
Chemokine-like Receptor 1 in Brain of Spontaneously Hypertensive Rats Mediates Systemic Hypertension


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Protein Expression of CMKLR1 around Brain Ventricles of Wistar Kyoto Rats (WKY) and SHR
2.2. Effects of CMKLR1 siRNA on Systolic Blood Pressure and Heart Rate in WKY and SHR
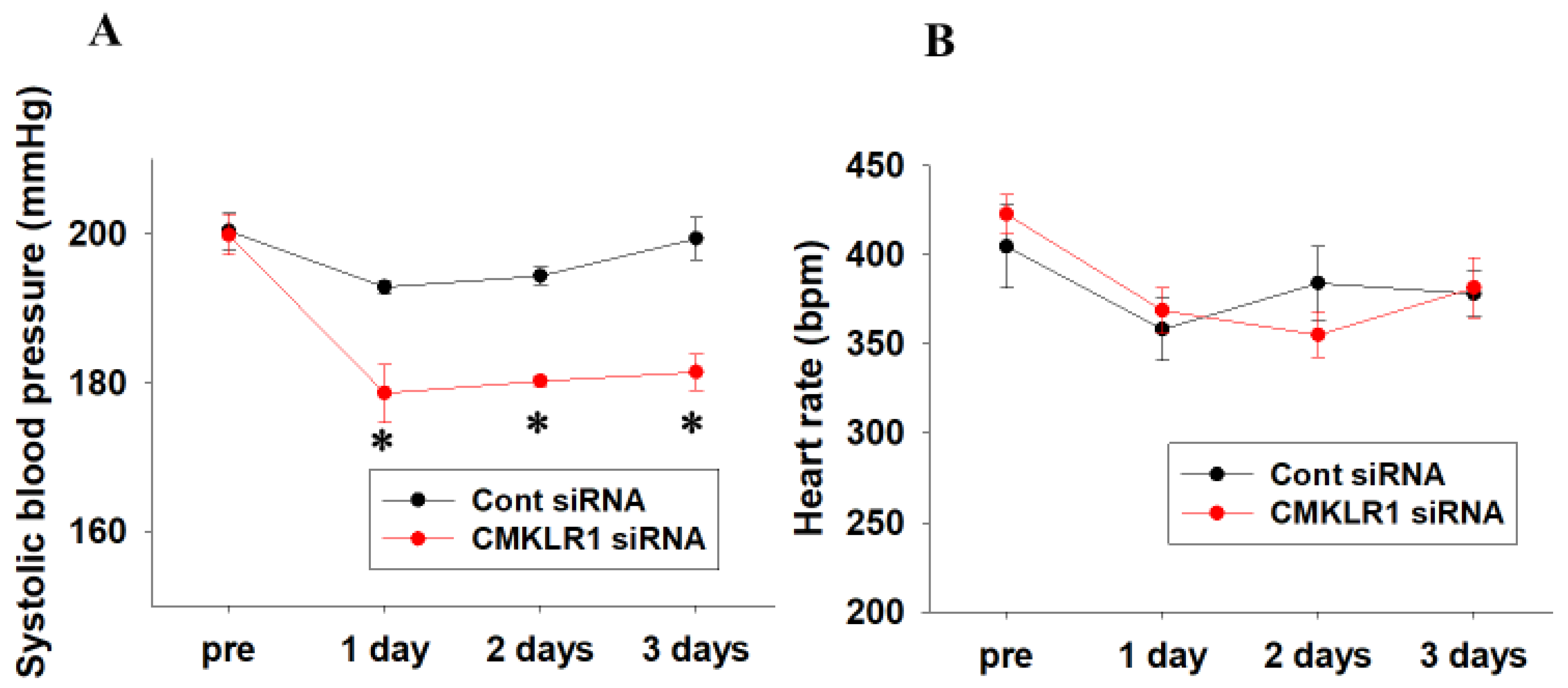
2.3. Short-Term Effects of CMKLR1 siRNA on Systolic Blood Pressure and Heart Rate in SHR
2.4. Effects of CMKLR1 siRNA on Protein Expression of CMKLR1 in PVN
2.5. Protein Expression of Chemerin around Brain Ventricles of WKY and SHR
2.6. Protein Expression of Peripheral Chemerin in WKY and SHR
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Western Blotting
4.4. siRNA Injection to Rats
4.5. Measurement of Systolic Blood Pressure and Heart Rate
4.6. Isolation of PVN
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CCRL2 | Chemokine (CC motif) receptor-like 2 |
| CMKLR1 | Chemokine-like receptor 1 |
| GABA | γ-aminobutyric acid |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GPR1 | G protein-receptor 1 |
| i.c.v. | Intracerebroventricular |
| LTCC | L-type calcium channel |
| NMDA | N-Methyl-D-aspartate |
| PVN | Paraventricular nucleus |
| PKC | Protein kinase C |
| RVLM | Rostral ventrolateral medulla |
| SHR | Spontaneously hypertensive rats |
| siRNA | Small interfering RNA |
| SNA | Sympathetic nerve activity |
| WKY | Wistar Kyoto rats |
References
- Yamawaki, H. Vascular effects of novel adipocytokines: Focus on vascular contractility and inflammatory responses. Biol. Pharm. Bull. 2011, 34, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes. Rev. 2013, 14, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Allen, S.J.; Kulig, P.; Allen, J.A.; Cichy, J.; Handel, T.M.; Butcher, E.C. Chemerin activation by serine proteases of the coagulation, fibrinolytic, and inflammatory cascades. J. Biol. Chem. 2005, 280, 34661–34666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helfer, G.; Wu, Q.F. Chemerin: A multifaceted adipokine involved in metabolic disorders. J. Endocrinol. 2018, 238, R79–R94. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.-g.; Song, S.-H.; Choi, K.-C.; Katoh, K.; Wittamer, V.; Parmentier, M.; Sasaki, S.-i. Chemerin—A new adipokine that modulates adipogenesis via its own receptor. Biochem. Biophys. Res. Commun. 2007, 362, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Wittamer, V.; Franssen, J.D.; Vulcano, M.; Mirjolet, J.F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.H.; Lee, S.K.; Song, N.Y.; Son, S.H.; Kim, K.R.; Chung, W.Y. Chemerin treatment inhibits the growth and bone invasion of breast cancer cells. Int. J. Mol. Sci. 2020, 21, 2871. [Google Scholar] [CrossRef] [Green Version]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef] [Green Version]
- Kostopoulos, C.G.; Spiroglou, S.G.; Varakis, J.N.; Apostolakis, E.; Papadaki, H.H. Chemerin and CMKLR1 expression in human arteries and periadventitial fat: A possible role for local chemerin in atherosclerosis? BMC Cardiovasc. Disord. 2014, 14, 56. [Google Scholar] [CrossRef]
- Kunimoto, H.; Kazama, K.; Takai, M.; Oda, M.; Okada, M.; Yamawaki, H. Chemerin promotes the proliferation and migration of vascular smooth muscle and increases mouse blood pressure. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1017–H1028. [Google Scholar] [CrossRef] [Green Version]
- Weng, C.; Shen, Z.; Li, X.; Jiang, W.; Peng, L.; Yuan, H.; Yang, K.; Wang, J. Effects of chemerin/CMKLR1 in obesity-induced hypertension and potential mechanism. Am. J. Transl. Res. 2017, 9, 3096–3104. [Google Scholar]
- De Henau, O.; Degroot, G.N.; Imbault, V.; Robert, V.; De Poorter, C.; McHeik, S.; Galés, C.; Parmentier, M.; Springael, J.Y. Signaling properties of chemerin receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Kennedy, A.J.; Davenport, A.P. International union of basic and clinical pharmacology CIII: Chemerin receptors CMKLR1 (Chemerin1) and GPR1 (Chemerin2) nomenclature, pharmacology, and function. Pharmacol. Rev. 2018, 70, 174–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirooka, Y.; Sunagawa, K. Hypertension, heart failure, and the sympathetic nervous system: From the regulatory abvormality as the pathophysiology to novel therapeutic aspects. Fukuoka Igaku Zasshi Hukuoka Acta Med. 2010, 101, 190–197. [Google Scholar]
- Kubo, T. Mechanisms of hypertension in the central nervous system. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2006, 126, 695–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, K.; Hiyama, T.Y.; Sakuta, H.; Matsuda, T.; Lin, C.H.; Kobayashi, K.; Kobayashi, K.; Kuwaki, T.; Takahashi, K.; Matsui, S.; et al. [Na+] Increases in Body Fluids Sensed by Central Na x Induce Sympathetically Mediated Blood Pressure Elevations via H + -Dependent Activation of ASIC1a. Neuron 2019, 101, 60–75.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshimizu, T.-a.; Nakamura, K.; Egashira, N.; Hiroyama, M.; Nonoguchi, H.; Tanoue, A. Vasopressin V1a and V1b receptors: From molecules to physiological systems. Physiol. Rev. 2012, 92, 1813–1864. [Google Scholar] [CrossRef] [Green Version]
- Kc, P.; Dick, T. Modulation of cardiorespiratory function mediated by the paraventricular nucleus. Respir. Physiol. Neurobiol. 2010, 174, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Ding, L.; Dai, H.; Wang, F.; Zhou, H.; Gao, Q.; Xiong, X.; Zhang, F.; Song, T.; Yuan, Y.; et al. Intermedin in paraventricular nucleus attenuates ang ii-induced sympathoexcitation through the inhibition of nadph oxidase-dependent ros generation in obese rats with hypertension. Int. J. Mol. Sci. 2019, 20, 4217. [Google Scholar] [CrossRef] [Green Version]
- Dampney, R.A.; Michelini, L.C.; Li, D.P.; Pan, H.L. Regulation of sympathetic vasomotor activity by the hypothalamic paraventricular nucleus in normotensive and hypertensive states. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1200–H1214. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Wong, J.; Brooks, V.L. Obesity: Sex and sympathetics. Biol. Sex Differ. 2020, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Rhea, E.M.; Salameh, T.S.; Logsdon, A.F.; Hanson, A.J.; Erickson, M.A.; Banks, W.A. Blood-Brain Barriers in Obesity. AAPS J. 2017, 19, 921–930. [Google Scholar] [CrossRef]
- Yamamoto, A.; Matsumoto, K.; Hori, K.; Kameshima, S.; Yamaguchi, N.; Okada, S.; Okada, M.; Yamawaki, H. Acute intracerebroventricular injection of chemerin-9 increases systemic blood pressure through activating sympathetic nerves via CMKLR1 in brain. Pflugers Arch. Eur. J. Physiol. 2020, 472, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, S.-R.; Chen, H.; Pan, H.-L. Endogenous AT1 Receptor–Protein Kinase C Activity in the Hypothalamus Augments Glutamatergic Input and Sympathetic Outflow in Hypertension. J. Physiol. 2019, 597, 4325–4340. [Google Scholar] [CrossRef] [PubMed]
- Estienne, A.; Ramé, C.; Ganier, P.; Chahnamian, M.; Barbe, A.; Grandhaye, J.; Dubois, J.P.; Batailler, M.; Migaud, M.; Lecompte, F.; et al. Chemerin impairs food intake and body weight in chicken: Focus on hypothalamic neuropeptides gene expression and AMPK signaling pathway. Gen. Comp. Endocrinol. 2021, 304, 113721. [Google Scholar] [CrossRef] [PubMed]
- Sell, H.; Divoux, A.; Poitou, C.; Basdevant, A.; Bouillot, J.L.; Bedossa, P.; Tordjman, J.; Eckel, J.; Clément, K. Chemerin correlates with markers for fatty liver in morbidly obese patients and strongly decreases after weight loss induced by bariatric surgery. J. Clin. Endocrinol. Metab. 2010, 95, 2892–2896. [Google Scholar] [CrossRef] [Green Version]
- Zabel, B.A.; Zuniga, L.; Ohyama, T.; Allen, S.J.; Cichy, J.; Handel, T.M.; Butcher, E.C. Chemoattractants, extracellular proteases, and the integrated host defense response. Exp. Hematol. 2006, 34, 1021–1032. [Google Scholar] [CrossRef]
- Chou, H.H.; Teng, M.S.; Hsu, L.A.; Er, L.K.; Wu, S.; Ko, Y.L. Circulating chemerin level is associated with metabolic, biochemical and haematological parameters—A population-based study. Clin. Endocrinol. 2021, 94, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Scalco, A.; Moro, N.; Mongillo, M.; Zaglia, T. Neurohumoral Cardiac Regulation: Optogenetics Gets Into the Groove. Front. Physiol. 2021, 12, 1323. [Google Scholar] [CrossRef]
- Brunetti, L.; Di Nisio, C.; Recinella, L.; Chiavaroli, A.; Leone, S.; Ferrante, C.; Orlando, G.; Vacca, M. Effects of vaspin, chemerin and omentin-1 on feeding behavior and hypothalamic peptide gene expression in the rat. Peptides 2011, 32, 1866–1871. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, N.; Ding, Y.; Zhang, Y.; Li, Q.; Flores, J.; Haghighiabyaneh, M.; Doycheva, D.; Tang, J.; Zhang, J.H. Chemerin suppresses neuroinflammation and improves neurological recovery via CaMKK2/AMPK/Nrf2 pathway after germinal matrix hemorrhage in neonatal rats. Brain. Behav. Immun. 2018, 70, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Helfer, G.; Ross, A.W.; Thomson, L.M.; Mayer, C.D.; Stoney, P.N.; McCaffery, P.J.; Morgan, P.J. A neuroendocrine role for chemerin in hypothalamic remodelling and photoperiodic control of energy balance. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.D.; Holmberg, C.; Kandola, S.; Steele, I.; Hegyi, P.; Tiszlavicz, L.; Jenkins, R.; Beynon, R.J.; Peeney, D.; Giger, O.T.; et al. Increased expression of chemerin in squamous esophageal cancer myofibroblasts and role in recruitment of mesenchymal stromal cells. PLoS ONE 2014, 9, e104877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.Q.; Liu, X.; Zhang, G.; Parelkar, N.K.; Arora, A.; Haines, M.; Fibuch, E.E.; Mao, L. Phosphorylation of Glutamate Receptors: A Potential Mechanism for the Regulation of Receptor Function and Psychostimulant Action. J. Neurosci. Res. 2006, 84, 1621–1629. [Google Scholar] [CrossRef]
- Yabuki, Y.; Wu, L.; Fukunaga, K. Cognitive enhancer ST101 improves schizophrenia-like behaviors in neonatal ventral hippocampus-lesioned rats in association with improved CaMKII/PKC pathway. J. Pharmacol. Sci. 2019, 140, 263–272. [Google Scholar] [CrossRef]
- Chen, B.S.; Roche, K.W. Regulation of NMDA receptors by phosphorylation. Neuropharmacology 2007, 53, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.H.; Chen, S.R.; Wang, L.; Huang, Y.; Deng, M.; Zhang, J.; Zhang, J.; Chen, H.; Yan, J.; Pan, H.L. Protein kinase C-mediated phosphorylation and a2d-1 interdependently regulate NMDA receptor trafficking and activity. J. Neurosci. 2021, 41, 6415–6429. [Google Scholar] [CrossRef]
- Lyu, Y.; Ren, X.K.; Zhang, H.F.; Tian, F.J.; Mu, J.B.; Zheng, J.P. Sub-chronic administration of benzo[a]pyrene disrupts hippocampal long-term potentiation via inhibiting CaMK II/PKC/PKA-ERK-CREB signaling in rats. Environ. Toxicol. 2020, 35, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, M.W.; Oliet, S.H.R.; Panatier, A. NMDARs, coincidence detectors of astrocytic and neuronal activities. Int. J. Mol. Sci. 2021, 22, 7258. [Google Scholar] [CrossRef]
- He, L.; Hu, X.-T.; Lai, Y.-J.; Long, Y.; Liu, L.; Zhu, B.-l.; Chen, G.-J. Regulation and the Mechanism of Estrogen on Cav1.2 Gene in Rat-Cultured Cortical Astrocytes. J. Mol. Neurosci. 2016, 60, 205–213. [Google Scholar] [CrossRef]
- Min, R.; Santello, M.; Nevian, T. The computational power of astrocyte mediated synaptic plasticity. Front. Comput. Neurosci. 2012, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Ferland, D.J.; Darios, E.S.; Neubig, R.R.; Sjögren, B.; Truong, N.; Torres, R.; Dexheimer, T.S.; Thompson, J.M.; Watts, S.W. Chemerin-induced arterial contraction is Gi- and calcium- dependent. Vasc. Pharmacol. 2017, 88, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cookson, T.A. Bacterial-Induced Blood Pressure Reduction: Mechanisms for the Treatment of Hypertension via the Gut. Front. Cardiovasc. Med. 2021, 8, 721393. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Muñoz, R.; Algieri, F.; Vezza, T.; Jiménez, R.; Gálvez, J.; et al. Changes to the gut microbiota induced by losartan contributes to its antihypertensive effects. Br. J. Pharmacol. 2020, 177, 2006–2023. [Google Scholar] [CrossRef]
- Dranse, H.J.; Zheng, A.; Comeau, A.M.; Langille, M.G.I.; Zabel, B.A.; Sinal, C.J. The impact of chemerin or chemokine-like receptor 1 loss on the mouse gut microbiome. PeerJ 2018, 6, e5494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, Y.; Otani, K.; Sekiguchi, A.; Kogane, T.; Kakuda, C.; Sakamoto, Y.; Kodama, T.; Okada, M.; Yamawaki, H. Vasculo-protective effect of BMS-309403 is independent of its specific inhibition of fatty acid-binding protein 4. Pflugers Arch. Eur. J. Physiol. 2017, 469, 1177–1188. [Google Scholar] [CrossRef]
- Otani, K.; Yokoya, M.; Kodama, T.; Hori, K.; Matsumoto, K.; Okada, M.; Yamawaki, H. Plasma exosomes regulate systemic blood pressure in rats. Biochem. Biophys. Res. Commun. 2018, 503, 776–783. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, A.; Otani, K.; Okada, M.; Yamawaki, H. Chemokine-like Receptor 1 in Brain of Spontaneously Hypertensive Rats Mediates Systemic Hypertension. Int. J. Mol. Sci. 2021, 22, 11812. https://doi.org/10.3390/ijms222111812
Yamamoto A, Otani K, Okada M, Yamawaki H. Chemokine-like Receptor 1 in Brain of Spontaneously Hypertensive Rats Mediates Systemic Hypertension. International Journal of Molecular Sciences. 2021; 22(21):11812. https://doi.org/10.3390/ijms222111812
Chicago/Turabian StyleYamamoto, Atsunori, Kosuke Otani, Muneyoshi Okada, and Hideyuki Yamawaki. 2021. "Chemokine-like Receptor 1 in Brain of Spontaneously Hypertensive Rats Mediates Systemic Hypertension" International Journal of Molecular Sciences 22, no. 21: 11812. https://doi.org/10.3390/ijms222111812
APA StyleYamamoto, A., Otani, K., Okada, M., & Yamawaki, H. (2021). Chemokine-like Receptor 1 in Brain of Spontaneously Hypertensive Rats Mediates Systemic Hypertension. International Journal of Molecular Sciences, 22(21), 11812. https://doi.org/10.3390/ijms222111812