
Involvement of GPR17 in Neuronal Fibre Outgrowth

 , , and
, , and
Abstract
:1. Introduction
2. Results
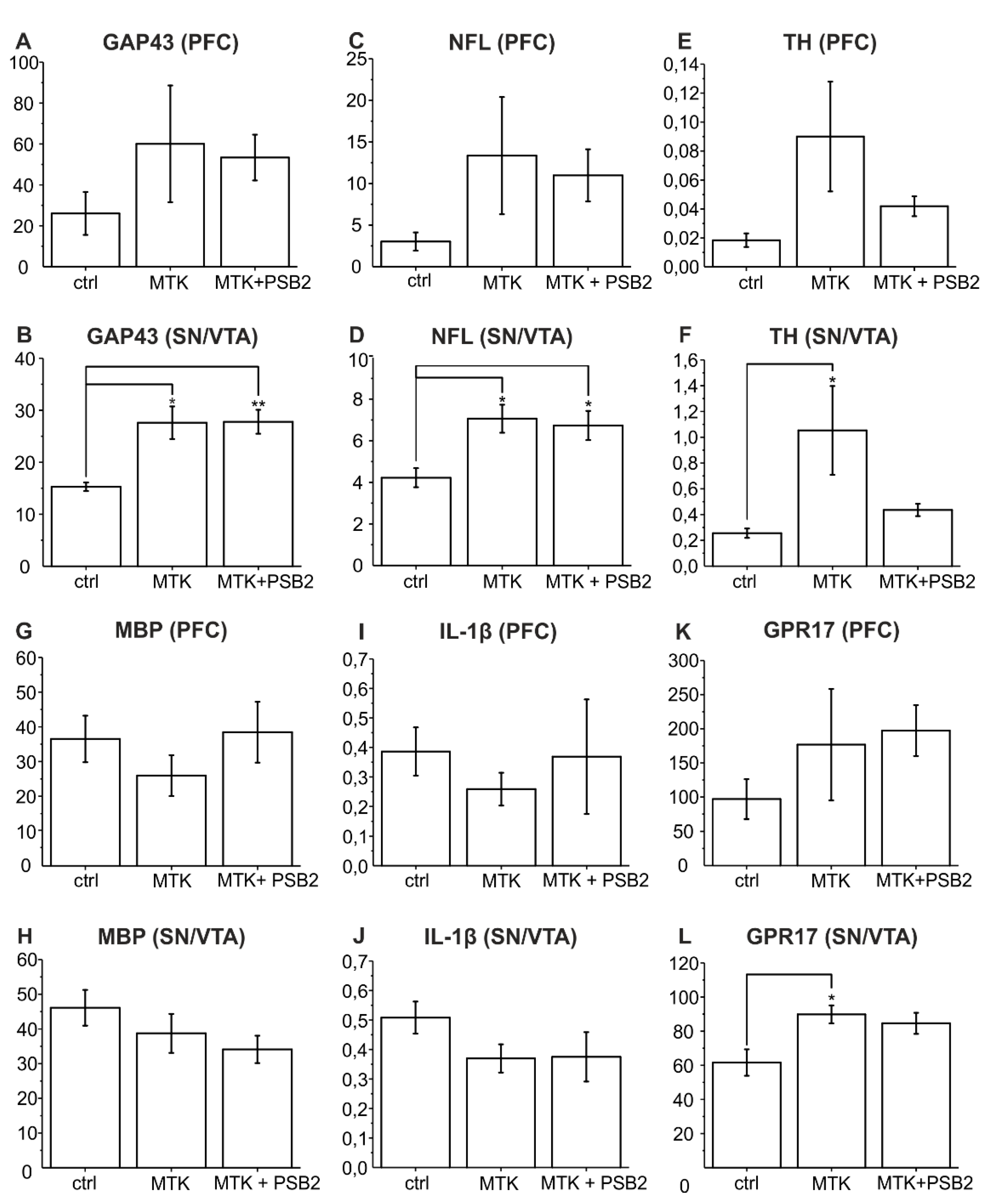
2.1. Neurite Fibre Outgrowth Modulation
2.2. Gene Expression Analysis of Genes Relevant for Neuronal Growth
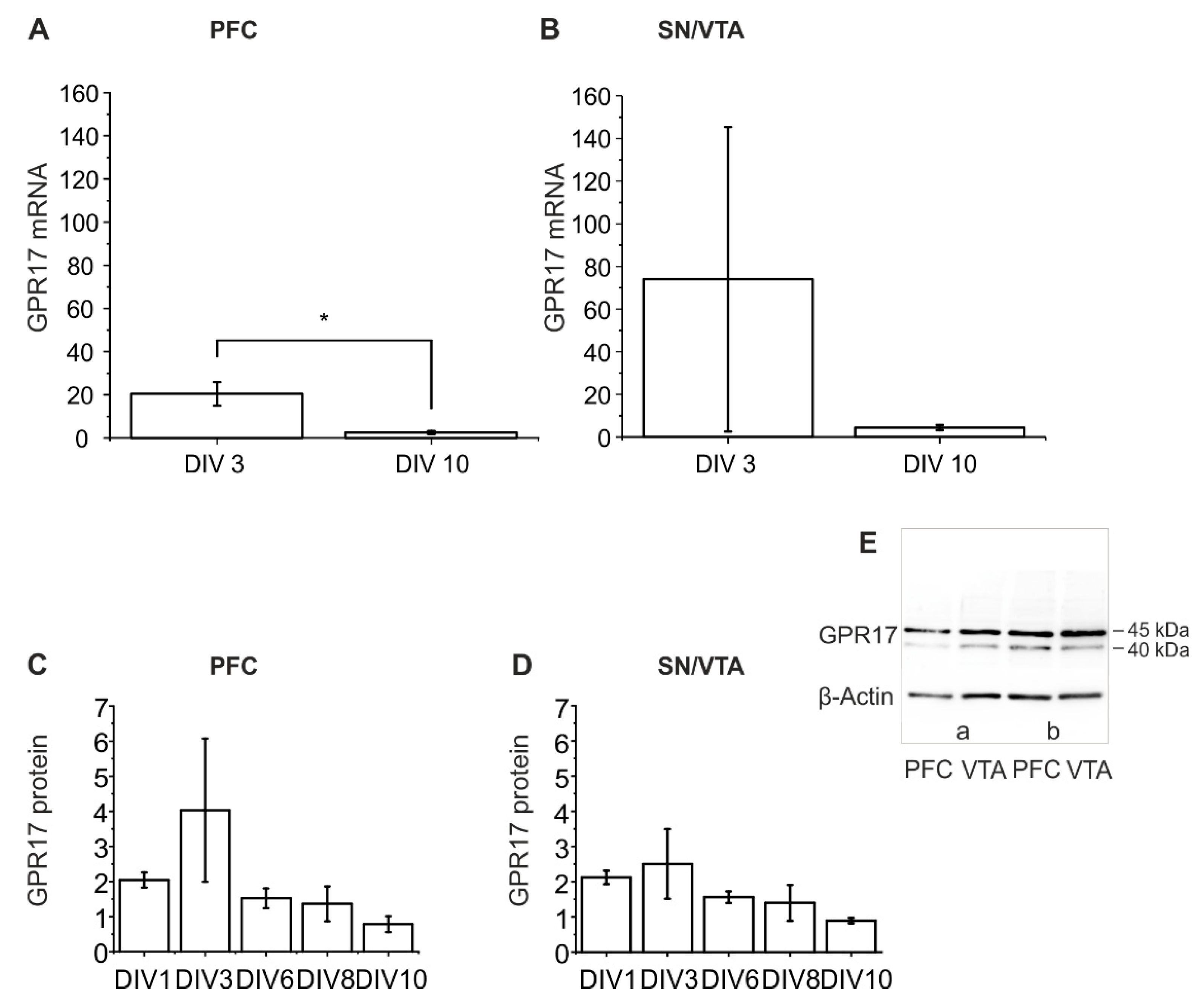
2.3. Dynamics of GPR17 Expression in Untreated Organotypic Dopaminergic Co-Cultures
2.3.1. RT-qPCR Analysis of GPR17 Expression
2.3.2. Western Blot Analysis
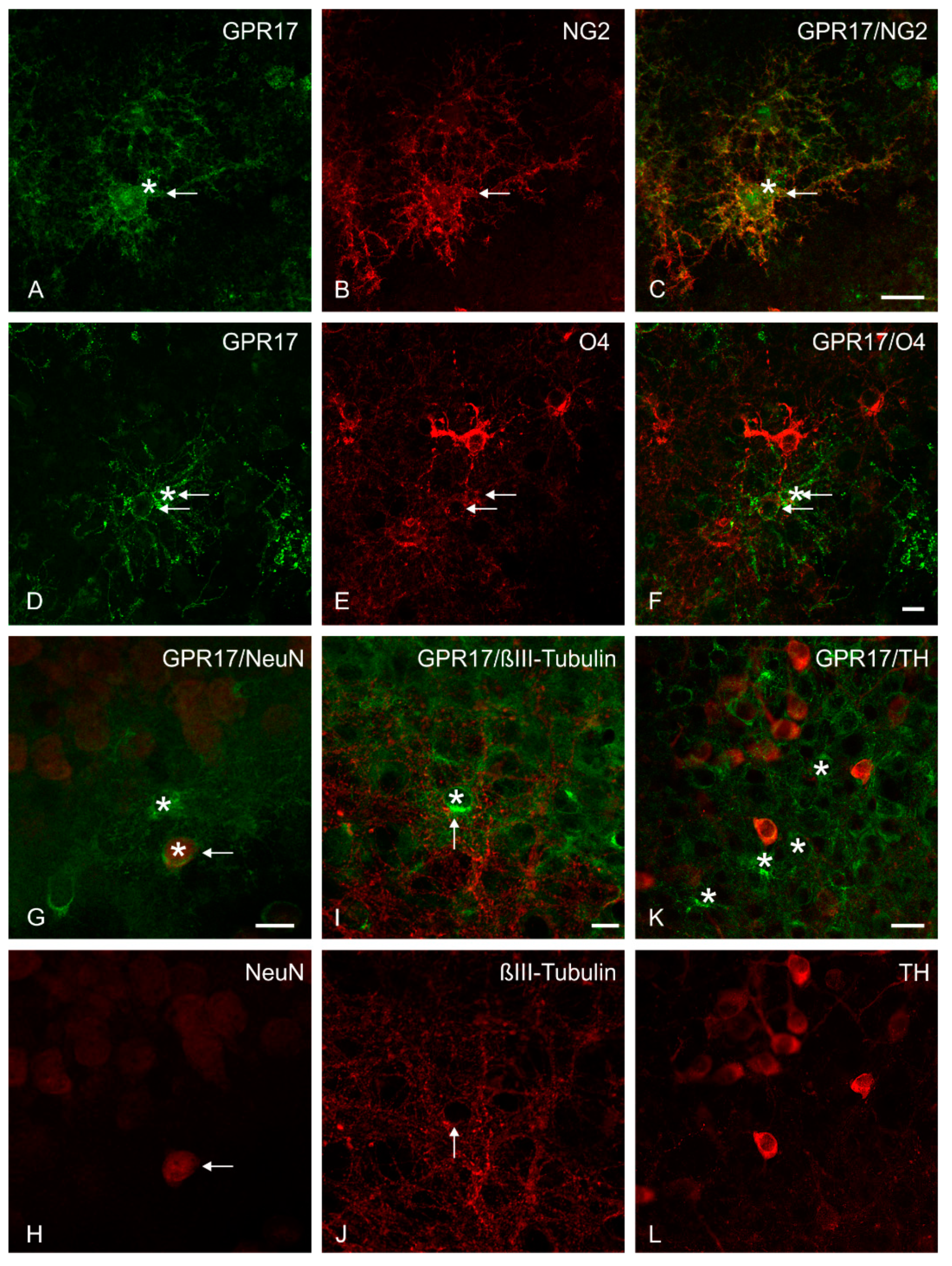
2.4. Immunohistochemical Analysis of GPR17 Expression
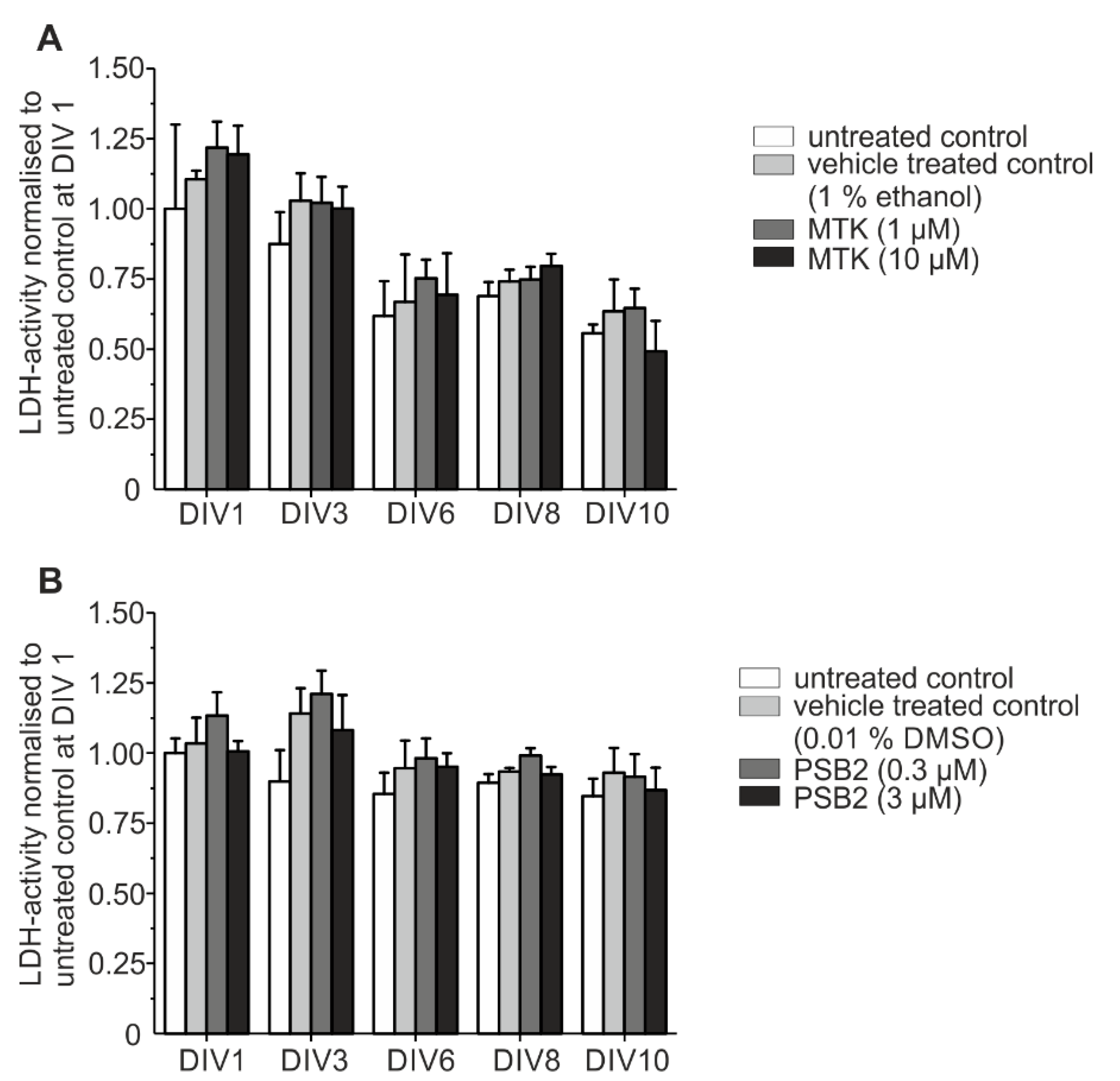
2.5. Toxicological Analysis
2.6. Electron Microscopy
3. Discussion
3.1. Pharmacological Inhibition of the GPR17 with MTK Can Promote Neurite Outgrowth
3.2. MTK Promotes Neurite Outgrowth by Elevating Neurotrophic Gene Expression
3.3. Is Neurite Outgrowth by MTK Stimulated by Modulation of Neuroinflammatory Pathways?
3.4. Is Neurite Outgrowth Stimulated by Targeting Oligodendrocytes in the Co-Culture?
4. Materials and Methods
4.1. Materials
4.2. Animals
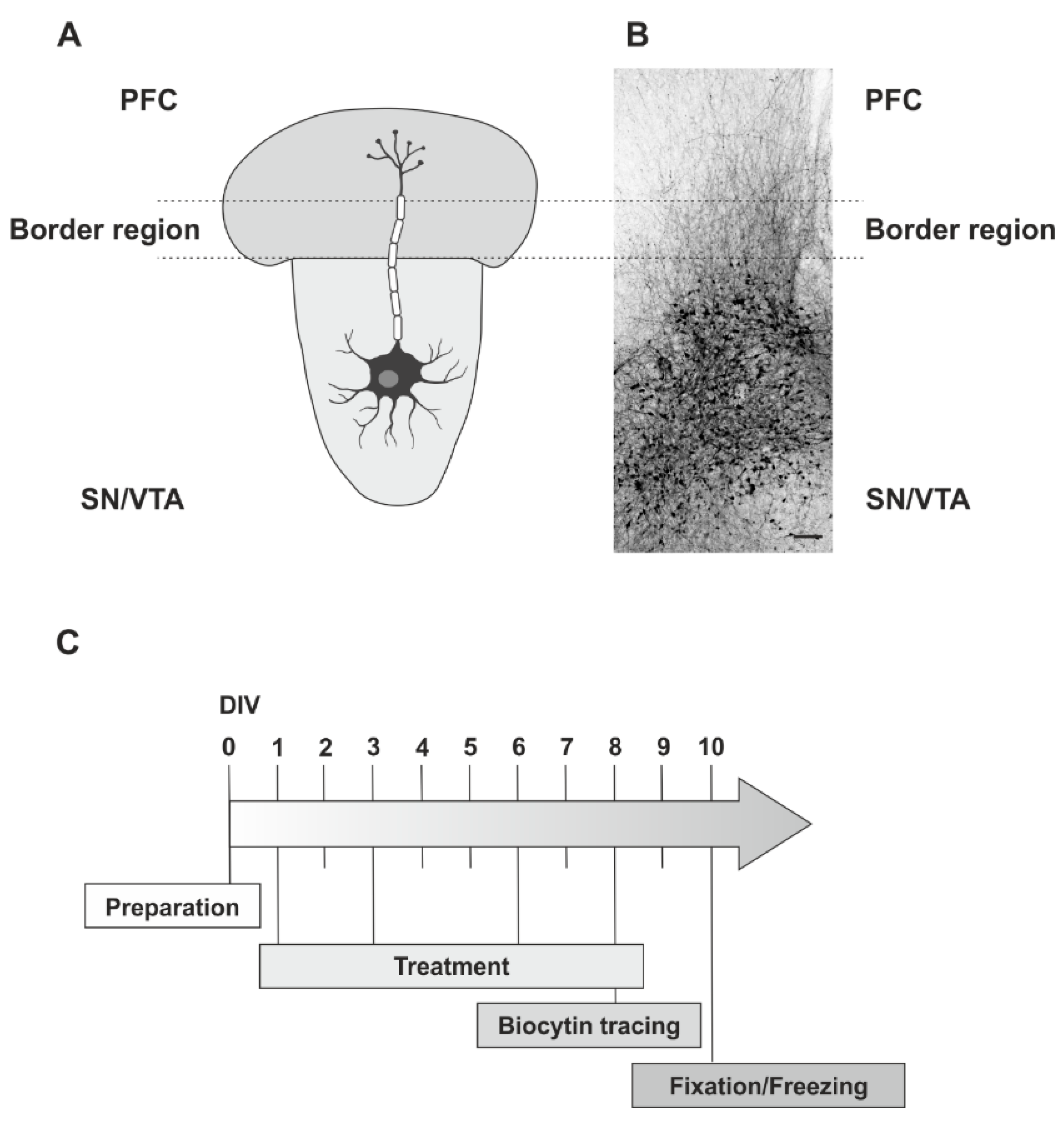
4.3. Preparation
4.4. Slice Co-Culture Treatment Procedure
4.5. Fixation of the Slice Co-Cultures
4.6. Neurite Fibre Tracing Procedure
4.7. Neurite Fibre Density Quantification
4.8. Multiple Immunofluorescence Labelling
4.9. Analysis of mRNA-Expression
4.10. Western Blot
4.11. Analysis of Cell Injury
4.12. Electron Microscopy
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACSF | artificial cerebrospinal fluid |
| CysLT-R | cysteinyl leukotriene receptor |
| GAP43 | growth associated protein 43 |
| GDNF | glial cell line-derived neurotrophic factor |
| GPR17 | G-protein coupled receptor 17 |
| IL-1β | interleukin-1β |
| LDH | lactate dehydrogenase |
| LTC/D4 | Leukotriene C/D4 |
| MBP | myelin basic protein |
| MTK | Montelukast |
| NFL | neurofilament light chain |
| NG2 | neural/glial antigen 2 |
| NeuN | neuronal nuclei antigen |
| OPC | oligodendrocyte precursor cell |
| PFC | prefrontal cortex |
| PSB-16282 | (PSB1) 3-(2-Carboxyethyl)-4-fluoro-6-(5-methylhexyloxy)-1H-indole-2-carboxylic acid |
| PSB-16484 | (PSB2) 3-(2-Carboxyethyl)-4-fluoro-6-iodo-1H-indole-2-carboxylic acid |
| RT-qPCR | reverse transcription-quantitative real-time PCR |
| SDS | sodium dodecyl sulfate |
| SN/VTA | substantia nigra/ventral tegmental area |
| TH | tyrosine hydroxylase |
References
- Feigin, V.L.; Abajobir, A.A.; Abate, K.H. Global, regional, and national burden of neurological disorders during 1990–2015: A systematic analysis for the global burden of disease study 2015. Lancet Neurol. 2017, 16, 877–897. [Google Scholar] [CrossRef] [Green Version]
- James, S.L.; Theadom, A.; Ellenbogen, R.G.; Bannick, M.S.; Montjoy-Venning, W.; Lucchesi, L.R.; Abbasi, N.; Abdulkader, R.; Abraha, H.N.; Adsuar, J.C.; et al. Global, regional, and national burden of traumatic brain injury and spinal cord injury, 1990--2016: A systematic analysis for the global burden of disease study 2016. Lancet Neurol. 2019, 18, 56–87. [Google Scholar] [CrossRef] [Green Version]
