
Towards the Idea of Molecular Brains


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Undoubtedly, only artists devote themselves to science…(Santiago Ramon y Cajal)
1. Introduction
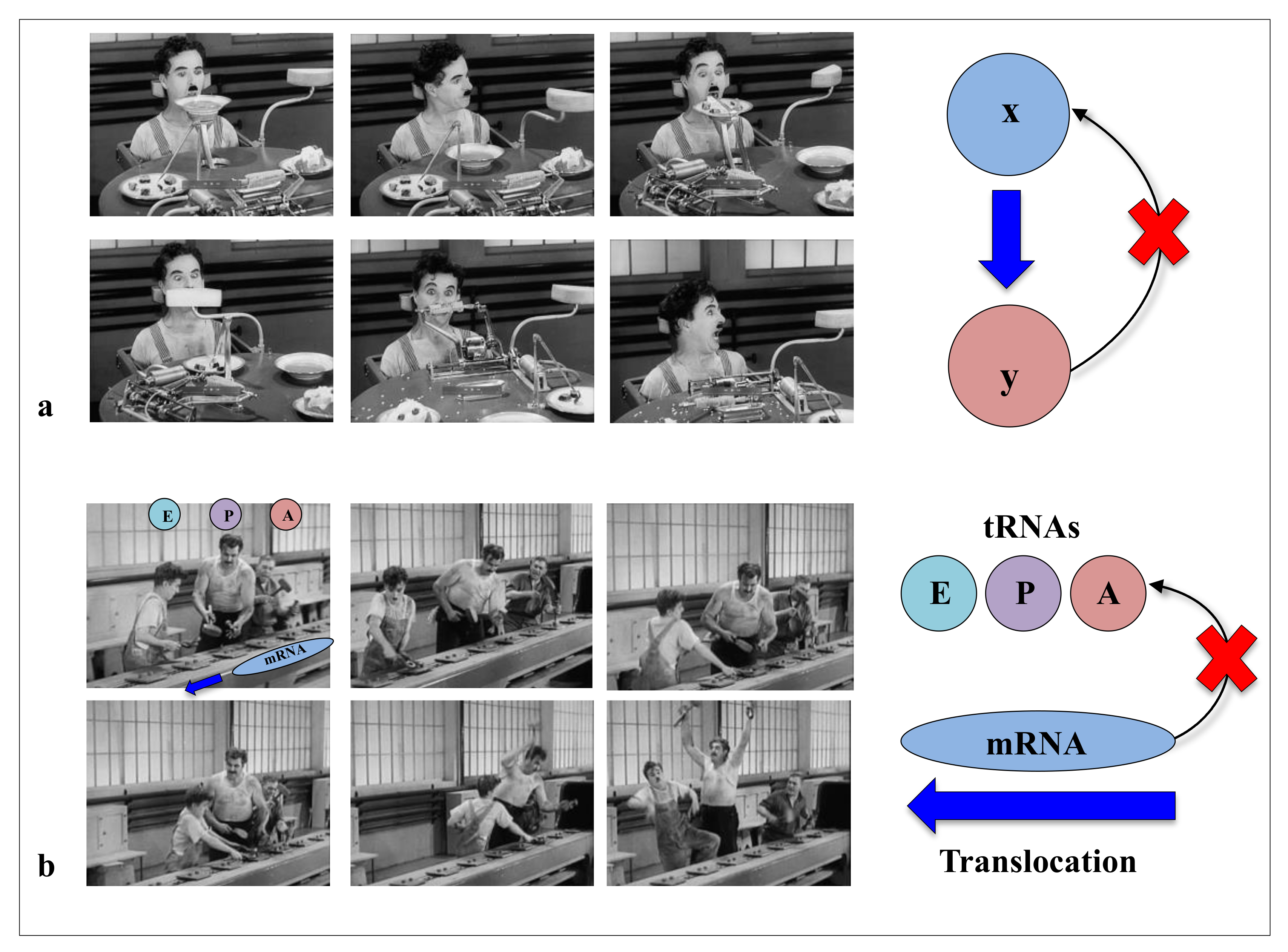
1.1. Feeding Without Feedback: Chaplin and the “Machine Metaphor” of Life
1.2. Information Transfer, Processing and Behaviours at the Molecular Scale
1.3. The Roots of the Molecular Brain Metaphor
1.4. Brains Beyond Connectomes
1.5. Where the “Molecular Brains” Could Take Us?
2. The Psychic Life of Microorganisms
2.1. Cajal and the Neuronal Turn of the Neurosciences
2.2. Binet, Jennings and Gelber: Toward Neuron-Free Neurosciences?
2.3. Finally, What Is Behaviour?
2.4. Categories of Molecular and Cellular Behaviours: From Sensorimotor Control to Decision Making
2.5. Cell Cognition and Consciousness
3. Cell Signalling and Sensory Motricity
3.1. The “Sensitive” Nature of Biological Molecules
3.2. Gated Channels
3.2.1. Voltage-Gated Channels
3.2.2. Temperature-Gated Channel
3.2.3. Mechanosensitive Ion Channels
3.2.4. Light-Sensitive Channels
3.2.5. Ligand-Gated Channels
3.3. Membrane Receptors
3.3.1. G Protein-Coupled Receptors (GPCR)
3.3.2. Kinases and Protein Phosphorylation
3.3.3. Photoreceptors
3.4. Other Molecular Sensors
3.4.1. Magnetoreception
3.4.2. Stress and Stressosome
3.4.3. Sensing with Nucleic Acids
3.5. The First Proto-Brains’ Ideas in Bacterial Chemotaxis
4. Ribosome Signalling and Primordial Molecular Brains
5. Evolution of Informational Systems across Scales
5.1. Comparison of Signalling Systems
5.2. Bayesian Brain Hypothesis
5.3. Beyond Allostery? Aromaticity in Signal Integration and Decision Making
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aristotle. Peri Psyches; Wentworth Press: London, UK, 2016. [Google Scholar]
- Levi, J. Tchouang-Tseu Maitre Du Tao; Pygmalion: Paris, France, 2006. [Google Scholar]
- Billeter, J.-F. Leçon Sur Thouang-Tseu; Allia: Paris, France, 2014. [Google Scholar]
- Timsit, M. Test de Rorschach et pathologie psychosomatique. Psychol. Clin. Et Proj. 1978, 31, 11–31. [Google Scholar] [CrossRef]
- Timsit-Berthier, M. Psychophysiologie cognitive et psychologie projective. Psychol. Clin. Et Proj. 1994, 38, 31–36. [Google Scholar] [CrossRef]
- Lagache, D. La rêverie imageante (Conduite adaptative au test de Rorschach). Psychol. Clin. Et Proj. 1957, 9, 3–11. [Google Scholar] [CrossRef]
- Guerra, P.A. The Monarch Butterfly as a Model for Understanding the Role of Environmental Sensory Cues in Long-Distance Migratory Phenomena. Front. Behav. Neurosci. 2020, 14, 600737. [Google Scholar] [CrossRef]
- Wan, G.; Hayden, A.N.; Iiams, S.E.; Merlin, C. Cryptochrome 1 Mediates Light-Dependent Inclination Magnetosensing in Monarch Butterflies. Nat. Commun. 2021, 12, 771. [Google Scholar] [CrossRef]
- Al-Khalili, J.; McFadden, J. Life on the Edge: The Coming of Age of Quantum Biology; Bantam Press: London, UK, 2014; ISBN 978-0-593-06932-5. [Google Scholar]
- Lorenz, E.N. Deterministic Nonperiodic Flow. J. Atmos. Sci. 1963, 20, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Trewavas, A.J.; Baluška, F. The Ubiquity of Consciousness. EMBO Rep. 2011, 12, 1221–1225. [Google Scholar] [CrossRef] [Green Version]
- Manley, D.B.; Taylor, C.S. Descartes’ Meditations—Trilingual Edition. 1996. Available online: https://corescholar.libraries.wright.edu/philosophy/8 (accessed on 21 October 2021).
- Bray, D. Protein Molecules as Computational Elements in Living Cells. Nature 1995, 376, 307–312. [Google Scholar] [CrossRef]
- Bray, D. Intrinsic Activity in Cells and the Brain. Mol. Biol. Cell 2014, 25, 737–738. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Lim, W. Cellular Perception and Misperception: Internal Models for Decision-Making Shaped by Evolutionary Experience. Bioessays 2016, 38, 845–849. [Google Scholar] [CrossRef] [Green Version]
- Okabe, K.; Yatani, A.; Brown, A.M. The Nature and Origin of Spontaneous Noise in G Protein-Gated Ion Channels. J. Gen. Physiol. 1991, 97, 1279–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, W.I.; Kobilka, B.K. The Molecular Basis of G Protein-Coupled Receptor Activation. Annu. Rev. Biochem. 2018, 87, 897–919. [Google Scholar] [CrossRef]
- Volinsky, N.; Kholodenko, B.N. Complexity of Receptor Tyrosine Kinase Signal Processing. Cold Spring Harb. Perspect. Biol. 2013, 5, a009043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutkeviciute, I.; Vilardaga, J.-P. Structural Insights into Emergent Signaling Modes of G Protein-Coupled Receptors. J. Biol. Chem. 2020, 295, 11626–11642. [Google Scholar] [CrossRef] [PubMed]
- Mast, F.D.; Ratushny, A.V.; Aitchison, J.D. Systems Cell Biology. J. Cell Biol. 2014, 206, 695–706. [Google Scholar] [CrossRef]
- Kitano, H. Computational Systems Biology. Nature 2002, 420, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.F. Pattern Formation and Complexity in Single Cells. Curr. Biol. 2020, 30, R544–R552. [Google Scholar] [CrossRef]
- Kirschner, M.; Gerhart, J.; Mitchison, T. Molecular “Vitalism”. Cell 2000, 100, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Fitch, W.T. Nano-Intentionality: A Defense of Intrinsic Intentionality. Biol. Philos. 2008, 23, 157–177. [Google Scholar] [CrossRef]
- Uversky, V.N. Dancing Protein Clouds: The Strange Biology and Chaotic Physics of Intrinsically Disordered Proteins*. J. Biol. Chem. 2016, 291, 6681–6688. [Google Scholar] [CrossRef] [Green Version]
- Marijuán, P.C.; Navarro, J.; del Moral, R. How the Living Is in the World: An Inquiry into the Informational Choreographies of Life. Prog. Biophys. Mol. Biol. 2015, 119, 469–480. [Google Scholar] [CrossRef]
- Ott, M. Cell Biology: Choreography of Protein Synthesis. Nature 2016, 533, 472–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemon, B.; Tjian, R. Orchestrated Response: A Symphony of Transcription Factors for Gene Control. Genes Dev. 2000, 14, 2551–2569. [Google Scholar] [CrossRef] [Green Version]
- Louppe, L. Écriture littéraire, écriture chorégraphique au XXe siècle: Une double révolution. Littérature 1998, 112, 88–99. [Google Scholar] [CrossRef]
- Louppe, L.; Dobbels, D.; Virilio, P.; Thom, R.; Laurenti, J.-N.; Preston-Dunlop, V. Danses Tracées. In Dessins et Notation des Chorégraphes; Dis Voir: New York, NY, USA, 2005. [Google Scholar]
- de Lahunta, S. Traces and Artefacts of Physical Intelligence. In The Performing Subject in the Space of Technology: Through the Virtual, Towards the Real; Causey, M., Meehan, E., O’Dwyer, N., Eds.; Palgrave Studies in Performance and Technology; Palgrave Macmillan UK: London, UK, 2015; pp. 220–231. ISBN 978-1-137-43816-4. [Google Scholar]
- Delahunta, R.S.; Barnard, P.; Nimmo-Smith, I.; Potts, J.; Ramponi, C. Densites of Agreement: Making Visible Some Intangible Properties of Dance. Danc. Theatre J. 2005, 21, 17–23. [Google Scholar]
- Timsit, Y. DNA-DNA Recognition: From Tight Contact to Fatal Attraction; IntechOpen: London, UK, 2012; ISBN 978-953-51-0079-9. [Google Scholar]
- Timsit, Y.; Westhof, E.; Fuchs, R.P.; Moras, D. Unusual Helical Packing in Crystals of DNA Bearing a Mutation Hot Spot. Nature 1989, 341, 459–462. [Google Scholar] [CrossRef]
- Várnai, P.; Timsit, Y. Differential Stability of DNA Crossovers in Solution Mediated by Divalent Cations. Nucleic Acids Res. 2010, 38, 4163–4172. [Google Scholar] [CrossRef] [Green Version]
- Makarova, T.M.; Bogdanov, A.A. The Ribosome as an Allosterically Regulated Molecular Machine. Biochem. Mosc. 2017, 82, 1557–1571. [Google Scholar] [CrossRef]
- Chen, J.; Choi, J.; O’Leary, S.E.; Prabhakar, A.; Petrov, A.; Grosely, R.; Puglisi, E.V.; Puglisi, J.D. The Molecular Choreography of Protein Synthesis: Translational Control, Regulation, and Pathways. Q. Rev. Biophys. 2016, 49, e11. [Google Scholar] [CrossRef]
- Nurse, P. Life, Logic and Information. Nature 2008, 454, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Bachelard, G. La Formation de L’Esprit Scientifique: Contribution À Une Psychanalyse de La Connaissance Objective; Vrin: Paris, France, 1947. [Google Scholar]
- Baluška, F.; Miller, W.B.; Reber, A.S. Biomolecular Basis of Cellular Consciousness via Subcellular Nanobrains. Int. J. Mol. Sci. 2021, 22, 2545. [Google Scholar] [CrossRef]
- Kholodenko, B.N. Cell-Signalling Dynamics in Time and Space. Nat. Rev. Mol. Cell Biol. 2006, 7, 165–176. [Google Scholar] [CrossRef]
- Azeloglu, E.U.; Iyengar, R. Signaling Networks: Information Flow, Computation, and Decision Making. Cold Spring Harb. Perspect. Biol. 2015, 7, a005934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.; Chauhan, P.; Saha, B.; Kubatzky, K.F. Conceptual Evolution of Cell Signaling. Int. J. Mol. Sci. 2019, 20, 3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timsit, Y.; Bennequin, D. Nervous-Like Circuits in the Ribosome Facts, Hypotheses and Perspectives. Int. J. Mol. Sci. 2019, 20, 2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirot, O.; Timsit, Y. Neuron-Like Networks Between Ribosomal Proteins Within the Ribosome. Sci. Rep. 2016, 6, 26485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timsit, Y.; Sergeant-Perthuis, G.; Bennequin, D. Evolution of Ribosomal Protein Network Architectures. Sci. Rep. 2021, 11, 625. [Google Scholar] [CrossRef] [PubMed]
- Barabási, A.-L.; Oltvai, Z.N. Network Biology: Understanding the Cell’s Functional Organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar] [CrossRef]
- Stock, J.; Levit, M. Signal Transduction: Hair Brains in Bacterial Chemotaxis. Curr. Biol. 2000, 10, R11–R14. [Google Scholar] [CrossRef] [Green Version]
- Stock, J.B.; Levit, M.N.; Wolanin, P.M. Information Processing in Bacterial Chemotaxis. Sci. Signal. 2002, 2002, pe25. [Google Scholar] [CrossRef]
- Webre, D.J.; Wolanin, P.M.; Stock, J.B. Bacterial Chemotaxis. Curr. Biol. 2003, 13, R47–R49. [Google Scholar] [CrossRef] [Green Version]
- Koshland, D.E. Bacterial Chemotaxis in Relation to Neurobiology. Annu. Rev. Neurosci. 1980, 3, 43–75. [Google Scholar] [CrossRef]
- Adler, J. Bacterial Chemotaxis and Molecular Neurobiology. Cold Spring Harb. Symp. Quant. Biol. 1983, 48, 803–804. [Google Scholar] [CrossRef] [PubMed]
- Baluška, F.; Mancuso, S.; Volkmann, D.; Barlow, P. The ‘Root-Brain’ Hypothesis of Charles and Francis Darwin. Plant. Signal. Behav. 2009, 4, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwin, C.; Darwin, S.F. The Power of Movement in Plants; John Murray: London, UK, 1880. [Google Scholar]
- Ben-Shem, A.; Garreau de Loubresse, N.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The Structure of the Eukaryotic Ribosome at 3.0 Å Resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef] [Green Version]
- Vion-Dury, J.; Mougin, G. Neuroscience sans Conscience n’est Que Ruine de l’âme. In Les Enjeux Cognitifs de l’artéfact Esthétique; L’Harmattan: Paris, France, 2018. [Google Scholar]
- Mazzarello, P. From Images to Physiology: A Strange Paradox at the Origin of Modern Neuroscience. Prog. Brain Res. 2018, 243, 233–256. [Google Scholar] [CrossRef]
- Fan, X.; Markram, H. A Brief History of Simulation Neuroscience. Front. Neuroinform 2019, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Budd, G.E. Early Animal Evolution and the Origins of Nervous Systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [Green Version]
