
Unexpected Interacting Effects of Physical (Radiation) and Chemical (Bisphenol A) Treatments on Male Reproductive Functions in Mice


Abstract
:1. Introduction
2. Results
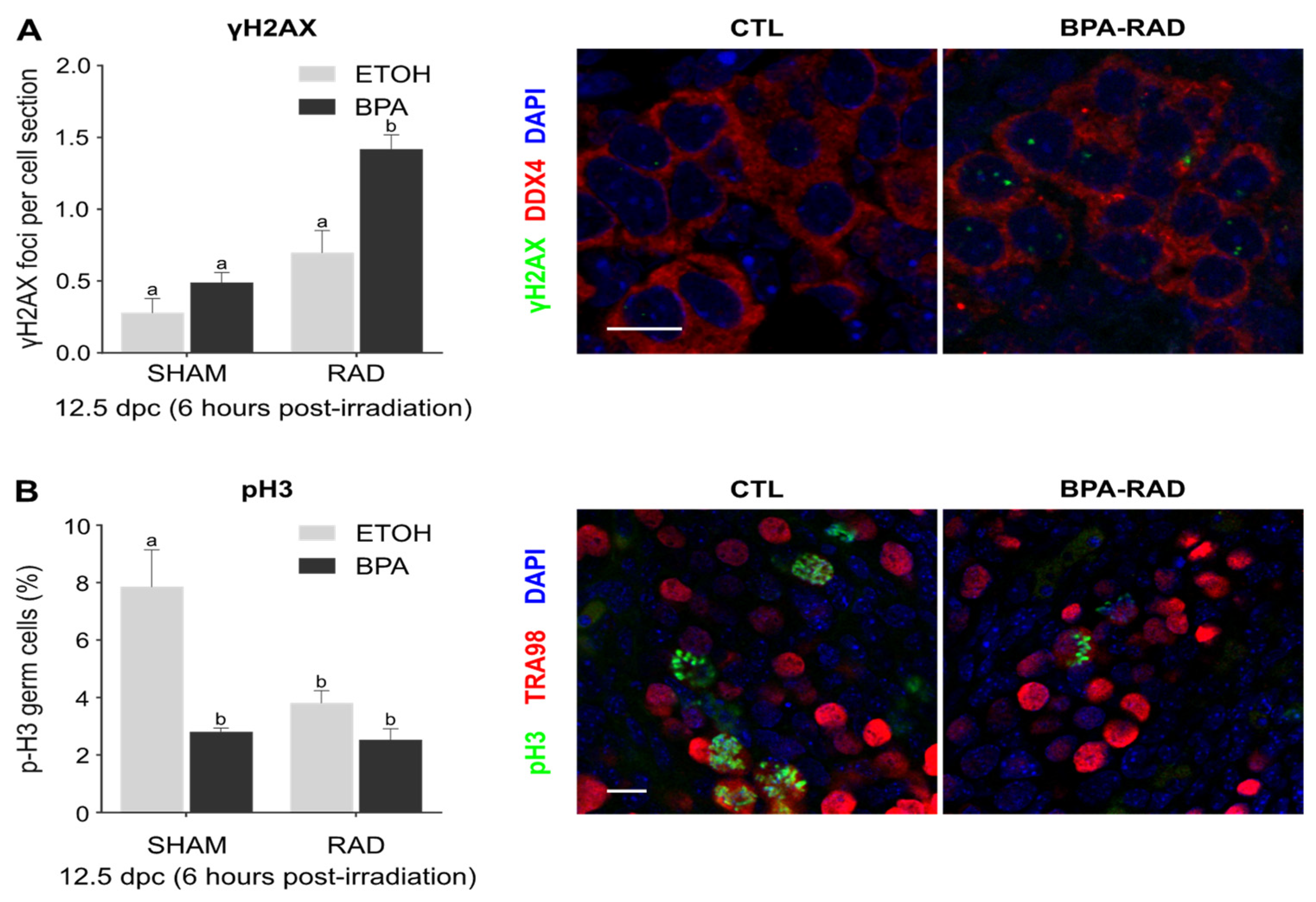
2.1. Synergistic Effects of BPA and RAD on DSBs and Consequences for Germ Cell Proliferation and Viability
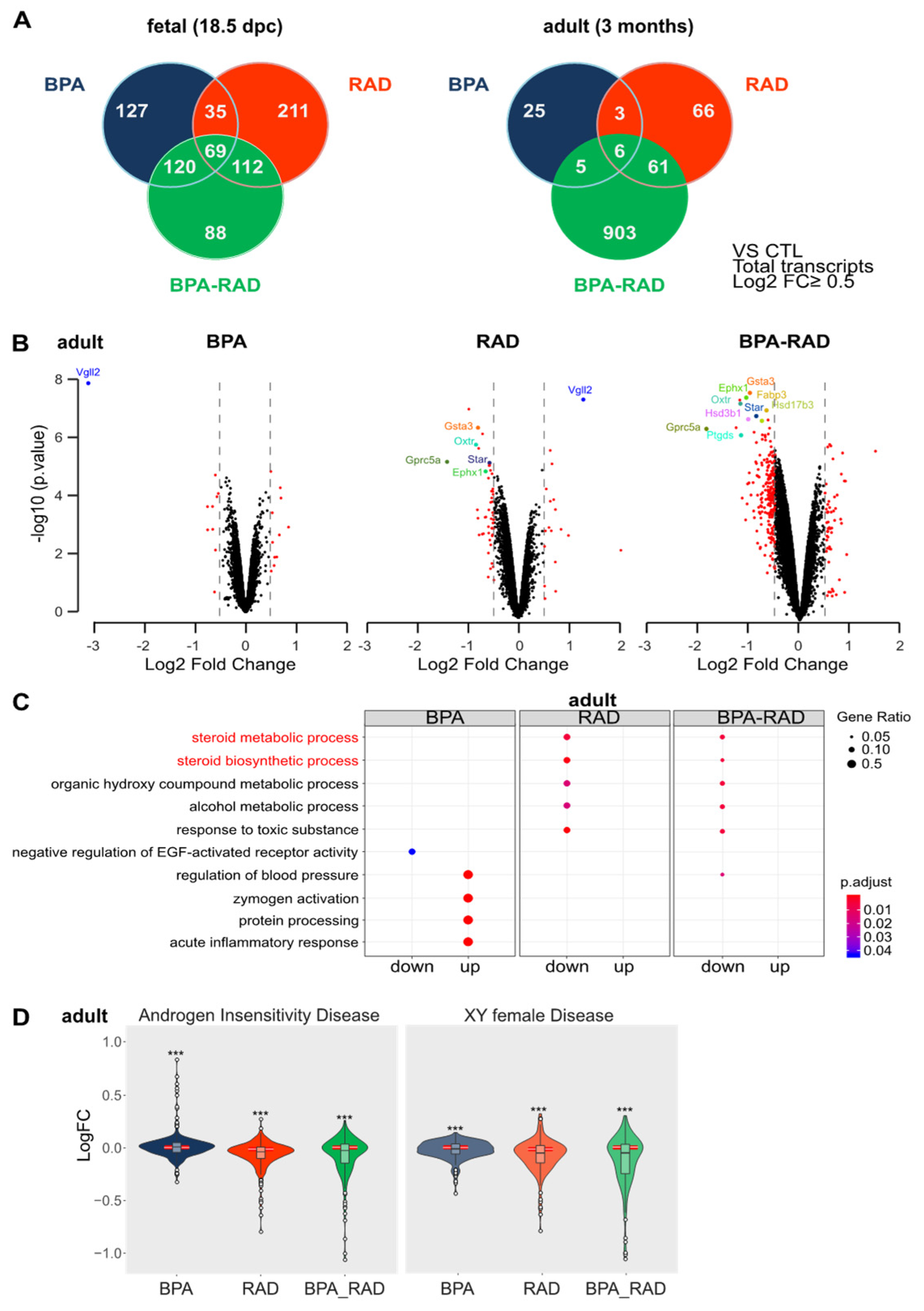
2.2. BPA and RAD Induced Long-Term Transcriptional Alterations in the Testis
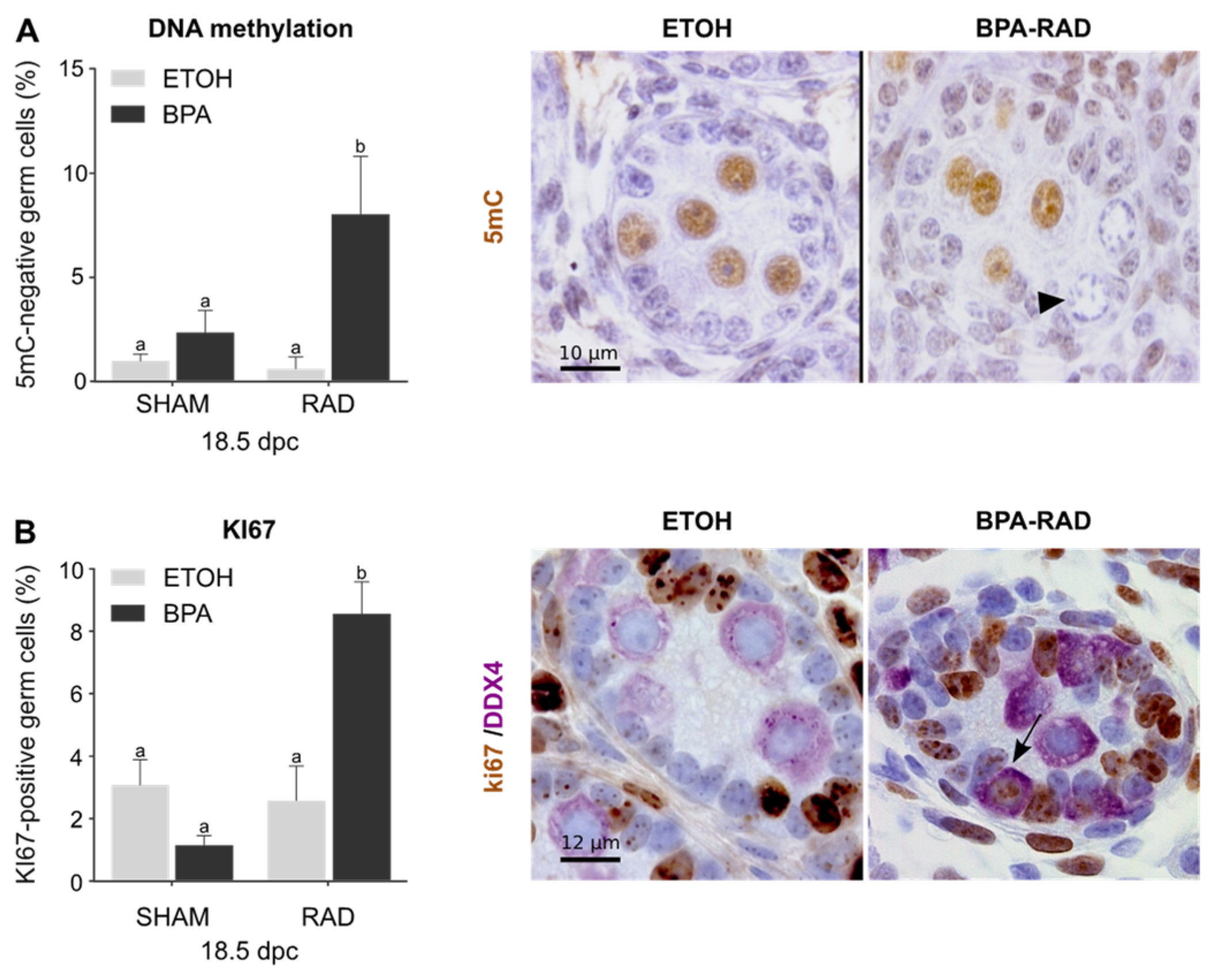
2.3. BPA and RAD Act Synergistically to Induce Gonocyte-to-Spermatogonia Transition
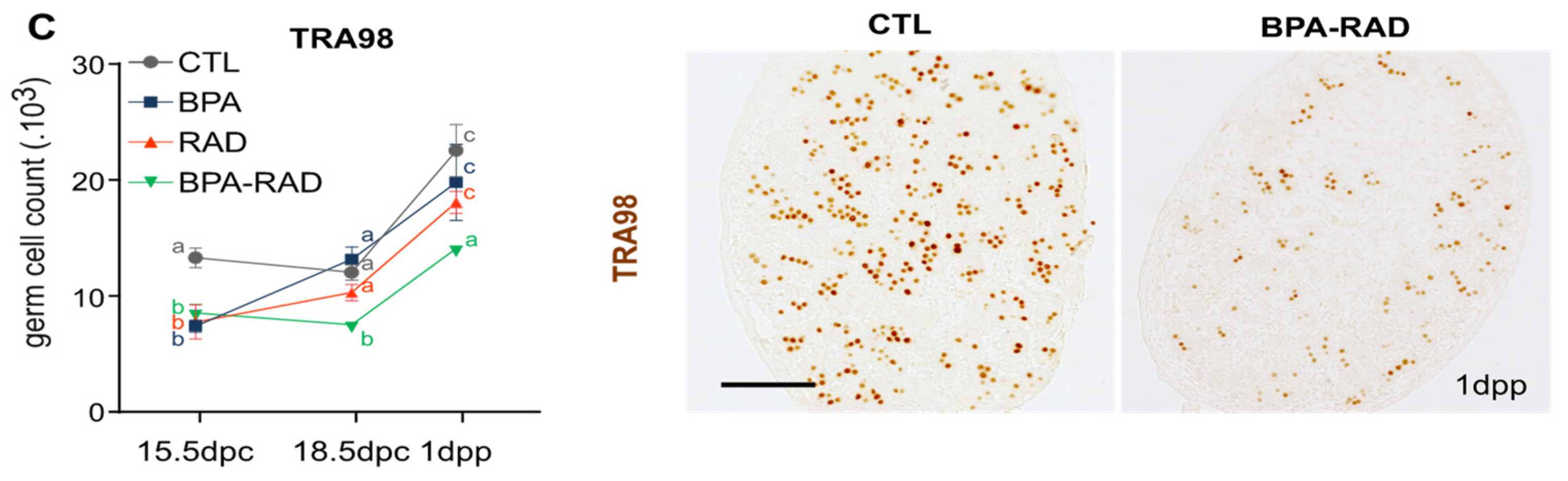
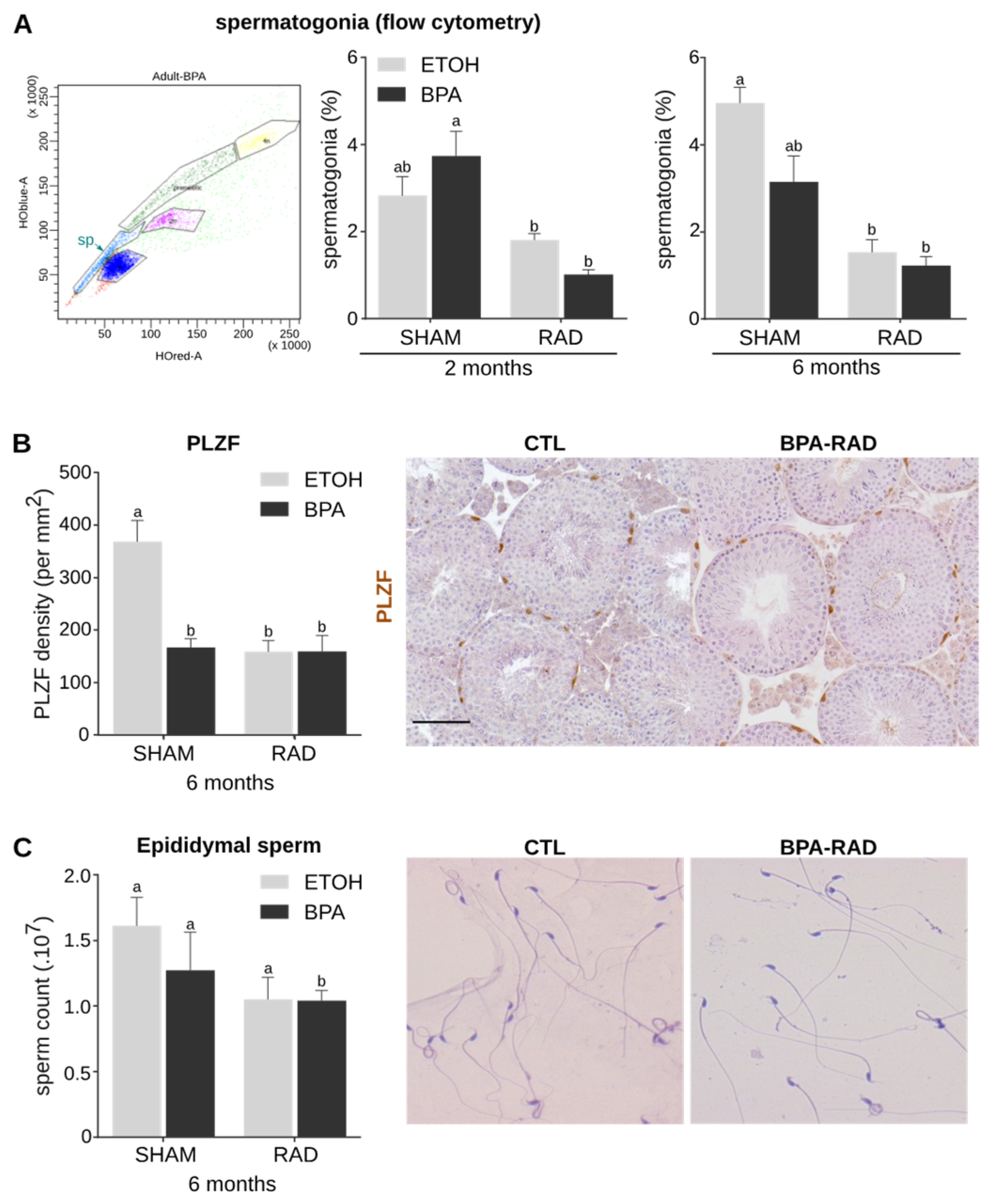
2.4. BPA and RAD Act Synergistically to Decrease Spermatogonia and Sperm Contents
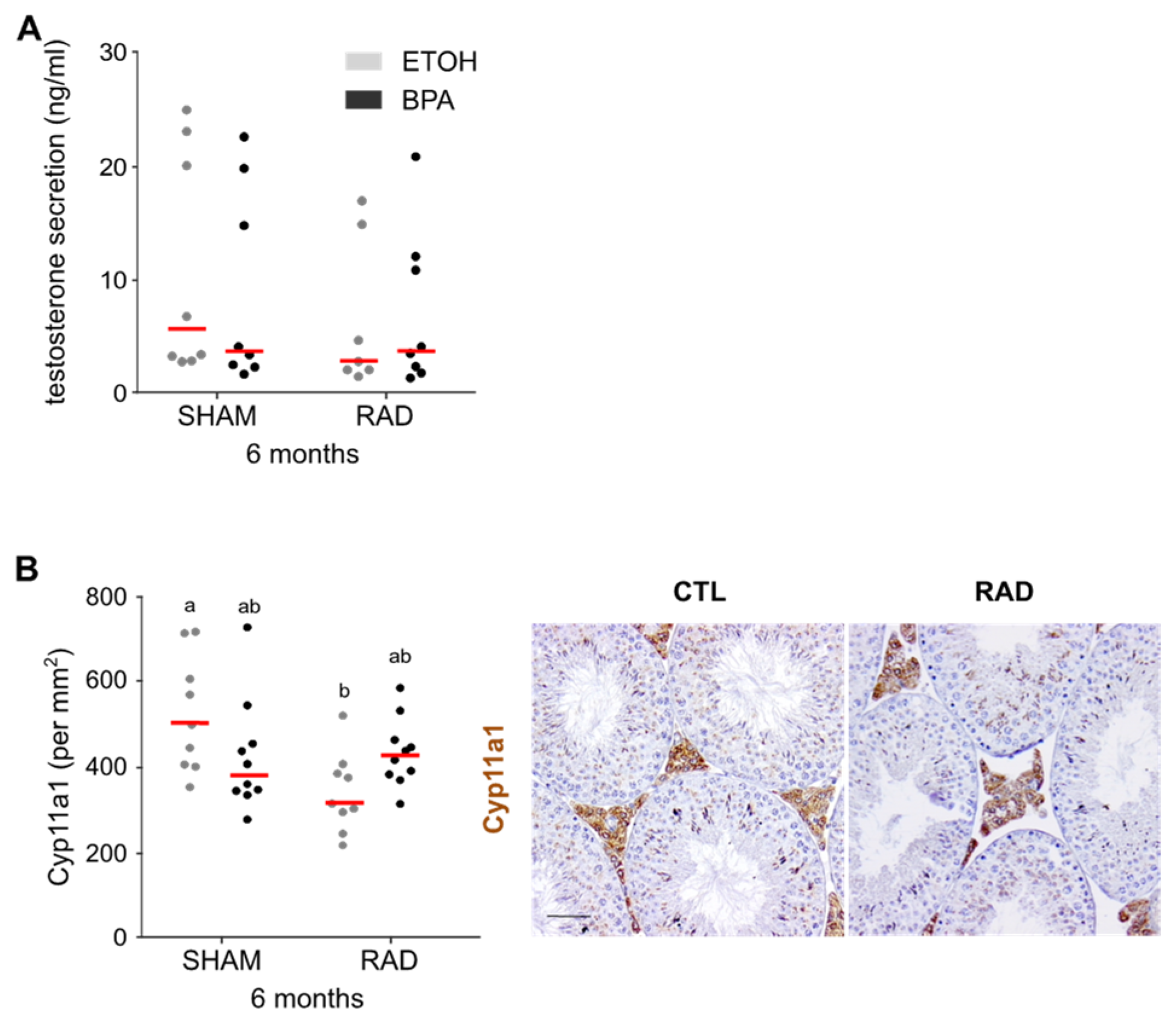
2.5. RAD Fetal Exposure Alone Decreases Androgen Production and the Number of Leydig Cells
2.6. RAD Fetal Exposure Decreases the Emission of Male Mouse Ultrasonic Vocalizations during Courtship
3. Discussion
4. Materials and Methods
4.1. BPA and IR Dose Selection after In Vitro Study
4.2. Animal Housing and Experimental Design
4.3. Histology and Immunostaining of Tissue Sections
4.4. RNA Extraction and Gene Chip Analyses
4.5. Pre-Processing and Microarray Analyses
4.6. Differentially Expressed Genes and Functional Enrichment Analyses
4.7. Interaction Study on Transcriptome Analyses
4.8. Hoechst 33342 and Propidium Iodide Staining and Flow Cytometry
4.9. Sperm Count
4.10. Testosterone Radioimmunoassay
4.11. Partner Preferences and Ultrasonic Vocalization Recordings
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Felice, F.; Marchetti, C.; Marampon, F.; Cascialli, G.; Muzii, L.; Tombolini, V. Radiation effects on male fertility. Andrology 2018, 7, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Hamer, G.; De Rooij, D.G. Mutations causing specific arrests in the development of mouse primordial germ cells and gonocytes. Biol. Reprod. 2018, 99, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Lambrot, R.; Coffigny, H.; Pairault, C.; Lécureuil, C.; Frydman, R.; Habert, R.; Rouiller-Fabre, V. High Radiosensitivity of Germ Cells in Human Male Fetus. J. Clin. Endocrinol. Metab. 2007, 92, 2632–2639. [Google Scholar] [CrossRef] [Green Version]
- Sapsford, C. The Effect of X-Rays on Gonocytes in the Testis of the Rat. Aust. J. Biol. Sci. 1965, 18, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowron, S. Radiosensitivity of Immature Male Gonads in Golden Hamster. II. Gonocytes and Spermatogonia A. Folia Biol. 1967, 15, 395–397. [Google Scholar]
- IRSN. Exposition de La Population Francaise Aux Rayonnements Ionisants: Bilan 2014–2019; IRSN: Fontenay-aux-Roses, France, 2021.
- Birioukov, A.; Meurer, M.; Peter, R.U.; Braun-Falco, O.; Plewig, G. Male Reproductive System in Patients Exposed to Ionizing Irradiation in the Chernobyl Accident. Arch. Androl. 1993, 30, 99–104. [Google Scholar] [CrossRef]
- Bai, H.; Sun, F.; Yang, G.; Wang, L.; Zhang, Q.; Zhang, Q.; Zhan, Y.; Chen, J.; Yu, M.; Li, C.; et al. CBLB502, a Toll-like receptor 5 agonist, offers protection against radiation-induced male reproductive system damage in mice. Biol. Reprod. 2018, 100, 281–291. [Google Scholar] [CrossRef]
- Belling, K.C.; Tanaka, M.; Dalgaard, M.D.; Nielsen, J.E.; Nielsen, H.B.; Brunak, S.; Almstrup, K.; Leffers, H. Transcriptome profiling of mice testes following low dose irradiation. Reprod. Biol. Endocrinol. 2013, 11, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedka, M.; Kuźba-Kryszak, T.; Nowikiewicz, T.; Zyromska, A. Fertility impairment in radiotherapy. Contemp. Oncol. 2016, 20, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colpi, G.; Contalbi, G.; Nerva, F.; Sagone, P.; Piediferro, G. Testicular function following chemo–radiotherapy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 113, S2–S6. [Google Scholar] [CrossRef]
- Delic, J.; Hendry, J.; Morris, I.; Shalet, S. Leydig cell function in the pubertal rat following local testicular irradiation. Radiother. Oncol. 1986, 5, 29–37. [Google Scholar] [CrossRef]
- Izard, M. Leydig cell function and radiation: A review of the literature. Radiother. Oncol. 1995, 34, 1–8. [Google Scholar] [CrossRef]
- Naeimi, R.A.; Amiri, F.T.; Khalatbary, A.R.; Ghasemi, A.; Zargari, M.; Ghesemi, M.; Hosseinimehr, S.J. Atorvastatin mitigates testicular injuries induced by ionizing radiation in mice. Reprod. Toxicol. 2017, 72, 115–121. [Google Scholar] [CrossRef]
- Rouiller-Fabre, V.; Guerquin, M.-J.; N’Tumba-Byn, T.; Muczynski, V.; Moison, D.; Tourpin, S.; Messiaen, S.; Habert, R.; Livera, G. Nuclear Receptors and Endocrine Disruptors in Fetal and Neonatal Testes: A Gapped Landscape. Front. Endocrinol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hunt, P.A.; Myers, J.P.; Saal, F.S.V. Human exposures to bisphenol A: Mismatches between data and assumptions. Rev. Environ. Health 2013, 28, 37–58. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.R.; Schönfelder, G. Urinary, Circulating, and Tissue Biomonitoring Studies Indicate Widespread Exposure to Bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.; Silva, J.; Santos, M.; Fardilha, M. Fighting Bisphenol A-Induced Male Infertility: The Power of Antioxidants. Antioxidants 2021, 10, 289. [Google Scholar] [CrossRef] [PubMed]
- Cariati, F.; Carbone, L.; Conforti, A.; Bagnulo, F.; Peluso, S.R.; Carotenuto, C.; Buonfantino, C.; Alviggi, E.; Alviggi, C.; Strina, I. Bisphenol A-Induced Epigenetic Changes and Its Effects on the Male Reproductive System. Front. Endocrinol. 2020, 11, 453. [Google Scholar] [CrossRef]
- Hart, R.J. The Impact of Prenatal Exposure to Bisphenol A on Male Reproductive Function. Front. Endocrinol. 2020, 11, 320. [Google Scholar] [CrossRef]
- Komarowska, M.D.; Grubczak, K.; Czerniecki, J.; Hermanowicz, A.; Hermanowicz, J.M.; Debek, W.; Matuszczak, E. Identification of the Bisphenol A (BPA) and the Two Analogues BPS and BPF in Cryptorchidism. Front. Endocrinol. 2021, 12, 694669. [Google Scholar] [CrossRef]
- Mammadov, E.; Uncu, M.; Dalkan, C. High Prenatal Exposure to Bisphenol A Reduces Anogenital Distance in Healthy Male Newborns. J. Clin. Res. Pediatr. Endocrinol. 2018, 10, 25–29. [Google Scholar] [CrossRef]
- Sun, X.; Li, D.; Liang, H.; Miao, M.; Song, X.; Wang, Z.; Zhou, Z.; Yuan, W. Maternal exposure to bisphenol A and anogenital distance throughout infancy: A longitudinal study from Shanghai, China. Environ. Int. 2018, 121, 269–275. [Google Scholar] [CrossRef]
- Eladak, S.; Moison, D.; Guerquin, M.-J.; Matilionyte, G.; Kilcoyne, K.; N’Tumba-Byn, T.; Messiaen, S.; Deceuninck, Y.; Pozzi-Gaudin, S.; Benachi, A.; et al. Effects of environmental Bisphenol A exposures on germ cell development and Leydig cell function in the human fetal testis. PLoS ONE 2018, 13, e0191934. [Google Scholar] [CrossRef]
- N’Tumba-Byn, T.; Moison, D.; Lacroix, M.; Lecureuil, C.; Lesage, L.; Prud’Homme, S.; Pozzi-Gaudin, S.; Frydman, R.; Benachi, A.; Livera, G.; et al. Differential Effects of Bisphenol A and Diethylstilbestrol on Human, Rat and Mouse Fetal Leydig Cell Function. PLoS ONE 2012, 7, e51579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, N.; Surendranath, A.R.; Pathak, J.L.; Yu, S.; Chung, C.Y. Bisphenol A (BPA) the mighty and the mutagenic. Toxicol. Rep. 2017, 5, 76–84. [Google Scholar] [CrossRef]
- Hamid, N.; Junaid, M.; Pei, D.-S. Combined toxicity of endocrine-disrupting chemicals: A review. Ecotoxicol. Environ. Saf. 2021, 215, 112136. [Google Scholar] [CrossRef]
- SCHER; SCCS. SCENIHR Opinion on the Toxicity and Assessment of Chemical Mixtures; European Commission: Brussels, Belgium, 2012. [Google Scholar]
- Cowell, W.J.; Wright, R.J. Sex-Specific Effects of Combined Exposure to Chemical and Non-chemical Stressors on Neuroendocrine Development: A Review of Recent Findings and Putative Mechanisms. Curr. Environ. Health Rep. 2017, 4, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyńska, M.M.; Jankowska-Steifer, E.A.; Tyrkiel, E.J.; Gajowik, A.; Radzikowska, J.; Pachocki, K.A. Comparison of the effects of bisphenol A alone and in a combination with X-irradiation on sperm count and quality in male adult and pubescent mice. Environ. Toxicol. 2013, 29, 1301–1313. [Google Scholar] [CrossRef]
- Dobrzyńska, M.M.; Gajowik, A.; Jankowska-Steifer, E.A.; Radzikowska, J.; Tyrkiel, E.J. Reproductive and developmental F1 toxicity following exposure of pubescent F0 male mice to bisphenol A alone and in a combination with X-rays irradiation. Toxicology 2018, 410, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyńska, M.M.; Radzikowska, J. Genotoxicity and reproductive toxicity of bisphenol A and X-ray/bisphenol A combination in male mice. Drug Chem. Toxicol. 2011, 36, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Heredia, L.; Bellés, M.; Llovet, M.I.; Domingo, J.L.; Linares, V. Behavioral effects in mice of postnatal exposure to low-doses of 137-cesium and bisphenol A. Toxicology 2016, 340, 10–16. [Google Scholar] [CrossRef]
- Kwan, W.S.; Nikezic, D.; Roy, V.A.L.; Yu, K.N. Multiple Stressor Effects of Radon and Phthalates in Children: Background Information and Future Research. Int. J. Environ. Res. Public Health 2020, 17, 2898. [Google Scholar] [CrossRef]
- Rider, C.; Boekelheide, K.; Catlin, N.; Gordon, C.J.; Morata, T.; Selgrade, M.K.; Sexton, K.; Simmons, J.E. Cumulative Risk: Toxicity and Interactions of Physical and Chemical Stressors. Toxicol. Sci. 2013, 137, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Sonavane, M.; Gassman, N.R. Bisphenol A co-exposure effects: A key factor in understanding BPA’s complex mechanism and health outcomes. Crit. Rev. Toxicol. 2019, 49, 371–386. [Google Scholar] [CrossRef]
- Thabet, N.M.; Abdel-Rafei, M.K.; Moustafa, E.M. Boswellic acid protects against Bisphenol-A and gamma radiation induced hepatic steatosis and cardiac remodelling in rats: Role of hepatic PPAR-α/P38 and cardiac Calcineurin-A/NFATc1/P38 pathways. Arch. Physiol. Biochem. 2020, 1–19. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pui, H.-P.; Saga, Y. NANOS2 acts as an intrinsic regulator of gonocytes-to-spermatogonia transition in the murine testes. Mech. Dev. 2018, 149, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Kikusui, T.; Shima, Y.; Sonobe, M.; Yoshida, Y.; Nagasawa, M.; Nomoto, K.; Mogi, K. Testosterone regulates the emission of ultrasonic vocalizations and mounting behavior during different developmental periods in mice. Dev. Psychobiol. 2020, 63, 725–733. [Google Scholar] [CrossRef]
- Guerquin, M.-J.; Duquenne, C.; Coffigny, H.; Rouiller-Fabre, V.; Lambrot, R.; Bakalska, M.; Frydman, R.; Habert, R.; Livera, G. Sex-specific differences in fetal germ cell apoptosis induced by ionizing radiation. Hum. Reprod. 2008, 24, 670–678. [Google Scholar] [CrossRef] [Green Version]
- Nakahira, R.; Ayabe, Y.; Iii, I.B.-T.; Tanaka, S.; Komura, J.-I. Effects of Continuous In Utero Low- and Medium-Dose-Rate Gamma-Ray Exposure on Fetal Germ Cells. Radiat. Res. 2020, 195, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Keating, A.F. Bisphenol A-Induced Ovotoxicity Involves DNA Damage Induction to Which the Ovary Mounts a Protective Response Indicated by Increased Expression of Proteins Involved in DNA Repair and Xenobiotic Biotransformation. Toxicol. Sci. 2016, 152, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Lombó, M.; Fernández-Díez, C.; González-Rojo, S.; Herráez, M.P. Genetic and epigenetic alterations induced by bisphenol A exposure during different periods of spermatogenesis: From spermatozoa to the progeny. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, T.X.; Hofmann, M.-C. NOTCH signaling in Sertoli cells regulates gonocyte fate. Cell Cycle 2013, 12, 2538–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Xie, J. Etomoxir regulates the differentiation of male germ cells by specifically reducing H3K27ac level. BMC Dev. Biol. 2021, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.-F.; Chen, W.S.C.; Chen, Y.-H.; Kao, T.-H.; Tseng, Y.-T.; Lee, C.-Y.; Chiu, Y.-C.; Lee, P.-L.; Lin, Q.-J.; Ching, Y.-H.; et al. DNMT3L promotes quiescence in postnatal spermatogonial progenitor cells. Development 2014, 141, 2402–2413. [Google Scholar] [CrossRef] [Green Version]
- Du, G.; Oatley, M.J.; Law, N.C.; Robbins, C.; Wu, X.; Oatley, J.M. Proper timing of a quiescence period in precursor prospermatogonia is required for stem cell pool establishment in the male germline. Development 2021, 148, dev.194571. [Google Scholar] [CrossRef]
- Kuroki, S.; Maeda, R.; Yano, M.; Kitano, S.; Miyachi, H.; Fukuda, M.; Shinkai, Y.; Tachibana, M. H3K9 Demethylases JMJD1A and JMJD1B Control Prospermatogonia to Spermatogonia Transition in Mouse Germline. Stem Cell Rep. 2020, 15, 424–438. [Google Scholar] [CrossRef]
- Moreno, S.; Attali, M.; Allemand, I.; Messiaen, S.; Fouchet, P.; Coffigny, H.; Romeo, P.-H.; Habert, R. TGFβ signaling in male germ cells regulates gonocyte quiescence and fertility in mice. Dev. Biol. 2010, 342, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-J.; Lee, W.-Y.; Do, J.T.; Park, C.; Song, H. Evaluation of testicular toxicity upon fetal exposure to bisphenol A using an organ culture method. Chemosphere 2021, 270, 129445. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.X.; DeFalco, T.; Capel, B.; Hofmann, M.-C. Constitutive activation of NOTCH1 signaling in Sertoli cells causes gonocyte exit from quiescence. Dev. Biol. 2013, 377, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Shima, Y.; Miyabayashi, K.; Sato, T.; Suyama, M.; Ohkawa, Y.; Doi, M.; Okamura, H.; Suzuki, K. Fetal Leydig cells dedifferentiate and serve as adult Leydig stem cells. Development 2018, 145, dev169136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Wen, X.; Ji, M.; Guan, X.; Chen, P.; Hao, X.; Chen, F.; Hu, Y.; Duan, P.; Ge, R.-S.; et al. Differentiation of seminiferous tubule-associated stem cells into leydig cell and myoid cell lineages. Mol. Cell. Endocrinol. 2021, 525, 111179. [Google Scholar] [CrossRef]
- Edelsztein, N.; Kashimada, K.; Schteingart, H.F.; Rey, R.A. CYP26B1 declines postnatally in Sertoli cells independently of androgen action in the mouse testis. Mol. Reprod. Dev. 2019, 87, 66–77. [Google Scholar] [CrossRef]
- Sararols, P.; Stévant, I.; Neirijnck, Y.; Rebourcet, D.; Darbey, A.; Curley, M.K.; Kühne, F.; Dermitzakis, E.; Smith, L.B.; Nef, S. Specific Transcriptomic Signatures and Dual Regulation of Steroidogenesis between Fetal and Adult Mouse Leydig Cells. Front. Cell Dev. Biol. 2021, 9, 695546. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y. Development of fetal and adult Leydig cells. Reprod. Med. Biol. 2019, 18, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhao, Y.; Li, L.; Xie, L.; Chen, X.; Liu, J.; Li, X.; Jin, L.; Li, X.; Ge, R.-S. Bisphenol A stimulates differentiation of rat stem Leydig cells in vivo and in vitro. Mol. Cell. Endocrinol. 2018, 474, 158–167. [Google Scholar] [CrossRef]
- Matsumoto, Y.K.; Okanoya, K. Mice modulate ultrasonic calling bouts according to sociosexual context. R. Soc. Open Sci. 2018, 5, 180378. [Google Scholar] [CrossRef] [Green Version]
- Musolf, K.; Meindl, S.; Larsen, A.; Kalcounis-Rueppell, M.C.; Penn, D.J. Ultrasonic Vocalizations of Male Mice Differ among Species and Females Show Assortative Preferences for Male Calls. PLoS ONE 2015, 10, e0134123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaba, A.; Osakada, T.; Touhara, K.; Kato, M.; Mogi, K.; Kikusui, T. Male mice ultrasonic vocalizations enhance female sexual approach and hypothalamic kisspeptin neuron activity. Horm. Behav. 2017, 94, 53–60. [Google Scholar] [CrossRef]
- Hammerschmidt, K.; Radyushkin, K.; Ehrenreich, H.; Fischer, J. Female mice respond to male ultrasonic ‘songs’ with approach behaviour. Biol. Lett. 2009, 5, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Chabout, J.; Sarkar, A.; Dunson, D.B.; Jarvis, E.D. Male mice song syntax depends on social contexts and influences female preferences. Front. Behav. Neurosci. 2015, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Mhaouty-Kodja, S. Courtship vocalizations: A potential biomarker of adult exposure to endocrine disrupting compounds? Mol. Cell. Endocrinol. 2019, 501, 110664. [Google Scholar] [CrossRef]
