
Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors


 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
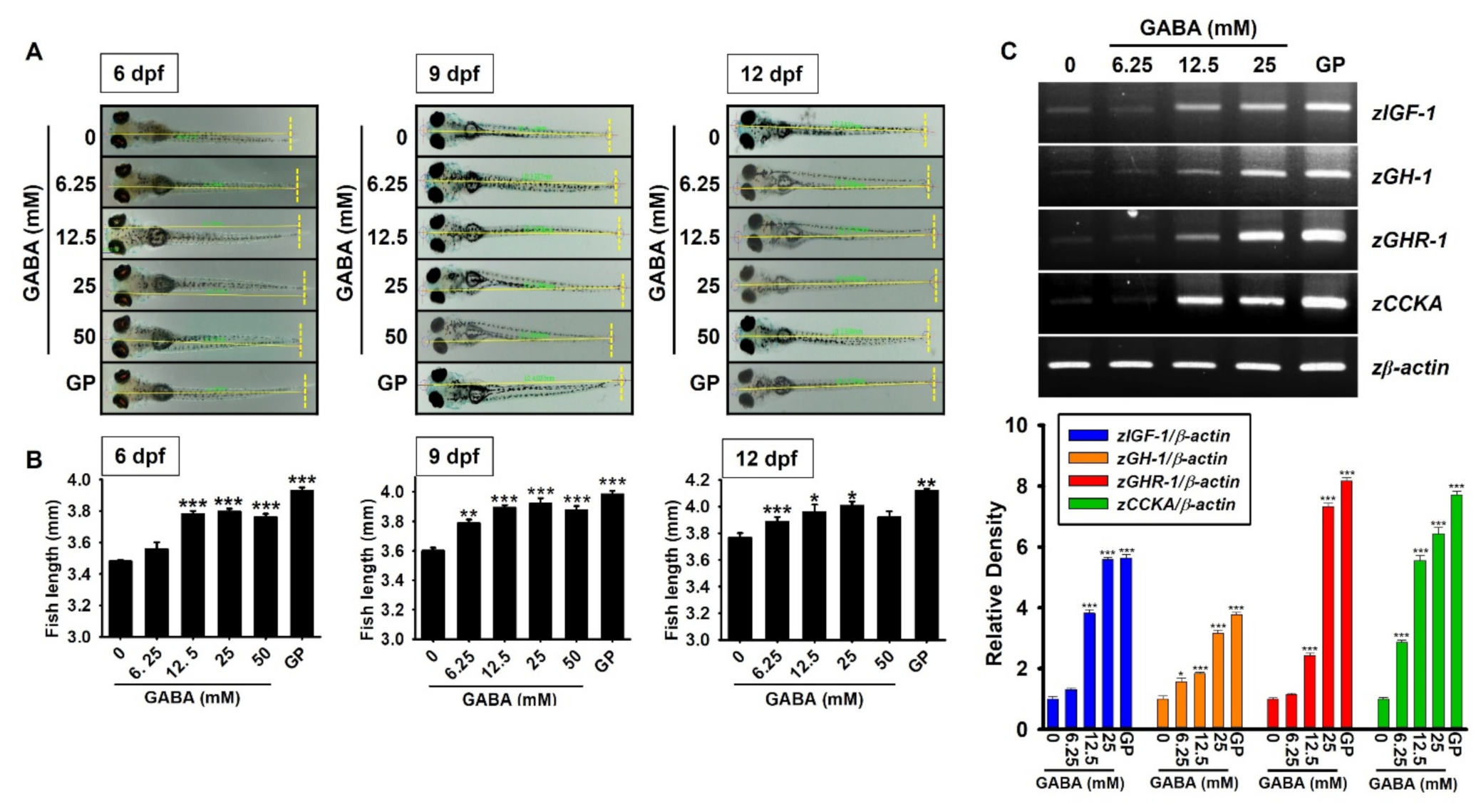
2.1. GABA Promotes Growth Rate in Zebrafish Larvae through the Upregulation of Growth-Stimulating Gene Expression
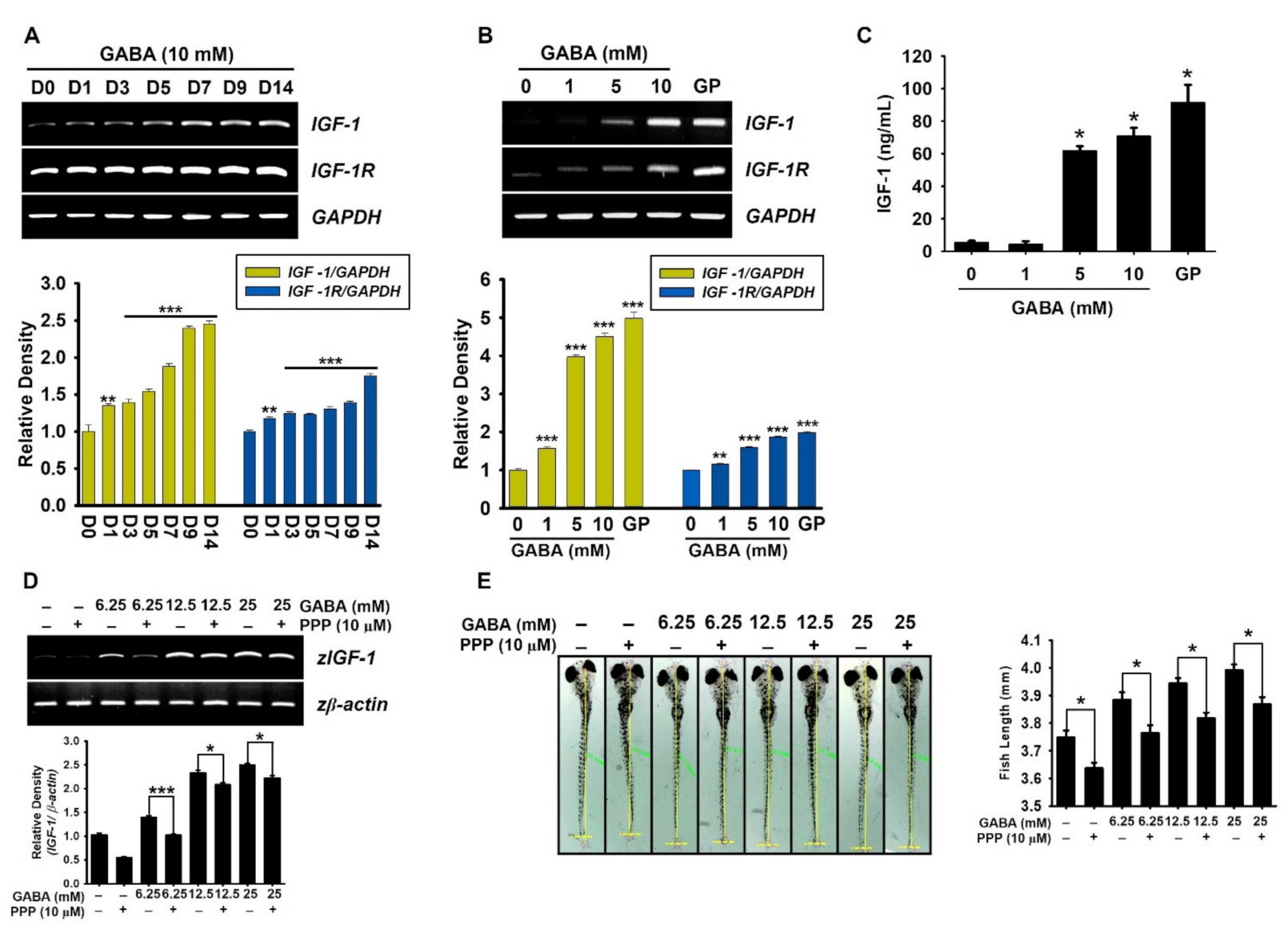
2.2. GABA Increases IGF-1-Mediated Growth Rate via IGF-1R
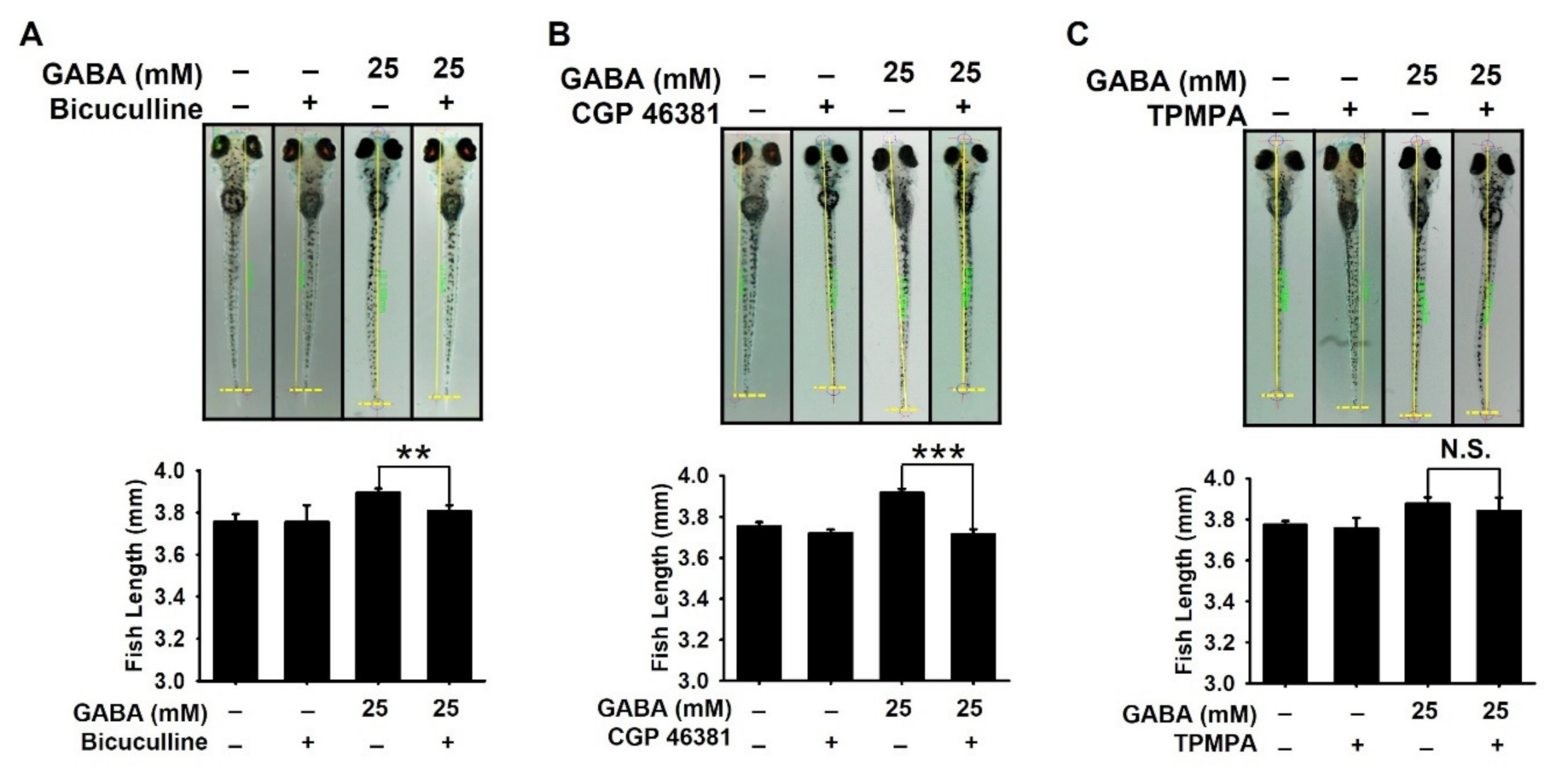
2.3. GABAA and GABAB Receptors Are Responsible for GABA-Mediated Growth Performances in Zebrafish Larvae
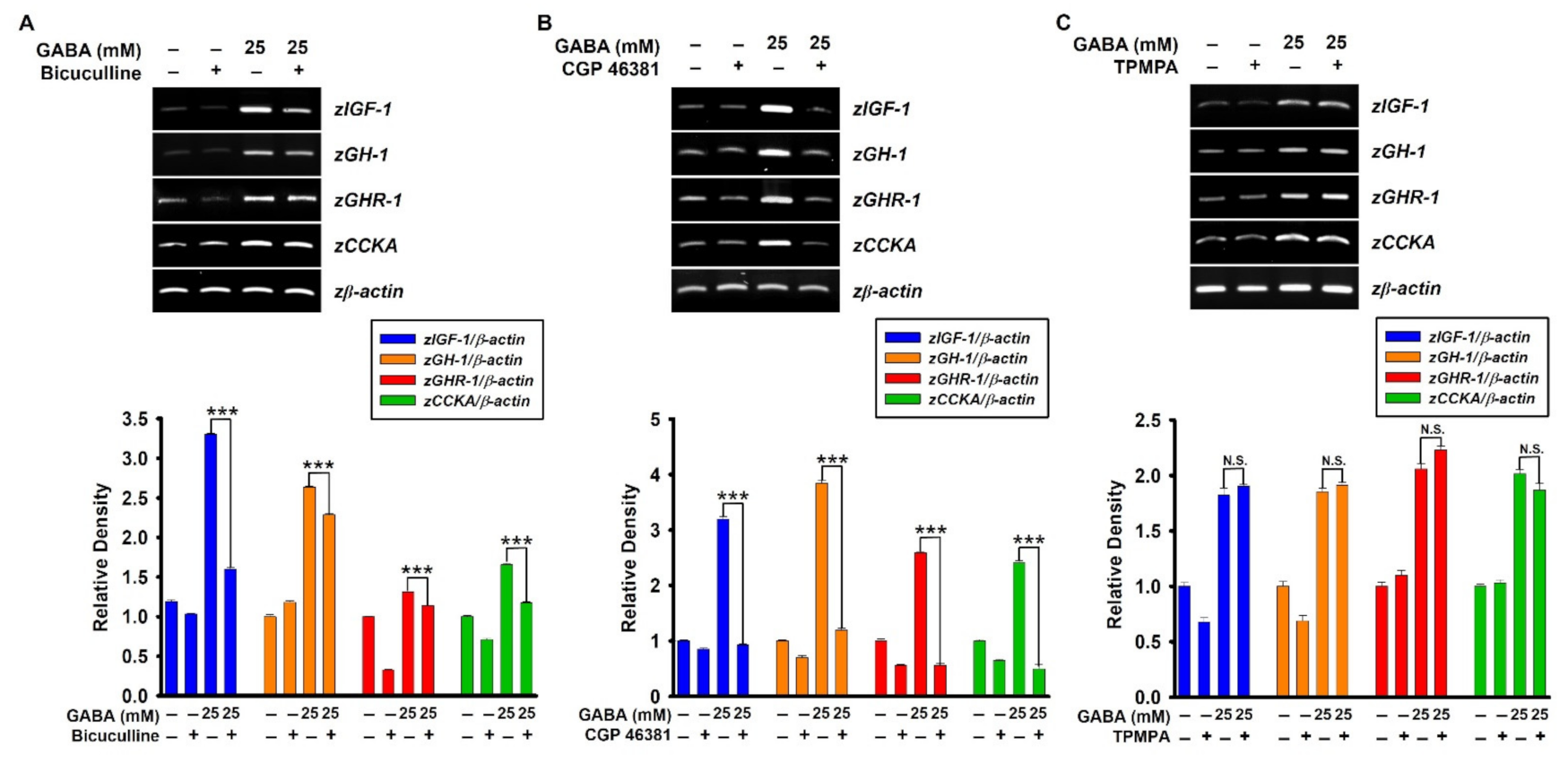
2.4. GABAA and GABAB Receptors Upregulate GABA-Mediated Growth-Stimulating Gene Expression in Zebrafish Larvae
2.5. GABAA and GABAB Receptors Upregulate GABA-Induced IGF-1 and IGF-1R Expression in Preosteoblast MC3T3-E1 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Total RNA Extraction from Preosteoblast MC3T3-E1 Cells and Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Maintenance of Zebrafish
4.6. Measurement of Total Body Length of Zebrafish Larvae and RT-PCR
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bidlingmaier, M.; Strasburger, C.J. Growth hormone. Handb. Exp. Pharmacol. 2010, 187–200. [Google Scholar] [CrossRef]
- Junnila, R.K.; List, E.O.; Berryman, D.E.; Murrey, J.W.; Kopchick, J.J. The GH/IGF-1 axis in ageing and longevity. Nat. Rev. Endocrinol. 2013, 9, 366–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakar, S.; Isaksson, O. Regulation of skeletal growth and mineral acquisition by the GH/IGF-1 axis: Lessons from mouse models. Growth Horm. IGF Res. 2016, 28, 26–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laron, Z. Insulin-like growth factor 1 (IGF-1): A growth hormone. Mol. Pathol. 