- Cartmill, M. Convergent? Minds? Some Questions about Mental Evolution. Interface Focus 2017, 7, 20160125. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Sporns, O. Network Neuroscience. Nat. Neurosci. 2017, 20, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Solé, R.; Moses, M.; Forrest, S. Liquid Brains, Solid Brains. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190040. [Google Scholar] [CrossRef] [PubMed]
- Perlovsky, L.I. Physics of the Mind. Front. Syst. Neurosci. 2016, 10, 84. [Google Scholar] [CrossRef]
- Friston, K. The Free-Energy Principle: A Unified Brain Theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Hameroff, S.; Penrose, R. Consciousness in the Universe: A Review of the “Orch OR” Theory. Phys. Life Rev. 2014, 11, 39–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinov, M.; Sporns, O. Complex Network Measures of Brain Connectivity: Uses and Interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Sporns, O. Rich-Club Organization of the Human Connectome. J. Neurosci. 2011, 31, 15775–15786. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Pantelis, C.; Bullmore, E.T. Schizophrenia, Neuroimaging and Connectomics. NeuroImage 2012, 62, 2296–2314. [Google Scholar] [CrossRef]
- Fornito, A.; Bullmore, E.T. Connectomic Intermediate Phenotypes for Psychiatric Disorders. Front. Psychiatry 2012, 3, 32. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2011; ISBN 978-0-262-01469-4. [Google Scholar]
- Izquierdo, E.J.; Beer, R.D. Connecting a Connectome to Behavior: An Ensemble of Neuroanatomical Models of C. Elegans Klinotaxis. PLoS Comput Biol. 2013, 9, e1002890. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O. Contributions and Challenges for Network Models in Cognitive Neuroscience. Nat. Neurosci. 2014, 17, 652–660. [Google Scholar] [CrossRef]
- Droulez, J.; Colliaux, D.; Houillon, A.; Bessière, P. Toward Biochemical Probabilistic Computation. arXiv 2015, arXiv:1511.02623. [Google Scholar]
- Colliaux, D.; Bessière, P.; Droulez, J. Cell Signaling as a Probabilistic Computer. Int. J. Approx. Reason. 2017, 83, 385–399. [Google Scholar] [CrossRef]
- Piccolino, M. Luigi Galvani and Animal Electricity: Two Centuries after the Foundation of Electrophysiology. Trends Neurosci. 1997, 20, 443–448. [Google Scholar] [CrossRef]
- Piccolino, M. Animal Electricity and the Birth of Electrophysiology: The Legacy of Luigi Galvani. Brain Res. Bull. 1998, 46, 381–407. [Google Scholar] [CrossRef]
- Scanziani, M.; Häusser, M. Electrophysiology in the Age of Light. Nature 2009, 461, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Van Hemmen, J.L.; Sejnowski, T.J. (Eds.) 23 Problems in Systems Neuroscience; Oxford University Press: Oxford, UK; New York, NY, USA, 2006; ISBN 978-0-19-514822-0. [Google Scholar]
- Gerstner, W.; Kistler, W.M.; Naud, R.; Paninski, L. Neuronal Dynamics: From Single Neurons to Networks and Models of Cognition; Cambridge University Press: Cambridge, MA, USA, 2014; ISBN 978-1-107-06083-8. [Google Scholar]
- Hahn, G.; Ponce-Alvarez, A.; Deco, G.; Aertsen, A.; Kumar, A. Portraits of Communication in Neuronal Networks. Nat. Rev. Neurosci. 2019, 20, 117–127. [Google Scholar] [CrossRef]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 978-1-118-58577-1. [Google Scholar]
- Adrian, E.D.; Zotterman, Y. The Impulses Produced by Sensory Nerve-Endings. J. Physiol. 1926, 61, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Panzeri, S.; Brunel, N.; Logothetis, N.K.; Kayser, C. Sensory Neural Codes Using Multiplexed Temporal Scales. Trends Neurosci. 2010, 33, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Panzeri, S.; Harvey, C.D.; Piasini, E.; Latham, P.E.; Fellin, T. Cracking the Neural Code for Sensory Perception by Combining Statistics, Intervention, and Behavior. Neuron 2017, 93, 491–507. [Google Scholar] [CrossRef] [Green Version]
- Gerstner, W.; Kreiter, A.K.; Markram, H.; Herz, A.V.M. Neural Codes: Firing Rates and Beyond. Proc. Natl. Acad. Sci. USA 1997, 94, 12740–12741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, R.B.; Gossen, E.R.; Jones, K.E. Neuronal Variability: Noise or Part of the Signal? Nat. Rev. Neurosci. 2005, 6, 389–397. [Google Scholar] [CrossRef]
- Lorenzo, P.M.D.; Chen, J.-Y.; Victor, J.D. Quality Time: Representation of a Multidimensional Sensory Domain through Temporal Coding. J. Neurosci. 2009, 29, 9227–9238. [Google Scholar] [CrossRef] [PubMed]
- Putney, J.; Conn, R.; Sponberg, S. Precise Timing Is Ubiquitous, Consistent, and Coordinated across a Comprehensive, Spike-Resolved Flight Motor Program. Proc. Natl. Acad. Sci. USA 2019, 116, 26951–26960. [Google Scholar] [CrossRef] [Green Version]
- Bialek, W.; de Steveninck, R.R.; de Steveninck, R.R.; Rieke, F.; Warland, D. Spikes: Exploring the Neural Code; Computational Neuroscience Series; A Bradford Book: Cambridge, MA, USA, 1996; ISBN 978-0-262-18174-7. [Google Scholar]
- Brette, R. Philosophy of the Spike: Rate-Based vs. Spike-Based Theories of the Brain. Front. Syst. Neurosci. 2015, 9, 151. [Google Scholar] [CrossRef]
- Brette, R. Is Coding a Relevant Metaphor for the Brain? Behav Brain Sci. 2018, 42, e215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pregowska, A.; Kaplan, E.; Szczepanski, J. How Far Can Neural Correlations Reduce Uncertainty? Comparison of Information Transmission Rates for Markov and Bernoulli Processes. Int. J. Neur. Syst. 2019, 29, 1950003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crumiller, M.; Knight, B.; Kaplan, E. The Measurement of Information Transmitted by a Neural Population: Promises and Challenges. Entropy 2013, 15, 3507–3527. [Google Scholar] [CrossRef] [Green Version]
- Grant, S.G.N. Synapse Molecular Complexity and the Plasticity Behaviour Problem. Brain Neurosci. Adv. 2018, 2, 2398212818810685. [Google Scholar] [CrossRef] [PubMed]
- Baas, P. Antoni Van Leeuwenhoek and His Observation on the Structure of the Woody Cell Wall. IAWA J. 1982, 3, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Sotelo, C. The History of the Synapse. Anat. Rec. 2020, 303, 1252–1279. [Google Scholar] [CrossRef]
- Llinás, R.R. The Contribution of Santiago Ramón y Cajal to Functional Neuroscience. Nat. Rev. Neurosci. 2003, 4, 77–80. [Google Scholar] [CrossRef]
- Sotelo, C. Viewing the Brain through the Master Hand of Ramón y Cajal. Nat. Rev. Neurosci. 2003, 4, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Delgado-García, J.M. Cajal and the Conceptual Weakness of Neural Sciences. Front. Neuroanat 2015, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Jermakowicz, W.J.; Casagrande, V.A. Neural Networks a Century after Cajal. Brain Res. Rev. 2007, 55, 264–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, F.R.M.; Nogueira, M.I.; Defelipe, J. The Influence of James and Darwin on Cajal and His Research into the Neuron Theory and Evolution of the Nervous System. Front. Neuroanat 2014, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- DeFelipe, J.; Garrido, E.; Markram, H. The Death of Cajal and the End of Scientific Romanticism and Individualism. Trends Neurosci. 2014, 37, 525–527. [Google Scholar] [CrossRef] [Green Version]
- Binet, A. The Psychic Life of Micro-Organisms. A Study in Experimental Psychology; The Open Court Publishing Company, London Agents Kegan Paul Trench, Trubner & Co LTD: London, UK, 1889. [Google Scholar]
- Galperin, M.Y. A Census of Membrane-Bound and Intracellular Signal Transduction Proteins in Bacteria: Bacterial IQ, Extroverts and Introverts. BMC Microbiol. 2005, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Jennings, H.S. Behavior of the Lower Organisms; Columbia University Press: New York, NY, USA, 1906. [Google Scholar]
- Loeb, J. The Mechanistic Conception of Life; Harvard University Press: Cambridge, MA, USA, 2013; ISBN 978-0-674-86431-3. [Google Scholar]
- Darwin, C.; Prodger, P. The Expression of the Emotions in Man and Animals; Oxford University Press: Oxford, UK, 1998; ISBN 978-0-19-515806-9. [Google Scholar]
- Gould, S.J. The Mismeasure of Man. Ethics 1983, 94, 153–155. [Google Scholar]
- Gelber, B. Food or Training Paramecium. Science 1957, 126, 1340–1341. [Google Scholar] [CrossRef] [PubMed]
- Gershman, S.J.; Balbi, P.E.; Gallistel, C.R.; Gunawardena, J. Reconsidering the Evidence for Learning in Single Cells. Elife 2021, 10, e61907. [Google Scholar] [CrossRef]
- Berman, G.J. How to Build a Behavior. Neuron 2018, 100, 1275–1277. [Google Scholar] [CrossRef] [Green Version]
- Berman, G.J. Measuring Behavior across Scales. BMC Biol. 2018, 16, 23. [Google Scholar] [CrossRef]
- Grant, S.G.N. The Molecular Evolution of the Vertebrate Behavioural Repertoire. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150051. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y. What Bacteria Want. Environ. Microbiol. 2018, 20, 4221–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jékely, G.; Godfrey-Smith, P.; Keijzer, F. Reafference and the Origin of the Self in Early Nervous System Evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20190764. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. The Computational Boundary of a “Self”: Developmental Bioelectricity Drives Multicellularity and Scale-Free Cognition. Front. Psychol. 2019, 10, 2688. [Google Scholar] [CrossRef] [Green Version]
- Fields, C.; Glazebrook, J.F.; Levin, M. Minimal Physicalism as a Scale-Free Substrate for Cognition and Consciousness. Neurosci. Conscious 2021, 2021, niab013. [Google Scholar] [CrossRef]
- Prossliner, T.; Skovbo Winther, K.; Sørensen, M.A.; Gerdes, K. Ribosome Hibernation. Annu. Rev. Genet. 2018, 52, 321–348. [Google Scholar] [CrossRef]
- Vind, A.C.; Genzor, A.V.; Bekker-Jensen, S. Ribosomal Stress-Surveillance: Three Pathways Is a Magic Number. Nucleic Acids Res. 2020, 48, 10648–10661. [Google Scholar] [CrossRef] [PubMed]
- Komar, A.A. A Pause for Thought along the Co-Translational Folding Pathway. Trends Biochem. Sci. 2009, 34, 16–24. [Google Scholar] [CrossRef]
- Wodak, S.J.; Paci, E.; Dokholyan, N.V.; Berezovsky, I.N.; Horovitz, A.; Li, J.; Hilser, V.J.; Bahar, I.; Karanicolas, J.; Stock, G.; et al. Allostery in Its Many Disguises: From Theory to Applications. Structure 2019, 27, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Erez, Z.; Steinberger-Levy, I.; Shamir, M.; Doron, S.; Stokar-Avihail, A.; Peleg, Y.; Melamed, S.; Leavitt, A.; Savidor, A.; Albeck, S.; et al. Communication between Viruses Guides Lysis-Lysogeny Decisions. Nature 2017, 541, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Calap, P.; Segredo-Otero, E.; Durán-Moreno, M.; Sanjuán, R. Social Evolution of Innate Immunity Evasion in a Virus. Nat. Microbiol. 2019, 4, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Krieger, I.V.; Kuznetsov, V.; Chang, J.-Y.; Zhang, J.; Moussa, S.H.; Young, R.F.; Sacchettini, J.C. The Structural Basis of T4 Phage Lysis Control: DNA as the Signal for Lysis Inhibition. J. Mol. Biol. 2020, 432, 4623–4636. [Google Scholar] [CrossRef]
- Dunigan, D.D.; Al-Sammak, M.; Al-Ameeli, Z.; Agarkova, I.V.; DeLong, J.P.; Van Etten, J.L. Chloroviruses Lure Hosts through Long-Distance Chemical Signaling. J. Virol. 2019, 93, e01688-18. [Google Scholar] [CrossRef] [Green Version]
