, 5, 738–749. [Google Scholar] [CrossRef]
- Streinu-Cercel, A.; Gane, E.; Cheng, W.; Sievert, W.; Roberts, S.; Ahn, S.H.; Kim, Y.J.; Agarwal, K.; Niforos, D.; Symonds, B.; et al. SAT-155: A phase 2a study evaluating the multi-dose activity of ARB-1467 in HBeAg positive and negative virally suppressed subjects with hepatitis B. J. Hepatol. 2017, 1, S688–S689. [Google Scholar] [CrossRef]
- Zeng, P.; Xu, Y.; Zeng, C.; Ren, H.; Peng, M. Chitosan-modified poly(d,l-lactide-co-glycolide) nanospheres for plasmid DNA delivery and HBV gene-silencing. Int. J. Pharm. 2011, 415, 259–266. [Google Scholar] [CrossRef]
- Wooddell, C.I.; Yuen, M.F.; Chan, H.L.; Gish, R.G.; Locarnini, S.A.; Chavez, D.; Ferrari, C.; Given, B.D.; Hamilton, J.; Kanner, S.B.; et al. RNAi-based treatment of chronically infected patients and chimpanzees reveals that integrated hepatitis B virus DNA is a source of HBsAg. Sci. Transl. Med. 2017, 9, eaan0241. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J. Overcoming the challenges of RNAi-based therapy: An interview with James Hamilton. Ther. Deliv. 2018, 9, 511–513. [Google Scholar] [CrossRef]
- Xia, C.; Chen, Y.C.; Gong, H.; Zeng, W.; Vu, G.P.; Trang, P.; Lu, S.; Wu, J.; Liu, F. Inhibition of hepatitis B virus gene expression and replication by ribonuclease P. Mol. Ther. 2013, 21, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.J.; Brown, C.R.; Shaikh, S.; Trapp, C.; Schlegel, M.K.; Qian, K.; Sehgal, A.; Rajeev, K.G.; Jadhav, V.; Manoharan, M.; et al. Advanced siRNA Designs Further Improve In Vivo Performance of GalNAc-siRNA Conjugates. Mol. Ther. 2018, 26, 708–717. [Google Scholar] [CrossRef]
- Janas, M.M.; Zlatev, I.; Liu, J.; Jiang, Y.; Barros, S.A.; Sutherland, J.E.; Davis, W.P.; Liu, J.; Brown, C.R.; Liu, X.; et al. Safety evaluation of 2′-deoxy-2′-fluoro nucleotides in GalNAc-siRNA conjugates. Nucleic Acids Res. 2019, 47, 3306–3320. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Mei, M.; Li, B.; Zhu, X.; Zu, W.; Tian, Y.; Wang, Q.; Guo, Y.; Dong, Y.; Tan, X. A non-viral CRISPR/Cas9 delivery system for therapeutically targeting HBV DNA and pcsk9 in vivo. Cell Res. 2017, 27, 440–443. [Google Scholar] [CrossRef]
- Suzuki, Y.; Onuma, H.; Sato, R.; Sato, Y.; Hashiba, A.; Maeki, M.; Tokeshi, M.; Kayesh, M.; Kohara, M.; Tsukiyama-Kohara, K.; et al. Lipid nanoparticles loaded with ribonucleoprotein-oligonucleotide complexes synthesized using a microfluidic device exhibit robust genome editing and hepatitis B virus inhibition. J. Control. Release 2021, 330, 61–71. [Google Scholar] [CrossRef]
- Asif-Ullah, M.; Levesque, M.; Robichaud, G.; Perreault, J.P. Development of ribozyme-based gene-inactivations: The example of the hepatitis delta virus ribozyme. Curr. Gene Ther. 2007, 7, 205–216. [Google Scholar] [CrossRef]
- Hou, W.; Wo, J.E.; Li, M.W.; Liu, K.Z. In vitro cleavage of hepatitis B virus C mRNA by 10-23 DNA enzyme. Hepatobiliary Pancreat. Dis. Int. 2005, 4, 573–576. [Google Scholar]
- Hou, W.; Ni, Q.; Wo, J.; Li, M.; Liu, K.; Chen, L.; Hu, Z.; Liu, R.; Hu, M. Inhibition of hepatitis B virus X gene expression by 10-23 DNAzymes. Antivir. Res. 2006, 72, 190–196. [Google Scholar] [CrossRef]
- Haussecker, D. Current issues of RNAi therapeutics delivery and development. J. Control. Release 2014, 195, 49–54. [Google Scholar] [CrossRef]