2001, 54, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, R. Growth hormone deficiency in patients with obesity. Endocrine 2015, 49, 304–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deijen, J.B.; Arwert, L.I. Impaired quality of life in hypopituitary adults with growth hormone deficiency: Can somatropin replacement therapy help? Treat. Endocrinol. 2006, 5, 243–250. [Google Scholar] [CrossRef]
- Malozowski, S. Reports of increased mortality and GH: Will this affect current clinical practice? J. Clin. Endocrinol. Metab. 2012, 97, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, A.; Menon, P.S. Growth hormone therapy. Indian J. Pediatrics 2005, 72, 139–144. [Google Scholar] [CrossRef]
- Leung, K.-C.; Ho, K.K.Y. Measurement of growth hormone, insulin-like growth factor I and their binding proteins: The clinical aspects. Clin. Chim. Acta 2001, 313, 119–123. [Google Scholar] [CrossRef]
- De, W.; Bréant, B.; Czernichow, P.; Asfari, M. Growth hormone (GH) and prolactin (PRL) regulate IGFBP-3 gene expression in rat beta-cells. Mol. Cell. Endocrinol. 1995, 114, 43–50. [Google Scholar] [CrossRef]
- Kim, B.; Huang, G.; Ho, W.-B.; Greenspan, D.S. Bone Morphogenetic Protein-1 Processes Insulin-like Growth Factor-binding Protein 3. J. Biol. Chem. 2011, 286, 29014–29025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, M.O.; Hwa, V.; David, A.; Rosenfeld, R.G.; Metherell, L.A. Genetic Defects in the Growth Hormone-IGF-I Axis Causing Growth Hormone Insensitivity and Impaired Linear Growth. Front. Endocrinol. 