
Chronic Effects of Imidacloprid on Honey Bee Worker Development—Molecular Pathway Perspectives



Abstract
:1. Introduction
2. The Effects of Sublethal Dosages of Imidacloprid Exposure during the Adult Stage from a Molecular Perspective
3. The Effects of Sublethal Dosages of Imidacloprid Exposure during the Larval Stage from a Molecular Perspective
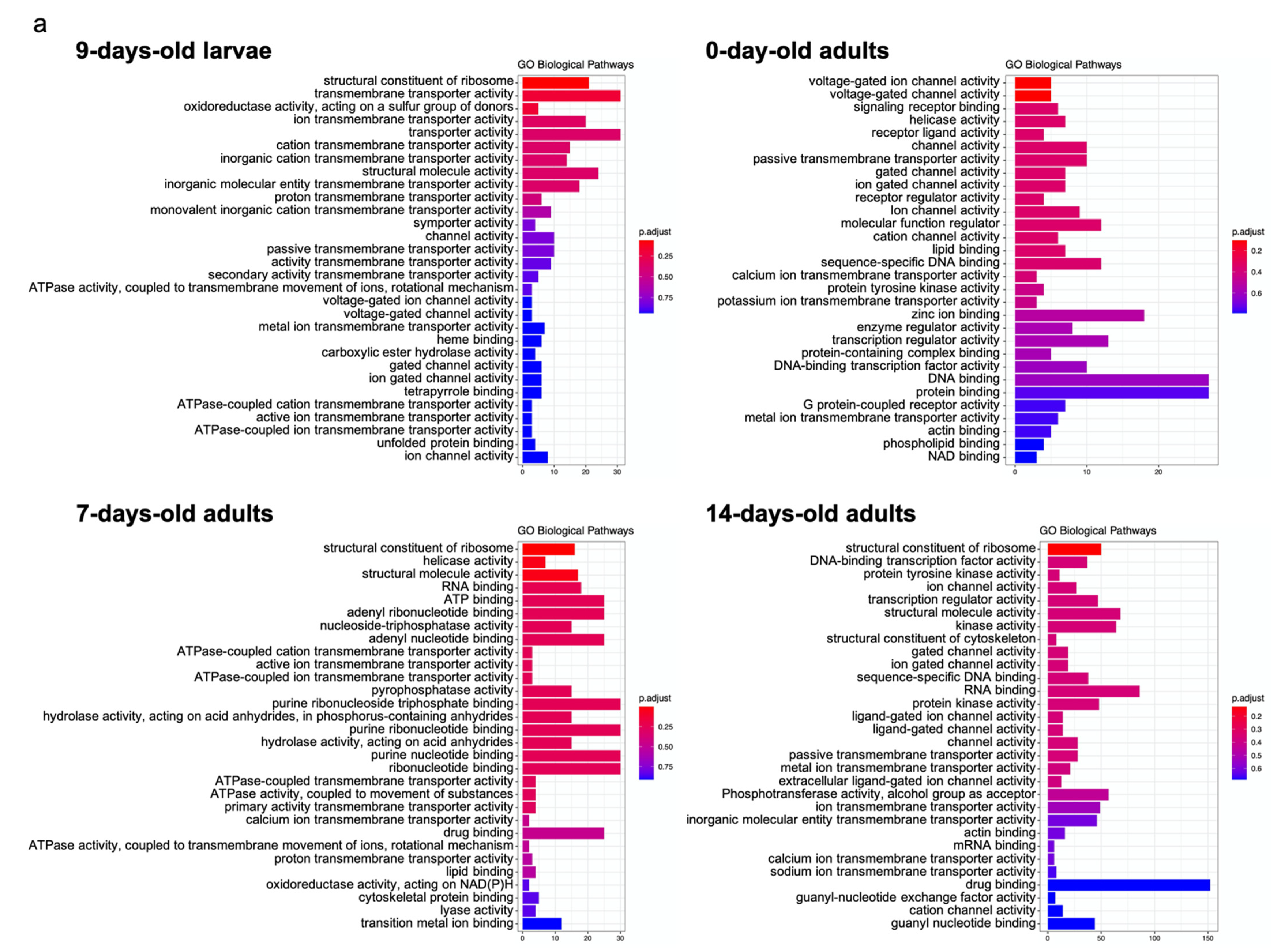
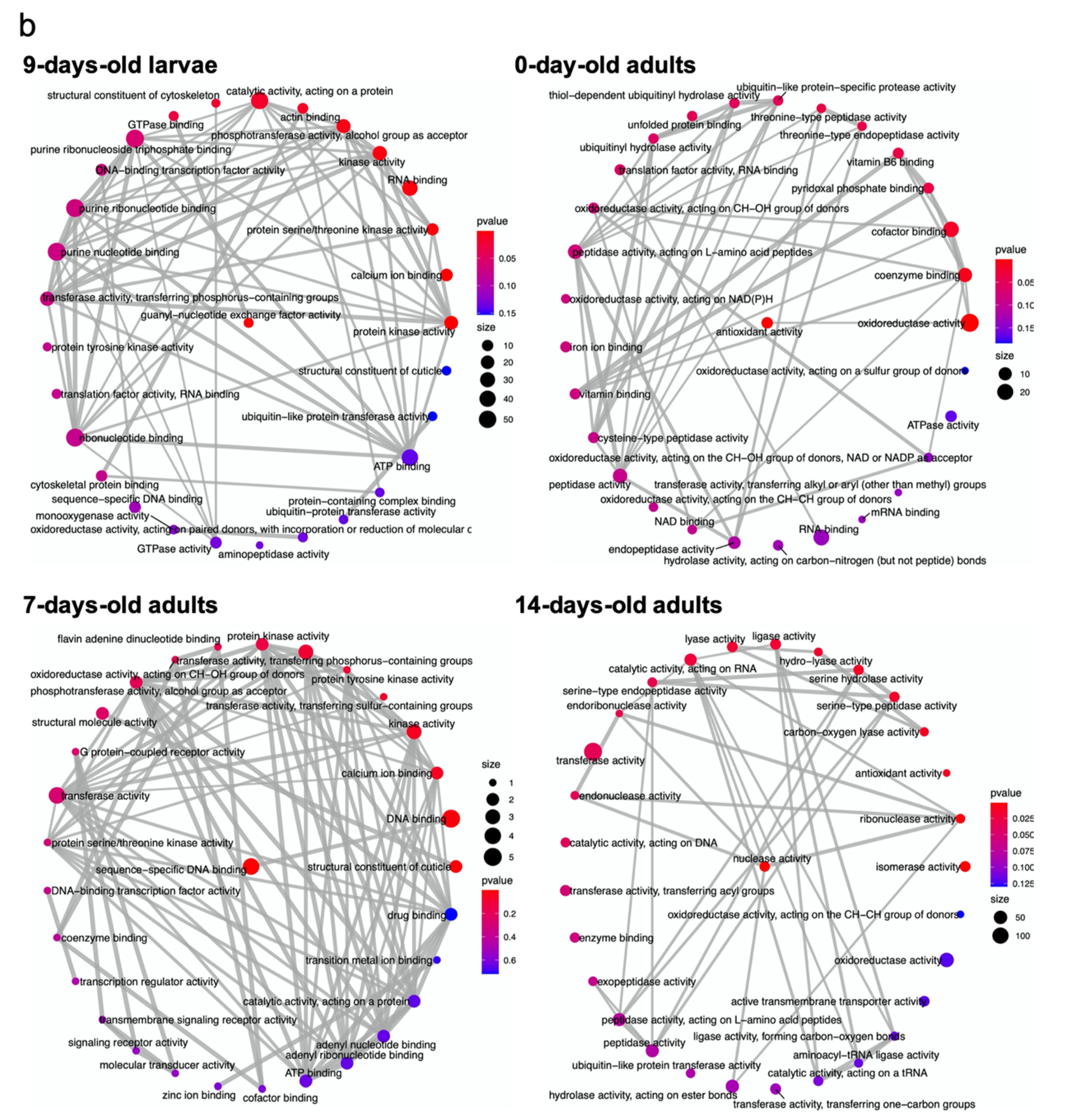
4. The Affected Molecular Pathways at Different Ages of Honey Bee Workers
5. The Molecular Effect of Imidacloprid on Other Pollinators
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schmuck, R. No causal relationship between Gaucho® seed dressing in sunflowers and the French bee syndrome. Pflanzenschutz Nachr. Bayer 1999, 52, 257–299. [Google Scholar]
- Schmuck, R.; Schöning, R.; Stork, A.; Schramel, O. Risk posed to honeybees (Apis mellifera L., Hymenoptera) by an imidacloprid seed dressing of sunflowers. Pest Manag. Sci. 2001, 57, 225–238. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Buckingham, S.; Lapied, B.; Corronc, H.; Sattelle, F. Imidacloprid actions on insect neuronal acetylcholine receptors. J. Exp. Biol. 1997, 200, 2685–2692. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Matsuda, K.; Shimomura, M.; Ihara, M.; Akamatsu, M.; Sattelle, D.B. Neonicotinoids show selective and diverse actions on their nicotinic receptor targets: Electrophysiology, molecular biology, and receptor modeling studies. Biosci. Biotechnol. Biochem. 2005, 69, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 2012, 491, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulson, D. Review: An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Lu, C.; Warchol, K.M.; Callahan, R.A. Sub-lethal exposure to neonicotinoids impaired honey bees winterization before proceeding to colony collapse disorder. Bull. Insectol. 2014, 67, 125–130. [Google Scholar]
- Sandrock, C.; Tanadini, M.; Tanadini, L.G.; Fauser-Misslin, A.; Potts, S.G.; Neumann, P. Impact of chronic neonicotinoid exposure on honeybee colony performance and queen supersedure. PLoS ONE 2014, 9, e103592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goñalongs, C.M.; Farina, W.M. Effects of sublethal doses of imidacloprid on young adult honeybee behaviour. PLoS ONE 2015, 10, e0140814. [Google Scholar]
- Woodcock, B.; Isaac, N.; Bullock, J.; Roy, D.B.; Garthwaite, D.G.; Crowe, A.; Pywell, R.F. Impacts of neonicotinoid use on long-term population changes in wild bees in England. Nat. Commun. 2016, 7, 12459. [Google Scholar] [CrossRef] [Green Version]
- Forfert, N.; Troxler, A.; Retschnig, G.; Gauthier, L.; Straub, L.; Moritz, R.F.A.; Neumann, P.; Williams, G.R. Neonicotinoid pesticides can reduce honeybee colony genetic diversity. PLoS ONE 2017, 12, e0186109. [Google Scholar] [CrossRef] [Green Version]
- Pisa, L.; Goulson, D.; Yang, E.C.; Gibbons, D.; Sánchez-Bayo, F.; Mitchell, E.; Aebi, A.; van der Sluijs, J.; MacQuarrie, C.; Giorio, C.; et al. An update of the Worldwide Integrated Assessment (WIA) on systemic insecticides. Part 2: Impacts on organisms and ecosystems. Environ. Sci. Pollut. Res. 2021, 28, 11749–11797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codling, G.; Naggar, Y.A.; Giesy, J.P.; Robertson, A.J. Concentrations of neonicotinoid insecticides in honey, pollen and honey bees (Apis mellifera) in central Saskatchewan, Canada. Chemosphere 2016, 144, 2321–2328. [Google Scholar] [CrossRef]
- Chauzat, M.P.; Faucon, J.P.; Martel, A.C.; Lachaize, J.; Cougoule, N.; Aubert, M. A survey of pesticide residues in pollen loads collected by honey bees in France. J. Econ. Entomol. 2006, 99, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Škerl, M.I.S.; Bolta, Š.V.; Česnik, H.B.; Gregorc, A. Residues of pesticides in honeybee (Apis mellifera carnica) bee bread and in pollen loads from treated apple orchards. Bull. Environ. Contam. Toxicol. 2009, 83, 374–377. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Mulhauser, B.; Mulot, M.; Mutabazi, A.; Glauser, G.; Aebi, A. A worldwide survey of neonicotinoids in honey. Science 2017, 358, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Böhme, F.; Bischoff, G.; Zebitz, C.P.W.; Rosenkranz, P.; Wallner, K. Pesticide residue survey of pollen loads collected honeybees (Apis mellifera) in daily internals at three agricultural sites in South Germany. PLoS ONE 2018, 13, e0199995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.J.; Kaplan, I.; Zhang, Y.; Szendrei, Z. Honeybee dietary neonicotinoid exposure is associated with pollen collection from agricultural weeds. Proc. R. Soc. B 2019, 286, 20190989. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A. Neonicotinoid insecticides. Banned by the European Union in 2018 after scientific studies concluded that harm honey bees. Sci. Rev. 2018, 37, 1–20. [Google Scholar]