- Dorsey, E.R.; Elbaz, A.; Nichols, E.; Abd-Allah, F.; Abdelalim, A.; Adsuar, J.C.; Ansha, M.G.; Brayne, C.; Choi, J.-Y.J.; Collado-Mateo, D.; et al. Global, regional, and national burden of parkinson’s disease, 1990--2016: A systematic analysis for the global burden of disease study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Nichols, E.; Szoeke, C.E.I.; Vollset, S.E.; Abbasi, N.; Abd-Allah, F.; Abdela, J.; Aichour, M.T.E.; Akinyemi, R.O.; Alahdab, F.; Asgedom, S.W.; et al. Global, regional, and national burden of alzheimer’s disease and other dementias, 1990--2016: A systematic analysis for the global burden of disease study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Olesen, J.; Gustavsson, A.; Svensson, M.; Wittchen, H.U.; Jönsson, B. The economic cost of brain disorders in europe. Eur. J. Neurol. 2012, 19, 155–162. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, H.; Wang, S.; Koito, H.; Li, J.; Ye, F.; Hoang, J.; Escobar, S.S.; Gow, A.; Arnett, H.A.; et al. The oligodendrocyte-specific g protein-coupled receptor gpr17 is a cell-intrinsic timer of myelination. Nat. Neurosci. 2009, 12, 1398–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennen, S.; Wang, H.; Peters, L.; Merten, N.; Simon, K.; Spinrath, A.; Blättermann, S.; Akkari, R.; Schrage, R.; Schröder, R.; et al. Decoding signaling and function of the orphan g protein-coupled receptor gpr17 with a small-molecule agonist. Sci. Signal. 2013, 6, ra93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfanti, E.; Gelosa, P.; Fumagalli, M.; Dimou, L.; Viganò, F.; Tremoli, E.; Cimino, M.; Sironi, L.; Abbracchio, M.P. The role of oligodendrocyte precursor cells expressing the gpr17 receptor in brain remodeling after stroke. Cell Death Dis. 2017, 8, e2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fumagalli, M.; Lecca, D.; Coppolino, G.T.; Parravicini, C.; Abbracchio, M.P. Pharmacological properties and biological functions of the gpr17 receptor, a potential target for neuro-regenerative medicine. Adv. Exp. Med. Biol. 2017, 1051, 169–192. [Google Scholar] [PubMed] [Green Version]
- Seyedsadr, M.S.; Ineichen, B.V. Gpr17, a player in lysolecithin-induced demyelination, oligodendrocyte survival, and differentiation. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 2273–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavi, M.S.; Karimi, G.; Roohbakhsh, A. The role of orphan g protein-coupled receptors in the pathophysiology of multiple sclerosis: A review. Life Sci. 2019, 224, 33–40. [Google Scholar] [CrossRef]
- Bonfanti, E.; Bonifacino, T.; Raffaele, S.; Milanese, M.; Morgante, E.; Bonanno, G.; Abbracchio, M.P.; Fumagalli, M. Abnormal upregulation of gpr17 receptor contributes to oligodendrocyte dysfunction in sod1 g93a mice. Int. J. Mol. Sci. 2020, 21, 2395. [Google Scholar] [CrossRef] [Green Version]