- Stokar-Avihail, A.; Tal, N.; Erez, Z.; Lopatina, A.; Sorek, R. Widespread Utilization of Peptide Communication in Phages Infecting Soil and Pathogenic Bacteria. Cell Host Microbe 2019, 25, 746–755.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego Del Sol, F.; Penadés, J.R.; Marina, A. Deciphering the Molecular Mechanism Underpinning Phage Arbitrium Communication Systems. Mol. Cell 2019, 74, 59–72.e3. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial Quorum Sensing in Complex and Dynamically Changing Environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Shiner, E.K.; Rumbaugh, K.P.; Williams, S.C. Inter-Kingdom Signaling: Deciphering the Language of Acyl Homoserine Lactones. FEMS Microbiol. Rev. 2005, 29, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Reading, N.C.; Sperandio, V. Quorum Sensing: The Many Languages of Bacteria. FEMS Microbiol. Lett. 2006, 254, 1–11. [Google Scholar] [CrossRef]
- Joint, I.; Tait, K.; Wheeler, G. Cross-Kingdom Signalling: Exploitation of Bacterial Quorum Sensing Molecules by the Green Seaweed Ulva. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1223–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reguera, G. When Microbial Conversations Get Physical. Trends Microbiol. 2011, 19, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Cifra, M.; Fields, J.Z.; Farhadi, A. Electromagnetic Cellular Interactions. Prog. Biophys. Mol. Biol. 2011, 105, 223–246. [Google Scholar] [CrossRef] [PubMed]
- Kučera, O.; Cifra, M. Cell-to-Cell Signaling through Light: Just a Ghost of Chance? Cell Commun. Signal. 2013, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholkmann, F.; Fels, D.; Cifra, M. Non-Chemical and Non-Contact Cell-to-Cell Communication: A Short Review. Am. J. Transl. Res. 2013, 5, 586–593. [Google Scholar] [PubMed]
- Humphries, J.; Xiong, L.; Liu, J.; Prindle, A.; Yuan, F.; Arjes, H.A.; Tsimring, L.; Süel, G.M. Species-Independent Attraction to Biofilms through Electrical Signaling. Cell 2017, 168, 200–209.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, J.W.; Zhai, X.; Kikuchi, K.; Redford, S.E.; Prindle, A.; Liu, J.; Greenfield, S.; Walczak, A.M.; Garcia-Ojalvo, J.; Mugler, A.; et al. Signal Percolation within a Bacterial Community. Cell Syst. 2018, 7, 137–145.e3. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.J.; Lee, S.K.; Kim, T.; Ghim, C.-M. Microbial Linguistics: Perspectives and Applications of Microbial Cell-to-Cell Communication. BMB Rep. 2011, 44, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Jacob, E.; Becker, I.; Shapira, Y.; Levine, H. Bacterial Linguistic Communication and Social Intelligence. Trends Microbiol. 2004, 12, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Pal, S. Information Transmission in Microbial and Fungal Communication: From Classical to Quantum. J. Cell Commun. Signal. 2018, 12, 491–502. [Google Scholar] [CrossRef]
- Fels, D. Endogenous Physical Regulation of Population Density in the Freshwater Protozoan Paramecium Caudatum. Sci. Rep. 2017, 7, 13800. [Google Scholar] [CrossRef] [Green Version]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell 2006, 125, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Combarnous, Y.; Nguyen, T.M.D. Cell Communications among Microorganisms, Plants, and Animals: Origin, Evolution, and Interplays. Int. J. Mol. Sci. 2020, 21, 8052. [Google Scholar] [CrossRef]
- Brunet, T.; King, N. The Origin of Animal Multicellularity and Cell Differentiation. Dev. Cell 2017, 43, 124–140. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, M.; Langston, L.; Stillman, B. Principles and Concepts of DNA Replication in Bacteria, Archaea, and Eukarya. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef]
- Srivastava, V.; Iglesias, P.A.; Robinson, D.N. Cytokinesis: Robust Cell Shape Regulation. Semin. Cell Dev. Biol. 2016, 53, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.M.; Fontaine-Bodin, L.; Bischofs, I.; Price, G.; Keasling, J.; Arkin, A.P. Memory in Microbes: Quantifying History-Dependent Behavior in a Bacterium. PLoS ONE 2008, 3, e1700. [Google Scholar] [CrossRef] [Green Version]
- Kandel, E.; Abel, T. Neuropeptides, Adenylyl Cyclase, and Memory Storage. Science 1995, 268, 825–826. [Google Scholar] [CrossRef] [Green Version]
- Dussutour, A. Learning in Single Cell Organisms. Biochem. Biophys. Res. Commun. 2021. [Google Scholar] [CrossRef]
- Marshall, W.F. Cellular Cognition: Sequential Logic in a Giant Protist. Curr. Biol. 2019, 29, R1303–R1305. [Google Scholar] [CrossRef]
- Dexter, J.P.; Prabakaran, S.; Gunawardena, J. A Complex Hierarchy of Avoidance Behaviors in a Single-Cell Eukaryote. Curr. Biol. 2019, 29, 4323–4329.e2. [Google Scholar] [CrossRef]
- Trinh, M.K.; Wayland, M.T.; Prabakaran, S. Behavioural Analysis of Single-Cell Aneural Ciliate, Stentor Roeseli, Using Machine Learning Approaches. J. R. Soc. Interface 2019, 16, 20190410. [Google Scholar] [CrossRef]
- Beekman, M.; Latty, T. Brainless but Multi-Headed: Decision Making by the Acellular Slime Mould Physarum Polycephalum. J. Mol. Biol. 2015, 427, 3734–3743. [Google Scholar] [CrossRef]
- De la Fuente, I.M.; Bringas, C.; Malaina, I.; Fedetz, M.; Carrasco-Pujante, J.; Morales, M.; Knafo, S.; Martínez, L.; Pérez-Samartín, A.; López, J.I.; et al. Evidence of Conditioned Behavior in Amoebae. Nat. Commun. 2019, 10, 3690. [Google Scholar] [CrossRef] [Green Version]
- Hennessey, T.M.; Rucker, W.B.; McDiarmid, C.G. Classical Conditioning in Paramecia. Anim. Learn. Behav. 1979, 7, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Trewavas, A.J. Plants Are Intelligent Too. EMBO Rep. 2012, 13, 772–773, author reply 773. [Google Scholar] [CrossRef] [Green Version]
- Trewavas, A. Plant Intelligence. Naturwissenschaften 2005, 92, 401–413. [Google Scholar] [CrossRef]
- Trewavas, A. Response to Alpi et al.: Plant Neurobiology--All Metaphors Have Value. Trends Plant. Sci. 2007, 12, 231–233. [Google Scholar] [CrossRef]
- Baluška, F.; Miller, W.B. Senomic View of the Cell: Senome versus Genome. Commun. Integr. Biol. 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lyon, P. The Cognitive Cell: Bacterial Behavior Reconsidered. Front. Microbiol. 2015, 6, 264. [Google Scholar] [CrossRef]
- Marijuán, P.C.; Navarro, J.; del Moral, R. On Prokaryotic Intelligence: Strategies for Sensing the Environment. Biosystems 2010, 99, 94–103. [Google Scholar] [CrossRef]
- Ben-Jacob, E.; Levine, H. Self-Engineering Capabilities of Bacteria. J. R. Soc. Interface 2006, 3, 197–214. [Google Scholar] [CrossRef] [Green Version]
- Baluška, F.; Levin, M. On Having No Head: Cognition throughout Biological Systems. Front. Psychol. 2016, 7, 902. [Google Scholar] [CrossRef] [Green Version]
- Keijzer, F. Moving and Sensing without Input and Output: Early Nervous Systems and the Origins of the Animal Sensorimotor Organization. Biol. Philos. 2015, 30, 311–331. [Google Scholar] [CrossRef] [Green Version]
- Pattee, H.H. Cell Phenomenology: The First Phenomenon. Prog. Biophys. Mol. Biol. 2015, 119, 461–468. [Google Scholar] [CrossRef]
- Richardson, K. Heritability Lost; Intelligence Found. Intelligence Is Integral to the Adaptation and Survival of All Organisms Faced with Changing Environments. EMBO Rep. 2012, 13, 591–595. [Google Scholar] [CrossRef] [Green Version]
- Baluska, F.; Mancuso, S. Deep Evolutionary Origins of Neurobiology: Turning the Essence of “neural” Upside-Down. Commun. Integr. Biol. 2009, 2, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Baluška, F. Conditions for Minimal Intelligence across Eukaryota: A Cognitive Science Perspective. Front. Psychol. 2015, 6, 1329. [Google Scholar] [CrossRef] [Green Version]
- van Duijn, M. Phylogenetic Origins of Biological Cognition: Convergent Patterns in the Early Evolution of Learning. Interface Focus 2017, 7, 20160158. [Google Scholar] [CrossRef] [Green Version]
- Jékely, G.; Keijzer, F.; Godfrey-Smith, P. An Option Space for Early Neural Evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Blaby-Haas, C.E.; Merchant, S.S.; Mains, R.E.; King, S.M.; Eipper, B.A. Early Eukaryotic Origins for Cilia-Associated Bioactive Peptide-Amidating Activity. J. Cell Sci. 2016, 129, 943–956. [Google Scholar] [CrossRef] [Green Version]
- Senatore, A.; Reese, T.S.; Smith, C.L. Neuropeptidergic Integration of Behavior in Trichoplax Adhaerens, an Animal without Synapses. J. Exp. Biol. 2017, 220, 3381–3390. [Google Scholar] [CrossRef] [Green Version]
- Jékely, G. Origin and Early Evolution of Neural Circuits for the Control of Ciliary Locomotion. Proc. Biol. Sci. 2011, 278, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Dukas, R. Cognitive Innovations and the Evolutionary Biology of Expertise. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160427. [Google Scholar] [CrossRef]
- Hein, A.M.; Carrara, F.; Brumley, D.R.; Stocker, R.; Levin, S.A. Natural Search Algorithms as a Bridge between Organisms, Evolution, and Ecology. Proc. Natl. Acad. Sci. USA 2016, 113, 9413–9420. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.-H.; Lu, B.; Evans, R.; Gutkind, J.S. Signals and Receptors. Cold Spring Harb. Perspect. Biol. 2016, 8, a005900. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.D.; Pawson, T. Cell Signaling in Space and Time: Where Proteins Come Together and When They’re Apart. Science 2009, 326, 1220–1224. [Google Scholar] [CrossRef] [Green Version]
- Thorner, J.; Hunter, T.; Cantley, L.C.; Sever, R. Signal Transduction: From the Atomic Age to the Post-Genomic Era. Cold Spring Harb. Perspect. Biol. 2014, 6, a022913. [Google Scholar] [CrossRef] [Green Version]
- Cashin, P.; Goldsack, L.; Hall, D.; O’Toole, R. Contrasting Signal Transduction Mechanisms in Bacterial and Eukaryotic Gene Transcription. FEMS Microbiol. Lett. 2006, 261, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Aravind, L.; Anantharaman, V.; Iyer, L.M. Evolutionary Connections between Bacterial and Eukaryotic Signaling Systems: A Genomic Perspective. Curr. Opin. Microbiol. 2003, 6, 490–497. [Google Scholar] [CrossRef]
- Ulrich, L.E.; Koonin, E.V.; Zhulin, I.B. One-Component Systems Dominate Signal Transduction in Prokaryotes. Trends Microbiol. 2005, 13, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Alm, E.; Huang, K.; Arkin, A. The Evolution of Two-Component Systems in Bacteria Reveals Different Strategies for Niche Adaptation. PLoS Comput. Biol. 2006, 2, e143. [Google Scholar] [CrossRef]
- Capra, E.J.; Laub, M.T. Evolution of Two-Component Signal Transduction Systems. Annu. Rev. Microbiol. 2012, 66, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Phyletic Distribution and Lineage-Specific Domain Architectures of Archaeal Two-Component Signal Transduction Systems. J. Bacteriol. 2018, 200, e00681-17. [Google Scholar] [CrossRef] [Green Version]
- Jacob-Dubuisson, F.; Mechaly, A.; Betton, J.-M.; Antoine, R. Structural Insights into the Signalling Mechanisms of Two-Component Systems. Nat. Rev. Microbiol. 2018, 16, 585–593. [Google Scholar] [CrossRef]
- West, A.H.; Stock, A.M. Histidine Kinases and Response Regulator Proteins in Two-Component Signaling Systems. Trends Biochem. Sci. 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Bourret, R.B.; Silversmith, R.E. Two-Component Signal Transduction. Curr. Opin. Microbiol. 2010, 13, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular Mechanisms of Two-Component Signal Transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Pawson, T. Modular Evolution of Phosphorylation-Based Signalling Systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2540–2555. [Google Scholar] [CrossRef]