- Laurino, D.; Manino, A.; Patetta, A.; Porporato, M. Toxicity of neonicotinoids insecticides on different honey bee genotype. Bull. Insectol. 2013, 66, 119–126. [Google Scholar]
- Suchail, S.; Guez, D.; Belzunces, L. Characteristics of imidacloprid toxicity in two Apis mellifera species. Environ. Toxicol. Chem. 2000, 19, 1901–1905. [Google Scholar] [CrossRef] [Green Version]
- Suchail, S.; Guez, D.; Belzunces, L.P. Discrepancy between acute and chronic toxicity induced by imidacloprid and its metabolites in Apis mellifera. Environ. Toxicol. Chem. 2001, 20, 2482–2486. [Google Scholar] [CrossRef]
- Nauen, R.; Ebbinghaus-Kintscherm, U.; Schmuck, R. Toxicity and nicotinic acetylcholine receptor interaction of imidacloprid and its metabolites in Apis mellifera (Hymenoptera: Apidae). Pest Manag. Sci. 2001, 57, 577–586. [Google Scholar] [CrossRef]
- Abbassy, M.A.; Nasr, H.M.; Abo-yousef, H.M.; Dawood, R.R. Acute toxicity of selected insecticides and their safety to honey bee (Apis mellifera L.) workers under laboratory conditions. Austin Environ. Sci. 2020, 5, 2. [Google Scholar]
- Dai, P.; Jack, C.J.; Mortensen, A.N.; Ellis, J.D. Acute toxicity of five pesticides to Apis mellifera larvae reared in vitro. Pest Manag. Sci. 2017, 73, 2282–2286. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.S.; Huang, Z.Y.; Milbrath, M.O. Neonicotinoid pesticides are more toxic to honey bees at lower temperatures: Implications for overwintering bees. Front. Ecol. Evol. 2020, 8, 556856. [Google Scholar] [CrossRef]
- Milone, J.P.; Rinkevich, F.D.; McAfee, A.; Foster, L.J.; Tarpy, D.R. Differences in larval pesticide tolerance and esterase activity across honey bee (Apis mellifera) stocks. Ecotoxicol. Environ. Saf. 2020, 206, 111213. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, F.D.; Margotta, J.W.; Pittman, J.M.; Danka, R.G.; Tarver, M.R.; Ottea, J.A.; Healy, K.B. Genetics, synergists, and age affect insecticide sensitivity of the honey bee, Apis mellifera. PLoS ONE 2015, 10, e0139841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the molecular determinants of bee sensitive to neonicotinoid insecticides. Curr. Biol. 2018, 28, 1137–1143.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, P.; Elias, M.; Silva-Neto, C.; Sujii, E.; Silva, D.; Gonçalves, B.; Franceschinelli, E. The effects of field-realistic doses of imidacloprid on Melipona quadrifasciata (Apidae: Meliponini) workers. Environ. Sci. Pollut. Res. 2020, 27, 38654–38661. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Genecque, E.; Menach, K.L.; Budzinski, H.; Cluzeau, S.; Pham-Delègue, M.H. Comparative sublethal toxicity of nine pesticides on olfactory learning performances of the honeybee Apis mellifera. Arch. Environ. Contam. Toxicol. 2005, 48, 242–250. [Google Scholar] [CrossRef]
- Dively, G.P.; Embrey, M.S.; Kamel, A.; Hawthorne, D.J.; Pettis, J.S. Assessment of chronic sublethal effects of imidacloprid on honey bee colony health. PLoS ONE 2015, 10, e0118748. [Google Scholar]
- Lawrence, T.J.; Culbert, E.M.; Felsot, A.S.; Hebert, V.R.; Sheppard, W.S. Survery and risk assessment of Apis mellifera (Hymenoptera: Apidea) exposure to neonicotinoid pestocodes in urban, rural, and agricultural settings. J. Econ. Entomol. 2016, 109, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Jepson, P.C.; Mayer, D.F. Limitation to use of topical toxicity data for prediction of pesticide side effect in the field. J. Econ. Entomol. 1995, 88, 1081–1088. [Google Scholar] [CrossRef]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- Decourtye, A.; Lacassie, E.; Pham-Delègue, M.H. Learning performances of honeybees (Apis mellifera) are differentially affected by imidacloprid according to the season. Pest Manag. Sci. 2003, 59, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Decourtye, A.; Devillers, J. Ecotoxicity of neonicotinoid insecticides to bees. In Insect Nicotinic Acetylcholine Receptors, 1st ed.; Thany, S.H., Ed.; Springer: New York, NY, USA, 2010; pp. 85–95. [Google Scholar]