- Gish, R.G.; Yuen, M.F.; Chan, H.L.; Given, B.D.; Lai, C.L.; Locarnini, S.A.; Lau, J.Y.; Wooddell, C.I.; Schluep, T.; Lewis, D.L. Synthetic RNAi triggers and their use in chronic hepatitis B therapies with curative intent. Antivir. Res. 2015, 121, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Torrecilla, J.; Rodriguez-Gascon, A.; Solinis, M.A.; Del, P.A. Lipid nanoparticles as carriers for RNAi against viral infections: Current status and future perspectives. Biomed. Res. Int. 2014, 2014, 161794. [Google Scholar] [CrossRef]
- Sajeesh, S.; Lee, T.Y.; Choe, J.Y.; Lee, D.K. An advanced liver targeting RNAi based therapeutic system using tripodal RNA-polymer nanoparticles. J. Control. Release 2015, 213, e95. [Google Scholar] [CrossRef]
- Springer, A.D.; Dowdy, S.F. GalNAc-siRNA Conjugates: Leading the Way for Delivery of RNAi Therapeutics. Nucleic Acid Ther. 2018, 28, 109–118. [Google Scholar] [CrossRef]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nat. Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef]
- Gagnon, K.T.; Watts, J.K. 10th Annual Meeting of the Oligonucleotide Therapeutics Society. Nucleic Acid Ther. 2014, 24, 428–434. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Yin, L.; Tang, C.; Yin, C. Multifunctional polymeric nanoparticles for oral delivery of TNF-alpha siRNA to macrophages. Biomaterials 2013, 34, 2843–2854. [Google Scholar] [CrossRef]
- Ravi, K.M.; Bakowsky, U.; Lehr, C.M. Preparation and characterization of cationic PLGA nanospheres as DNA carriers. Biomaterials 2004, 25, 1771–1777. [Google Scholar] [CrossRef]
- Yuen, M.F.; Schiefke, I.; Yoon, J.H.; Ahn, S.H.; Heo, J.; Kim, J.H.; Lik, Y.C.H.; Yoon, K.T.; Klinker, H.; Manns, M.; et al. RNA Interference Therapy With ARC-520 Results in Prolonged Hepatitis B Surface Antigen Response in Patients with Chronic Hepatitis B Infection. Hepatology 2020, 72, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Cradick, T.J.; Keck, K.; Bradshaw, S.; Jamieson, A.C.; McCaffrey, A.P. Zinc-finger nucleases as a novel therapeutic strategy for targeting hepatitis B virus DNAs. Mol. Ther. 2010, 18, 947–954. [Google Scholar] [CrossRef]
- Aravalli, R.N.; Steer, C.J. Gene editing technology as an approach to the treatment of liver diseases. Expert Opin. Biol. Ther. 2016, 16, 595–608. [Google Scholar] [CrossRef]
- Ely, A.; Moyo, B.; Arbuthnot, P. Progress with Developing Use of Gene Editing to Cure Chronic Infection with Hepatitis B Virus. Mol. Ther. 2016, 24, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ely, A.; Singh, P.; Smith, T.S.; Arbuthnot, P. In vitro transcribed mRNA for expression of designer nucleases: Advantages as a novel therapeutic for the management of chronic HBV infection. Adv. Drug Deliv. Rev. 2021, 168, 134–146. [Google Scholar] [CrossRef]
- Cui, X. Molecular Aspects of Zinc Finger Nucleases (ZFNs)-Mediated Gene Editing in Rat Embryos. Methods Mol. Biol. 2019, 1874, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, J.; Niu, Y.; Qin, D.; Liu, H.; Li, G.; Hu, Y.; Wang, J.; Lu, Y.; Kang, Y.; et al. Modeling Rett Syndrome Using TALEN-Edited MECP2 Mutant Cynomolgus Monkeys. Cell 2017, 169, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Sorek, R.; Kunin, V.; Hugenholtz, P. CRISPR—A widespread system that provides acquired resistance against phages in bacteria and archaea. Nat. Rev. Microbiol. 2008, 6, 181–186. [Google Scholar] [CrossRef]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, H.; Yoshida, K.; Hozumi, A.; Sasakura, Y. CRISPR/Cas9-mediated gene knockout in the ascidian Ciona intestinalis. Dev. Growth Differ. 2014, 56, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.V.; Nunez, J.K.; Doudna, J.A. Biology and Applications of CRISPR Systems: Harnessing Nature’s Toolbox for Genome Engineering. Cell 2016, 164, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Bloom, K.; Maepa, M.B.; Ely, A.; Arbuthnot, P. Gene Therapy for Chronic HBV-Can We Eliminate cccDNA? Genes 2018, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Ramanan, V.; Shlomai, A.; Cox, D.B.; Schwartz, R.E.; Michailidis, E.; Bhatta, A.; Scott, D.A.; Zhang, F.; Rice, C.M.; Bhatia, S.N. CRISPR/Cas9 cleavage of viral DNA efficiently suppresses hepatitis B virus. Sci. Rep. 2015, 5, 10833. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.R.; Yang, H.C.; Kuo, Y.T.; Liu, C.J.; Yang, T.Y.; Sung, K.C.; Lin, Y.Y.; Wang, H.Y.; Wang, C.C.; Shen, Y.C.; et al. The CRISPR/Cas9 System Facilitates Clearance of the Intrahepatic HBV Templates In Vivo. Mol. Ther. Nucleic Acids 2014, 3, e186. [Google Scholar] [CrossRef]
- Liu, X.; Hao, R.; Chen, S.; Guo, D.; Chen, Y. Inhibition of hepatitis B virus by the CRISPR/Cas9 system via targeting the conserved regions of the viral genome. J. Gen. Virol. 2015, 96, 2252–2261. [Google Scholar] [CrossRef]
- Seeger, C.; Sohn, J.A. Targeting Hepatitis B Virus With CRISPR/Cas9. Mol. Ther. Nucleic Acids 2014, 3, e216. [Google Scholar] [CrossRef]
- Dong, C.; Qu, L.; Wang, H.; Wei, L.; Dong, Y.; Xiong, S. Targeting hepatitis B virus cccDNA by CRISPR/Cas9 nuclease efficiently inhibits viral replication. Antivir. Res. 2015, 118, 110–117. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Bassit, L.C.; Mueller, H.; Kornepati, A.; Bogerd, H.P.; Nie, T.; Chatterjee, P.; Javanbakht, H.; Schinazi, R.F.; Cullen, B.R. Suppression of hepatitis B virus DNA accumulation in chronically infected cells using a bacterial CRISPR/Cas RNA-guided DNA endonuclease. Virology 2015, 476, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Sheng, C.; Liu, H.; Liu, G.; Du, X.; Du, J.; Zhan, L.; Li, P.; Yang, C.; Qi, L.; et al. An Effective Molecular Target Site in Hepatitis B Virus S Gene for Cas9 Cleavage and Mutational Inactivation. Int. J. Biol. Sci. 2016, 12, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Lucifora, J.; Protzer, U. Attacking hepatitis B virus cccDNA—The holy grail to hepatitis B cure. J. Hepatol. 2016, 64, S41–S48. [Google Scholar] [CrossRef]
- Yang, H.C.; Chen, P.J. The potential and challenges of CRISPR-Cas in eradication of hepatitis B virus covalently closed circular DNA. Virus Res. 2018, 244, 304–310. [Google Scholar] [CrossRef]
- Senis, E.; Fatouros, C.; Grosse, S.; Wiedtke, E.; Niopek, D.; Mueller, A.K.; Borner, K.; Grimm, D. CRISPR/Cas9-mediated genome engineering: An adeno-associated viral (AAV) vector toolbox. Biotechnol. J. 2014, 9, 1402–1412. [Google Scholar] [CrossRef]
- Moreno, A.M.; Fu, X.; Zhu, J.; Katrekar, D.; Shih, Y.V.; Marlett, J.; Cabotaje, J.; Tat, J.; Naughton, J.; Lisowski, L.; et al. In Situ Gene Therapy via AAV-CRISPR-Cas9-Mediated Targeted Gene Regulation. Mol. Ther. 2018, 26, 1818–1827. [Google Scholar] [CrossRef] [Green Version]