- Lecca, D.; Trincavelli, M.L.; Gelosa, P.; Sironi, L.; Ciana, P.; Fumagalli, M.; Villa, G.; Verderio, C.; Grumelli, C.; Guerrini, U.; et al. The recently identified p2y-like receptor gpr17 is a sensor of brain damage and a new target for brain repair. PLoS ONE 2008, 3, e3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viganò, F.; Dimou, L. The heterogeneous nature of ng2-glia. Brain Res. 2016, 1638, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, H.; Parravicini, C.; Lecca, D.; Zanier, E.R.; Heine, C.; Bremicker, K.; Fumagalli, M.; Rosa, P.; Longhi, L.; Stocchetti, N.; et al. Changes of the gpr17 receptor, a new target for neurorepair, in neurons and glial cells in patients with traumatic brain injury. Purinergic Signal. 2013, 9, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Viganò, F.; Schneider, S.; Cimino, M.; Bonfanti, E.; Gelosa, P.; Sironi, L.; Abbracchio, M.P.; Dimou, L. Gpr17 expressing ng2-glia: Oligodendrocyte progenitors serving as a reserve pool after injury. Glia 2016, 64, 287–299. [Google Scholar] [CrossRef]
- Coppolino, G.T.; Marangon, D.; Negri, C.; Menichetti, G.; Fumagalli, M.; Gelosa, P.; Dimou, L.; Furlan, R.; Lecca, D.; Abbracchio, M.P. Differential local tissue permissiveness influences the final fate of gpr17-expressing oligodendrocyte precursors in two distinct models of demyelination. Glia 2018, 66, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Lecca, D.; Raffaele, S.; Abbracchio, M.P.; Fumagalli, M. Regulation and signaling of the gpr17 receptor in oligodendroglial cells. Glia 2020, 68, 1957–1967. [Google Scholar] [CrossRef]
- Benned-Jensen, T.; Rosenkilde, M.M. Distinct expression and ligand-binding profiles of two constitutively active gpr17 splice variants. Br. J. Pharmacol. 2010, 159, 1092–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köse, M.; Ritter, K.; Thiemke, K.; Gillard, M.; Kostenis, E.; Müller, C.E. Development of (3)h2-carboxy-4,6-dichloro-1h-indole-3-propionic acid ((3)hpsb-12150): A useful tool for studying gpr17. ACS Med. Chem. Lett. 2014, 5, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Ciana, P.; Fumagalli, M.; Trincavelli, M.L.; Verderio, C.; Rosa, P.; Lecca, D.; Ferrario, S.; Parravicini, C.; Capra, V.; Gelosa, P.; et al. The orphan receptor gpr17 identified as a new dual uracil nucleotides/cysteinyl-leukotrienes receptor. EMBO J. 2006, 25, 4615–4627. [Google Scholar] [CrossRef] [PubMed]
- Qi, A.-D.; Harden, T.K.; Nicholas, R.A. Is gpr17 a p2y/leukotriene receptor? Examination of uracil nucleotides, nucleotide sugars, and cysteinyl leukotrienes as agonists of gpr17. J. Pharmacol. Exp. Ther. 2013, 347, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, K.; Merten, N.; Schröder, R.; Hennen, S.; Preis, P.; Schmitt, N.-K.; Peters, L.; Schrage, R.; Vermeiren, C.; Gillard, M.; et al. The orphan receptor gpr17 is unresponsive to uracil nucleotides and cysteinyl leukotrienes. Mol. Pharmacol. 