- Yang, E.C.; Chang, H.C.; We, W.Y.; Chen, Y.W. Impaired olfactory associative behavior of honeybee workers due to contamination of imidacloprid in the larval stage. PLoS ONE 2012, 7, e49472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decourtye, A.; Armengaud, C.; Renou, M.; Devillers, J.; Cluzeau, S.; Gauthier, M.; Pham-Delegue, M.H. Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pestic. Biochem. Physiol. 2004, 78, 83–92. [Google Scholar] [CrossRef]
- Medrzycki, P.; Montanari, R.; Bortolotti, L.; Maini, S.; Porrini, C. Effects of imidacloprid administered in sub-lethal doses on honey bee behaviour. Laboratory tests. Bull. Insectol. 2003, 56, 59–62. [Google Scholar]
- Forfert, N.; Moritz, R.F.A. Thiacloprid alters social interactions among honey bee workers (Apis mellifera). J. Apic. Res. 2017, 56, 467–474. [Google Scholar] [CrossRef]
- Hatjina, F.; Papaefthimiou, C.; Charistos, L.; Dogaroglu, T.; Bouga, M.; Emmanouil, C.; Arnold, G. Sublethal doses of imidacloprid decreased size of hypopharyngeal glands and respiratory rhythm of honeybees in vivo. Apidologie 2013, 44, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Decourtye, A.; Devillers, J.; Cluzeau, S.; Charreton, M.; Pham-Delègue, M.H. Effects of imidacloprid and deltamethrin on associative learning in honeybees under semi-field and laboratory conditions. Ecotoxicol. Environ. Saf. 2004, 57, 410–419. [Google Scholar] [CrossRef]
- Faucon, J.P.; Aurières, C.; Drajnudel, P.; Mathieu, L.; Ribière, M.; Martel, A.-C.; Zeggane, S.; Chauzat, M.-P.; Aubert, M.F.A. Experimental study on the toxicity of imidacloprid given in syrup to honey bee (Apis mellifera) colonies. Pest Manag. Sci. 2005, 6, 111–125. [Google Scholar] [CrossRef]
- Yang, E.C.; Chuang, Y.C.; Chen, Y.L.; Chang, L.H. Abnormal foraging behavior induced by sublethal dosage of imidacloprid in the honey bee. J. Econ. Entomol. 2008, 101, 1743–1748. [Google Scholar] [CrossRef]
- Schneider, C.W.; Tautz, J.; Grünewald, B.; Fuchs, S. RFID tracking of sublethal effects of two neonicotinoid insecticides on the foraging behavior of Apis mellifera. PLoS ONE 2012, 7, e30023. [Google Scholar] [CrossRef] [Green Version]
- Williamson, S.M.; Wright, G.A. Exposure to multiple cholinergic pesticides impairs olfactory learning and memory in honeybees. J. Exp. Biol. 2013, 216, 083931. [Google Scholar] [CrossRef] [Green Version]
- Karahan, A.; Çakmak, I.; Hranitz, J.; Karaca, I.; Wells, H. Sublethal imidacloprid effects on honey bee flower choices when foraging. Ecotoxicology 2015, 24, 2017–2025. [Google Scholar] [CrossRef]
- Zhang, E.; Nieh, J.C. The neonicotinoid imidacloprid impairs honey bee aversive learning of simulated predation. J. Exp. Biol. 2015, 218, 3199–3205. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, T.; Chen, Y.; Heerman, M.; He, J.; Huang, J.; Nie, H.; Su, S. Brain transcriptome of honey bees (Apis mellifera) exhibiting impaired olfactory learning induced by a sublethal dose of imidacloprid. Pestic. Biochem. Physiol. 2019, 156, 36–43. [Google Scholar] [CrossRef]
- Gooley, Z.C.; Gooley, A.C. Exposure to field realistic concentrations of imidacloprid at different ambient temperatures disrupts non-flight metabolic rate in honey bee (Apis mellifera) foragers. Bull. Insectol. 2020, 73, 161–170. [Google Scholar]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Brunet, J.-L.; Dussaubat, C.; Mondet, F.; Tchamitchan, S.; Cousin, M.; Brillard, J.; Baldy, A.; Belzunces, L.P.; le Conte, Y. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environ. Microbiol. 2010, 12, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Pettis, J.S.; van Engelsdorp, D.; Johnson, J.; Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften 2012, 99, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.C.; Yang, E.C. Sublethal dosage of imidacloprid reduces the microglomerular density of honey bee mushroom bodies. Sci. Rep. 2016, 6, 19298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alburaki, M.; Steckel, S.J.; Chen, D.; Mcdermott, E.; Weiss, M.; Skinner, J.A.; Kelly, H.; Lorenz, G.; Tarpy, D.R.; Meikle, W.G.; et al. Landscape and pesticide effects on honey bees: Forager survival and expression of acetylcholinesterase and brain oxidative genes. Apidologie 2017, 48, 556–571. [Google Scholar] [CrossRef] [Green Version]