- Chew, W.L.; Tabebordbar, M.; Cheng, J.K.; Mali, P.; Wu, E.Y.; Ng, A.H.; Zhu, K.; Wagers, A.J.; Church, G.M. A multifunctional AAV-CRISPR-Cas9 and its host response. Nat. Methods 2016, 13, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Hou, X.; Zuo, F.; Li, X.; Pang, Y.; Jiang, G. Application of nanoparticle-based siRNA and CRISPR/Cas9 delivery systems in gene-targeted therapy. Nanomedicine 2019, 14, 511–514. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, C.; Zheng, Y.; Zhao, Y.; Wang, Y.; Hao, J.; Zhao, X.; Yi, K.; Shi, L.; Kang, C.; et al. Virus-like nanoparticle as a co-delivery system to enhance efficacy of CRISPR/Cas9-based cancer immunotherapy. Biomaterials 2020, 258, 120275. [Google Scholar] [CrossRef]
- Wang, Y.; Shahi, P.K.; Xie, R.; Zhang, H.; Abdeen, A.A.; Yodsanit, N.; Ma, Z.; Saha, K.; Pattnaik, B.R.; Gong, S. A pH-responsive silica-metal-organic framework hybrid nanoparticle for the delivery of hydrophilic drugs, nucleic acids, and CRISPR-Cas9 genome-editing machineries. J. Control. Release 2020, 324, 194–203. [Google Scholar] [CrossRef]
- Chou, S.J.; Yang, P.; Ban, Q.; Yang, Y.P.; Wang, M.L.; Chien, C.S.; Chen, S.J.; Sun, N.; Zhu, Y.; Liu, H.; et al. Dual Supramolecular Nanoparticle Vectors Enable CRISPR/Cas9-Mediated Knockin of Retinoschisin 1 Gene-A Potential Nonviral Therapeutic Solution for X-Linked Juvenile Retinoschisis. Adv. Sci. 2020, 7, 1903432. [Google Scholar] [CrossRef]
- Chen, G.; Abdeen, A.A.; Wang, Y.; Shahi, P.K.; Robertson, S.; Xie, R.; Suzuki, M.; Pattnaik, B.R.; Saha, K.; Gong, S. A biodegradable nanocapsule delivers a Cas9 ribonucleoprotein complex for in vivo genome editing. Nat. Nanotechnol. 2019, 14, 974–980. [Google Scholar] [CrossRef]
- Finn, J.D.; Smith, A.R.; Patel, M.C.; Shaw, L.; Youniss, M.R.; van Heteren, J.; Dirstine, T.; Ciullo, C.; Lescarbeau, R.; Seitzer, J.; et al. A Single Administration of CRISPR/Cas9 Lipid Nanoparticles Achieves Robust and Persistent In Vivo Genome Editing. Cell Rep. 2018, 22, 2227–2235. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.M.; Simon, J.K.; Baker, J.J. Applications of nanotechnology for immunology. Nat. Rev. Immunol. 2013, 13, 592–605. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.; Chen, Y.; Huang, K. Nano-based approaches in the development of antiviral agents and vaccines. Life Sci. 2021, 265, 118761. [Google Scholar] [CrossRef]
- Roldao, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, X.; Bian, Y.; Wang, S.; Chai, Q.; Guo, Z.; Wang, Z.; Zhu, P.; Peng, H.; Yan, X.; et al. Dual-targeting nanoparticle vaccine elicits a therapeutic antibody response against chronic hepatitis B. Nat. Nanotechnol. 2020, 15, 406–416. [Google Scholar] [CrossRef]
- Zhu, J.; Qin, F.; Ji, Z.; Fei, W.; Tan, Z.; Hu, Y.; Zheng, C. Mannose-Modified PLGA Nanoparticles for Sustained and Targeted Delivery in Hepatitis B Virus Immunoprophylaxis. AAPS PharmSciTech 2019, 21, 13. [Google Scholar] [CrossRef]
- Yong, C.Y.; Yeap, S.K.; Goh, Z.H.; Ho, K.L.; Omar, A.R.; Tan, W.S. Induction of humoral and cell-mediated immune responses by hepatitis B virus epitope displayed on the virus-like particles of prawn nodavirus. Appl. Environ. Microbiol. 2015, 81, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Ninyio, N.N.; Ho, K.L.; Yong, C.Y.; Chee, H.Y.; Hamid, M.; Ong, H.K.; Mariatulqabtiah, A.R.; Tan, W.S. Chimeric Virus-Like Particles of Prawn Nodavirus Displaying Hepatitis B Virus Immunodominant Region: Biophysical Properties and Cytokine Response. Int. J. Mol. Sci. 2021, 22, 1922. [Google Scholar] [CrossRef]
- Mobini, S.; Chizari, M.; Mafakher, L.; Rismani, E.; Rismani, E. Computational Design of a Novel VLP-Based Vaccine for Hepatitis B Virus. Front. Immunol. 2020, 11, 2074. [Google Scholar] [CrossRef]
- Whitacre, D.C.; Peters, C.J.; Sureau, C.; Nio, K.; Li, F.; Su, L.; Jones, J.E.; Isogawa, M.; Sallberg, M.; Frelin, L.; et al. Designing a therapeutic hepatitis B vaccine to circumvent immune tolerance. Hum. Vaccines Immunother. 2020, 16, 251–268. [Google Scholar] [CrossRef]
- Chan, H.L.; Thompson, A.; Martinot-Peignoux, M.; Piratvisuth, T.; Cornberg, M.; Brunetto, M.R.; Tillmann, H.L.; Kao, J.H.; Jia, J.D.; Wedemeyer, H.; et al. Hepatitis B surface antigen quantification: Why and how to use it in 2011—A core group report. J. Hepatol. 2011, 55, 1121–1131. [Google Scholar] [CrossRef]
- Shouval, D.; Roggendorf, H.; Roggendorf, M. Enhanced immune response to hepatitis B vaccination through immunization with a Pre-S1/Pre-S2/S vaccine. Med. Microbiol. Immunol. 2015, 204, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus-like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nanopreparation | Nanocarrier Composition | Ligand | Drug | Preparation Method | Particle Size (nm) | Encapsulation Efficiency (%) | Drug Loading (%) | Ref. |
|---|---|---|---|---|---|---|---|---|
| Polymeric nanoparticle | Stearic acid-grafted chitosan oligosaccharide (grafting rate 3.79%) | N/A | Lamivudine (ester linkage to stearic acid) | Dialysis method | 273.8 ± 8.5 | 99.48 ± 0.04 | 39.04 ± 0.51 | [20] |
| Low-molecular-weight chitosan | Glycyrrhizin | Lamivudine | Ionotropic gelation method | 145.8 ± 4.2 | 65.89 ± 1.58 | 71.37 ± 1.19 | [21] | |
| Solid lipid nanoparticle | Monostearin (poloxamer 188 as surfactant) | N/A | Adefovir dipivoxil | Solvent diffusion method | 389.4 ± 166.5 | 15.32 ± 2.58 | 3.06 ± 0.51 | [19] |
| Multiple lipid nanoparticle | Compritol® ATO 888, Miglyol®-812 (Span-80 as surfactant) | N/A | Lamivudine | Hot homogenization method combined with high shear and ultrasonication | 450 ± 10 | 20 ± 2 | 1.08 ± 0.06 | [22] |
| Nanopreparation | Nanocarrier Composition | Ligand | Drugs | Method of Preparation | Particle Size (nm) | Encapsulation Efficiency (%) | Drug Loading (%) | Refs. |
|---|---|---|---|---|---|---|---|---|
| Ribozyme technology | ||||||||
| Polymeric micelle | Chitosan oligosaccharide-grafted stearic acid | N/A | 10–23 DNAzyme specific to e-gene ORF A1816UG | Self-aggregation | 164.0 ± 2.1 | N/A | N/A | [47] |
| Chitosan oligosaccharide-SS-Octadecylamine | N/A | 10–23 DNAzyme specific to e-gene ORF A1816UG | Self-aggregation | 214.75 ± 3.43 | 96.48 ± 0.27 | 1.582 ± 0.004 | [48] | |
| 10–23 DNAzyme specific to s-gene ORF A157UG | 230.70 ± 6.16 | 96.45 ± 0.33 | 1.581 ± 0.005 | |||||
| RNA interference technology | ||||||||
| Lipid nanoparticle | Cationic cholesteryl polyamine N1-cholesteryloxycarbonyl-3,7-diazanonane-1,9-diamine and the neutral colipid dioleoyl-l,r-phosphatidyl ethanolamine | Polyethylene glycol | siRNA | Film dispersion– sonication method | 80–100 | N/A | N/A | [49] |
| Proprietary lipid nanoparticle platform (Arbutus Biopharma) | N/A | ARB-1740 | Spontaneous vesicle formation | 65–80 | 92–98 | N/A | [50,51] | |
| ARB-1467 | N/A | N/A | N/A | [52] | ||||
| Polymeric nanoparticle | Poly(d,l-lactide-co-glycolide)-grafted chitosan (PLGA–CHS) | N/A | Plasmid DNA (pDNA) | Spontaneous emulsion diffusion method | 59.43 ± 14 | N/A | Nearly 100% at the ratio of 100:1 (PLGA–CHS NS to pDNA) | [53] |
| ARC-EX1 containing hepatocyte-targeted N-acetylgalactosamine-conjugated melittin-like peptide | N/A | ARC-520 along with a related ARC-521 | N/A | N/A | N/A | N/A | [54,55] | |
| Conjugate nanoparticle | N-acetylgalactosamine (GalNAc)–siRNA conjugates | N/A | ALN-HBV | N/A | N/A | N/A | N/A | [56,57,58] |
| Gene editing technology | ||||||||
| Lipid-like nanoparticle | Tris(2-aminoethyl) benzene-1,3,5-tricarboxamide | N/A | CRISPR/Cas9 | N/A | N/A | N/A | N/A | [59] |
| Lipid nanoparticle | Cationic lipid, phospholipid, cholesterol | Polyethylene glycol | CRISPR/Cas ribonucleoprotein | Mixer-equipped microfluidic device | <200 | >80 | N/A | [60] |
| Nanopreparation | Nanocarrier Composition | Vaccine | Method of Preparation | Particle Size (nm) | Encapsulation Efficiency (%) | Drug Loading Capability | Ref. |
|---|---|---|---|---|---|---|---|
| Polymeric nanoparticle | SpyTag–ferritin | PreS1 | Self-assembly | N/A | N/A | N/A | [106] |
| Mannose-modified poly d,l-lactide-co-glycolic acid | HBsAg | Double emulsion solvent evaporation technique | 186.6 ± 3.7 | 63.7 ± 4.5 | 1.5 ± 0.1 μg/mg | [107] | |
| Virus-like particles (VLPs) | Macrobrachium rosenbergii nodavirus (MrNV) | MrNV VLPs | Self-assembly | 30 | N/A | N/A | [108] |
| HBV “a” determinant (aD) displayed on the prawn nodavirus capsid (Nc) and expressed in Spodoptera frugiperda (Sf9) cells (Nc-aD-Sf9) | Nc-aD-Sf9 VLPs | Self-assembly | 56.4 | N/A | N/A | [109] | |
| HBcAg including Myrcludex and the “a” determinant sequence of HBsAg | VLP-based vaccine by placing the antibody-binding fragments of HBsAg in the major immunodominant region epitope of HBcAg | Self-assembly | N/A | N/A | N/A | [110] | |
| The woodchuck hepatitis core antigen (WHcAg) | PreS1-WHc VLPs | Self-assembly | N/A | N/A | N/A | [111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, J.; Gao, P.; Li, Q.; He, K.; Zhang, L.; Wang, J.; Huang, L. Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis B Virus Therapy: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 11227. https://doi.org/10.3390/ijms222011227
Miao J, Gao P, Li Q, He K, Zhang L, Wang J, Huang L. Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis B Virus Therapy: A Narrative Review. International Journal of Molecular Sciences. 2021; 22(20):11227. https://doi.org/10.3390/ijms222011227
Chicago/Turabian StyleMiao, Jing, Peng Gao, Qian Li, Kaifeng He, Liwen Zhang, Junyan Wang, and Lingfei Huang. 2021. "Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis B Virus Therapy: A Narrative Review" International Journal of Molecular Sciences 22, no. 20: 11227. https://doi.org/10.3390/ijms222011227
APA StyleMiao, J., Gao, P., Li, Q., He, K., Zhang, L., Wang, J., & Huang, L. (2021). Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis B Virus Therapy: A Narrative Review. International Journal of Molecular Sciences, 22(20), 11227. https://doi.org/10.3390/ijms222011227