- Babonis, L.S.; Martindale, M.Q. Phylogenetic Evidence for the Modular Evolution of Metazoan Signalling Pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [Green Version]
- Mayer, B.J. Clues to the Evolution of Complex Signaling Machinery. Proc. Natl. Acad. Sci. USA 2008, 105, 9453–9454. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Yaffe, M.B. Protein Regulation in Signal Transduction. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Valiunas, A. Richard Feynman and the Pleasure Principle. New Atlantis 2018, 50, 61–84. [Google Scholar]
- Wruck, F.; Katranidis, A.; Nierhaus, K.H.; Büldt, G.; Hegner, M. Translation and Folding of Single Proteins in Real Time. Proc. Natl. Acad. Sci. USA 2017, 114, E4399–E4407. [Google Scholar] [CrossRef] [Green Version]
- Rodnina, M.V.; Wintermeyer, W. Protein Elongation, Co-Translational Folding and Targeting. J. Mol. Biol. 2016, 428, 2165–2185. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Røgen, P.; Paci, E.; Vendruscolo, M.; Dobson, C.M. Protein Folding and the Organization of the Protein Topology Universe. Trends Biochem. Sci. 2005, 30, 13–19. [Google Scholar] [CrossRef]
- Fersht, A.R.; Daggett, V. Protein Folding and Unfolding at Atomic Resolution. Cell 2002, 108, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Rothman, J.E.; Schekman, R. Molecular Mechanism of Protein Folding in the Cell. Cell 2011, 146, 851–854. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.L.; Rose, G.D. Molten Globules, Entropy-Driven Conformational Change and Protein Folding. Curr. Opin. Struct. Biol. 2013, 23, 4–10. [Google Scholar] [CrossRef]
- Henzler-Wildman, K.; Kern, D. Dynamic Personalities of Proteins. Nature 2007, 450, 964–972. [Google Scholar] [CrossRef]
- Karplus, M.; McCammon, J.A. Molecular Dynamics Simulations of Biomolecules. Nat. Struct. Biol. 2002, 9, 646–652. [Google Scholar] [CrossRef]
- van der Kamp, M.W.; Schaeffer, R.D.; Jonsson, A.L.; Scouras, A.D.; Simms, A.M.; Toofanny, R.D.; Benson, N.C.; Anderson, P.C.; Merkley, E.D.; Rysavy, S.; et al. Dynameomics: A Comprehensive Database of Protein Dynamics. Structure 2010, 18, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, M.H.; Corces, V.G. A Tethered-Inchworm Model of SMC DNA Translocation. Nat. Struct. Mol. Biol. 2018, 25, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Ulianov, S.V.; Zakharova, V.V.; Galitsyna, A.A.; Kos, P.I.; Polovnikov, K.E.; Flyamer, I.M.; Mikhaleva, E.A.; Khrameeva, E.E.; Germini, D.; Logacheva, M.D.; et al. Order and Stochasticity in the Folding of Individual Drosophila Genomes. Nat. Commun. 2021, 12, 41. [Google Scholar] [CrossRef]
- Shaban, H.A.; Barth, R.; Bystricky, K. Navigating the Crowd: Visualizing Coordination between Genome Dynamics, Structure, and Transcription. Genome Biol. 2020, 21, 278. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Koenderink, G.H.; MacKintosh, F.C.; Weitz, D.A. Cytoplasmic Diffusion: Molecular Motors Mix It Up. J. Cell Biol. 2008, 183, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Amos, L.A.; Löwe, J. Overview of the Diverse Roles of Bacterial and Archaeal Cytoskeletons. Subcell Biochem. 2017, 84, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Margulis, L.; Chapman, M.; Guerrero, R.; Hall, J. The Last Eukaryotic Common Ancestor (LECA): Acquisition of Cytoskeletal Motility from Aerotolerant Spirochetes in the Proterozoic Eon. Proc. Natl. Acad. Sci. USA 2006, 103, 13080–13085. [Google Scholar] [CrossRef] [Green Version]
- Löwe, J.; Amos, L.A. Evolution of Cytomotive Filaments: The Cytoskeleton from Prokaryotes to Eukaryotes. Int. J. Biochem. Cell Biol. 2009, 41, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Cooper, J.A. Actin, a Central Player in Cell Shape and Movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [Green Version]
- Akıl, C.; Tran, L.T.; Orhant-Prioux, M.; Baskaran, Y.; Manser, E.; Blanchoin, L.; Robinson, R.C. Insights into the Evolution of Regulated Actin Dynamics via Characterization of Primitive Gelsolin/Cofilin Proteins from Asgard Archaea. Proc. Natl. Acad. Sci. USA 2020, 117, 19904–19913. [Google Scholar] [CrossRef]
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; Plastino, J. Actin Dynamics, Architecture, and Mechanics in Cell Motility. Physiol. Rev. 2014, 94, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Carlier, M.-F.; Le Clainche, C.; Wiesner, S.; Pantaloni, D. Actin-Based Motility: From Molecules to Movement. Bioessays 2003, 25, 336–345. [Google Scholar] [CrossRef]
- Carlier, M.-F.; Pantaloni, D. Control of Actin Assembly Dynamics in Cell Motility. J. Biol. Chem. 2007, 282, 23005–23009. [Google Scholar] [CrossRef] [Green Version]
- Trépout, S.; Wehenkel, A.M. Bacterial Tubulins: A Eukaryotic-Like Microtubule Cytoskeleton. Trends Microbiol. 2017, 25, 782–784. [Google Scholar] [CrossRef] [PubMed]
- Yutin, N.; Koonin, E.V. Archaeal Origin of Tubulin. Biol. Direct. 2012, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, E.T.; Voter, W.A.; Erickson, H.P. GTP Hydrolysis during Microtubule Assembly. Biochemistry 1987, 26, 4148–4156. [Google Scholar] [CrossRef]
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef] [PubMed]
- Beeby, M.; Ferreira, J.L.; Tripp, P.; Albers, S.-V.; Mitchell, D.R. Propulsive Nanomachines: The Convergent Evolution of Archaella, Flagella and Cilia. FEMS Microbiol. Rev. 2020, 44, 253–304. [Google Scholar] [CrossRef]
- Miyata, M.; Robinson, R.C.; Uyeda, T.Q.P.; Fukumori, Y.; Fukushima, S.-I.; Haruta, S.; Homma, M.; Inaba, K.; Ito, M.; Kaito, C.; et al. Tree of Motility—A Proposed History of Motility Systems in the Tree of Life. Genes Cells 2020, 25, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Armitage, J.P.; Berry, R.M. Assembly and Dynamics of the Bacterial Flagellum. Annu. Rev. Microbiol. 2020, 74, 181–200. [Google Scholar] [CrossRef]
- Ishikawa, T.; Ueno, H.; Omori, T.; Kikuchi, K. Cilia and Centrosomes: Ultrastructural and Mechanical Perspectives. Semin. Cell Dev. Biol. 2021, 110, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.R. The Evolution of Eukaryotic Cilia and Flagella as Motile and Sensory Organelles. Adv. Exp. Med. Biol. 2007, 607, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Wan, K.Y.; Jékely, G. On the Unity and Diversity of Cilia. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadadhar, S.; Bodakuntla, S.; Natarajan, K.; Janke, C. The Tubulin Code at a Glance. J. Cell Sci. 2017, 130, 1347–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadadhar, S.; Alvarez Viar, G.; Hansen, J.N.; Gong, A.; Kostarev, A.; Ialy-Radio, C.; Leboucher, S.; Whitfield, M.; Ziyyat, A.; Touré, A.; et al. Tubulin Glycylation Controls Axonemal Dynein Activity, Flagellar Beat, and Male Fertility. Science 2021, 371. [Google Scholar] [CrossRef]
- Goldschen-Ohm, M.P.; Chanda, B. SnapShot: Channel Gating Mechanisms. Cell 2017, 170, 594.e1. [Google Scholar] [CrossRef]
- Kim, D.M.; Nimigean, C.M. Voltage-Gated Potassium Channels: A Structural Examination of Selectivity and Gating. Cold Spring Harb. Perspect. Biol. 2016, 8, a029231. [Google Scholar] [CrossRef] [Green Version]
- Bezanilla, F. Voltage-Gated Ion Channels. IEEE Trans. NanoBiosci. 2005, 4, 34–48. [Google Scholar] [CrossRef]
- Catterall, W.A.; Lenaeus, M.J.; Gamal El-Din, T.M. Structure and Pharmacology of Voltage-Gated Sodium and Calcium Channels. Annu. Rev. Pharm. Toxicol. 2020, 60, 133–154. [Google Scholar] [CrossRef] [Green Version]
- Payandeh, J.; Scheuer, T.; Zheng, N.; Catterall, W.A. The Crystal Structure of a Voltage-Gated Sodium Channel. Nature 2011, 475, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Lee, A.; Chen, J.; Ruta, V.; Cadene, M.; Chait, B.T.; MacKinnon, R. X-Ray Structure of a Voltage-Dependent K+ Channel. Nature 2003, 423, 33–41. [Google Scholar] [CrossRef]
- Okamura, Y.; Okochi, Y. Molecular Mechanisms of Coupling to Voltage Sensors in Voltage-Evoked Cellular Signals. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 111–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, X.; Lee, A.; Limapichat, W.; Dougherty, D.A.; MacKinnon, R. A Gating Charge Transfer Center in Voltage Sensors. Science 2010, 328, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Long, S.B.; Tao, X.; Campbell, E.B.; MacKinnon, R. Atomic Structure of a Voltage-Dependent K+ Channel in a Lipid Membrane-like Environment. Nature 2007, 450, 376–382. [Google Scholar] [CrossRef]
- Long, S.B.; Campbell, E.B.; Mackinnon, R. Voltage Sensor of Kv1.2: Structural Basis of Electromechanical Coupling. Science 2005, 309, 903–908. [Google Scholar] [CrossRef] [Green Version]
- Heer, F.T.; Posson, D.J.; Wojtas-Niziurski, W.; Nimigean, C.M.; Bernèche, S. Mechanism of Activation at the Selectivity Filter of the KcsA K+ Channel. Elife 2017, 6, e25844. [Google Scholar] [CrossRef] [Green Version]
- Tao, X.; MacKinnon, R. Cryo-EM Structure of the KvAP Channel Reveals a Non-Domain-Swapped Voltage Sensor Topology. Elife 2019, 8, e52164. [Google Scholar] [CrossRef] [PubMed]
- Bignucolo, O.; Bernèche, S. The Voltage-Dependent Deactivation of the KvAP Channel Involves the Breakage of Its S4 Helix. Front. Mol. Biosci. 2020, 7, 162. [Google Scholar] [CrossRef]
- Murata, Y.; Iwasaki, H.; Sasaki, M.; Inaba, K.; Okamura, Y. Phosphoinositide Phosphatase Activity Coupled to an Intrinsic Voltage Sensor. Nature 2005, 435, 1239–1243. [Google Scholar] [CrossRef]
- Kohout, S.C.; Ulbrich, M.H.; Bell, S.C.; Isacoff, E.Y. Subunit Organization and Functional Transitions in Ci-VSP. Nat. Struct. Mol. Biol. 2008, 15, 106–108. [Google Scholar] [CrossRef]
- Okamura, Y. Biodiversity of Voltage Sensor Domain Proteins. Pflug. Arch. 2007, 454, 361–371. [Google Scholar] [CrossRef]
- Villalba-Galea, C.A. Voltage-Controlled Enzymes: The New JanusBifrons. Front. Pharm. 2012, 3, 161. [Google Scholar] [CrossRef] [Green Version]
- Knyazev, D.G.; Kuttner, R.; Bondar, A.-N.; Zimmerman, M.; Siligan, C.; Pohl, P. Voltage Sensing in Bacterial Protein Translocation. Biomolecules 2020, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faouri, R.A.; Krueger, E.; Govind Kumar, V.; Fologea, D.; Straub, D.; Alismail, H.; Alfaori, Q.; Kight, A.; Ray, J.; Henry, R.; et al. An Effective Electric Dipole Model for Voltage-Induced Gating Mechanism of Lysenin. Sci. Rep. 2019, 9, 11440. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Nilius, B.; Voets, T. Peripheral Thermosensation in Mammals. Nat. Rev. Neurosci. 2014, 15, 573–589. [Google Scholar] [CrossRef]
- Schneider, E.R.; Anderson, E.O.; Gracheva, E.O.; Bagriantsev, S.N. Temperature Sensitivity of Two-Pore (K2P) Potassium Channels. Curr. Top. Membr. 2014, 74, 113–133. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Franulic, I.; Poblete, H.; Miño-Galaz, G.; González, C.; Latorre, R. Allosterism and Structure in Thermally Activated Transient Receptor Potential Channels. Annu. Rev. Biophys. 2016, 45, 371–398. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, R.B.; Orellana, L.; Esteban-Martín, S.; Orozco, M.; Salvatella, X. Correlated Motions Are a Fundamental Property of β-Sheets. Nat. Commun. 2014, 5, 4070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 Structures in Distinct Conformations Reveal Activation Mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.; Qin, F.; Julius, D. TRPV1 Channels Are Intrinsically Heat Sensitive and Negatively Regulated by Phosphoinositide Lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Leitner, D.M. Energy Flow in Proteins. Annu. Rev. Phys. Chem. 2008, 59, 233–259. [Google Scholar] [CrossRef] [PubMed]