- Chaimanee, V.; Evans, J.D.; Chen, Y.; Jackson, C.; Pettis, J.S. Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 2016, 89, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, G.; de Graaf, D.C. Stress indicator gene expression profiles, colony dynamics and tissue development of honey bees exposed to sub-lethal doses of imidacloprid in laboratory and field experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Alburaki, M.; Rinderer, N.; Sampson, B.; Knight, P.R.; Karim, S.; Adamczyk, J. Effects of coumaphos and imidacloprid on honey bee (Hymenoptera: Apidae) lifespan and antioxidant gene regulations in laboratory experiments. Sci. Rep. 2018, 8, 15003. [Google Scholar] [CrossRef]
- Aufauvre, J.; Misme-Aucouturier, B.; Viguès, B.; Texier, C.; Delbac, F.; Blot, N. Transcriptome Analyses of the Honeybee Response to Nosema ceranae and Insecticides. PLoS ONE 2014, 9, e91686. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Luo, Q.-H.; Hou, C.-S.; Wang, Q.; Dai, P.-L.; Gao, J.; Liu, Y.-J.; Diao, Q.-Y. Sublethal effects of imidacloprid on targeting muscle and ribosomal protein related genes in the honey bee Apis mellifera L. Sci. Rep. 2017, 7, 15943. [Google Scholar] [CrossRef] [PubMed]
- Christen, V.; Shirrmann, M.; Frey, J.E.; Fent, K. Global Transcriptomic Effects of Environmentally Relevant Concentrations of the Neonicotinoids Clothianidin, Imidacloprid, and Thiamethoxam in the Brain of Honey Bees (Apis mellifera). Environ. Sci. Technol. 2018, 52, 7534–7544. [Google Scholar] [CrossRef]
- Wu, M.C.; Chang, Y.W.; Lu, K.H.; Yang, E.C. Gene expression changes in honey bees induced by sublethal imidacloprid exposure during the larval stage. Insect Biochem. Mol. Biol. 2017, 88, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Tesovnik, T.; Zora, M.; Gregorc, A.; Rinehart, T.; Adamczyk, J.; Narat, M. Immune gene expression in developing honey bees (Apis mellifera L.) simultaneously exposed to imidacloprid and Varroa destructor in laboratory conditions. J. Apicul. Res. 2019, 58, 730–739. [Google Scholar] [CrossRef]
- Williams, G.; Troxler, A.; Retschnig, G.; Roth, K.; Yañez, O.; Shutler, D.; Neumann, P.; Gauthier, L. Neonicotinoid pesticides severely affect honey bee queens. Sci. Rep. 2015, 5, 14621. [Google Scholar] [CrossRef] [Green Version]
- Wu-Smart, J.; Spivak, M. Sub-lethal effects of dietary neonicotinoid insecticide exposure on honey bee queen fecundity and colony development. Sci. Rep. 2016, 6, 32108. [Google Scholar] [CrossRef]
- Vergara-Amado, J.; Manzi, C.; Franco, L.M.; Contecha, S.C.; Marquez, S.J.; Solano-Iguaran, J.J.; Haro, R.E.; Silva, A.X. Effects of residual doses of neonicotinoid (imidacloprid) on metabolic rate of queen honey bees Apis mellifera (Hymenoptera: Apidae). Apidologie 2020, 51, 1091–1099. [Google Scholar] [CrossRef]
- Woyciechowski, M.; Moroń, D. Life expectancy and onset of foraging in the honeybee (Apis mellifera). Insectes Soc. 2009, 56, 193–201. [Google Scholar] [CrossRef]
- Wu, J.Y.; Anelli, C.M.; Sheppard, W.S. Sub-lethal effects of pesticide residues in brood comb on worker honey bee (Apis mellifera) development and longevity. PLoS ONE 2011, 6, e14720. [Google Scholar] [CrossRef] [Green Version]
- Colin, T.; Meikle, W.G.; Wu, X.; Barron, A.B. Traces of a Neonicotinoid Induce Precocious Foraging and Reduce Foraging Performance in Honey Bees. Environ. Sci. Technol. 2019, 53, 8252–8261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippers, M.P.; Dukas, R.; Smith, R.W.; Wang, J.; Smolen, K.; McClellandet, G.B. Lifetime performance in foraging honeybees: Behaviour and physiology. J. Exp. Biol. 2006, 209, 3828–3836. [Google Scholar] [CrossRef] [Green Version]
- Vance, J.T.; Williams, J.B.; Elekonich, M.M.; Roberts, S.P. The effects of age and behavioral development on honey bee (Apis mellifera) flight performance. J. Exp. Biol. 2009, 212, 2604–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippers, M.P.; Dukas, R.; McClelland, G.B. Lifetime- and caste-specific changes in flight metabolic rate and muscle biochemistry of honeybees, Apis mellifera. J. Comp. Physiol. B 2010, 180, 45–55. [Google Scholar] [CrossRef]
- Perry, C.J.; Søvik, E.; Myerscough, M.R.; Arron, A.B. Behavioral Maturation Accelerates Failure of Stressed Honey Bee Colonies. Proc. Natl. Acad. Sci. USA 2015, 112, 3427–3432. [Google Scholar] [CrossRef] [Green Version]