2017, 91, 518–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceruti, S.; Viganò, F.; Boda, E.; Ferrario, S.; Magni, G.; Boccazzi, M.; Rosa, P.; Buffo, A.; Abbracchio, M.P. Expression of the new p2y-like receptor gpr17 during oligodendrocyte precursor cell maturation regulates sensitivity to atp-induced death. Glia 2011, 59, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, S.; Villa, G.; Genovese, T.; Mazzon, E.; Longhi, R.; Rosa, P.; Bramanti, P.; Cuzzocrea, S.; Abbracchio, M.P. The p2y-like receptor gpr17 as a sensor of damage and a new potential target in spinal cord injury. Brain J. Neurol. 2009, 132, 2206–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marschallinger, J.; Schäffner, I.; Klein, B.; Gelfert, R.; Rivera, F.J.; Illes, S.; Grassner, L.; Janssen, M.; Rotheneichner, P.; Schmuckermair, C.; et al. Structural and functional rejuvenation of the aged brain by an approved anti-asthmatic drug. Nat. Commun. 2015, 6, 8466. [Google Scholar] [CrossRef] [Green Version]
- Daniele, S.; Lecca, D.; Trincavelli, M.L.; Ciampi, O.; Abbracchio, M.P.; Martini, C. Regulation of pc12 cell survival and differentiation by the new p2y-like receptor gpr17. Cell. Signal. 2010, 22, 697–706. [Google Scholar] [CrossRef]
- Mao, F.-X.; Li, W.-J.; Chen, H.-J.; Qian, L.-H.; Buzby, J.S. Periventricular leukomalacia long-term prognosis may be improved by treatment with udp-glucose, gdnf, and memantine in neonatal rats. Brain Res. 2012, 1486, 112–120. [Google Scholar] [CrossRef]
- Franke, H.; Schelhorn, N.; Illes, P. Dopaminergic neurons develop axonal projections to their target areas in organotypic co-cultures of the ventral mesencephalon and the striatum/prefrontal cortex. Neurochem. Int. 2003, 42, 431–439. [Google Scholar] [CrossRef]
- Heine, C.; Sygnecka, K.; Franke, H. Purines in neurite growth and astroglia activation. Neuropharmacology 2016, 104, 255–271. [Google Scholar] [CrossRef]
- Cho, S.; Wood, A.; Bowlby, M.R. Brain slices as models for neurodegenerative disease and screening platforms to identify novel therapeutics. Curr. Neuropharmacol. 2007, 5, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Humpel, C. Organotypic brain slice cultures: A review. Neuroscience 2015, 305, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Heine, C.; Sygnecka, K.; Scherf, N.; Grohmann, M.; Bräsigk, A.; Franke, H. P2y(1) receptor mediated neuronal fibre outgrowth in organotypic brain slice co-cultures. Neuropharmacology 2015, 93, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Baqi, Y.; Pillaiyar, T.; Abdelrahman, A.; Kaufmann, O.; Alshaibani, S.; Rafehi, M.; Ghasimi, S.; Akkari, R.; Ritter, K.; Simon, K.; et al. 3-(2-carboxyethyl)indole-2-carboxylic acid derivatives: Structural requirements and properties of potent agonists of the orphan g protein-coupled receptor gpr17. J. Med. Chem. 2018, 61, 8136–8154. [Google Scholar] [CrossRef]
- Baqi, Y.; Alshaibani, S.; Ritter, K.; Abdelrahman, A.; Spinrath, A.; Kostenis, E.; Müller, C.E. Improved synthesis of 4-/6-substituted 2-carboxy-1h-indole-3-propionic acid derivatives and structure--activity relationships as gpr17 agonists. Med. Chem. Commun. 2014, 5, 86–92. [Google Scholar] [CrossRef]
- af Bjerkén, S.; Boger, H.A.; Nelson, M.; Hoffer, B.J.; Granholm, A.-C.; Strömberg, I. Effects of glial cell line-derived neurotrophic factor deletion on ventral mesencephalic organotypic tissue cultures. Brain Res. 2007, 1133, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wu, M.; Zhan, C.; Ma, E.; Yang, M.; Yang, X.; Li, Y. Neurofilament proteins in axonal regeneration and neurodegenerative diseases. Neural Regen. Res. 2012, 7, 620–626. [Google Scholar]
- Ma, T.C.; Willis, D.E. What makes a rag regeneration associated? Front. Mol. Neurosci. 2015, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benowitz, L.I.; Routtenberg, A. Gap-43: An intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997, 20, 84–91. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.V.; Nixon, R.A. Neurofilaments at a glance. J. Cell Sci. 2012, 125, 3257–3263. [Google Scholar] [CrossRef] [PubMed]
- Bademci, G.; Vance, J.M.; Wang, L. Tyrosine hydroxylase gene: Another piece of the genetic puzzle of parkinson’s disease. CNS Neurol. Disord. Drug Targets 2012, 11, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Lecca, D.; Abbracchio, M.P. Cns remyelination as a novel reparative approach to neurodegenerative diseases: The roles of purinergic signaling and the p2y-like receptor gpr17. Neuropharmacology 2016, 104, 82–93. [Google Scholar] [CrossRef]
- Ou, Z.; Sun, Y.; Lin, L.; You, N.; Liu, X.; Li, H.; Ma, Y.; Cao, L.; Han, Y.; Liu, M.; et al. Olig2-targeted g-protein-coupled receptor gpr17 regulates oligodendrocyte survival in response to lysolecithin-induced demyelination. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 10560–10573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendiola, A.S.; Cardona, A.E. The il-1\textgreek{b} phenomena in neuroinflammatory diseases. J. Neural Transm. 2018, 125, 781–795. [Google Scholar] [CrossRef]
- Fratangeli, A.; Parmigiani, E.; Fumagalli, M.; Lecca, D.; Benfante, R.; Passafaro, M.; Buffo, A.; Abbracchio, M.P.; Rosa, P. The regulated expression, intracellular trafficking, and membrane recycling of the p2y-like receptor gpr17 in oli-neu oligodendroglial cells. J. Biol. Chem. 2013, 288, 5241–5256. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, M.; Daniele, S.; Lecca, D.; Lee, P.R.; Parravicini, C.; Fields, R.D.; Rosa, P.; Antonucci, F.; Verderio, C.; Trincavelli, M.L.; et al. Phenotypic changes, signaling pathway, and functional correlates of gpr17-expressing neural precursor cells during oligodendrocyte differentiation. J. Biol. Chem. 2011, 286, 10593–10604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sygnecka, K.; Heine, C.; Scherf, N.; Fasold, M.; Binder, H.; Scheller, C.; Franke, H. Nimodipine enhances neurite outgrowth in dopaminergic brain slice co-cultures. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2015, 40, 1–11. [Google Scholar] [CrossRef]
- Boccazzi, M.; Lecca, D.; Marangon, D.; Guagnini, F.; Abbracchio, M.P.; Ceruti, S. A new role for the p2y-like gpr17 receptor in the modulation of multipotency of oligodendrocyte precursor cells in vitro. Purinergic Signal. 2016, 12, 661–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gachet, C. P2y(12) receptors in platelets and other hematopoietic and non-hematopoietic cells. Purinergic Signal. 2012, 8, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Tsuda, M.; Tozaki-Saitoh, H.; Inoue, K.; Kiyama, H. Nerve injury-activated microglia engulf myelinated axons in a p2y12 signaling-dependent manner in the dorsal horn. Glia 2010, 58, 1838–1846. [Google Scholar] [CrossRef]
- Zhan, T.-W.; Tian, Y.-X.; Wang, Q.; Wu, Z.-X.; Zhang, W.-P.; Lu, Y.-B.; Wu, M. Cangrelor alleviates pulmonary fibrosis by inhibiting gpr17-mediated inflammation in mice. Int. Immunopharmacol. 2018, 62, 261–269. [Google Scholar] [CrossRef]
- Maekawa, A.; Balestrieri, B.; Austen, K.F.; Kanaoka, Y. Gpr17 is a negative regulator of the cysteinyl leukotriene 1 receptor response to leukotriene d4. Proc. Natl. Acad. Sci. USA 2009, 106, 11685–11690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisel, M.; Herr, A.; Milosevic, J.; Hermann, A.; Habisch, H.-J.; Schwarz, S.; Kirsch, M.; Antoniadis, G.; Brenner, R.; Hallmeyer-Elgner, S.; et al. Transcription profiling of adult and fetal human neuroprogenitors identifies divergent paths to maintain the neuroprogenitor cell state. Stem Cells 2007, 25, 1231–1240. [Google Scholar] [CrossRef]
- Huber, C.; Marschallinger, J.; Tempfer, H.; Furtner, T.; Couillard-Despres, S.; Bauer, H.-C.; Rivera, F.J.; Aigner, L. Inhibition of leukotriene receptors boosts neural progenitor proliferation. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2011, 28, 793–804. [Google Scholar] [CrossRef]
- Kabba, J.A.; Xu, Y.; Christian, H.; Ruan, W.; Chenai, K.; Xiang, Y.; Zhang, L.; Saavedra, J.M.; Pang, T. Microglia: Housekeeper of the central nervous system. Cell. Mol. Neurobiol. 2017, 38, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Podbielska, M.; Das, A.; Smith, A.W.; Chauhan, A.; Ray, S.K.; Inoue, J.; Azuma, M.; Nozaki, K.; Hogan, E.L.; Banik, N.L. Neuron-microglia interaction induced bi-directional cytotoxicity associated with calpain activation. J. Neurochem. 2016, 139, 440–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Wang, H.; Li, C.-X.; Song, S.-W.; Fang, S.-H.; Wei, E.-Q.; Shi, Q.-J. Gpr17 mediates ischemia-like neuronal injury via microglial activation. Int. J. Mol. Med. 2018, 42, 2750–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; He, X.; Meng, H.; Li, Y.; Dmitriev, P.; Tian, F.; Page, J.C.; Lu, Q.R.; He, Z. Robust myelination of regenerated axons induced by combined manipulations of gpr17 and microglia. Neuron 2020, 108, 876–886.e874. [Google Scholar] [CrossRef]
- Nave, K.-A. Myelination and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.-W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Fumagalli, M.; Bonfanti, E.; Daniele, S.; Zappelli, E.; Lecca, D.; Martini, C.; Trincavelli, M.L.; Abbracchio, M.P. The ubiquitin ligase mdm2 controls oligodendrocyte maturation by intertwining mtor with g protein-coupled receptor kinase 2 in the regulation of gpr17 receptor desensitization. Glia 2015, 63, 2327–2339. [Google Scholar] [CrossRef]
- Simon, K.; Hennen, S.; Merten, N.; Blättermann, S.; Gillard, M.; Kostenis, E.; Gomeza, J. The orphan g protein-coupled receptor gpr17 negatively regulates oligodendrocyte differentiation via g\textgreek{a}i/o and its downstream effector molecules. J. Biol. Chem. 2016, 291, 705–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelosa, P.; Bonfanti, E.; Castiglioni, L.; Delgado-Garcia, J.M.; Gruart, A.; Fontana, L.; Gotti, M.; Tremoli, E.; Lecca, D.; Fumagalli, M.; et al. Improvement of fiber connectivity and functional recovery after stroke by montelukast, an available and safe anti-asthmatic drug. Pharmacol. Res. 2019, 142, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Tamura, Y.; Yamato, M.; Kume, S.; Eguchi, A.; Takata, K.; Watanabe, Y.; Kataoka, Y. Ng2 glial cells regulate neuroimmunological responses to maintain neuronal function and survival. Sci. Rep. 2017, 7, 42041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Hawthorne, A.L.; Bai, L.; Miller, R.H.; Silver, J. Adult ng2+ cells are permissive to neurite outgrowth and stabilize sensory axons during macrophage-induced axonal dieback after spinal cord injury. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Boda, E.; Viganò, F.; Rosa, P.; Fumagalli, M.; Labat-Gest, V.; Tempia, F.; Abbracchio, M.P.; Dimou, L.; Buffo, A. The gpr17 receptor in ng2 expressing cells: Focus on in vivo cell maturation and participation in acute trauma and chronic damage. Glia 2011, 59, 1958–1973. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2018, 18, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Buhl, R.; Bals, R.; Baur, X.; Berdel, D.; Criée, C.P.; Gappa, M.; Gillissen, A.; Greulich, T.; Haidl, P.; Hamelmann, E.; et al. S2k-leitlinie zur diagnostik und therapie von patienten mit asthma. Pneumologie 2017, 71, 849–919. [Google Scholar]
- Pillaiyar, T.; Funke, M.; Al-Hroub, H.; Weyler, S.; Ivanova, S.; Schlegel, J.; Abdelrahman, A.; Müller, C.E. Design, synthesis and biological evaluation of suramin-derived dual antagonists of the proinflammatory g protein-coupled receptors p2y2 and gpr17. Eur. J. Med. Chem. 2020, 186, 111789. [Google Scholar] [CrossRef] [PubMed]
- Parravicini, C.; Lecca, D.; Marangon, D.; Coppolino, G.T.; Daniele, S.; Bonfanti, E.; Fumagalli, M.; Raveglia, L.; Martini, C.; Gianazza, E.; et al. Development of the first in vivo gpr17 ligand through an iterative drug discovery pipeline: A novel disease-modifying strategy for multiple sclerosis. PLoS ONE 2020, 15, e0231483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, C.; Franke, H. Organotypic slice co-culture systems to study axon regeneration in the dopaminergic system ex vivo. Methods Mol. Biol. 2014, 1162, 97–111. [Google Scholar] [PubMed]
- Hudson, J.; Granholm, A.C.; Gerhardt, G.A.; Henry, M.A.; Hoffman, A.; Biddle, P.; Leela, N.S.; Mackerlova, L.; Lile, J.D.; Collins, F. Glial cell line-derived neurotrophic factor augments midbrain dopaminergic circuits in vivo. Brain Res. Bull. 1995, 36, 425–432. [Google Scholar] [CrossRef]
- Heine, C.; Wegner, A.; Grosche, J.; Allgaier, C.; Illes, P.; Franke, H. P2 receptor expression in the dopaminergic system of the rat brain during development. Neuroscience 2007, 149, 165–181. [Google Scholar] [CrossRef]
- Heine, C.; Sygnecka, K.; Scherf, N.; Berndt, A.; Egerland, U.; Hage, T.; Franke, H. Phosphodiesterase 2 inhibitors promote axonal outgrowth in organotypic slice co-cultures. Neuro-Signals 2013, 21, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.Y.; Choi, D.W. Quantitative determination of glutamate mediated cortical neuronal injury in cell culture by lactate dehydrogenase efflux assay. J. Neurosci. Methods 1987, 20, 83–90. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Target | Forward | Reverse |
|---|---|---|---|
| NM_017195.3 | rat GAP43 | ACCACTGATAACTCGCCGTC | TGGCTTCATCTACAGCTTCTTTCT |
| NM_001071777.1 | rat GPR17 | ACTTGTCCTGTGTGCTGGTC | CCCAAAAGGCCAGTGATTGC |
| NM_031512.2 | rat IL-1β | TAGCAGCTTTCGACAGTGAGG | TCTGGACAGCCCAAGTCAAG |
| NM_001025293.1 | rat MBP | TGTGCCACATGTACAAGGACT | TTCATCTTGGGTCCTCTGCG |
| NM_001106116.1 | rat MrpL32 | TTCCGGACCGCTACATAGGTG | CTAGTGCTGGTGCCCACTGAG |
| NM_031783.2 | rat NFL | GCAGCTTACAGGAAACTCTTGG | ACCTGCGAGCTCTGAGAGTA |
| NM_012740.4 | rat TH | TTCTTGAAGGAGCGGACTGG | TGCATTGAAACACGCGGAAG |
| Marker | GPR17-Co-Expression |
|---|---|
| Iba1 | + |
| GFAP | − |
| TH | − |
| NeuN | + |
| βIII-Tubulin | + |
| NFL | − |
| NG2 | +++ |
| O4 | + |
| CNPase | + |
| MBP | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braune, M.; Scherf, N.; Heine, C.; Sygnecka, K.; Pillaiyar, T.; Parravicini, C.; Heimrich, B.; Abbracchio, M.P.; Müller, C.E.; Franke, H. Involvement of GPR17 in Neuronal Fibre Outgrowth. Int. J. Mol. Sci. 2021, 22, 11683. https://doi.org/10.3390/ijms222111683
Braune M, Scherf N, Heine C, Sygnecka K, Pillaiyar T, Parravicini C, Heimrich B, Abbracchio MP, Müller CE, Franke H. Involvement of GPR17 in Neuronal Fibre Outgrowth. International Journal of Molecular Sciences. 2021; 22(21):11683. https://doi.org/10.3390/ijms222111683
Chicago/Turabian StyleBraune, Max, Nico Scherf, Claudia Heine, Katja Sygnecka, Thanigaimalai Pillaiyar, Chiara Parravicini, Bernd Heimrich, Maria P. Abbracchio, Christa E. Müller, and Heike Franke. 2021. "Involvement of GPR17 in Neuronal Fibre Outgrowth" International Journal of Molecular Sciences 22, no. 21: 11683. https://doi.org/10.3390/ijms222111683
APA StyleBraune, M., Scherf, N., Heine, C., Sygnecka, K., Pillaiyar, T., Parravicini, C., Heimrich, B., Abbracchio, M. P., Müller, C. E., & Franke, H. (2021). Involvement of GPR17 in Neuronal Fibre Outgrowth. International Journal of Molecular Sciences, 22(21), 11683. https://doi.org/10.3390/ijms222111683