- Miño-Galaz, G.A. Allosteric Communication Pathways and Thermal Rectification in PDZ-2 Protein: A Computational Study. J. Phys. Chem. B 2015, 119, 6179–6189. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, C.; Rohaim, A.; Shaya, D.; Findeisen, F.; Stein, R.A.; Nurva, S.R.; Mishra, S.; Mchaourab, H.S.; Minor, D.L. Unfolding of a Temperature-Sensitive Domain Controls Voltage-Gated Channel Activation. Cell 2016, 164, 922–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigoni, C.; Minor, D.L. Global versus Local Mechanisms of Temperature Sensing in Ion Channels. Pflug. Arch. 2018, 470, 733–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Idikuda, V.; Chowdhury, S.; Chanda, B. Activation of the Archaeal Ion Channel MthK Is Exquisitely Regulated by Temperature. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Anishkin, A.; Kung, C. Microbial Mechanosensation. Curr. Opin. Neurobiol. 2005, 15, 397–405. [Google Scholar] [CrossRef]
- Kung, C.; Martinac, B.; Sukharev, S. Mechanosensitive Channels in Microbes. Annu. Rev. Microbiol. 2010, 64, 313–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perozo, E. Gating Prokaryotic Mechanosensitive Channels. Nat. Rev. Mol. Cell Biol. 2006, 7, 109–119. [Google Scholar] [CrossRef]
- Haswell, E.S.; Phillips, R.; Rees, D.C. Mechanosensitive Channels: What Can They Do and How Do They Do It? Structure 2011, 19, 1356–1369. [Google Scholar] [CrossRef] [Green Version]
- Bruni, G.N.; Weekley, R.A.; Dodd, B.J.T.; Kralj, J.M. Voltage-Gated Calcium Flux Mediates Escherichia Coli Mechanosensation. Proc. Natl. Acad. Sci. USA 2017, 114, 9445–9450. [Google Scholar] [CrossRef] [Green Version]
- Blount, P.; Iscla, I. Life with Bacterial Mechanosensitive Channels, from Discovery to Physiology to Pharmacological Target. Microbiol. Mol. Biol. Rev. 2020, 84, e00055-19. [Google Scholar] [CrossRef]
- Johnson, S.C.; Veres, J.; Malcolm, H.R. Exploring the Diversity of Mechanosensitive Channels in Bacterial Genomes. Eur. Biophys. J. 2021, 50, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.D.; Bavi, N.; Martinac, B. Biophysical Principles of Ion-Channel-Mediated Mechanosensory Transduction. Cell Rep. 2019, 29, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Delmas, P.; Hao, J.; Rodat-Despoix, L. Molecular Mechanisms of Mechanotransduction in Mammalian Sensory Neurons. Nat. Rev. Neurosci. 2011, 12, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Kefauver, J.M.; Ward, A.B.; Patapoutian, A. Discoveries in Structure and Physiology of Mechanically Activated Ion Channels. Nature 2020, 587, 567–576. [Google Scholar] [CrossRef]
- Bass, R.B.; Strop, P.; Barclay, M.; Rees, D.C. Crystal Structure of Escherichia Coli MscS, a Voltage-Modulated and Mechanosensitive Channel. Science 2002, 298, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Steinbacher, S.; Bass, R.; Strop, P.; Rees, D.C. Structures of the Prokaryotic Mechanosensitive Channels MscL and MscS. In Current Topics in Membranes; Mechanosensitive Ion Channels, Part A; Academic Press: Cambridge, MA, USA, 2007; Volume 58, pp. 1–24. [Google Scholar]
- Zhang, Y.; Daday, C.; Gu, R.-X.; Cox, C.D.; Martinac, B.; de Groot, B.L.; Walz, T. Visualization of the Mechanosensitive Ion Channel MscS under Membrane Tension. Nature 2021, 590, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhou, H.; Chi, S.; Wang, Y.; Wang, J.; Geng, J.; Wu, K.; Liu, W.; Zhang, T.; Dong, M.-Q.; et al. Structure and Mechanogating Mechanism of the Piezo1 Channel. Nature 2018, 554, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, H.; Zhang, M.; Liu, W.; Deng, T.; Zhao, Q.; Li, Y.; Lei, J.; Li, X.; Xiao, B. Structure and Mechanogating of the Mammalian Tactile Channel PIEZO2. Nature 2019, 573, 225–229. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Guo, Y.R.; Miyagi, A.; Levring, J.; MacKinnon, R.; Scheuring, S. Force-Induced Conformational Changes in PIEZO1. Nature 2019, 573, 230–234. [Google Scholar] [CrossRef]
- Brohawn, S.G.; Campbell, E.B.; MacKinnon, R. Physical Mechanism for Gating and Mechanosensitivity of the Human TRAAK K+ Channel. Nature 2014, 516, 126–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, G.; Ollig, D.; Fuhrmann, M.; Kateriya, S.; Musti, A.M.; Bamberg, E.; Hegemann, P. Channelrhodopsin-1: A Light-Gated Proton Channel in Green Algae. Science 2002, 296, 2395–2398. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, K.; Hegemann, P. The Form and Function of Channelrhodopsin. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, H.E. Structure-Function Relationship of Channelrhodopsins. Adv. Exp. Med. Biol. 2021, 1293, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Grimm, C.; Hegemann, P. Biophysics of Channelrhodopsin. Annu. Rev. Biophys. 2015, 44, 167–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lórenz-Fonfría, V.A.; Heberle, J. Channelrhodopsin Unchained: Structure and Mechanism of a Light-Gated Cation Channel. Biochim. Biophys. Acta 2014, 1837, 626–642. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Nomura, T.; Nakane, T.; Yamashita, K.; Inoue, K.; Ito, S.; Vierock, J.; Hirata, K.; Maturana, A.D.; Katayama, K.; et al. Time-Resolved Serial Femtosecond Crystallography Reveals Early Structural Changes in Channelrhodopsin. Elife 2021, 10, e62389. [Google Scholar] [CrossRef]
- Govorunova, E.G.; Sineshchekov, O.A.; Janz, R.; Liu, X.; Spudich, J.L. NEUROSCIENCE. Natural Light-Gated Anion Channels: A Family of Microbial Rhodopsins for Advanced Optogenetics. Science 2015, 349, 647–650. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Kato, H.E.; Yamashita, K.; Ito, S.; Inoue, K.; Ramakrishnan, C.; Fenno, L.E.; Evans, K.E.; Paggi, J.M.; Dror, R.O.; et al. Crystal Structure of the Natural Anion-Conducting Channelrhodopsin GtACR1. Nature 2018, 561, 343–348. [Google Scholar] [CrossRef]
- Sineshchekov, O.A.; Govorunova, E.G.; Li, H.; Spudich, J.L. Bacteriorhodopsin-like Channelrhodopsins: Alternative Mechanism for Control of Cation Conductance. Proc. Natl. Acad. Sci. USA 2017, 114, E9512–E9519. [Google Scholar] [CrossRef] [Green Version]
- Zabelskii, D.; Alekseev, A.; Kovalev, K.; Rankovic, V.; Balandin, T.; Soloviov, D.; Bratanov, D.; Savelyeva, E.; Podolyak, E.; Volkov, D.; et al. Viral Rhodopsins 1 Are an Unique Family of Light-Gated Cation Channels. Nat. Commun. 2020, 11, 5707. [Google Scholar] [CrossRef]
- Smart, T.G.; Paoletti, P. Synaptic Neurotransmitter-Gated Receptors. Cold Spring Harb. Perspect. Biol. 2012, 4, a009662. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.J.; Baenziger, J.E. Ion Channels as Lipid Sensors: From Structures to Mechanisms. Nat. Chem. Biol. 2020, 16, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Changeux, J.-P.; Christopoulos, A. Allosteric Modulation as a Unifying Mechanism for Receptor Function and Regulation. Cell 2016, 166, 1084–1102. [Google Scholar] [CrossRef] [PubMed]
- Dent, J.A. The Evolution of Pentameric Ligand-Gated Ion Channels. Adv. Exp. Med. Biol. 2010, 683, 11–23. [Google Scholar] [CrossRef]
- Jaiteh, M.; Taly, A.; Hénin, J. Evolution of Pentameric Ligand-Gated Ion Channels: Pro-Loop Receptors. PLoS ONE 2016, 11, e0151934. [Google Scholar] [CrossRef] [Green Version]
- Hilf, R.J.; Dutzler, R. A Prokaryotic Perspective on Pentameric Ligand-Gated Ion Channel Structure. Curr. Opin. Struct. Biol. 2009, 19, 418–424. [Google Scholar] [CrossRef]
- Tasneem, A.; Iyer, L.M.; Jakobsson, E.; Aravind, L. Identification of the Prokaryotic Ligand-Gated Ion Channels and Their Implications for the Mechanisms and Origins of Animal Cys-Loop Ion Channels. Genome Biol. 2005, 6, R4. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C.R. The Structural Basis of Function in Cys-Loop Receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [Green Version]
- Corringer, P.-J.; Poitevin, F.; Prevost, M.S.; Sauguet, L.; Delarue, M.; Changeux, J.-P. Structure and Pharmacology of Pentameric Receptor Channels: From Bacteria to Brain. Structure 2012, 20, 941–956. [Google Scholar] [CrossRef] [Green Version]
- Hilf, R.J.C.; Dutzler, R. X-Ray Structure of a Prokaryotic Pentameric Ligand-Gated Ion Channel. Nature 2008, 452, 375–379. [Google Scholar] [CrossRef]
- Hilf, R.J.C.; Dutzler, R. Structure of a Potentially Open State of a Proton-Activated Pentameric Ligand-Gated Ion Channel. Nature 2009, 457, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, N.; Prado de Carvalho, L.; Cartaud, J.; Neyton, J.; Le Poupon, C.; Taly, A.; Grutter, T.; Changeux, J.-P.; Corringer, P.-J. A Prokaryotic Proton-Gated Ion Channel from the Nicotinic Acetylcholine Receptor Family. Nature 2007, 445, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Cheng, H.; Jiang, Y.; Melcher, K.; Xu, H.E. Ion Channels Gated by Acetylcholine and Serotonin: Structures, Biology, and Drug Discovery. Acta Pharmacol. Sin. 2015, 36, 895–907. [Google Scholar] [CrossRef]
- Scott, S.; Aricescu, A.R. A Structural Perspective on GABAA Receptor Pharmacology. Curr. Opin. Struct. Biol. 2019, 54, 189–197. [Google Scholar] [CrossRef]
- Hibbs, R.E.; Gouaux, E. Principles of Activation and Permeation in an Anion-Selective Cys-Loop Receptor. Nature 2011, 474, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.S.; Aricescu, A.R. Crystal Structure of a Human GABAA Receptor. Nature 2014, 512, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Masiulis, S.; Desai, R.; Uchański, T.; Serna Martin, I.; Laverty, D.; Karia, D.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Kotecha, A.; et al. GABAA Receptor Signalling Mechanisms Revealed by Structural Pharmacology. Nature 2019, 565, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Brejc, K.; van Dijk, W.J.; Klaassen, R.V.; Schuurmans, M.; van der Oost, J.; Smit, A.B.; Sixma, T.K. Crystal Structure of an ACh-Binding Protein Reveals the Ligand-Binding Domain of Nicotinic Receptors. Nature 2001, 411, 269–276. [Google Scholar] [CrossRef]
- Purohit, P.; Mitra, A.; Auerbach, A. A Stepwise Mechanism for Acetylcholine Receptor Channel Gating. Nature 2007, 446, 930–933. [Google Scholar] [CrossRef]
- Gupta, S.; Chakraborty, S.; Vij, R.; Auerbach, A. A Mechanism for Acetylcholine Receptor Gating Based on Structure, Coupling, Phi, and Flip. J. Gen. Physiol. 2017, 149, 85–103. [Google Scholar] [CrossRef]
- Polovinkin, L.; Hassaine, G.; Perot, J.; Neumann, E.; Jensen, A.A.; Lefebvre, S.N.; Corringer, P.-J.; Neyton, J.; Chipot, C.; Dehez, F.; et al. Conformational Transitions of the Serotonin 5-HT3 Receptor. Nature 2018, 563, 275–279. [Google Scholar] [CrossRef]
- Chen, Y.; Reilly, K.; Chang, Y. Evolutionarily Conserved Allosteric Network in the Cys Loop Family of Ligand-Gated Ion Channels Revealed by Statistical Covariance Analyses. J. Biol. Chem. 2006, 281, 18184–18192. [Google Scholar] [CrossRef] [Green Version]
- Calimet, N.; Simoes, M.; Changeux, J.-P.; Karplus, M.; Taly, A.; Cecchini, M. A Gating Mechanism of Pentameric Ligand-Gated Ion Channels. Proc. Natl. Acad. Sci. USA 2013, 110, E3987–E3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Huang, Y.; Whiteaker, P. Mechanism of Allosteric Modulation of the Cys-Loop Receptors. Pharmaceuticals 2010, 3, 2592–2609. [Google Scholar] [CrossRef] [Green Version]