- Dombret, C.; Capela, D.; Poissenot, K.; Parmentier, C.; Bergsten, E.; Pionneau, C.; Chardonnet, S.; Hardin-Pouzet, H.; Grange-Messent, V.; Keller, M.; et al. Neural Mechanisms Underlying the Disruption of Male Courtship Behavior by Adult Exposure to Di(2-ethylhexyl) Phthalate in Mice. Environ. Health Perspect. 2017, 125, 097001. [Google Scholar] [CrossRef] [Green Version]
- Vergouwen, R.P.F.A.; Huiskamp, R.; Bas, R.J.; Roepers-Gajadien, H.L.; Davids, J.A.G.; De Rooij, D.G. Radiosensitivity of Testicular Cells in the Fetal Mouse. Radiat. Res. 1995, 141, 66. [Google Scholar] [CrossRef]
- Jansz, G.F.; Pomerantz, D.K. Fetal Irradiation Increases Androgen Production by Dispersed Leydig Cells of the Rat. J. Androl. 1984, 5, 344–350. [Google Scholar] [CrossRef]
- Keiko, S.; Masakazu, T.; Hiroko, I.-O.; Kiyomi, I.; Hiroshi, I. Steroidogenesis in the testes and the adrenals of adult male rats after γ-irradiation in utero at late pregnancy. J. Steroid Biochem. 1990, 35, 301–305. [Google Scholar] [CrossRef]
- Mahaney, B.L.; Meek, K.; Lees-Miller, S.P. Repair of ionizing radiation-induced DNA double-strand breaks by non-homologous end-joining. Biochem. J. 2009, 417, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, B.S.; Irizarry, R.A. A framework for oligonucleotide microarray preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Xu, Y.; Li, M.; Li, D.; Lu, Y.; Yu, D.; Du, W. Bisphenol A accelerates meiotic progression in embryonic chickens via the estrogen receptor β signaling pathway. Gen. Comp. Endocrinol. 2018, 259, 66–75. [Google Scholar] [CrossRef]
- Bastos, H.; Lassalle, B.; Chicheportiche, A.; Riou, L.; Testart, J.; Allemand, I.; Fouchet, P. Flow cytometric characterization of viable meiotic and postmeiotic cells by Hoechst 33342 in mouse spermatogenesis. Cytom. Part A 2005, 65A, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Picon, R.; Ktorza, A. Effect of LH on testosterone production by foetal rat testes in vitro. FEBS Lett. 1976, 68, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Hafner, S.D.; Katz, J. MonitoR: Acoustic Template Detection in R. 2018. Available online: http://www.uvm.edu/rsenr/vtcfwru/R/?Page=monitoR/monitoR.htm (accessed on 27 October 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Transcript * | Type of Interaction (% of Misregulated Transcripts) | ||||
|---|---|---|---|---|---|
| Potentiation | Additivity | Antagonism | Unexpected | ||
| Fetal | |||||
| All transcripts | 762 | 9.8 | 26.6 | 57.9 | 5.7 |
| Protein coding mRNA | 238 | 18.1 | 7.1 | 64.3 | 10.5 |
| Adult | |||||
| All transcripts | 1069 | 71.2 | 16.3 | 11.2 | 1.2 |
| Protein coding mRNA | 308 | 59.7 | 26.9 | 12.3 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wieckowski, M.; Ranga, S.; Moison, D.; Messiaen, S.; Abdallah, S.; Granon, S.; Habert, R.; Rouiller-Fabre, V.; Livera, G.; Guerquin, M.-J. Unexpected Interacting Effects of Physical (Radiation) and Chemical (Bisphenol A) Treatments on Male Reproductive Functions in Mice. Int. J. Mol. Sci. 2021, 22, 11808. https://doi.org/10.3390/ijms222111808
Wieckowski M, Ranga S, Moison D, Messiaen S, Abdallah S, Granon S, Habert R, Rouiller-Fabre V, Livera G, Guerquin M-J. Unexpected Interacting Effects of Physical (Radiation) and Chemical (Bisphenol A) Treatments on Male Reproductive Functions in Mice. International Journal of Molecular Sciences. 2021; 22(21):11808. https://doi.org/10.3390/ijms222111808
Chicago/Turabian StyleWieckowski, Margaux, Stéphanie Ranga, Delphine Moison, Sébastien Messiaen, Sonia Abdallah, Sylvie Granon, René Habert, Virginie Rouiller-Fabre, Gabriel Livera, and Marie-Justine Guerquin. 2021. "Unexpected Interacting Effects of Physical (Radiation) and Chemical (Bisphenol A) Treatments on Male Reproductive Functions in Mice" International Journal of Molecular Sciences 22, no. 21: 11808. https://doi.org/10.3390/ijms222111808
APA StyleWieckowski, M., Ranga, S., Moison, D., Messiaen, S., Abdallah, S., Granon, S., Habert, R., Rouiller-Fabre, V., Livera, G., & Guerquin, M. -J. (2021). Unexpected Interacting Effects of Physical (Radiation) and Chemical (Bisphenol A) Treatments on Male Reproductive Functions in Mice. International Journal of Molecular Sciences, 22(21), 11808. https://doi.org/10.3390/ijms222111808