- Ushitani, T.; Perry, C.J.; Cheng, K.; Barron, A.B. Accelerated behavioural development changes fine-scale search behaviour and spatial memory in honey bees (Apis mellifera L.). J. Exp. Biol. 2016, 219, 412–418. [Google Scholar]
- Chen, Y.R.; Tzeng, D.T.W.; Ting, C.; Hsu, P.S.; Wu, T.H.; Zhong, S.; Yang, E.C. Missins nurse bees—Early transcriptomic switch from nurse bee to forager induced by subltheal imidacloprid. Front. Genet. 2021, 12, 665927. [Google Scholar] [CrossRef]
- Belenky, P.; Racette, F.G.; Bogan, K.L.; McClure, J.M.; Smith, J.S.; Brenner, C. Nicotinamide riboside promotes sir2 silencing and extends lifespan via Nrk and Urh1/Pnp1/Meu1 pathways to NAD(+). Cell 2007, 129, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; Zhu, Y.; Liu, P.; Ye, L.; Jiang, X.; Cao, H.; Yu, L. Age and Behavior-Dependent Differential miRNAs Expression in the Hypopharyngeal Glands of Honeybees (Apis mellifera L.). Insects 2021, 12, 764. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, fat metabolism, and life span: What is the connection? Cell Metabol. 2013, 17, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walden, H.; Deans, A.J. The Fanconi anemia DNA repair pathway: Structural and functional insights into a complex disorder. Annu. Rev. Biophys. 2014, 43, 257–278. [Google Scholar] [CrossRef]
- Ben-Shahar, Y.; Leung, H.-T.; Pak, W.L.; Sokolowski, M.B.; Robinson, G.E. cGMP-dependent changes in phototaxis: A possible role for the foraging gene in honey bee division of labor. J. Exp. Biol. 2003, 206, 2507–2515. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological theme among gene clusters. OMICS J. Integrat. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Liu, S.; Bo, X.; Yu, G. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar]
- Seeley, T.D. Adaptive significance of the age polyethism schedule in honeybee colonies. Behav. Ecol. Sociobiol. 1982, 11, 287–293. [Google Scholar] [CrossRef]
- Paleolog, J.; Wilde, J.; Siuda, M.; Bąk, B.; Wójcik, Ł.; Strachecka, A. Imidacloprid markedly affects hemolymph proteolysis, biomarkers, DNA global methylation, and the cuticle proteolytic layer in western honeybees. Apidologie 2020, 51, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Colgan, T.J.; Fletcher, I.K.; Arce, A.N.; Gill, R.J.; Rodrigues, A.R.; Stolle, E.; Chittka, L.; Wurm, Y. Caste- and pesticide-specific effects of neonicotinoid pesticide exposure on gene expression in bumblebees. Mol. Ecol. 2019, 28, 1964–1974. [Google Scholar] [CrossRef] [Green Version]
- Bebane, P.S.A.; Hunt, J.H.; Pegoraro, M.; Jones, A.R.C.; Marchall, H.; Rosato, E.; Mallon, E.B. The effects of the neonicotinoid imidacloprid on gene expression and DNA methylation in the buff-tailed bumblebee Bumbus terrestris. Proc. R. Soc. B 2019, 286, 20190718. [Google Scholar] [CrossRef]
- Gao, J.; Jin, S.S.; He, Y.; Luo, J.H.; Xu, C.Q.; Wu, Y.Y.; Hou, C.S.; Wang, Q.; Diao, Q.Y. Physilogical analysis and transcriptome analysis of Asian honey bee (Apis cerana cerana) in response to sublethal neonicotinoid imidacloprid. Insect 2020, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Beadle, K.; Singh, K.S.; Troczka, B.J.; Randall, E.; Zaworra, M.; Zimmer, C.T.; Hayward, A.; Reid, R.; Kor, L.; Kohler, M.; et al. Genomic insights into neonicotinoid sensitivity in the solitary bee Osmia bicornis. PLoS Genet. 2019, 15, e1007903. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oral LD50a (ng Per Bee) | Contact LD50 (ng Per Bee) | Oral LC50a (ppm) | Test Period | Honey Bee | Tested Areas | Hours | Reference | |
|---|---|---|---|---|---|---|---|---|
| adults | NA | 40 | NA | NA | NA | USA | 24 | [37] |
| 5.4 (5.2–1.6) | 23.8 (22.3–25.3) | NA | NA | Apis mellifera mellifera | France | 24 | [24] | |
| 6.6 (5.1–8.1) | 15.1 (11.9–18.3) | Apis mellifera caucasica | ||||||
| 4.8 (4.5–5.1) | 24.3 (22.0–26.6) | NA | NA | Apis mellifera mellifera | 48 | |||
| 6.5 (4.7–8.3) | 12.8 (9.7–15.9) | Apis mellifera caucasica | ||||||
| 41 | NA | NA | Jul-99 | NA | Germany I | 72 | [26] | |
| 20 | 104 (83.0–130) | Jul-99 | The Netherlands I | |||||
| 81 | 61.0 (26.0–90.0) | May-00 | Germany II | |||||
| 81 | 50.0 (9.1–71.0) | May-00 | United Kingdom I | |||||
| 81 | 42.0 (20.0–59.0) | May-00 | Germany III | |||||
| 81 | 42.9 (34.6–53.2) | May-00 | Germany IV | |||||
| 81 | 74.9 (61.8–90.9) | Jul-00 | Germany V | |||||
| 57 ± 28 | NA | NA | NA | NA | France | 48 | [25] | |
| 37 ± 10 | 72 | |||||||
| 37 ± 10 | 96 | |||||||
| 3.7 (2.6–5.3) | 81 (55.0–119.0) | NA | NA | NA | UK | 48 | [2] | |
| > 21.0 | 230.3 | Netherlands | ||||||
| 40.9 | nt | Germany | ||||||
| 11.6 (7.3-18.3) | 242.6 (173.3–353.4) | |||||||
| 21.2 (15.0-29.6) | 59.7 (39.1–92.7) | |||||||
| NA | 17.9 (9.2–31.5) | NA | Jun to Sep-99 | NA | USA | 24 | [38] | |
| 30.6 | NA | NA | NA | NA | France | 48 | [39] | |
| 25.4 ± 22.8 | NA | NA | NA | NA | France | 48 | [40] | |
| 118.74 | NA | NA | NA | NA | Italy | 24 | [23] | |
| 90.09 | 48 | |||||||
| 69.68 | 72 | |||||||
| NA | 29 | 0.003 | NA | NA | Egypt | 24 | [27] | |
| 26 | 0.0006 | 48 | ||||||
| larvae | 4170 (2960–5850) | NA | 138.84 (98.20–196.30) | NA | NA | USA. | 72 | [28] |
| 1400 | NA | NA | NA | Taiwan | 268 | [41] | ||
| (a) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Imidacloprid | Treatment | Approaches | Treatment Stages | Volume | Affected Functions | Gene id/Name | Gene Regulatory Trend | Treatment Period (d) | Sampling d (d Post First Day of Treatment) | References | |||
| Treatment | Raised Environment | Up | Down | ||||||||||
| 20 ppb | feeding (syrup) | NA | adults | NA | NA | Lim 3 homeobix (down), | 0 | 1 | 7 | 7 | [63] | ||
| vanin-like protein 1-like (2) (down) | 0 | 1 | 11 | ||||||||||
| 20 ppb | topical treatment | caged | qPCR | adults | 1 μL | P450 superfamily genes | CYP6AS14 | 0 | 1 | 1 | 1 | [60] | |
| immunity | thioredoxin peroxidase, Amel/LRR | 0 | 1 | 1 | |||||||||
| eater, Amel/LRR | 0 | 2 | 7 | ||||||||||
| development and antioxidant enzyme-coding gene | Catalase (down), SOD (up) | 1 | 1 | 1 | |||||||||
| 5 ppb | feeding (pollen patty and sugar solution) | field | qPCR | adults | NA | immunity | Apisimin (down), Defensin 2 (down), Vitellogenin (up) | 1 | 2 | 10 | 10 | [61] | |
| caged | Abaecin (down), Apisimin (down), Defensin 1 (down), Defensin 2 (down) | 0 | 4 | ||||||||||
| field | Abaecin (down), Apisimin (up) | 1 | 1 | 20 | 20 | ||||||||
| caged | Abaecin (down), Defensin 1 (up), Vitellogenin (down) | 1 | 2 | ||||||||||
| 200 ppb | field | Defensin 2 (up), Vitellogenin (up) | 2 | 0 | 10 | 10 | |||||||
| caged | Abaecin (down), Apisimin (down), Defensin 1 (down), Defensin 2 (down), Vitellogenin (down) | 0 | 5 | ||||||||||
| field | Abaecin (up), Defensin 2 (up), Vitellogenin (up) | 3 | 20 | 20 | |||||||||
| caged | Abaecin (down), Apisimin (down) | 0 | 2 | ||||||||||
| 5 ppb | field | detoxification genes | AChE-1 (down), CYP6AS3 (down), CYP9Q3 (down) | 0 | 3 | 10 | 10 | ||||||
| caged | AChE-1 (down), AChE-2 (down), CYP6AS4 (down), CYP6AS10 (down), CYP9Q1 (down), CYP9Q2 (down), CYP9Q3 (down), CYT p450 (down) | 0 | 8 | ||||||||||
| field | AChE-2 (down), CYP6AS3 (down) | 0 | 2 | 20 | 20 | ||||||||
| caged | AChE-1 (down), CYP6AS3 (down), CYP6AS4 (down), CYP9Q1 (up), CYP9Q2 (down), CYP9Q3 (up), CYT p450 (up) | 3 | 4 | ||||||||||
| 200 ppb | field | AChE-2 (down), CYP6AS10 (down) | 0 | 2 | 10 | 10 | |||||||
| caged | AChE-1 (down), AChE-2 (down), CYP6AS4 (up), CYP6AS10 (down), CYP9Q1 (down), CYP9Q2 (down), CYP9Q3 (down) | 1 | 6 | ||||||||||
| field | AChE-1 (up), AChE-2 (up), CYP6AS3 (up), CYP9Q1 (up), CYP9Q2 (up), CYP9Q3 (up), CYT p450 (up) | 7 | 0 | 20 | 20 | ||||||||
| caged | AChE-2 (up), CYP6AS3 (down), CYP6AS10 (down), CYP9Q1 (up), CYP9Q2 (down), CYT p450 (down) | 2 | 4 | ||||||||||
| 5 ppb | sugar syrup | caged | aqPCR | adults | NA | detoxification enzyme | GstD1 | 1 | 0 | 7 weeks | 7 weeks | [59] | |
| 20 ppb | |||||||||||||
| 100 ppb | |||||||||||||
| 10 ppb | feeding, (sucrose solution, 50% wt/wt) | caged | NGS | adults | NA | total DEGs 509 | NA | 160 | 349 | 8 | 8 | [64] | |
| ribosomal protein | 28 | 0 | |||||||||||
| phototransduction | NA | ||||||||||||
| visual perception | |||||||||||||
| photoreceptor | |||||||||||||
| actin binding | |||||||||||||
| actin cytoskeleton | |||||||||||||
| muscle attachment | |||||||||||||
| somatic muscle development | |||||||||||||
| 5 ppb | feeding (food patty) | caged | qPCR | adults | NA | antioxidant genes | Cat (down), MsrA (down), TrxR1 (down) | 0 | 3 | 10 | 10 | [62] | |
| 10 ppb | antioxidant genes | Cat (up), TrxR1 (up), SelK (up), MsrB (up), Sod2 (down) | 4 | 1 | |||||||||
| 20 ppb | feeding (sucrose solution, 30% wt/wt) | caged | NGS | adults | NA | total DEGs | 131 | 1 | 130 | 11 | 11 | [53] | |
| chemosensory-related genes | GB46225, GB46227, GB46230, GB53372, GB16006, GB50003 | 0 | 6 | ||||||||||
| immune and detoxification response | abaecin (GB18323), apisimin (GB53576), defensin 1 (GB41428), glucose dehydrogenase (GB43007), glucose dehydrogenase-like (GB51446), leucine-rich repeat-containing protein 26-like (GB44192), phenoloxidase subunit A3 (GB43738), serine protease easter (GB45700) and tyrosine aminotransferase (GB45969) | 0 | 8 | ||||||||||
| insecticide resistance-related gene | cuticular protein 14 (GB46297), cytochrome b561 (GB40148), cytochrome P450 6a2 (GB49876), cytochrome P450 9e2 (GB43713), esterase A2 (GB43571), cytochrome P450 6a17 (GB49885) and UDP-glucuronosyltransferase 2C1 (GB52179) | 0 | 7 | ||||||||||
| oxidation-reduction | GB52785, GB44549, GB43007, GB55515, GB49876, GB50655, GB43713, GB51446, GB50178, GB49885 | 0 | 10 | ||||||||||
| iron ion | GB55515, GB49876, GB50655, GB43713, GB49885 | 0 | 5 | ||||||||||
| oxidoreductase activity | GB52785, GB44549, GB43007, GB55515, GB49876, GB50655, GB43713, GB51446, GB50178, GB49885 | 0 | 10 | ||||||||||
| behavioral response | SLC18A2 (GB50003), melanopsin (GB41643), aquaporin 4 (GB41240), PRKACB (GB48362) | 0 | 4 | ||||||||||
| phototransduction | GB41297, GB51068 | 0 | 2 | ||||||||||
| 0.3 ng/bee | sucrose solution | caged | NGS | adults | 100 μL/bee in average per 24 h | metabolic pathways | DN74754_c11_g2 (down) | 7 | 19 | 2 | 2 | [65] | |
| starch and sucrose metabolism | DN74754_c11_g2 (down) | ||||||||||||
| purine metabolism | DN75806_c0_g2 (down) | ||||||||||||
| 3 ng/bee | metabolic pathways | DN75371_c1_g1 (down), DN178528_c0_g1 (up), DN74754_c11_g2 (down) | 36 | 77 | |||||||||
| pentose phosphate pathway | DN75371_c1_g1 (down) | ||||||||||||
| purine metabolism | DN75806_c0_g2 (down) | ||||||||||||
| glycine, serine, and threoninemetabolism | DN178528_c0_g1 (up) | ||||||||||||
| porphyrin metabolism | DN178528_c0_g1 (up) | ||||||||||||
| starch and sucrose metabolism | DN74754_c11_g2 (down) | ||||||||||||
| (b) | |||||||||||||
| imidacloprid | treatment | Approaches | treatment stages | volume | affected functions | Gene ID/name | DEGs trend | treatment period (days) | sampling days (days post first day of treatment) | references | |||
| treatment | raised environment | up | down | ||||||||||
| 20 ppb | topical treatment | caged | qPCR | queen, adults | 2 μL | P450 superfamily genes | CYP306A1 (down) | 0 | 1 | 1 | 1 | [60] | |
| CYP4G11 (up), CYP6AS14 (down) | 1 | 1 | 7 | ||||||||||
| immunity | thioredoxin peroxidase (up), apidaecin (down), eater (down), Amel/LRR (down), VgMC (down) | 1 | 4 | 1 | |||||||||
| thioredoxin peroxidase (up), Amel/LRR (down), VgMC (down) | 1 | 2 | 7 | ||||||||||
| development and antioxidant enzyme-coding gene | SOD (down), hexamerin 70b (down) | 0 | 2 | 1 | |||||||||
| hexamerin 70b (down) | 0 | 1 | 7 | ||||||||||
| (c) | |||||||||||||
| imidacloprid | treatment | Approaches | treatment stages | collected stages | volume | affected functions | Gene ID/name | DEGs trend | treatment period (d) | sampling d (d post first day of treatment) | references | ||
| treatment | raised environment | up | down | ||||||||||
| 500 ppb | feeding (water) | field | NGS | larvae | adults | 1 μL | total DEGs | 578 | 4 | 21 | [66] | ||
| detoxification | GB48993 (up), GB49876 (up), GB43693 (up), GB44513 (up), GB55257 (up), GB49877 (up), GB49614 (up), GB48905 (down), GB49875 (down), GB40287 (down), GB43728 (down), GB49887 (down), GB52023 (down), GB43716 (down), GB43727 (down), GB51356 (down), GB47279 (down), GB49885 (down), GB55669 (down), GB49626 (down), GB46814 (down), GB43713 (down), GB49886 (down), GB43715 (down), | 7 | 17 | ||||||||||
| immunity | GB51223 (down), GB47318 (down), GB47546 (down), GB40164 (down), GB53576 (down) | 0 | 5 | ||||||||||
| mitochondria | GB44116 (up), GB42580 (up), GB50970 (up), GB42141 (up), GB49306 (up), GB46083 (up), GB53201 (up), GB49942 (down), GB47970 (down), GB42550 (down), GB51583 (down) | 7 | 4 | ||||||||||
| metabolism | GB42460 (up), GB55040 (up), GB54404 (up), GB49562 (down), GB51236 (down), GB54302 (down), GB49336 (down), GB48172 (down), GB53312 (down), GB51247 (down), GB51580 (down), GB51815 (down), GB45596 (down), GB53525 (down), GB54401 (down), GB48850 (down), GB49380 (down), GB51814 (down), GB53579 (down), GB54396 (down), GB43006 (down), GB44548 (down), GB43247 (down), GB53872 (down) | 3 | 21 | ||||||||||
| neuron development | GB50170 (up), GB50061 (up), GB42798 (up), GB41270 (up), GB49750 (up), GB47563 (up), GB47918 (up), GB55389 (up), GB41630 (up), GB47565 (up), GB51612 (up), GB52630 (up), GB46091 (up), GB49726 (up), GB41126 (down), GB52454 (down), GB41856 (down), GB43778 (down), GB40356 (down), GB43504 (down), GB49109 (down), GB43788 (down), GB49708 (down) | 4 | 19 | ||||||||||
| sensory processing | GB50936 (up), GB55547 (up), GB45850 (up), GB52326 (up), GB41643 (down), GB46229 (down), GB40616 (down), GB44550 (down), GB46225 (down),GB46230 (down), GB53367 (down), GB51369 (down), GB46224 (down), GB46228 (down), GB53368 (down), GB53372 (down), GB54970 (down),GB51189 (down) | 4 | 14 | ||||||||||
| signaling pathway | GB49937 (up), GB41862 (down), GB50705 (down), GB47566 (down), GB47961 (down), GB48815 (down), GB41220 (down), GB51125 (down) | 2 | 6 | ||||||||||
| structural protein | GB47903 (up), GB42581 (up), GB50453 (up), GB44002 (up), GB49021 (up), GB50438 (up), GB53565 (up), GB53119 (up), GB50236 (up), GB45073 (up), GB40566 (up), GB49845 (up), GB41227 (up), GB41203 (up), GB45174 (up), GB41311 (up), GB47902 (up), GB41310 (up), GB45968 (up), GB41308 (up), GB44214 (up), GB52194 (up), GB45211 (up), GB41946 (up), GB45943 (up), GB52014 (up), GB52161 (up), GB43173 (up), GB40253 (up), GB44074 (up), GB42571 (up), GB49219 (up), GB43377 (down), GB51391 (down), GB40304 (down), GB41757 (down) | 32 | 4 | ||||||||||
| transcription factors | GB50933 (up), GB52658 (up), GB50795 (down), GB42049 (down), GB52761 (down) | 2 | 3 | ||||||||||
| transporters and receptors | GB47513 (up), GB42142 (up), GB53053 (up), GB54918 (up), GB42802 (up), GB43870 (up), GB42865 (up), GB40973 (up), GB47391 (up), GB49727 (up), GB40867 (down), GB43672 (down), GB50423 (down), GB54881 (down), GB47942 (down), GB44824 (down), GB48790 (down), GB49801 (down), GB44987 (down), GB54942 (down), GB49396 (down), GB46030 (down), GB41240 (down), GB50098 (down), GB48330 (down), GB51487 (down), GB47278 (down), GB49473 (down), GB40818 (down), GB42942 (down), GB47931 (down), GB45986 (down), GB54467 (down), GB43963 (down), GB41182 (down), GB50890 (down), GB55098 (down), GB55239 (down), GB51504 (down), GB53124 (down), GB50457 (down), GB52097 (down), GB41815 (down), GB55503 (down), GB46597 (down), GB45159 (down) | 10 | 36 | ||||||||||
| 20 ppb | feeding (larvae diet) | caged | qPCR | larvae | white eye pupae | 10 μL | Toll pathway | Cactus (down), Dorsan (down) | 0 | 2 | 4 | 9 | [67] |
| IMD pathway | PGRP LC710 (down) | 0 | 1 | ||||||||||
| JNK pathway | Kayak (down) | 0 | 1 | ||||||||||
| Antimicrobial peptides | Abeacin (down) | 0 | 1 | ||||||||||
| Melanization | PPO (up) | 1 | 0 | ||||||||||
| brown eye pupae | Toll pathway | PGRP SC 4300 (down), Spaetzle (down), Cactus (down), Dorsan (down) | 0 | 4 | 12 | ||||||||
| JAK/STAT | Domless (down) | 0 | 1 | ||||||||||
| IMD pathway | PGRP LC710 (down), Relush (down) | 0 | 2 | ||||||||||
| JNK pathway | Basket (down), Kayak (down) | 0 | 2 | ||||||||||
| Antimicrobial peptides | Apidaecin (down), Hymenoptaecin (down), Defensin 1 (down) | 0 | 3 | ||||||||||
| Melanization | PPO (down) | 0 | 1 | ||||||||||
| newly emerged adults | Toll pathway | Cactuc (up) | 1 | 0 | 16 | ||||||||
| IMD pathway | Relish (up) | 1 | 0 | ||||||||||
| JNK pathway | Kayak (up) | 1 | 0 | ||||||||||
| Antimicrobial peptides | Hymenoptaecin (up), Lysozyme 2 (up) | 2 | 0 | ||||||||||
| Melanization | PPO (up) | 1 | 0 | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-R.; Tzeng, D.T.W.; Yang, E.-C. Chronic Effects of Imidacloprid on Honey Bee Worker Development—Molecular Pathway Perspectives. Int. J. Mol. Sci. 2021, 22, 11835. https://doi.org/10.3390/ijms222111835
Chen Y-R, Tzeng DTW, Yang E-C. Chronic Effects of Imidacloprid on Honey Bee Worker Development—Molecular Pathway Perspectives. International Journal of Molecular Sciences. 2021; 22(21):11835. https://doi.org/10.3390/ijms222111835
Chicago/Turabian StyleChen, Yun-Ru, David T. W. Tzeng, and En-Cheng Yang. 2021. "Chronic Effects of Imidacloprid on Honey Bee Worker Development—Molecular Pathway Perspectives" International Journal of Molecular Sciences 22, no. 21: 11835. https://doi.org/10.3390/ijms222111835
APA StyleChen, Y.-R., Tzeng, D. T. W., & Yang, E.-C. (2021). Chronic Effects of Imidacloprid on Honey Bee Worker Development—Molecular Pathway Perspectives. International Journal of Molecular Sciences, 22(21), 11835. https://doi.org/10.3390/ijms222111835