- Gharpure, A.; Noviello, C.M.; Hibbs, R.E. Progress in Nicotinic Receptor Structural Biology. Neuropharmacology 2020, 171, 108086. [Google Scholar] [CrossRef] [PubMed]
- Lummis, S.C.R.; McGonigle, I.; Ashby, J.A.; Dougherty, D.A. Two Amino Acid Residues Contribute to a Cation-π Binding Interaction in the Binding Site of an Insect GABA Receptor. J. Neurosci. 2011, 31, 12371–12376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiu, X.; Puskar, N.L.; Shanata, J.A.P.; Lester, H.A.; Dougherty, D.A. Nicotine Binding to Brain Receptors Requires a Strong Cation-Pi Interaction. Nature 2009, 458, 534–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, J.; Mayer, M.L. Functional Insights from Glutamate Receptor Ion Channel Structures. Annu. Rev. Physiol. 2013, 75, 313–337. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.L. Glutamate Receptors at Atomic Resolution. Nature 2006, 440, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Greger, I.H.; Watson, J.F.; Cull-Candy, S.G. Structural and Functional Architecture of AMPA-Type Glutamate Receptors and Their Auxiliary Proteins. Neuron 2017, 94, 713–730. [Google Scholar] [CrossRef]
- Chen, S.; Gouaux, E. Structure and Mechanism of AMPA Receptor—Auxiliary Protein Complexes. Curr. Opin. Struct. Biol. 2019, 54, 104–111. [Google Scholar] [CrossRef]
- Kamalova, A.; Nakagawa, T. AMPA Receptor Structure and Auxiliary Subunits. J. Physiol. 2021, 599, 453–469. [Google Scholar] [CrossRef]
- Meyerson, J.R.; Kumar, J.; Chittori, S.; Rao, P.; Pierson, J.; Bartesaghi, A.; Mayer, M.L.; Subramaniam, S. Structural Mechanism of Glutamate Receptor Activation and Desensitization. Nature 2014, 514, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Sobolevsky, A.I.; Rosconi, M.P.; Gouaux, E. X-Ray Structure, Symmetry and Mechanism of an AMPA-Subtype Glutamate Receptor. Nature 2009, 462, 745–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Chen, S.; Swensen, A.C.; Qian, W.-J.; Gouaux, E. Architecture and Subunit Arrangement of Native AMPA Receptors Elucidated by Cryo-EM. Science 2019, 364, 355–362. [Google Scholar] [CrossRef]
- Puljung, M.C. Cryo-Electron Microscopy Structures and Progress toward a Dynamic Understanding of KATP Channels. J. Gen. Physiol. 2018, 150, 653–669. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.G. KATP Channels as Molecular Sensors of Cellular Metabolism. Nature 2006, 440, 470–476. [Google Scholar] [CrossRef]
- Lee, K.P.K.; Chen, J.; MacKinnon, R. Molecular Structure of Human KATP in Complex with ATP and ADP. Elife 2017, 6. [Google Scholar] [CrossRef]
- Martin, G.M.; Sung, M.W.; Yang, Z.; Innes, L.M.; Kandasamy, B.; David, L.L.; Yoshioka, C.; Shyng, S.-L. Mechanism of Pharmacochaperoning in a Mammalian KATP Channel Revealed by Cryo-EM. Elife 2019, 8, e46417. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-Transmembrane Receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef]
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal Structure of Rhodopsin: A G Protein-Coupled Receptor. Science 2000, 289, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Gushchin, I.; Shevchenko, V.; Polovinkin, V.; Kovalev, K.; Alekseev, A.; Round, E.; Borshchevskiy, V.; Balandin, T.; Popov, A.; Gensch, T.; et al. Crystal Structure of a Light-Driven Sodium Pump. Nat. Struct. Mol. Biol. 2015, 22, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.E.; Inoue, K.; Abe-Yoshizumi, R.; Kato, Y.; Ono, H.; Konno, M.; Hososhima, S.; Ishizuka, T.; Hoque, M.R.; Kunitomo, H.; et al. Structural Basis for Na(+) Transport Mechanism by a Light-Driven Na(+) Pump. Nature 2015, 521, 48–53. [Google Scholar] [CrossRef]
- Shalaeva, D.N.; Galperin, M.Y.; Mulkidjanian, A.Y. Eukaryotic G Protein-Coupled Receptors as Descendants of Prokaryotic Sodium-Translocating Rhodopsins. Biol. Direct. 2015, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Mendoza, A.; Sebé-Pedrós, A.; Ruiz-Trillo, I. The Evolution of the GPCR Signaling System in Eukaryotes: Modularity, Conservation, and the Transition to Metazoan Multicellularity. Genome Biol. Evol. 2014, 6, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Almén, M.S.; Fredriksson, R.; Schiöth, H.B. The Origin of GPCRs: Identification of Mammalian like Rhodopsin, Adhesion, Glutamate and Frizzled GPCRs in Fungi. PLoS ONE 2012, 7, e29817. [Google Scholar] [CrossRef]
- Strotmann, R.; Schröck, K.; Böselt, I.; Stäubert, C.; Russ, A.; Schöneberg, T. Evolution of GPCR: Change and Continuity. Mol. Cell Endocrinol 2011, 331, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Kooistra, A.J.; Mordalski, S.; Pándy-Szekeres, G.; Esguerra, M.; Mamyrbekov, A.; Munk, C.; Keserű, G.M.; Gloriam, D.E. GPCRdb in 2021: Integrating GPCR Sequence, Structure and Function. Nucleic Acids Res. 2021, 49, D335–D343. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Schiöth, H.B. The Role of G Protein-Coupled Receptors in the Early Evolution of Neurotransmission and the Nervous System. J. Exp. Biol. 2015, 218, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G Protein Activation by G-Protein-Coupled Receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and Dynamics of GPCR Signaling Complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.G.F.; DeVree, B.T.; Zou, Y.; Kruse, A.C.; Chung, K.Y.; Kobilka, T.S.; Thian, F.S.; Chae, P.S.; Pardon, E.; Calinski, D.; et al. Crystal Structure of the Β2 Adrenergic Receptor-Gs Protein Complex. Nature 2011, 477, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.Y.; Rasmussen, S.G.F.; Liu, T.; Li, S.; DeVree, B.T.; Chae, P.S.; Calinski, D.; Kobilka, B.K.; Woods, V.L.; Sunahara, R.K. Conformational Changes in the G Protein Gs Induced by the Β2 Adrenergic Receptor. Nature 2011, 477, 611–615. [Google Scholar] [CrossRef] [Green Version]
- García-Nafría, J.; Tate, C.G. Cryo-EM Structures of GPCRs Coupled to Gs, Gi and Go. Mol. Cell Endocrinol. 2019, 488, 1–13. [Google Scholar] [CrossRef]
- Glukhova, A.; Draper-Joyce, C.J.; Sunahara, R.K.; Christopoulos, A.; Wootten, D.; Sexton, P.M. Rules of Engagement: GPCRs and G Proteins. ACS Pharm. Transl. Sci. 2018, 1, 73–83. [Google Scholar] [CrossRef]
- Grisshammer, R. The Quest for High-Resolution G Protein-Coupled Receptor–G Protein Structures. Proc. Natl. Acad. Sci. USA 2020, 117, 6971–6973. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, L.; Zhao, S. Ligands of Adrenergic Receptors: A Structural Point of View. Biomolecules 2021, 11, 936. [Google Scholar] [CrossRef]
- Ognjenović, J.; Grisshammer, R.; Subramaniam, S. Frontiers in Cryo Electron Microscopy of Complex Macromolecular Assemblies. Annu. Rev. Biomed. Eng. 2019, 21, 395–415. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common Activation Mechanism of Class A GPCRs. Elife 2019, 8, e50279. [Google Scholar] [CrossRef]
- Knight, K.M.; Ghosh, S.; Campbell, S.L.; Lefevre, T.J.; Olsen, R.H.J.; Smrcka, A.V.; Valentin, N.H.; Yin, G.; Vaidehi, N.; Dohlman, H.G. A Universal Allosteric Mechanism for G Protein Activation. Mol. Cell 2021, 81, 1384–1396.e6. [Google Scholar] [CrossRef]
- Nakashima, A.; Takeuchi, H.; Imai, T.; Saito, H.; Kiyonari, H.; Abe, T.; Chen, M.; Weinstein, L.S.; Yu, C.R.; Storm, D.R.; et al. Agonist-Independent GPCR Activity Regulates Anterior-Posterior Targeting of Olfactory Sensory Neurons. Cell 2013, 154, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, J.P.; Sunahara, R.K. Mechanistic Insights into GPCR-G Protein Interactions. Curr. Opin. Struct. Biol. 2016, 41, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Dror, R.O.; Pan, A.C.; Arlow, D.H.; Borhani, D.W.; Maragakis, P.; Shan, Y.; Xu, H.; Shaw, D.E. Pathway and Mechanism of Drug Binding to G-Protein-Coupled Receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 13118–13123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawson, T. Specificity in Signal Transduction: From Phosphotyrosine-SH2 Domain Interactions to Complex Cellular Systems. Cell 2004, 116, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Marasco, M.; Carlomagno, T. Specificity and Regulation of Phosphotyrosine Signaling through SH2 Domains. J. Struct. Biol. X 2020, 4, 100026. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell Signaling by Receptor Tyrosine Kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [Green Version]
- Schlessinger, J.; Lemmon, M.A. SH2 and PTB Domains in Tyrosine Kinase Signaling. Sci. STKE 2003, 2003, RE12. [Google Scholar] [CrossRef]
- Yaffe, M.B. Phosphotyrosine-Binding Domains in Signal Transduction. Nat. Rev. Mol. Cell Biol. 2002, 3, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Braatsch, S.; Klug, G. Blue Light Perception in Bacteria. Photosynth Res. 2004, 79, 45–57. [Google Scholar] [CrossRef]
- Colley, N.J.; Nilsson, D.-E. Photoreception in Phytoplankton. Integr. Comp. Biol. 2016, 56, 764–775. [Google Scholar] [CrossRef]
- Häder, D.P. Photosensory Behavior in Procaryotes. Microbiol. Rev. 1987, 51, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hegemann, P. Algal Sensory Photoreceptors. Annu. Rev. Plant. Biol. 2008, 59, 167–189. [Google Scholar] [CrossRef]
- Herrou, J.; Crosson, S. Function, Structure and Mechanism of Bacterial Photosensory LOV Proteins. Nat. Rev. Microbiol. 2011, 9, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.-Y.; Soulier, N.T.; Canniffe, D.P.; Shen, G.; Bryant, D.A. Light Regulation of Pigment and Photosystem Biosynthesis in Cyanobacteria. Curr. Opin. Plant. Biol. 2017, 37, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Ishizuka, T. Cyanobacteriochromes: A New Superfamily of Tetrapyrrole-Binding Photoreceptors in Cyanobacteria. PhotoChem. Photobiol. Sci. 2008, 7, 1159–1167. [Google Scholar] [CrossRef]
- Jékely, G. Evolution of Phototaxis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2795–2808. [Google Scholar] [CrossRef] [PubMed]
- Kottke, T.; Xie, A.; Larsen, D.S.; Hoff, W.D. Photoreceptors Take Charge: Emerging Principles for Light Sensing. Annu. Rev. Biophys. 2018, 47, 291–313. [Google Scholar] [CrossRef]
- Ueki, N.; Matsunaga, S.; Inouye, I.; Hallmann, A. How 5000 Independent Rowers Coordinate Their Strokes in Order to Row into the Sunlight: Phototaxis in the Multicellular Green Alga Volvox. BMC Biol. 2010, 8, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möglich, A. Signal Transduction in Photoreceptor Histidine Kinases. Protein Sci. 2019, 28, 1923–1946. [Google Scholar] [CrossRef]
- Wilde, A.; Mullineaux, C.W. Light-Controlled Motility in Prokaryotes and the Problem of Directional Light Perception. FEMS Microbiol. Rev. 2017, 41, 900–922. [Google Scholar] [CrossRef] [Green Version]
- Schuergers, N.; Lenn, T.; Kampmann, R.; Meissner, M.V.; Esteves, T.; Temerinac-Ott, M.; Korvink, J.G.; Lowe, A.R.; Mullineaux, C.W.; Wilde, A. Cyanobacteria Use Micro-Optics to Sense Light Direction. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavelis, G.S.; Hayakawa, S.; White, R.A.; Gojobori, T.; Suttle, C.A.; Keeling, P.J.; Leander, B.S. Eye-like Ocelloids Are Built from Different Endosymbiotically Acquired Components. Nature 2015, 523, 204–207. [Google Scholar] [CrossRef]
- Möglich, A.; Yang, X.; Ayers, R.A.; Moffat, K. Structure and Function of Plant Photoreceptors. Annu. Rev. Plant. Biol. 2010, 61, 21–47. [Google Scholar] [CrossRef] [Green Version]
- Conrad, K.S.; Manahan, C.C.; Crane, B.R. Photochemistry of Flavoprotein Light Sensors. Nat. Chem. Biol. 2014, 10, 801–809. [Google Scholar] [CrossRef]
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and Animal Rhodopsins: Structures, Functions, and Molecular Mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef]
- Butryn, A.; Raza, H.; Rada, H.; Moraes, I.; Owens, R.J.; Orville, A.M. Molecular Basis for GTP Recognition by Light-Activated Guanylate Cyclase RhGC. FEBS J. 2020, 287, 2797–2807. [Google Scholar] [CrossRef] [Green Version]
- Avelar, G.M.; Schumacher, R.I.; Zaini, P.A.; Leonard, G.; Richards, T.A.; Gomes, S.L. A Rhodopsin-Guanylyl Cyclase Gene Fusion Functions in Visual Perception in a Fungus. Curr. Biol. 2014, 24, 1234–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikuta, T.; Shihoya, W.; Sugiura, M.; Yoshida, K.; Watari, M.; Tokano, T.; Yamashita, K.; Katayama, K.; Tsunoda, S.P.; Uchihashi, T.; et al. Structural Insights into the Mechanism of Rhodopsin Phosphodiesterase. Nat. Commun. 2020, 11, 5605. [Google Scholar] [CrossRef] [PubMed]
- Borgstahl, G.E.O.; Williams, D.R.; Getzoff, E.D. 1.4.ANG. Structure of Photoactive Yellow Protein, a Cytosolic Photoreceptor: Unusual Fold, Active Site, and Chromophore. Biochemistry 1995, 34, 6278–6287. [Google Scholar] [CrossRef]
- Pellequer, J.L.; Wager-Smith, K.A.; Kay, S.A.; Getzoff, E.D. Photoactive Yellow Protein: A Structural Prototype for the Three-Dimensional Fold of the PAS Domain Superfamily. Proc. Natl. Acad. Sci. USA 1998, 95, 5884–5890. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.L.; Zhulin, I.B. PAS Domains: Internal Sensors of Oxygen, Redox Potential, and Light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. A Short History of Structure Based Research on the Photocycle of Photoactive Yellow Protein. Struct. Dyn. 2017, 4, 032201. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.O.; Lee, J.H.; Kim, J.; Schmidt, M.; Moffat, K.; Srajer, V.; Ihee, H. Volume-Conserving Trans-Cis Isomerization Pathways in Photoactive Yellow Protein Visualized by Picosecond X-Ray Crystallography. Nat. Chem. 2013, 5, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigala, P.A.; Tsuchida, M.A.; Herschlag, D. Hydrogen Bond Dynamics in the Active Site of Photoactive Yellow Protein. Proc. Natl. Acad. Sci. USA 2009, 106, 9232–9237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glantz, S.T.; Carpenter, E.J.; Melkonian, M.; Gardner, K.H.; Boyden, E.S.; Wong, G.K.-S.; Chow, B.Y. Functional and Topological Diversity of LOV Domain Photoreceptors. Proc. Natl. Acad. Sci. USA 2016, 113, E1442–E1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.M.; Kaiser, J.; Ziegler, T.; Pilsl, S.; Renzl, C.; Sixt, L.; Pietruschka, G.; Moniot, S.; Kakoti, A.; Juraschitz, M.; et al. A Blue Light Receptor That Mediates RNA Binding and Translational Regulation. Nat. Chem. Biol. 2019, 15, 1085–1092. [Google Scholar] [CrossRef]
- Harper, S.M.; Neil, L.C.; Gardner, K.H. Structural Basis of a Phototropin Light Switch. Science 2003, 301, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Okajima, K. Molecular Mechanism of Phototropin Light Signaling. J. Plant. Res. 2016, 129, 149–157. [Google Scholar] [CrossRef]
- Möglich, A.; Moffat, K. Structural Basis for Light-Dependent Signaling in the Dimeric LOV Domain of the Photosensor YtvA. J. Mol. Biol. 2007, 373, 112–126. [Google Scholar] [CrossRef] [Green Version]
- Barends, T.R.M.; Hartmann, E.; Griese, J.J.; Beitlich, T.; Kirienko, N.V.; Ryjenkov, D.A.; Reinstein, J.; Shoeman, R.L.; Gomelsky, M.; Schlichting, I. Structure and Mechanism of a Bacterial Light-Regulated Cyclic Nucleotide Phosphodiesterase. Nature 2009, 459, 1015–1018. [Google Scholar] [CrossRef]
- Kennis, J.T.M.; Mathes, T. Molecular Eyes: Proteins That Transform Light into Biological Information. Interface Focus 2013, 3, 20130005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Masuda, S. Light-Induced Chromophore and Protein Responses and Mechanical Signal Transduction of BLUF Proteins. Biophys. Rev. 2018, 10, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Tame, J.R.H. Seeing the Light with BLUF Proteins. Biophys. Rev. 2017, 9, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutomi, O.; Yamamoto, R.; Hirose, K.; Mizuno, K.; Nakagiri, Y.; Imai, H.; Noga, A.; Obbineni, J.M.; Zimmermann, N.; Nakajima, M.; et al. A Dynein-Associated Photoreceptor Protein Prevents Ciliary Acclimation to Blue Light. Sci. Adv. 2021, 7, eabf3621. [Google Scholar] [CrossRef]
- Jung, A.; Domratcheva, T.; Tarutina, M.; Wu, Q.; Ko, W.-H.; Shoeman, R.L.; Gomelsky, M.; Gardner, K.H.; Schlichting, I. Structure of a Bacterial BLUF Photoreceptor: Insights into Blue Light-Mediated Signal Transduction. Proc. Natl. Acad. Sci. USA 2005, 102, 12350–12355. [Google Scholar] [CrossRef] [Green Version]
- Masuda, S.; Bauer, C.E. AppA Is a Blue Light Photoreceptor That Antirepresses Photosynthesis Gene Expression in Rhodobacter Sphaeroides. Cell 2002, 110, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Winkler, A.; Heintz, U.; Lindner, R.; Reinstein, J.; Shoeman, R.L.; Schlichting, I. A Ternary AppA-PpsR-DNA Complex Mediates Light Regulation of Photosynthesis-Related Gene Expression. Nat. Struct. Mol. Biol. 2013, 20, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Ohki, M.; Sugiyama, K.; Kawai, F.; Tanaka, H.; Nihei, Y.; Unzai, S.; Takebe, M.; Matsunaga, S.; Adachi, S.-I.; Shibayama, N.; et al. Structural Insight into Photoactivation of an Adenylate Cyclase from a Photosynthetic Cyanobacterium. Proc. Natl. Acad. Sci. USA 2016, 113, 6659–6664. [Google Scholar] [CrossRef] [Green Version]
- Lindner, R.; Hartmann, E.; Tarnawski, M.; Winkler, A.; Frey, D.; Reinstein, J.; Meinhart, A.; Schlichting, I. Photoactivation Mechanism of a Bacterial Light-Regulated Adenylyl Cyclase. J. Mol. Biol. 2017, 429, 1336–1351. [Google Scholar] [CrossRef]
- Brautigam, C.A.; Smith, B.S.; Ma, Z.; Palnitkar, M.; Tomchick, D.R.; Machius, M.; Deisenhofer, J. Structure of the Photolyase-like Domain of Cryptochrome 1 from Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 12142–12147. [Google Scholar] [CrossRef] [Green Version]
- Mei, Q.; Dvornyk, V. Evolutionary History of the Photolyase/Cryptochrome Superfamily in Eukaryotes. PLoS ONE 2015, 10, e0135940. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.; Jaskolski, M.; Davis, S.J. Circadian Oscillator Proteins across the Kingdoms of Life: Structural Aspects. BMC Biol. 2019, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Zoltowski, B.D. Resolving Cryptic Aspects of Cryptochrome Signaling. Proc. Natl. Acad. Sci. USA 2015, 112, 8811–8812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Wang, X.; Zhang, M.; Bian, M.; Deng, W.; Zuo, Z.; Yang, Z.; Zhong, D.; Lin, C. Trp Triad-Dependent Rapid Photoreduction Is Not Required for the Function of Arabidopsis CRY1. Proc. Natl. Acad. Sci. USA 2015, 112, 9135–9140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Lin, C. A Structural View of Plant CRY2 Photoactivation and Inactivation. Nat. Struct. Mol. Biol. 2020, 27, 401–403. [Google Scholar] [CrossRef]
- Wang, Q.; Lin, C. Mechanisms of Cryptochrome-Mediated Photoresponses in Plants. Annu. Rev. Plant. Biol. 2020, 71, 103–129. [Google Scholar] [CrossRef] [Green Version]
- Palayam, M.; Ganapathy, J.; Guercio, A.M.; Tal, L.; Deck, S.L.; Shabek, N. Structural Insights into Photoactivation of Plant Cryptochrome-2. Commun. Biol. 2021, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.R.; Brunzelle, J.S.; Forest, K.T.; Vierstra, R.D. A Light-Sensing Knot Revealed by the Structure of the Chromophore-Binding Domain of Phytochrome. Nature 2005, 438, 325–331. [Google Scholar] [CrossRef]
- Essen, L.-O.; Mailliet, J.; Hughes, J. The Structure of a Complete Phytochrome Sensory Module in the Pr Ground State. Proc. Natl. Acad. Sci. USA 2008, 105, 14709–14714. [Google Scholar] [CrossRef] [Green Version]
- Rockwell, N.C.; Lagarias, J.C. Phytochrome Evolution in 3D: Deletion, Duplication, and Diversification. New Phytol. 2020, 225, 2283–2300. [Google Scholar] [CrossRef] [Green Version]
- Nagano, S. From Photon to Signal in Phytochromes: Similarities and Differences between Prokaryotic and Plant Phytochromes. J. Plant. Res. 2016, 129, 123–135. [Google Scholar] [CrossRef]
- Woitowich, N.C.; Halavaty, A.S.; Waltz, P.; Kupitz, C.; Valera, J.; Tracy, G.; Gallagher, K.D.; Claesson, E.; Nakane, T.; Pandey, S.; et al. Structural Basis for Light Control of Cell Development Revealed by Crystal Structures of a Myxobacterial Phytochrome. IUCrJ 2018, 5, 619–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgie, E.S.; Zhang, J.; Vierstra, R.D. Crystal Structure of Deinococcus Phytochrome in the Photoactivated State Reveals a Cascade of Structural Rearrangements during Photoconversion. Structure 2016, 24, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Takala, H.; Björling, A.; Berntsson, O.; Lehtivuori, H.; Niebling, S.; Hoernke, M.; Kosheleva, I.; Henning, R.; Menzel, A.; Ihalainen, J.A.; et al. Signal Amplification and Transduction in Phytochrome Photosensors. Nature 2014, 509, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakemore, R.P. Magnetotactic Bacteria. Annu. Rev. Microbiol. 1982, 36, 217–238. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Wu, L.-F. Flagella and Swimming Behavior of Marine Magnetotactic Bacteria. Biomolecules 2020, 10, 460. [Google Scholar] [CrossRef] [Green Version]
- Vincenti, B.; Ramos, G.; Cordero, M.L.; Douarche, C.; Soto, R.; Clement, E. Magnetotactic Bacteria in a Droplet Self-Assemble into a Rotary Motor. Nat. Commun. 2019, 10, 5082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uebe, R.; Schüler, D. Magnetosome Biogenesis in Magnetotactic Bacteria. Nat. Rev. Microbiol. 2016, 14, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Lower, B.H.; Bazylinski, D.A. The Bacterial Magnetosome: A Unique Prokaryotic Organelle. J. Mol. Microbiol. Biotechnol. 2013, 23, 63–80. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Wu, L.-F. Evolution of the Bacterial Organelle Responsible for Magnetotaxis. Trends Microbiol. 2013, 21, 534–543. [Google Scholar] [CrossRef]
- Popp, F.; Armitage, J.P.; Schüler, D. Polarity of Bacterial Magnetotaxis Is Controlled by Aerotaxis through a Common Sensory Pathway. Nat. Commun. 2014, 5, 5398. [Google Scholar] [CrossRef] [Green Version]
- Davidov, G.; Müller, F.D.; Baumgartner, J.; Bitton, R.; Faivre, D.; Schüler, D.; Zarivach, R. Crystal Structure of the Magnetobacterial Protein MtxA C-Terminal Domain Reveals a New Sequence-Structure Relationship. Front. Mol. Biosci. 2015, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hore, P.J.; Mouritsen, H. The Radical-Pair Mechanism of Magnetoreception. Annu. Rev. Biophys. 2016, 45, 299–344. [Google Scholar] [CrossRef]
- Evans, E.W.; Dodson, C.A.; Maeda, K.; Biskup, T.; Wedge, C.J.; Timmel, C.R. Magnetic Field Effects in Flavoproteins and Related Systems. Interface Focus 2013, 3, 20130037. [Google Scholar] [CrossRef] [Green Version]
- Pané-Farré, J.; Quin, M.B.; Lewis, R.J.; Marles-Wright, J. Structure and Function of the Stressosome Signalling Hub. Subcell Biochem. 2017, 83, 1–41. [Google Scholar] [CrossRef]
- Marles-Wright, J.; Grant, T.; Delumeau, O.; van Duinen, G.; Firbank, S.J.; Lewis, P.J.; Murray, J.W.; Newman, J.A.; Quin, M.B.; Race, P.R.; et al. Molecular Architecture of the “Stressosome,” a Signal Integration and Transduction Hub. Science 2008, 322, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Redzej, A.; Rolhion, N.; Costa, T.R.D.; Rifflet, A.; Waksman, G.; Cossart, P. The Cryo-Electron Microscopy Supramolecular Structure of the Bacterial Stressosome Unveils Its Mechanism of Activation. Nat. Commun. 2019, 10, 3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, P.; Garrity, P. Sensing Temperature. Curr. Biol. 2013, 23, R304–R307. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.; Triandafillou, C.; Drummond, D.A. Cellular Sensing by Phase Separation: Using the Process, Not Just the Products. J. Biol. Chem. 2019, 294, 7151–7159. [Google Scholar] [CrossRef] [Green Version]
- Browse, J.; Xin, Z. Temperature Sensing and Cold Acclimation. Curr. Opin. Plant. Biol. 2001, 4, 241–246. [Google Scholar] [CrossRef]
- Morita, M.; Kanemori, M.; Yanagi, H.; Yura, T. Heat-Induced Synthesis of Sigma32 in Escherichia Coli: Structural and Functional Dissection of RpoH MRNA Secondary Structure. J. Bacteriol 1999, 181, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso-Becera, M.V.; Servín-González, L.; Soberón-Chávez, G. RNA Structures Are Involved in the Thermoregulation of Bacterial Virulence-Associated Traits. Trends Microbiol. 2015, 23, 509–518. [Google Scholar] [CrossRef]
- Righetti, F.; Narberhaus, F. How to Find RNA Thermometers. Front. Cell Infect. Microbiol. 2014, 4, 132. [Google Scholar] [CrossRef] [Green Version]
- Kortmann, J.; Narberhaus, F. Bacterial RNA Thermometers: Molecular Zippers and Switches. Nat. Rev. Microbiol. 2012, 10, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Loh, E.; Righetti, F.; Eichner, H.; Twittenhoff, C.; Narberhaus, F. RNA Thermometers in Bacterial Pathogens. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Dong, X.; Qu, G.; Piazza, C.L.; Belfort, M. Group II Intron as Cold Sensor for Self-Preservation and Bacterial Conjugation. Nucleic Acids Res. 2020, 48, 6198–6209. [Google Scholar] [CrossRef]
- Bastet, L.; Turcotte, P.; Wade, J.T.; Lafontaine, D.A. Maestro of Regulation: Riboswitches Orchestrate Gene Expression at the Levels of Translation, Transcription and MRNA Decay. RNA Biol. 2018, 15, 679–682. [Google Scholar] [CrossRef]
- Breaker, R.R. Riboswitches and Translation Control. Cold Spring Harb. Perspect. Biol. 2018, 10, a032797. [Google Scholar] [CrossRef] [PubMed]
- McCown, P.J.; Corbino, K.A.; Stav, S.; Sherlock, M.E.; Breaker, R.R. Riboswitch Diversity and Distribution. RNA 2017, 23, 995–1011. [Google Scholar] [CrossRef]
- Henkin, T.M. Riboswitch RNAs: Using RNA to Sense Cellular Metabolism. Genes Dev. 2008, 22, 3383–3390. [Google Scholar] [CrossRef] [Green Version]
- Grojean, J.; Downes, B. Riboswitches as Hormone Receptors: Hypothetical Cytokinin-Binding Riboswitches in Arabidopsis Thaliana. Biol. Direct. 2010, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Michnicka, M.; Zhang, Y.; Wang, Y.-X.; Nikonowicz, E.P. Capture and Release of TRNA by the T-Loop Receptor in the Function of the T-Box Riboswitch. Biochemistry 2017, 56, 3549–3558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, E.C. RNA Sensors and Riboswitches: Self-Regulating Messages. Curr. Biol. 2003, 13, R285–R291. [Google Scholar] [CrossRef] [Green Version]
- Bębenek, A.; Ziuzia-Graczyk, I. Fidelity of DNA Replication-a Matter of Proofreading. Curr. Genet. 2018, 64, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Timsit, Y. DNA Structure and Polymerase Fidelity. J. Mol. Biol. 1999, 293, 835–853. [Google Scholar] [CrossRef]
- Timsit, Y. Local Sensing of Global DNA Topology: From Crossover Geometry to Type II Topoisomerase Processivity. Nucleic Acids Res. 2011, 39, 8665–8676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwang, T.J.; Tse, E.C.M.; Barton, J.K. Sensing DNA through DNA Charge Transport. ACS Chem. Biol. 2018, 13, 1799–1809. [Google Scholar] [CrossRef]
- Sontz, P.A.; Muren, N.B.; Barton, J.K. DNA Charge Transport for Sensing and Signaling. Acc. Chem. Res. 2012, 45, 1792–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvez, S.; Long, M.J.C.; Poganik, J.R.; Aye, Y. Redox Signaling by Reactive Electrophiles and Oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Adler, J.; Tso, W.W. “Decision”-Making in Bacteria: Chemotactic Response of Escherichia Coli to Conflicting Stimuli. Science 1974, 184, 1292–1294. [Google Scholar] [CrossRef]
- Adler, J. Effects of Amino Acids and Oxygen on Chemotaxis in Escherichia coli. J. Bacteriol. 1966, 92, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshland, D.E. A Response Regulator Model in a Simple Sensory System. Science 1977, 196, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Koshland, D.E.; Goldbeter, A.; Stock, J.B. Amplification and Adaptation in Regulatory and Sensory Systems. Science 1982, 217, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, J.J.E.; Groisman, E.A. Making Informed Decisions: Regulatory Interactions between Two-Component Systems. Trends Microbiol. 2003, 11, 359–366. [Google Scholar] [CrossRef]
- Baker, M.D.; Wolanin, P.M.; Stock, J.B. Signal Transduction in Bacterial Chemotaxis. Bioessays 2006, 28, 9–22. [Google Scholar] [CrossRef]
- Micali, G.; Endres, R.G. Bacterial Chemotaxis: Information Processing, Thermodynamics, and Behavior. Curr. Opin. Microbiol. 2016, 30, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Hellingwerf, K.J. Bacterial Observations: A Rudimentary Form of Intelligence? Trends Microbiol. 2005, 13, 152–158. [Google Scholar] [CrossRef]
- Hazelbauer, G.L.; Falke, J.J.; Parkinson, J.S. Bacterial Chemoreceptors: High-Performance Signaling in Networked Arrays. Trends Biochem. Sci. 2008, 33, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Falke, J.J.; Piasta, K.N. Architecture and Signal Transduction Mechanism of the Bacterial Chemosensory Array: Progress, Controversies, and Challenges. Curr. Opin. Struct. Biol. 2014, 29, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkinson, J.S.; Hazelbauer, G.L.; Falke, J.J. Signaling and Sensory Adaptation in Escherichia Coli Chemoreceptors: 2015 Update. Trends Microbiol. 2015, 23, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Wadhams, G.H.; Armitage, J.P. Making Sense of It All: Bacterial Chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Sourjik, V.; Wingreen, N.S. Responding to Chemical Gradients: Bacterial Chemotaxis. Curr. Opin. Cell Biol. 2012, 24, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Muok, A.R.; Briegel, A.; Crane, B.R. Regulation of the Chemotaxis Histidine Kinase CheA: A Structural Perspective. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183030. [Google Scholar] [CrossRef]
- Cassidy, C.K.; Himes, B.A.; Sun, D.; Ma, J.; Zhao, G.; Parkinson, J.S.; Stansfeld, P.J.; Luthey-Schulten, Z.; Zhang, P. Structure and Dynamics of the E. Coli Chemotaxis Core Signaling Complex by Cryo-Electron Tomography and Molecular Simulations. Commun. Biol. 2020, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briegel, A.; Wong, M.L.; Hodges, H.L.; Oikonomou, C.M.; Piasta, K.N.; Harris, M.J.; Fowler, D.J.; Thompson, L.K.; Falke, J.J.; Kiessling, L.L.; et al. New Insights into Bacterial Chemoreceptor Array Structure and Assembly from Electron Cryotomography. Biochemistry 2014, 53, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Briegel, A.; Ortega, D.R.; Tocheva, E.I.; Wuichet, K.; Li, Z.; Chen, S.; Müller, A.; Iancu, C.V.; Murphy, G.E.; Dobro, M.J.; et al. Universal Architecture of Bacterial Chemoreceptor Arrays. Proc. Natl. Acad. Sci. USA 2009, 106, 17181–17186. [Google Scholar] [CrossRef] [Green Version]
- Wuichet, K.; Zhulin, I.B. Origins and Diversification of a Complex Signal Transduction System in Prokaryotes. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuichet, K.; Zhulin, I.B. Molecular Evolution of Sensory Domains in Cyanobacterial Chemoreceptors. Trends Microbiol. 2003, 11, 200–203. [Google Scholar] [CrossRef]
- Belousoff, M.J.; Davidovich, C.; Zimmerman, E.; Caspi, Y.; Wekselman, I.; Rozenszajn, L.; Shapira, T.; Sade-Falk, O.; Taha, L.; Bashan, A.; et al. Ancient Machinery Embedded in the Contemporary Ribosome. Biochem. Soc. Trans. 2010, 38, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Fox, G.E. Origin and Evolution of the Ribosome. Cold Spring Harb. Perspect. Biol. 2010, 2, a003483. [Google Scholar] [CrossRef] [Green Version]
- Opron, K.; Burton, Z.F. Ribosome Structure, Function, and Early Evolution. Int. J. Mol. Sci. 2018, 20, 40. [Google Scholar] [CrossRef] [Green Version]
- Melnikov, S.; Ben-Shem, A.; Garreau de Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One Core, Two Shells: Bacterial and Eukaryotic Ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Lecompte, O.; Ripp, R.; Thierry, J.-C.; Moras, D.; Poch, O. Comparative Analysis of Ribosomal Proteins in Complete Genomes: An Example of Reductive Evolution at the Domain Scale. Nucleic Acids Res. 2002, 30, 5382–5390. [Google Scholar] [CrossRef]
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.V.; et al. History of the Ribosome and the Origin of Translation. Proc. Natl. Acad. Sci. USA 2015, 112, 15396–15401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosjean, H.; Westhof, E. An Integrated, Structure- and Energy-Based View of the Genetic Code. Nucleic Acids Res. 2016, 44, 8020–8040. [Google Scholar] [CrossRef] [Green Version]
- Root-Bernstein, M.; Root-Bernstein, R. The Ribosome as a Missing Link in the Evolution of Life. J. Theor. Biol. 2015, 367, 130–158. [Google Scholar] [CrossRef] [Green Version]
- Root-Bernstein, R.; Root-Bernstein, M. The Ribosome as a Missing Link in Prebiotic Evolution II: Ribosomes Encode Ribosomal Proteins That Bind to Common Regions of Their Own MRNAs and RRNAs. J. Biol. 2016, 397, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Root-Bernstein, R.; Root-Bernstein, M. The Ribosome as a Missing Link in Prebiotic Evolution III: Over-Representation of TRNA- and RRNA-Like Sequences and Plieofunctionality of Ribosome-Related Molecules Argues for the Evolution of Primitive Genomes from Ribosomal RNA Modules. Int. J. Mol. Sci. 2019, 20, 140. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, R.M.; Ramakrishnan, V. Structural Basis of the Translational Elongation Cycle. Annu. Rev. Biochem. 2013, 82, 203–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The Complete Atomic Structure of the Large Ribosomal Subunit at 2.4 A Resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.N.; Doudna Cate, J.H. The Structure and Function of the Eukaryotic Ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef] [PubMed]
- Rhodin, M.H.J.; Dinman, J.D. An Extensive Network of Information Flow through the B1b/c Intersubunit Bridge of the Yeast Ribosome. PLoS ONE 2011, 6, e20048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meskauskas, A.; Dinman, J.D. A Molecular Clamp Ensures Allosteric Coordination of Peptidyltransfer and Ligand Binding to the Ribosomal A-Site. Nucleic Acids Res. 2010, 38, 7800–7813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulima, S.O.; Gülay, S.P.; Anjos, M.; Patchett, S.; Meskauskas, A.; Johnson, A.W.; Dinman, J.D. Eukaryotic RpL10 Drives Ribosomal Rotation. Nucleic Acids Res. 2014, 42, 2049–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulay, S.P.; Bista, S.; