
Ezrin Peptide Therapy from HIV to COVID: Inhibition of Inflammation and Amplification of Adaptive Anti-Viral Immunity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Treatment of COVID and Prevention of SARS-CoV-2 Reinfection
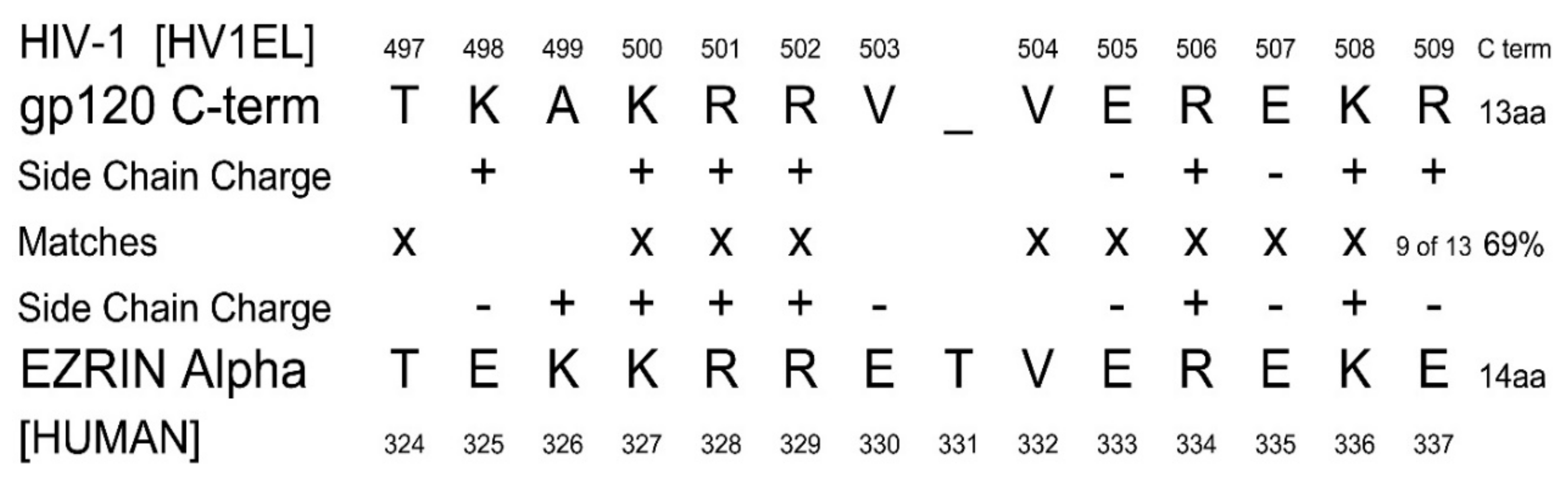
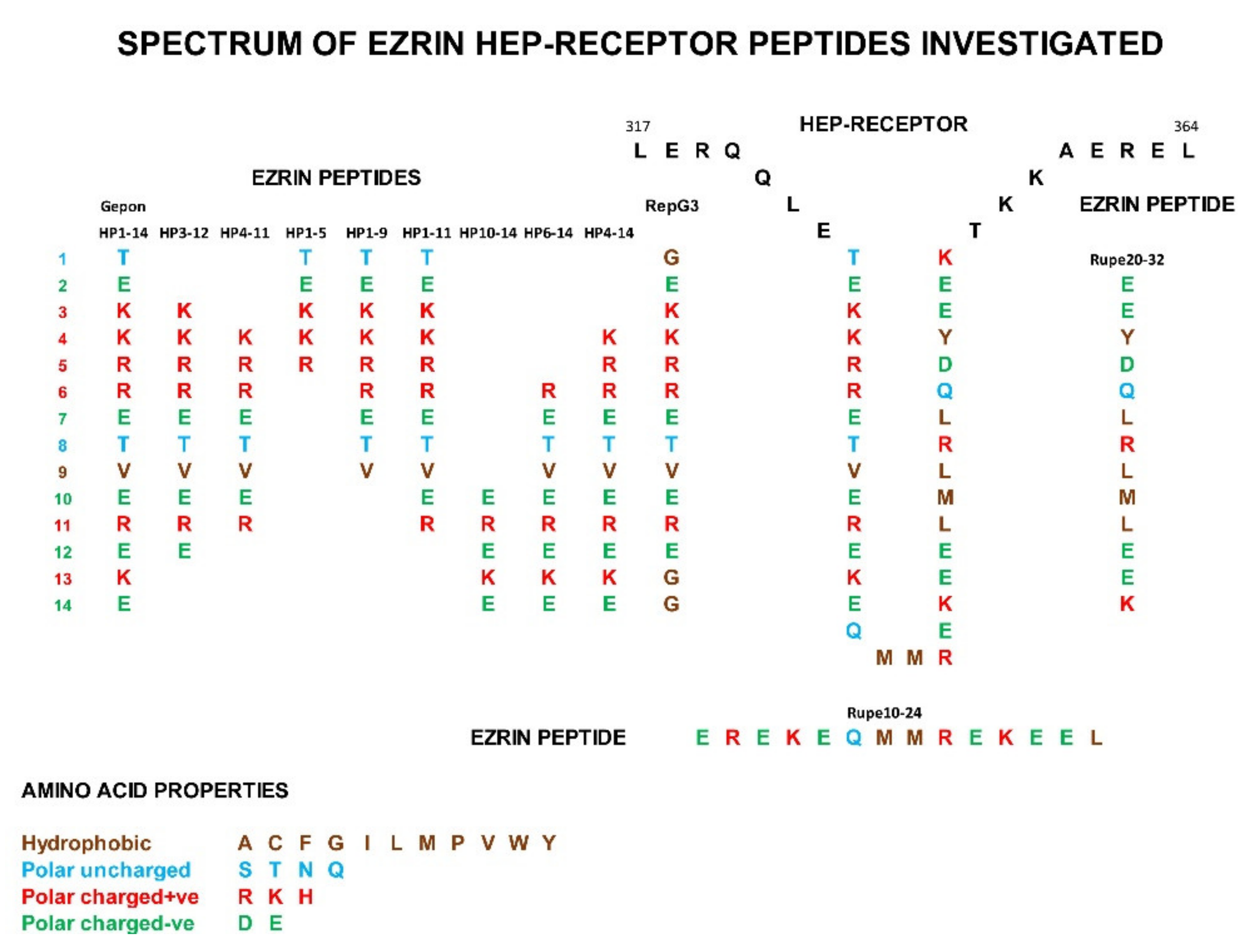
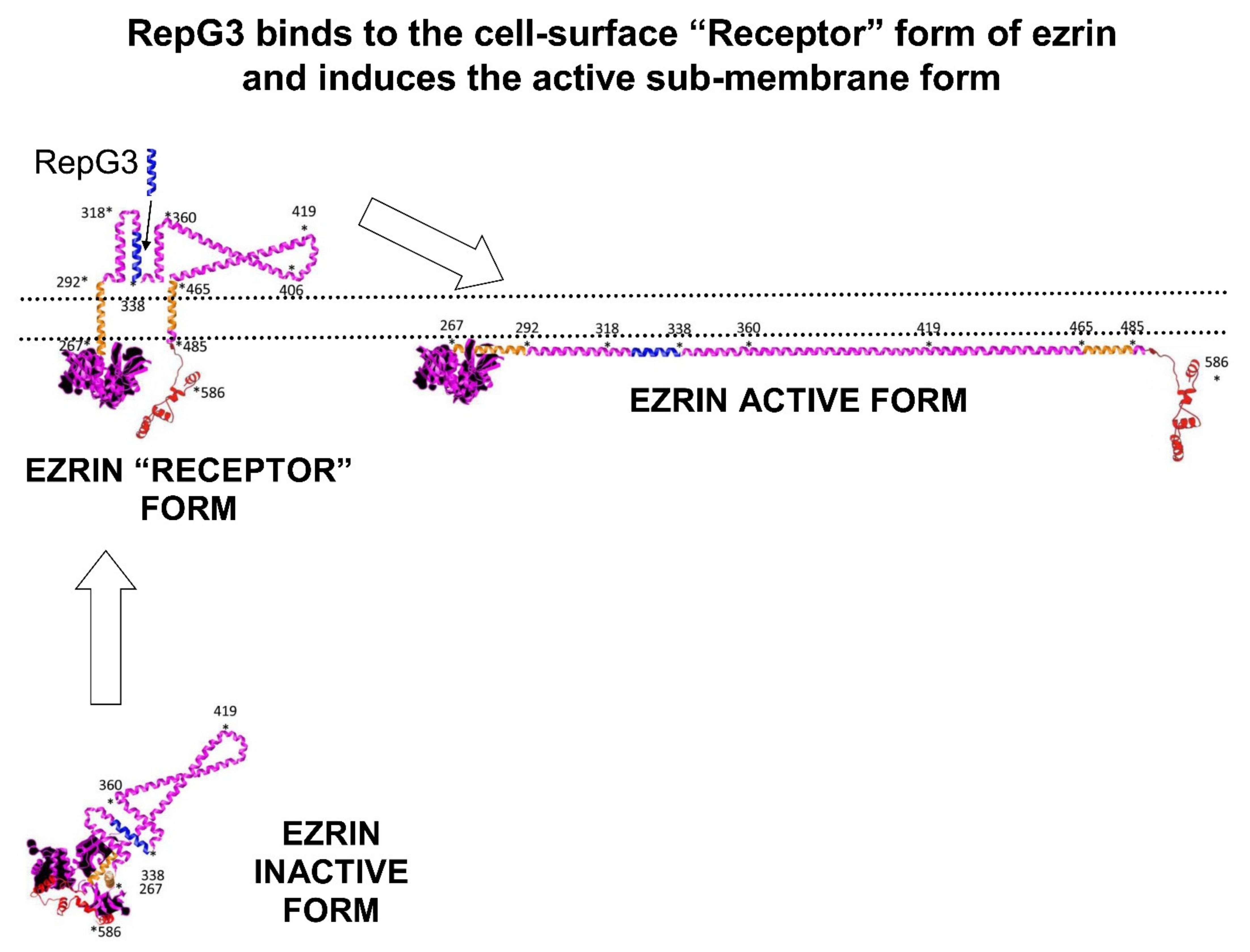
1.2. Human Ezrin Peptides
1.3. Ezrin and the B-Cell Receptor (BCR)
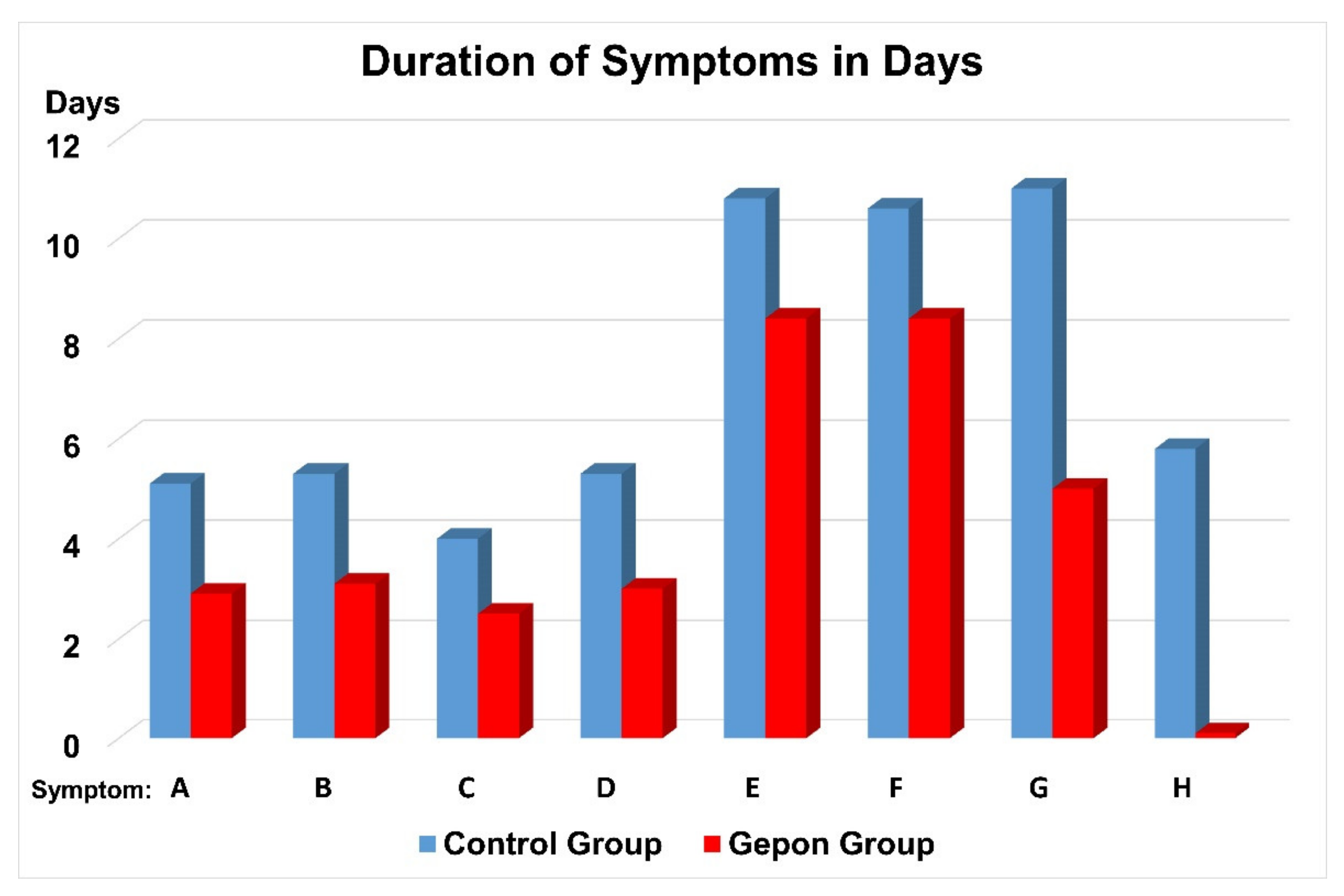
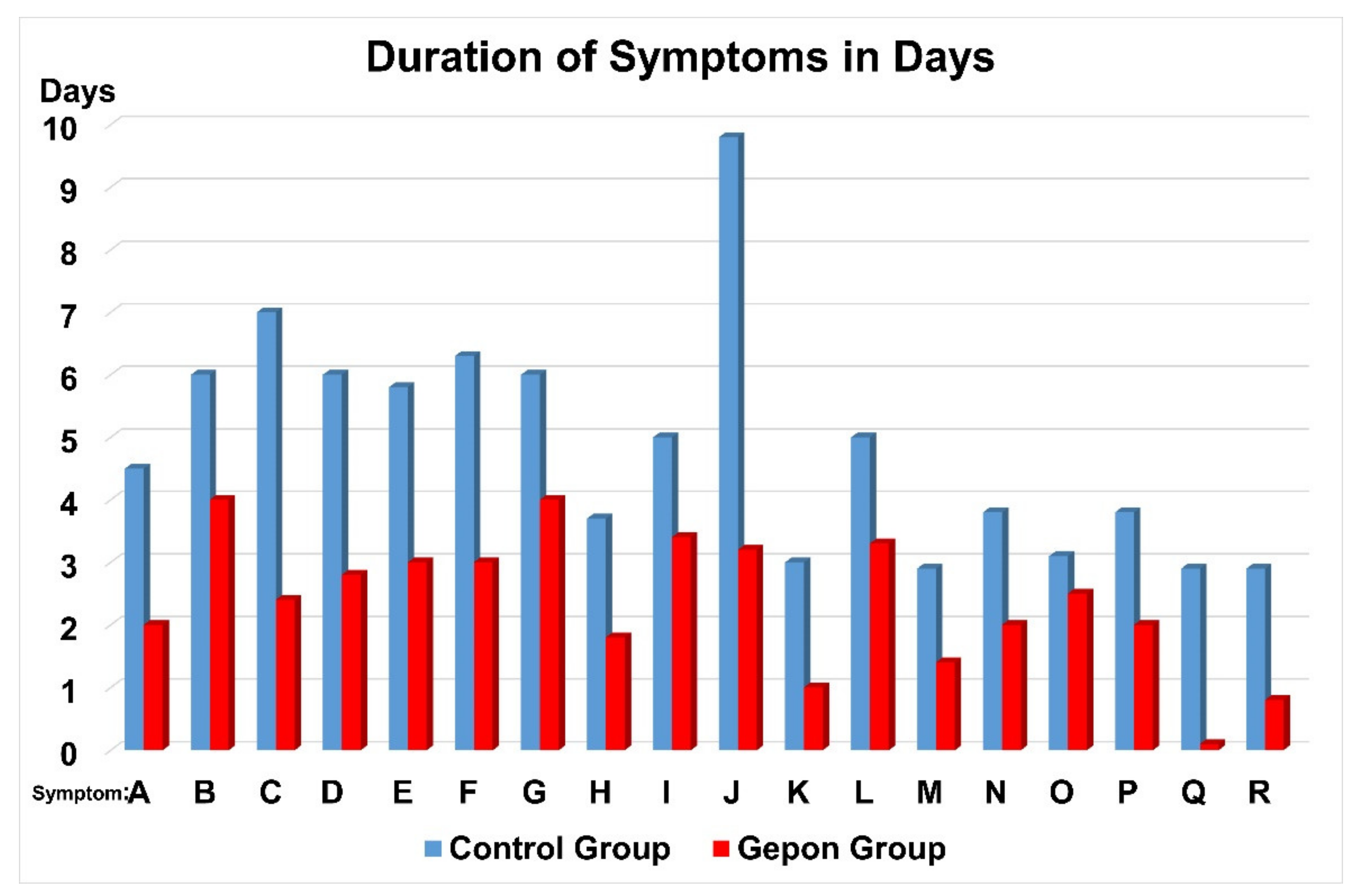
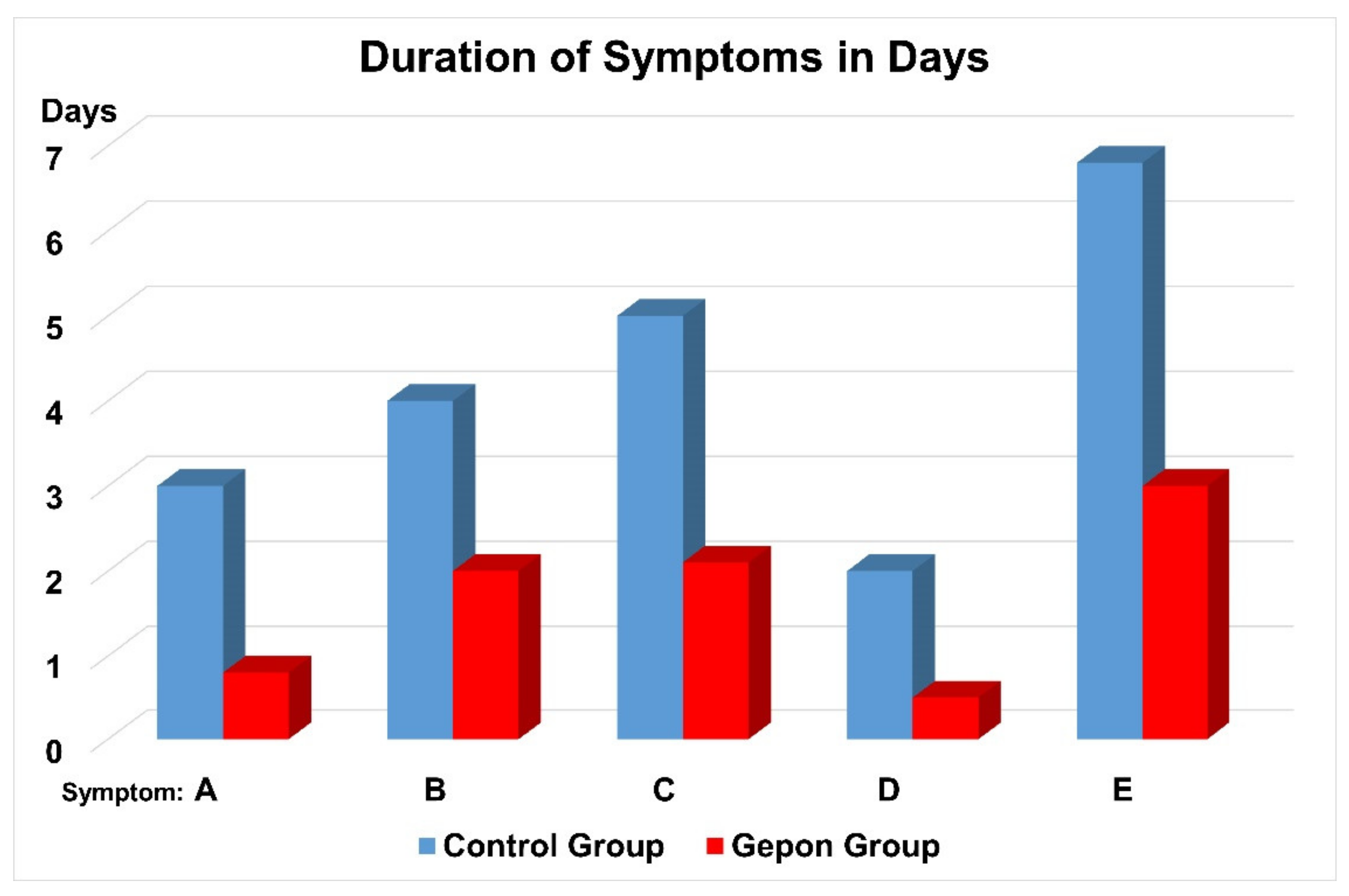
2. Clinical Trials of Ezrin Peptide Treatment of Acute Viral Respiratory Infection (AVRI)
3. Clinical Trials of Ezrin Peptide Treatment of AVRI with Inflammation in Children
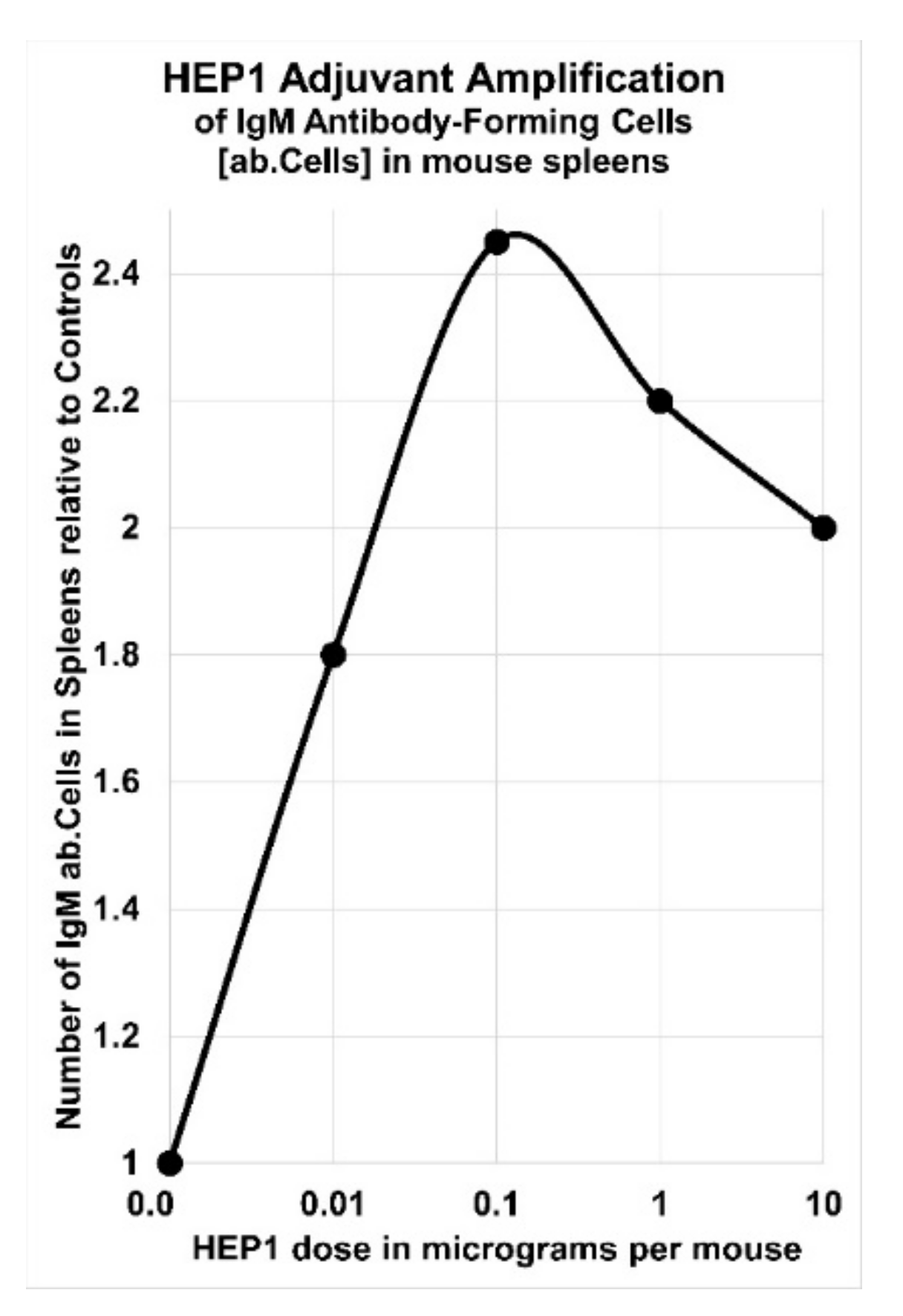
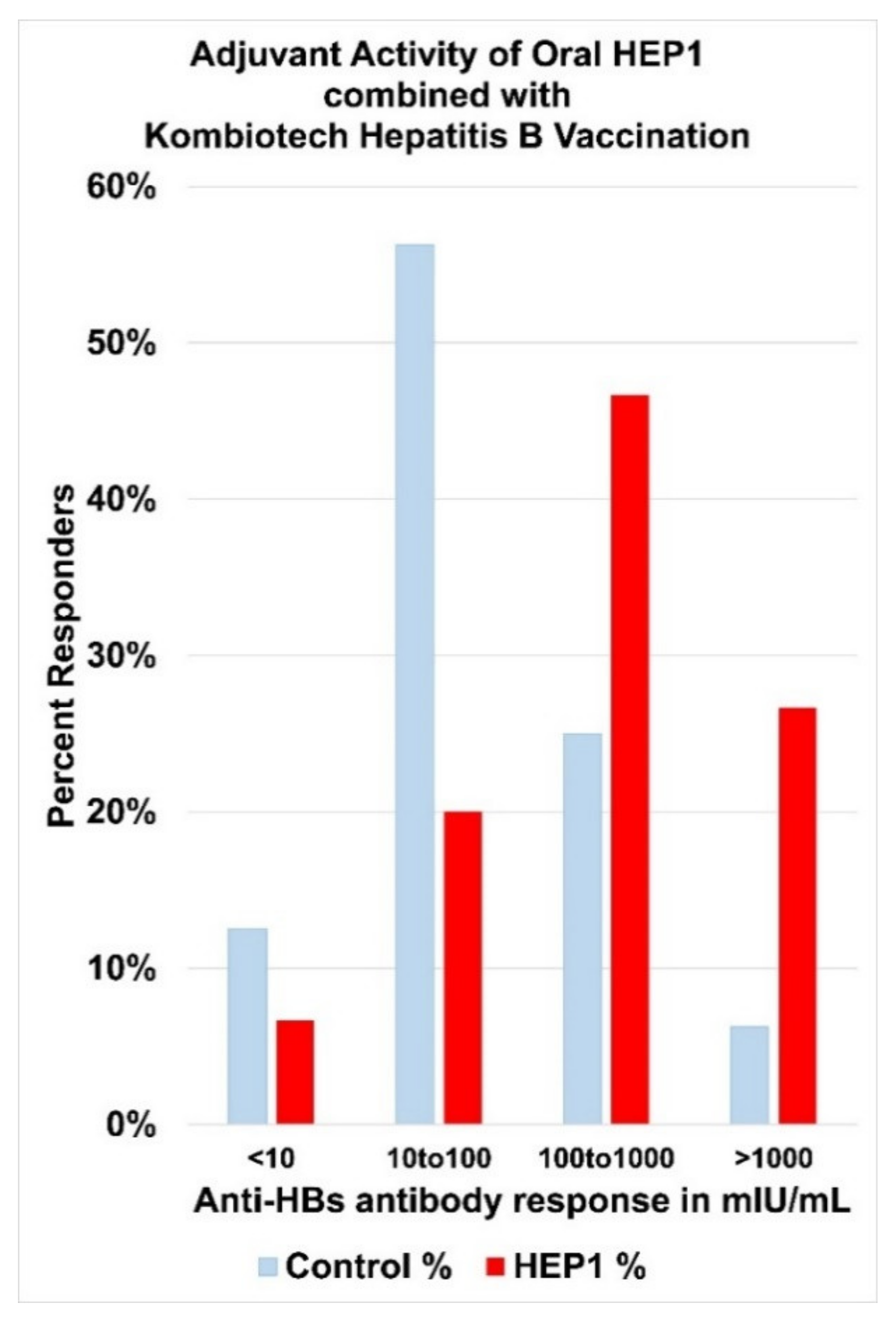
4. Human Ezrin Peptides Enhance Antibody Formation in Mice and Humans
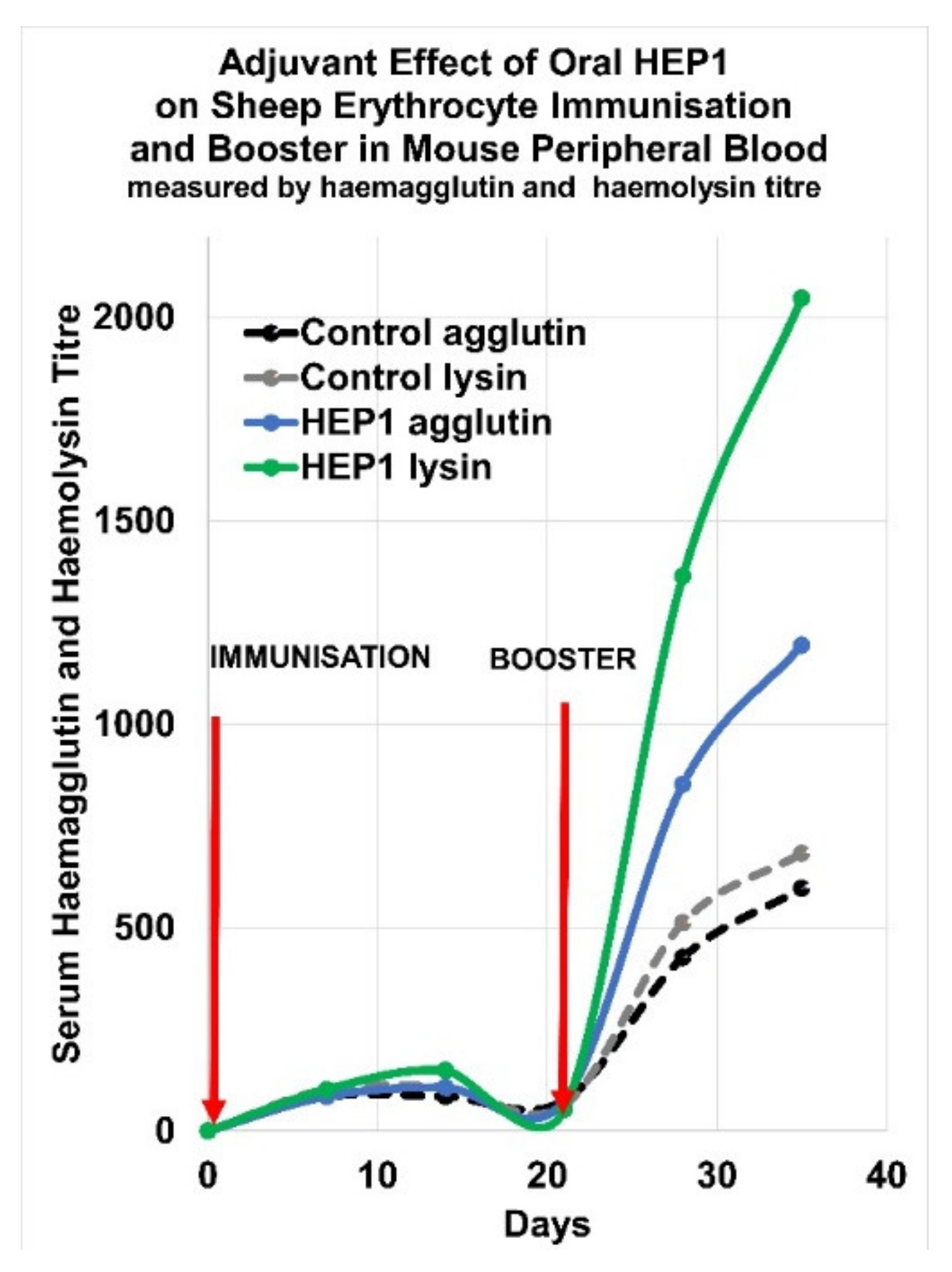
5. Adjuvant Effect of HEP1 on the Production of IgM Antibodies to a Cellular Antigen
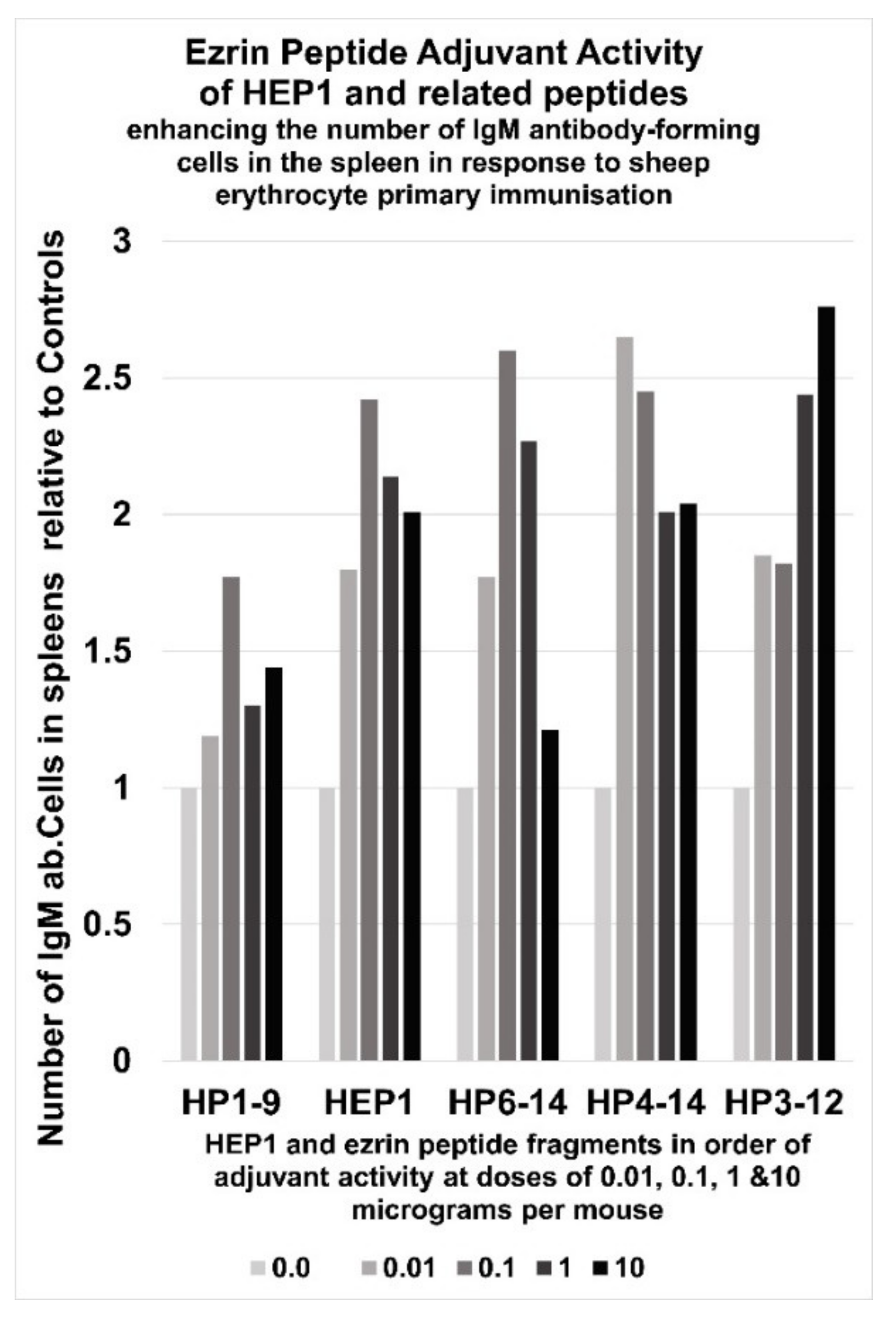
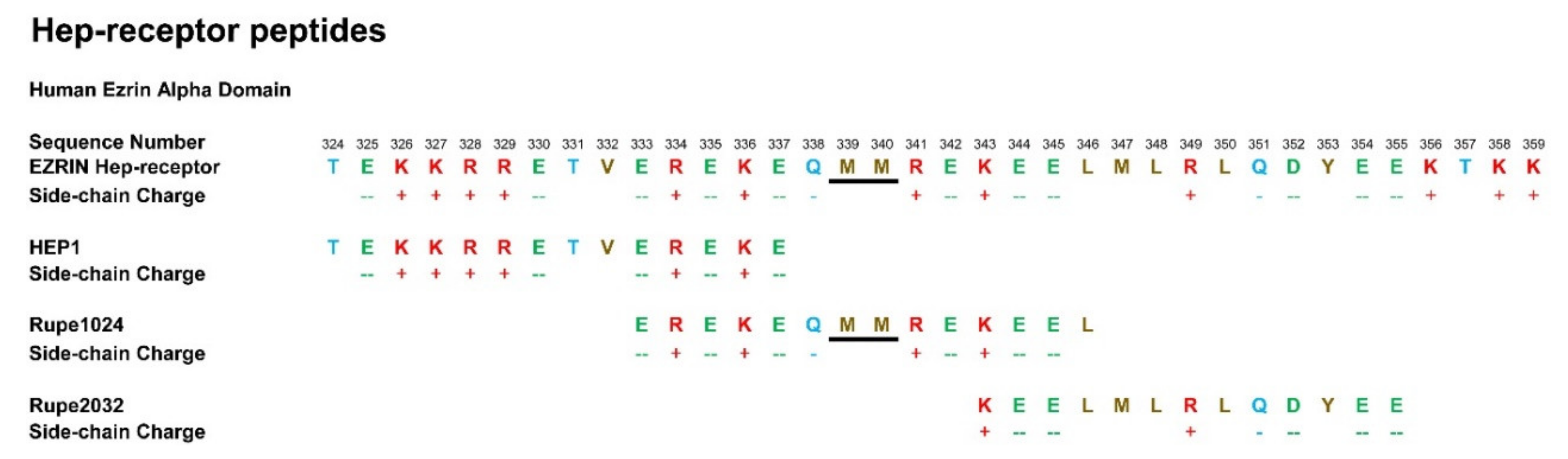
6. Immunoadjuvant Effect of Homologue Peptides of HEP1
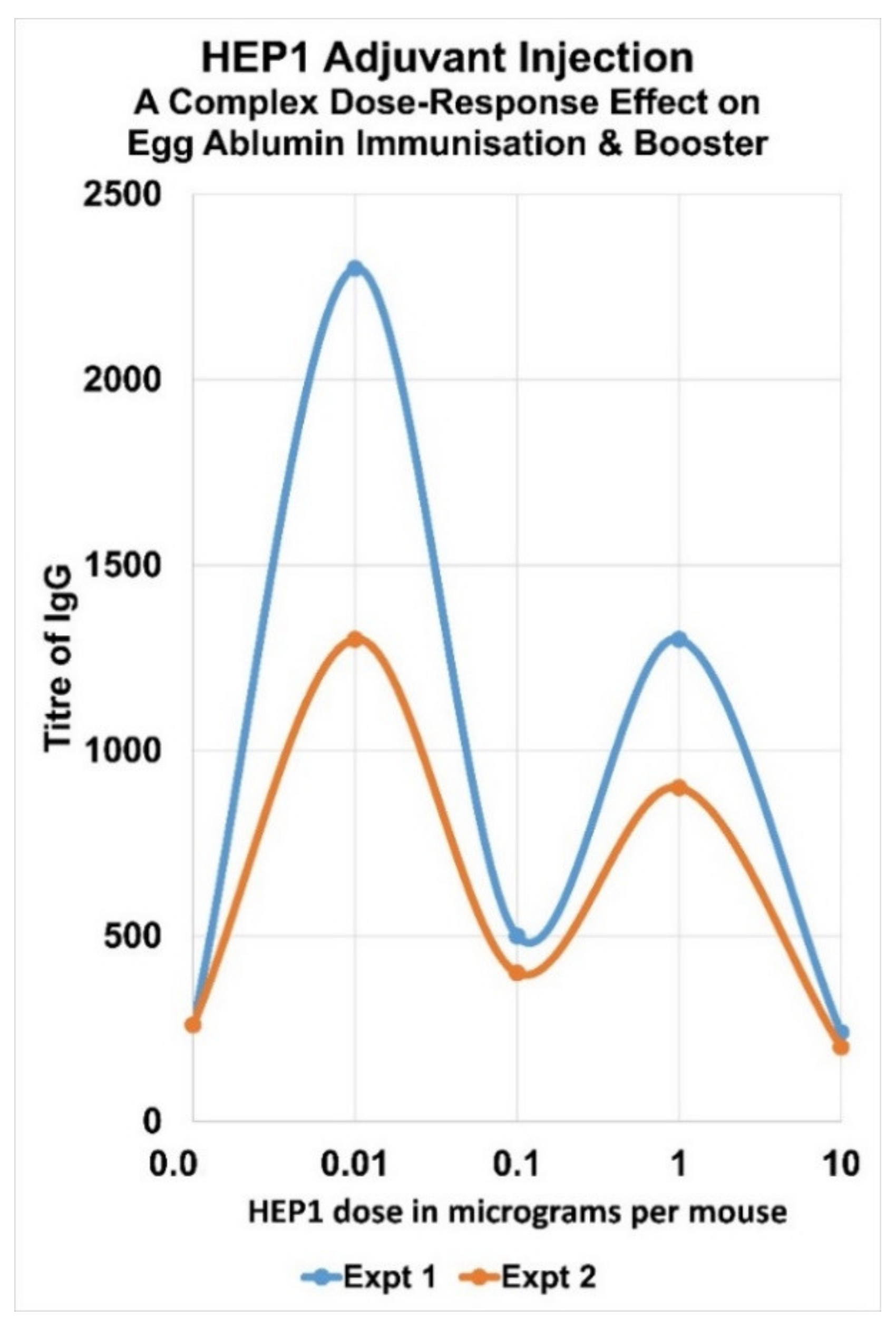
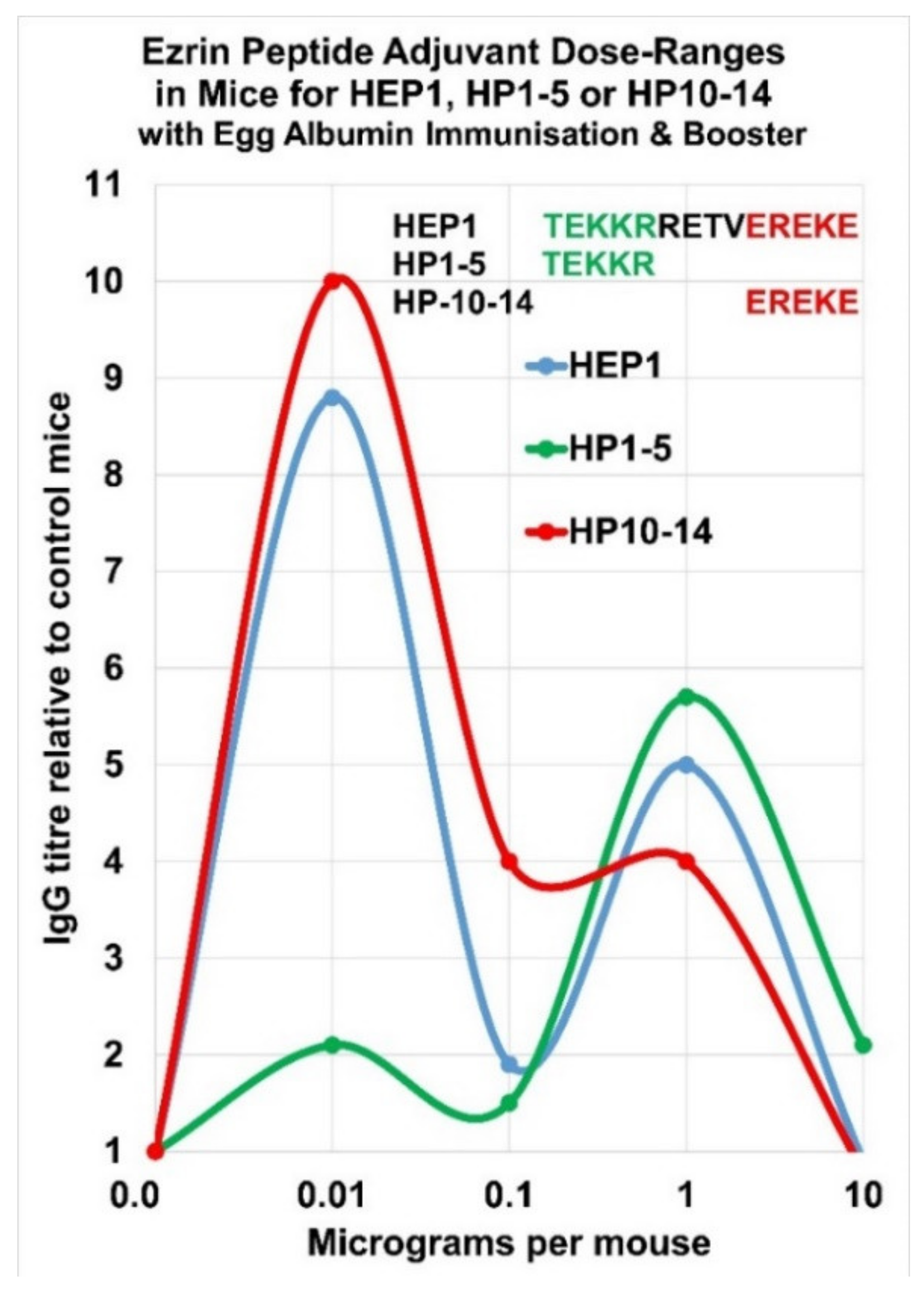
7. Complex Adjuvant Effect of Ezrin Peptides with Egg-Albumin Antigen in Mice
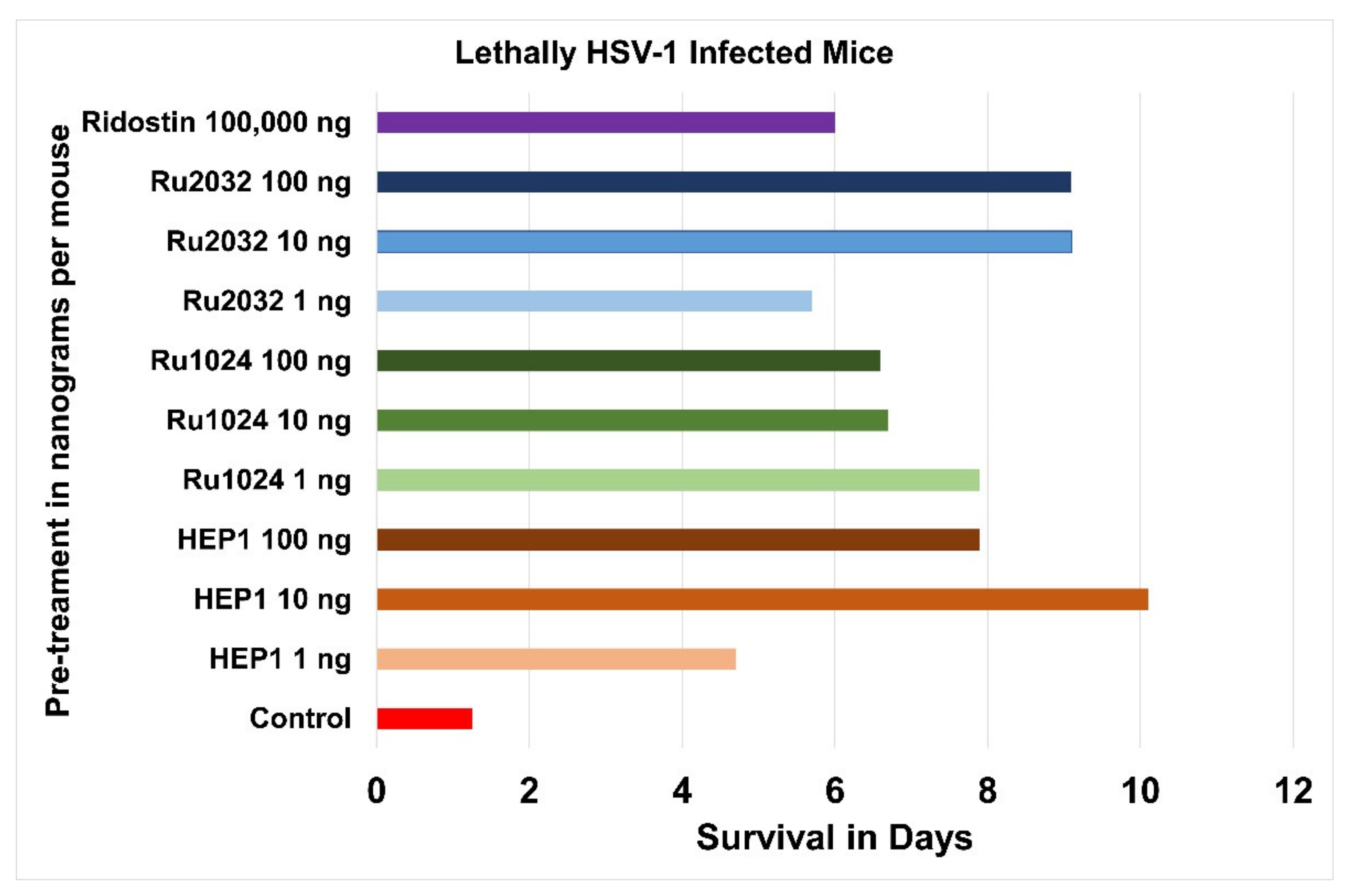
8. Immune Protection with Ezrin Peptides from Lethal Herpes Infection in Mice
9. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Solovev, A.M. Conservative Therapy of Lesions caused by Papiloma Virus (HPV). Prescr. Physician 2003, 22–26. (In Russian) [Google Scholar]
- Kulikov, I.A. Promising treatment for recurrent vulvovaginal candidiasis. Meditinsky Sov. 2020, 13, 61–69. (In Russian) [Google Scholar] [CrossRef]
- Pore, D.; Gupta, N. The ezrin-radixin-moesin family of proteins in the regulation of B-cell immune response. Crit. Rev. Immunol. 2015, 35, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parameswaran, N.; Enyindah-Asonye, G.; Bagheri, N.; Shah, N.B.; Gupta, N. Spatial Coupling of JNK Activation to the B Cell Antigen Receptor by Tyrosine-Phosphorylated Ezrin. J. Immunol. 2013, 190, 2017–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katlinskii, A.; Ataullakhanov, R.; Holms, R.; Pichugin, A.; Masternak, T.; Malkina, E.; Shishkova, N. Dependence of immune-adjuvant action of immune-modulator Gepon on peptide structure. Allergy Asthma Clin. Immunol. 2002, 6, 8–12. (In Russian) [Google Scholar]
- Holms, R.; Katlinskii, A.; Ataullakhanov, R.; Pichugin, A.; Masternak, T.; Malkina, E.; Shishkova, N. Immuno-adjuvant properties of synthetic peptides derived from the hinge region of ezrin. Allergy Asthma Clin. Immunol. 2002, 6, 13–16. (In Russian) [Google Scholar]
- Katlinskii, A.; Ataullakhanov, R.; Holms, R.; Narovlyanskii, A.; Mezentseva, M.; Sherbenko, V.; Yershov, F. Anti-viral activity of Gepon and its structural homologues in human cell culture infected with encephalomyocarditis virus. Allergy Asthma Clin. Immunol. 2002, 6, 23–25. (In Russian) [Google Scholar]
- Holms, R.; Katlinskii, A.; Ataullakhanov, R.; Narovlyanskii, A.; Mezentseva, M.; Sherbenko, V.; Yershov, F. Synthetic peptides mimicking the hinge region of human ezrin have anti-viral activity in cell culture infected with encephalomyocarditis virus. Allergy Asthma Clin. Immunol. 2002, 6, 26–28. (In Russian) [Google Scholar]
- Shamsheva, O.V.; Ospinova, M.A.; Polyakov, V.A. Protective Vaccination against Hepatitis B in Children with Malignant Tumours. Child. Infect. 2003, 4, 10–13. [Google Scholar]
- Aziz, M.; Fatima, R.; Assaly, R. Elevated interleukin-6 and severe COVID-19: A meta-analysis. J. Med. Virol. 2020, 92, 2283–2285. [Google Scholar] [CrossRef]
- Lowery, S.A.; Sariol, A.; Perlman, S. Innate immune and inflammatory responses to SARS-CoV-2: Implications for COVID-19. Cell Host Microbe 2021, 29, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Subramanian, A.; Taia, T.; Wang, T.T. An aberrant inflammatory response in severe COVID-19. Cell Host Microbe 2021, 29, 1043–1047. [Google Scholar] [CrossRef]
- Hsu, A.C.-Y.; Wang, G.; Reid, A.T.; Veerati, P.C.; Pathinayake, P.S.; Daly, K.; Mayall, J.R.; Hansbro, P.M.; Horvat, J.C.; Wang, F.; et al. SARS-CoV-2 Spike protein promotes hyper-inflammatory response that can be ameliorated by Spike-antagonistic peptide and FDA-approved ER stress and MAP kinase inhibitors in vitro. bioRxiv 2020. [Google Scholar] [CrossRef]
- Patra, T.; Meyer, K.; Geerling, L.; Isbell, T.S.; Hoft, D.F.; Brien, J.; Pinto, A.K.; Ray, R.B.; Ray, R. SARS-CoV-2 spike protein promotes IL-6 trans-signaling by activation of angiotensin II receptor signaling in epithelial cells. PLoS Pathog. 2020, 16, e1009128. [Google Scholar] [CrossRef]
- Campbell, G.R.; To, R.K.; Hanna, J.; Spector, S.A. SARS-CoV-2, SARS-CoV-1, and HIV-1 derived ssRNA sequences activate the NLRP3 inflammasome in human macrophages through a non-classical pathway. iScience 2021, 24, 102295. [Google Scholar] [CrossRef] [PubMed]
- Campochiaro, C.; Dagna, L. The conundrum of interleukin-6 blockade in COVID-19. Lancet Rheumatol. 2020, 2, e579–e580. [Google Scholar] [CrossRef]
- Chen, L.Y.C.; Hoiland, R.L.; Stukas, S.; Wellington, C.L.; Sekhon, M.S. Assessing the importance of interleukin-6 in COVID-19. Lancet Respir. Med. 2021, 9, e13. [Google Scholar] [CrossRef]
- Simon, A.; Jones, S.A.; Christopher, A.; Hunter, C.A. Is IL-6 a key cytokine target for therapy in COVID-19? Nat. Rev. Immunol. 2021, 21, 337–339. [Google Scholar] [CrossRef]
- Rubin, E.J.; Dan, L.; Longo, D.L.; Lindsey, R.; Baden, L.R. Interleukin-6 Receptor Inhibition in Covid-19—Cooling the In-flammatory Soup. N. Engl. J. Med. 2021, 384, 1564–1565. [Google Scholar] [CrossRef] [PubMed]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the human innate immune system. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef] [PubMed]
- Holms, R.D.; Ataullakhanov, R.I. Ezrin-Derived Peptides and Uses. UK Patent Application No 2100996.4, 25 January 2021. [Google Scholar]
- Holms, R.D.; Bessler, W.G.; Konopleva, M.V.; Ataullakhanov, R.I. Review of Russian Ezrin Peptide Treatment of Acute Viral Respiratory Disease and Virus Induced Pneumonia; A Potential Treatment for COVID-19. 2020. Available online: https://www.researchgate.net/publication/340600041_Review_of_Russian_ezrin_peptide_treatment_of_acute_viral_respiratory_disease_and_virus_induced_pneumonia_a_potential_treatment_for_covid-19 (accessed on 13 August 2021).
- Holms, R.D.; Ataullakhanov, R.I. Review of a Potential Cure for COVID with Human Ezrin Peptides: An Ideal Combination of Anti-Inflammatory and Anti-Viral Immune Activity; ResearchGate: Berlin, Germany, 2021. [Google Scholar] [CrossRef]
- Chanh, C.T.; Dreesman, G.R.; Kanda, P.; Linette, G.P.; Sparrow, J.T.; Ho, D.D.; Kennedy, R.C. Induction of anti-HIV neutralizing antibodies by synthetic peptides. EMBO J. 1986, 5, 3065–3071. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Kien, F.; Cheung, C.-Y.; Siu, Y.-L.; Chan, W.-L.; Li, H.; Leung, H.-L.; Jaume, M.; Bruzzone, R.; Peiris, J.S.M.; et al. Ezrin Interacts with the SARS Coronavirus Spike Protein and Restrains Infection at the Entry Stage. PLoS ONE 2012, 7, e49566. [Google Scholar] [CrossRef]
- Holms, R.D. Aids Prophylactics. International Patent Application No. PCT/GB95/001285, 2 June 1995. [Google Scholar]
- Holms, R.D. Regulatory/Unfolding Peptides of Ezrin. International Patent Application No. PCT/GB00/03566, 15 September 2000. [Google Scholar]
- Holms, R.D.; Ataullakhanov, R.I. HCV Combination Therapy. International Patent Application No. PCT/GB2004/000330, 27 January 2004. [Google Scholar]
- Holms, R.D.; Ataullakhanov, R.I. The Use of Peptides In Anti-Ulcer Therapy. International Patent Application No. PCT/GB2006/004390, 23 November 2006. [Google Scholar]
- Holms, R.D.; Ataullakhanov, R.I.; Ataullakhanov, R.; Sayadyan, K. Ezrin-Derived Peptides and Pharmaceutical Compositions Thereof. International Patent Application No. PCT/EP2016/062336, 1 June 2016. [Google Scholar]
- Gepon’ Preparation is Registered by the Russian Ministry of Health (Registration Documents R No: 000015/01-2000 of 12.07.2000, FSP 42-0012-0015-00). The Instruction on the Medical Application of the ‘Gepon’ Preparation was Approved with the Decision of the Pharmacology Committee of the Ministry of Health of the Russian Federation (Minutes No: 8 of 7 October 1999). The Preparation Belongs to the Group of Immunomodulators and is Prescribed to Increase the Immune Defence Mechanism, to Treat and to Prevent Bacterial, Viral and Fungal Infections. Available online: https://avexima.ru/medicines/gepon/ (accessed on 13 August 2021).
- Gepon Medical Instructions. Available online: https://295222.selcdn.ru/avexima.ru/iblock/3d6/3d60905d8aeaeaaf4f5c1fd5c1d77525/f690b9c70c05d60181093820b56e5c07.pdf (accessed on 13 August 2021).
- Ataullakhanov, R.I.; Holms, R.D.; Katlinsky, A.V.; Pichugin, A.V.; Papuashvili, M.N.; Shishkova, N.M. Treatment with Gepon immunomodulator increases the efficacy of the immune control of opportunistic infections in HIV-infected patients. Allergy Asthma Clin. Immunol. 2002, 3–11. (In Russian) [Google Scholar]
- Safonova, O.A.; Pichukin, A.B.; Kozhemyakina, E.S.; Malshev, N.A.; Ataullakhanov, R.I. Treatment of Acute Respiratory Disease and its Complications (Pneumonia). Chapter 7 Immunotherapy of Respiratory Disease. Section 2 Immunotherapy of Diseases of the Respiratory Tract. In Immunotherapia; Khaitov, R.M., Ataullakhanov, R.I., Eds.; Izdatekskaya Group, GEOTAR-Media: Moscow, Russia, 2011; pp. 122–158. [Google Scholar]
- Kadova, O.V.; Kharlamova, F.S.; Uchaikin, V.F.; Sherbakova, A.A.; Legkova, T.P.; Feldfiks, L.I. Effective treatment of Croup Syndrome with immunomodulator Gepon. Russ. Med. J. 2002, 147, 138–141. [Google Scholar]
- Polyakova, T.O.; Magomedov, M.M.; Artemyev, M.E.; Surikov, E.V.; Palchun, V.T. A new approach in the treatment of chronic diseases of the pharynx. Attend. Dr. 2002, 64–65. (In Russian) [Google Scholar]
- Kladova, O.V.; Kharlamova, F.S.; Shcherbakova, A.A.; Legkova, T.P.; Feldfix, L.I.; Znamenskaya, A.A.; Ovchinnikova, G.S.; Uchaikin, V.F. The first experience of Hepon intranasal application in children with respiratory infections. Pediatrics 2002, 86–88. (In Russian) [Google Scholar]
- Kladova, O.V.; Uchiakin, V.F. Clinical Study Report on intranasal application of Gepon in children with the respiratory infections: Safety and efficacy of Gepon in Acute Respiratory Virus Infection [AVRI], Virus Induced Inflammation [VII], Laryngo-Tracheo-Bronchitis with Laryngeal Stenosis [SLTB] and Recurrent Croup [RC] syndrome. Unpublished.
- Salamov, G.; Holms, R.D.; Bessler, W.; Ataullakhanov, R. Treatment of Hepatitis C Virus Infection with Human Ezrin Peptide One (HEP1) in HIV Infected Patients. Arzneim. Forschung Drug Res. 2007, 57, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Gorbarets, I.P.; Voronkova, N.V.; Lopatina, T.V.; Ivanovskaya, V.N.; Braginsky, D.M.; Blokhina, N.P.; Malyshev, N.A. The combined use of Hepon drug product and recombinant interferon-alpha in the patients with chronic hepatitis C increases the efficiency of antiviral treatment and reduces side effects of the therapy. Hepatology 2003, 23–28. (In Russian) [Google Scholar]
- Cherednichenko, T.V.; Uchaikin, V.F.; Chaplygina, G.V.; Kurbanova, G.M. A new efficient treatment of viral hepatitis. Attend. Dr. 2003, 82–83. (In Russian) [Google Scholar]
- Marchenko, L.A.; Lushkova, I.L.; Shurshalina, A.V. Treatment of patients with recurrent genital herpes. Quest. Gynecol. Obstet. Perinatol. 2004, 3, 90–93. (In Russian) [Google Scholar]
- Tishchenko, A.L. A new approach in the treatment of recurrent urogenital candidiasis. Attend. Dr. 2002, 46–47. (In Russian) [Google Scholar]
- Telunts, A.V. Treatment of candidiasis in infants. Quest. Gynecol. Obstet. Perinatol. 2004, 3, 89–90. (In Russian) [Google Scholar]
- Lazebnik, L.B.; Zvenigorodskaya, L.A.; Firsakova, V.Y.; Pichugin, A.V.; Ataullakhanov, R.I. The application of Hepon immunomodulator in the treatment of erosive ulcerous lesions of gastroduodenal zone. Exp. Clin. Gastroenterol. 2003, 17–20. (In Russian) [Google Scholar]
- Malakhova, N.S.; Pichugin, A.V.; Khalif, I.L.; Ataullakhanov, R.I. The application of Hepon immunomodulator for the treatment of nonspecific ulcerative colitis. Farmateka 2005, 105–108. (In Russian) [Google Scholar]
- Bardychev, M.S. Treatment of local radiation injuries with the activator of local immunity. Russ. Med. J. 2003, 11, 646–647. (In Russian) [Google Scholar]
- Parfenov, A.I. The activator of the local immunity Hepon in the complex treatment of dysbiotic disorders of the intestine. Exp. Clin. Gastroenterol. 2003, 66–69. (In Russian) [Google Scholar]
- Novokshonov, A.A.; Uchaikin, V.F.; Sokolova, N.V.; Tikhonova, O.N.; Portnykh, O.Y. Biocenosis-protecting therapy of intestinal infections in children. Russ. Med. J. 2004, 6, 1. (In Russian) [Google Scholar]
- Dudchenko, M.A.; Lysenko, B.F.; Chelishvili, A.L.; Katlinsky, A.V.; Ataullakhanov, R.R. Complex treatment of trophic ulcers. Attend. Dr. 2002, 72–75. (In Russian) [Google Scholar]
- Chulkina, M.; Negmadjanov, U.; Lebedeva, E.; Pichugin, A.; Mazurova, D.; Ataullakhanov, R.; Holmuhamedov, E. Synthetic peptide TEKKRRETVEREKE derived from ezrin induces differentiation of NIH/3T3 fibroblasts. Eur. J. Pharmacol. 2017, 811, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Ataullakhanov, R.I.; Katlinsky, A.V.; Holms, R.D.; Masternak, T.B.; Larin, A.S.; Malkina, E.Y.; Shishkova, N.M. Amplification of antibody formation under the influence of immunomodulator Gepon. Immunlogy 2003, 1, 9–11. [Google Scholar]
- Katlinky, A.V.; Ataullakhanov, R.I.; Holms, R.D.; Pichugin, A.V.; Masternak, T.B.; Malkina, E.Y.; Shishkova, N.M. Immuno-adjuvant action of structural homologues of immunomodulator Gepon. Immunology 2003, 1, 12–14. [Google Scholar]
- García-Ortiz, A.; Serrador, J.M. ERM Proteins at the Crossroad of Leukocyte Polarization, Migration and Intercellular Adhesion. Int. J. Mol. Sci. 2020, 21, 1502. [Google Scholar] [CrossRef] [Green Version]
- Rozsnyayac, Z.; Sarmaya, G.; Zöllerc, M.; Gergelyb, J. Membrane-bound ezrin is involved in B-cell receptor-mediated signalling: Potential role of an ITAM-like ezrin motif. Immunol. Lett. 1996, 54, 163–169. [Google Scholar] [CrossRef]
- Harwood, N.E.; Batista, F.D. The Cytoskeleton Coordinates the Early Events of B-cell Activation. Cold Spring Harb. Perspect. Biol. 2011, 3, a002360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treanor, B.; Depoil, D.; Bruckbauer, A.; Batista, F.D. Dynamic cortical actin remodeling by ERM proteins controls BCR microcluster organization and integrity. J. Exp. Med. 2011, 208, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Liu, C.; Upadhyaya, A. The pivotal position of the actin cytoskeleton in the initiation and regulation of B cell receptor activation. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kladova, O.; Kharlamova, F.; Sherbakova, A.; Legkova, T.; Feldfiks, L.; Uchaikin, V. Immunomodulator Gepon: An Effective Treatment for Croup Syndrome. Russ. Med. J. 2002, 147, 138–141. [Google Scholar]
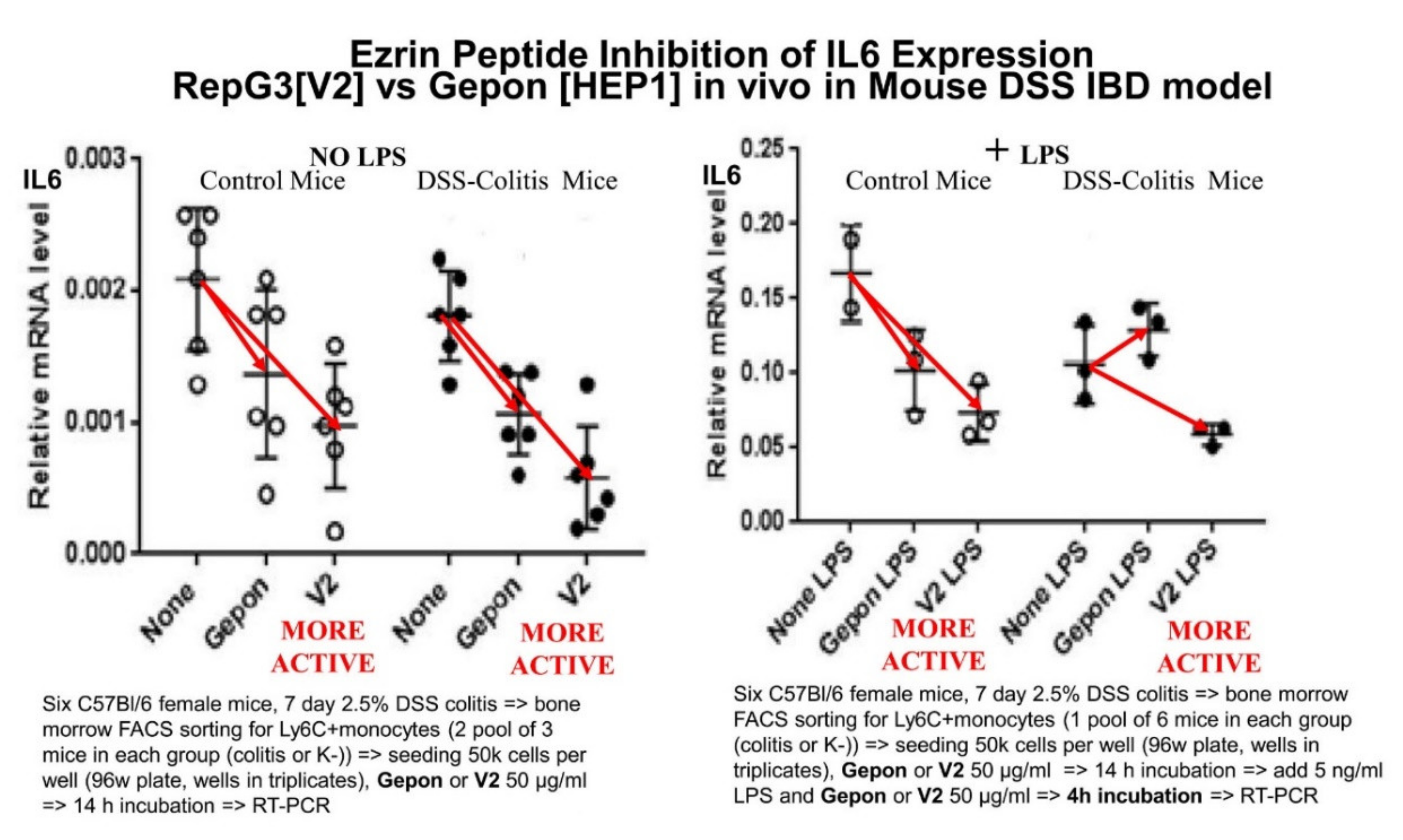
- Chulkina, M.M.; Pichugin, A.V.; Ataullakhanov, R.I. Pharmaceutical grade synthetic peptide Thr-Glu-Lys-Lys-Arg-Arg-Glu-Thr-Val-Glu-Arg-Glu-Lys-Glu ameliorates DSS-induced murine colitis by reducing the number and pro-inflammatory activity of colon tissue-infiltrating Ly6G+ granulocytes and Ly6C+ monocytes. Peptides 2020, 132, 170364. [Google Scholar] [CrossRef] [PubMed]
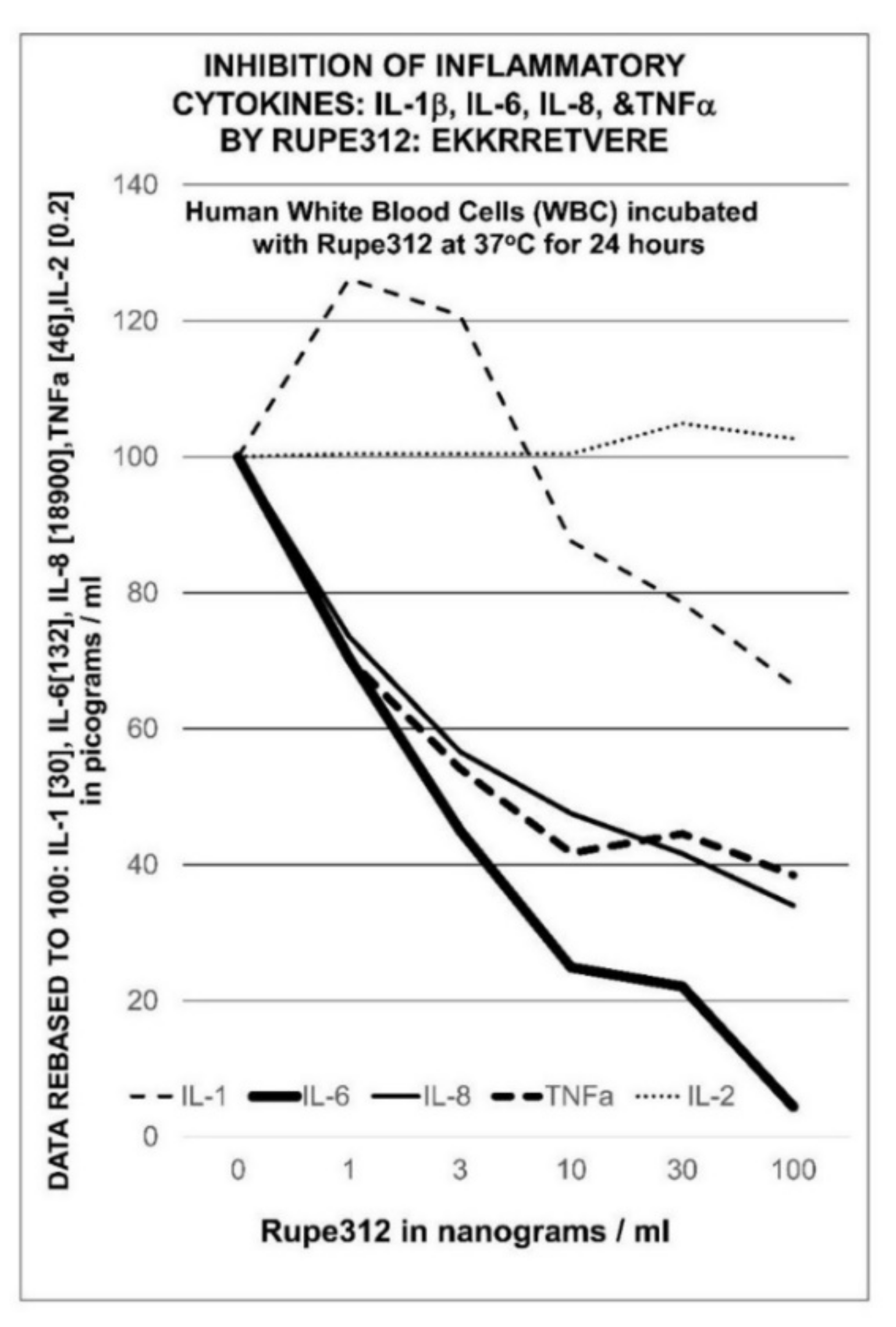
- Ataullakhanov, R.I.; Holms, R.D.; Narovlanskii, A.N.; Katlinskii, A.B.; Mezentseva, M.V.; Sherbennko, V.E.; Farfarovskii, V.C.; Yershov, F.I. Change in cytokine gene transcription in human cells under the influence of immune-modulator Gepon. Allergy Asthma Clin. Immunol. 2002, 6, 17–22. (In Russian) [Google Scholar]
- Ataullakhanov, R.I.; Katlinskii, A.B.; Holms, R.D.; Masternak, T.B.; Lapin, A.S.; Makina, E.U.; Shishkova, N.M. Immuno-modular Gepon enhances anti-body production. Allergy Asthma Clin. Immunol. 2002, 6, 3–7. (In Russian) [Google Scholar]
- Danilenk, E.D.; Belkina, A.O.; Sysoeva, G.M. Development of Drugs Based on High-Polymeric Double-Stranded RNA for Antiviral and Antitumor Therapy. Biochem. Suppl. Ser. B Biomed. Chem. 2019, 4, 308–323. [Google Scholar] [CrossRef]
- Barinsky, I.F.; Alimbarova, L.M.; Lazarenko, A.A.; Ataullakhanov, R.I. Antiviral activity of the peptide immunomodulator Gepon in experimental infections caused by herpes simplex virus types 1 and 2. Probl. Virol. 2003, 5, 30–33. [Google Scholar]
- Available online: https://www.abcam.com/products?keywords=6F1A9++ab205381 (accessed on 13 August 2021).
- Li, J.; Yin, W.; Jing, Y.; Kang, D.; Yang, L.; Cheng, J.; Yu, Z.; Peng, Z.; Li, X.; Wen, Y.; et al. The Coordination Between B Cell Receptor Signaling and the Actin Cytoskeleton During B Cell Activation. Front. Immunol. 2019, 9, 3096. Available online: https://doi.org/10.3389/fimmu.2018.03096 (accessed on 13 August 2021).
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor CREB in Immune Function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [PubMed] [Green Version]

















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holms, R.D.; Ataullakhanov, R.I. Ezrin Peptide Therapy from HIV to COVID: Inhibition of Inflammation and Amplification of Adaptive Anti-Viral Immunity. Int. J. Mol. Sci. 2021, 22, 11688. https://doi.org/10.3390/ijms222111688
Holms RD, Ataullakhanov RI. Ezrin Peptide Therapy from HIV to COVID: Inhibition of Inflammation and Amplification of Adaptive Anti-Viral Immunity. International Journal of Molecular Sciences. 2021; 22(21):11688. https://doi.org/10.3390/ijms222111688
Chicago/Turabian StyleHolms, Rupert D., and Ravshan I. Ataullakhanov. 2021. "Ezrin Peptide Therapy from HIV to COVID: Inhibition of Inflammation and Amplification of Adaptive Anti-Viral Immunity" International Journal of Molecular Sciences 22, no. 21: 11688. https://doi.org/10.3390/ijms222111688
APA StyleHolms, R. D., & Ataullakhanov, R. I. (2021). Ezrin Peptide Therapy from HIV to COVID: Inhibition of Inflammation and Amplification of Adaptive Anti-Viral Immunity. International Journal of Molecular Sciences, 22(21), 11688. https://doi.org/10.3390/ijms222111688