2011, 2, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, C.M.; Perry, J.K.; Vickers, M.H. Manipulation of the Growth Hormone-Insulin-Like Growth Factor (GH-IGF) Axis: A Treatment Strategy to Reverse the Effects of Early Life Developmental Programming. Int. J. Mol. Sci. 2017, 18, 1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazeli, P.K.; Klibanski, A. Determinants of GH resistance in malnutrition. J. Endocrinol. 2014, 220, R57–R65. [Google Scholar] [CrossRef] [Green Version]
- Seid, E.; Derseh, L.; Derso, T.; Assefa, M.; Gonete, K.A.; Tariku, A. Nutrient consumption and associated factors among school age children in Dewa Chefe District, northeast Ethiopia: A cross-sectional study. BMC Res. Notes 2018, 11, 669. [Google Scholar] [CrossRef]
- Hinton, T.; Johnston, G.A. GABA, The Major Inhibitory Neurotransmitter in the Brain. Ref. Modul. Biomed. Sci. 2018, 1–15. [Google Scholar] [CrossRef]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as food supplements: The effects of GABA on brain and behavior. Front. Psychol. 2015, 6, 1520. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological stress-reducing effect of chocolate enriched with γ-aminobutyric acid (GABA) in humans: Assessment of stress using heart rate variability and salivary chromogranin A. Int. J. Food Sci. Nutr. 2009, 60, 106–113. [Google Scholar] [CrossRef]
- Yamatsu, A.; Yamashita, Y.; Pandharipande, T.; Maru, I.; Kim, M. Effect of oral γ-aminobutyric acid (GABA) administration on sleep and its absorption in humans. Food Sci. Biotechnol. 2016, 25, 547–551. [Google Scholar] [CrossRef]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef]
- Acs, Z.; Szabó, B.; Kapócs, G.; Makara, G.B. gamma-Aminobutyric acid stimulates pituitary growth hormone secretion in the neonatal rat. A superfusion study. Endocrinology 1987, 120, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Powers, M.E.; Yarrow, J.F.; McCoy, S.C.; Borst, S.E. Growth Hormone Isoform Responses to GABA Ingestion at Rest and after Exercise. Med. Sci. Sports Exerc. 2008, 40, 104. [Google Scholar] [CrossRef] [PubMed]
- Grönbladh, A.; Johansson, J.; Nyberg, F.; Hallberg, M. Recombinant Human Growth Hormone Affects the Density and Functionality of GABAB Receptors in the Male Rat Brain. Neuroendocrinology 2013, 97, 203–211. [Google Scholar] [CrossRef]
- Baloucoune, G.A.; Chun, L.; Zhang, W.; Xu, C.; Huang, S.; Sun, Q.; Wang, Y.; Tu, H.; Liu, J. GABAB Receptor Subunit GB1 at the Cell Surface Independently Activates ERK1/2 through IGF-1R Transactivation. PLoS ONE 2012, 7, e39698. [Google Scholar] [CrossRef]
- Molagoda, I.M.N.; Jayasingha, J.; Choi, Y.H.; Park, E.K.; Jeon, Y.J.; Lee, B.J.; Kim, G.Y. Fermented Oyster Extract Promotes Insulin-Like Growth Factor-1-Mediated Osteogenesis and Growth Rate. Mar. Drugs 2020, 18, 472. [Google Scholar] [CrossRef]
- Messripour, M.; Mesripour, A. Effects of vitamin B6 on age associated changes of rat brain glutamate decarboxylase activity. Afr. J. Pharm. Pharmacol. 2011, 5, 454–456. [Google Scholar] [CrossRef] [Green Version]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary Neurotransmitters: A Narrative Review on Current Knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.A.; Mitchell, R. Effects of gamma-aminobutyric acid receptor agonists on the secretion of growth hormone, luteinizing hormone, adrenocorticotrophic hormone and thyroid-stimulating hormone from the rat pituitary gland in vitro. J. Endocrinol. 1986, 108, 1–8. [Google Scholar] [CrossRef]
- Saleh, A.A.; Rashad, A.M.A.; Hassanine, N.; Sharaby, M.A.; Zhao, Y. Comparative analysis of IGFBP-3 gene sequence in Egyptian sheep, cattle, and buffalo. BMC Res. Notes 2019, 12, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xuan, S.; Bouxsein, M.L.; von Stechow, D.; Akeno, N.; Faugere, M.C.; Malluche, H.; Zhao, G.; Rosen, C.J.; Efstratiadis, A.; et al. Osteoblast-specific Knockout of the Insulin-like Growth Factor (IGF) Receptor Gene Reveals an Essential Role of IGF Signaling in Bone Matrix Mineralization *. J. Biol. Chem. 2002, 277, 44005–44012. [Google Scholar] [CrossRef] [Green Version]
- Reible, B.; Schmidmaier, G.; Moghaddam, A.; Westhauser, F. Insulin-Like Growth Factor-1 as a Possible Alternative to Bone Morphogenetic Protein-7 to Induce Osteogenic Differentiation of Human Mesenchymal Stem Cells in Vitro. Int. J. Mol. Sci. 2018, 19, 1674. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.Y.; Hou, X.Y.; Yan, F.B.; Sun, G.R.; Han, R.L.; Kang, X.T. Effect of gamma-aminobutyric acid on growth performance and immune function in chicks under beak trimming stress. Anim Sci J. 2013, 84, 121–129. [Google Scholar] [CrossRef]
- Ncho, C.M.; Jeong, C.; Gupta, V.; Goel, A. The effect of gamma-aminobutyric acid supplementation on growth performances, immune responses, and blood parameters of chickens reared under stressful environment: A meta-analysis. Environ. Sci. Pollut. Res. Int. 2021, 28, 45019–45028. [Google Scholar] [CrossRef] [PubMed]
- Minuk, G.Y.; Kaita, K.; Gauthier, T.; Dembinski, T.; Murphy, L.J. Effect of exogenous gamma-aminobutyric acid (GABA) on hepatic insulin-like growth factor I (IGF-I) and IGF-I binding protein (IGFBP-I) mRNA abundance following partial hepatectomy in rats. Can. J. Physiol Pharm. 1995, 73, 1546–1551. [Google Scholar] [CrossRef]
- Chebib, M.; Johnston, G.A. The ’ABC’ of GABA receptors: A brief review. Clin. Exp. Pharm. Physiol 1999, 26, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.G.; Bottiglieri, T.; Snead, O.C., 3rd. GABA, gamma-hydroxybutyric acid, and neurological disease. Ann. Neurol. 2003, 54 (Suppl. 6), S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Noviello, C.M.; Teng, J.; Walsh, R.M., Jr.; Kim, J.J.; Hibbs, R.E. Structure of a human synaptic GABAA receptor. Nature 2018, 559, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Bettler, B. Regulation of neuronal GABA(B) receptor functions by subunit composition. Nat. Rev. Neurosci. 2012, 13, 380–394. [Google Scholar] [CrossRef]
- Naffaa, M.M.; Hung, S.; Chebib, M.; Johnston, G.A.R.; Hanrahan, J.R. GABA-rho receptors: Distinctive functions and molecular pharmacology. Br. J. Pharm. 2017, 174, 1881–1894. [Google Scholar] [CrossRef] [Green Version]
- Tamayama, T.; Maemura, K.; Kanbara, K.; Hayasaki, H.; Yabumoto, Y.; Yuasa, M.; Watanabe, M. Expression of GABA(A) and GABA(B) receptors in rat growth plate chondrocytes: Activation of the GABA receptors promotes proliferation of mouse chondrogenic ATDC5 cells. Mol. Cell Biochem. 2005, 273, 117–126. [Google Scholar] [CrossRef]
- Takahata, Y.; Takarada, T.; Hinoi, E.; Nakamura, Y.; Fujita, H.; Yoneda, Y. Osteoblastic γ-Aminobutyric Acid, Type B Receptors Negatively Regulate Osteoblastogenesis toward Disturbance of Osteoclastogenesis Mediated by Receptor Activator of Nuclear Factor κB Ligand in Mouse Bone. J. Biol. Chem. 2011, 286, 32906–32917. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, E.T.; McCann, S.M. Effects of intraventricular injection of gamma-aminobutyric acid (GABA) on plasma growth hormone and thyrotroprin in conscious ovariectomized rats. Endocrinology 1978, 103, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, J.O.; Jervois, P.M.; Menadue, M.F.; Blessing, W.W. Activation of GABA receptors in the hypothalamus stimulates secretion of growth hormone and prolactin. Brain Res. 1986, 374, 119–125. [Google Scholar] [CrossRef]
- Gamel-Didelon, K.; Corsi, C.; Pepeu, G.; Jung, H.; Gratzl, M.; Mayerhofer, A. An autocrine role for pituitary GABA: Activation of GABA-B receptors and regulation of growth hormone levels. Neuroendocrinology 2002, 76, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. J. Physiol 2020, 598, 3793–3801. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene * | Primer Sequence (5′-3′) | Size | Tm |
|---|---|---|---|
| IGF-1 | F: 5′-GGACCAGAGACCCTTTGCGGG-3′ | 171 bp | 63 °C |
| R: 5′-GGCTGCTTTTGACCCTTCAGTGG-3′ | |||
| IGF-1R | F: 5′-ACTGACCTCATGCGCATGTGCTGG-3′ | 198 bp | 63 °C |
| R: 5′-CTCGTTCTTGCGGCCCCCGTTCAT-3′ | |||
| GAPDH | F: 5′-ACCACAGTCCATGCCATCAC-3′ | 480 bp | 63 °C |
| R: 5′-CACCACCCTGTTGCTGTAGC-3′ |
| Gene * | Primer Sequence (5′-3′) | Size | Tm |
|---|---|---|---|
| zIGF1 | F 5′-GAGTACCCACACCCTCTCAC-3′ | 213 bp | 56 °C |
| R 5′-TGAAAGCAGCATTCGTCCAC-3′ | |||
| zGH-1 | F 5′-GGTGGTGGTTAGTTTGCTGG-3′ | 157 bp | 56 °C |
| R 5′-CAACTGTCTGCGTTCCTCAG-3′ | |||
| zGHR-1 | F 5′-TCAGTCCGACTCAGAAACCG-3′ | 178 bp | 56 °C |
| R 5′-TTCTGAAGCACGGGACCATA-3′ | |||
| zCCKA | F 5′-GATGAAGAACCTCGCAGCAG-3′ | 154 bp | 56 °C |
| R 5′-GGCCAAAATCCATCCATCCC-3′ | |||
| zβ-actin | F 5′-CGAGCGTGGCTACAGCTTCA-3′ | 155 bp | 60 °C |
| R 5′-GACCGTCAGGCAGCTCATAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Athapaththu, A.M.G.K.; Molagoda, I.M.N.; Jayasooriya, R.G.P.T.; Choi, Y.H.; Jeon, Y.-J.; Park, J.-H.; Lee, B.-J.; Kim, G.-Y. Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors. Int. J. Mol. Sci. 2021, 22, 11254. https://doi.org/10.3390/ijms222011254
Athapaththu AMGK, Molagoda IMN, Jayasooriya RGPT, Choi YH, Jeon Y-J, Park J-H, Lee B-J, Kim G-Y. Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors. International Journal of Molecular Sciences. 2021; 22(20):11254. https://doi.org/10.3390/ijms222011254
Chicago/Turabian StyleAthapaththu, Athapaththu Mudiyanselage Gihan Kavinda, Ilandarage Menu Neelaka Molagoda, Rajapaksha Gedara Prasad Tharanga Jayasooriya, Yung Hyun Choi, You-Jin Jeon, Joung-Hyun Park, Bae-Jin Lee, and Gi-Young Kim. 2021. "Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors" International Journal of Molecular Sciences 22, no. 20: 11254. https://doi.org/10.3390/ijms222011254
APA StyleAthapaththu, A. M. G. K., Molagoda, I. M. N., Jayasooriya, R. G. P. T., Choi, Y. H., Jeon, Y. -J., Park, J. -H., Lee, B. -J., & Kim, G. -Y. (2021). Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors. International Journal of Molecular Sciences, 22(20), 11254. https://doi.org/10.3390/ijms222011254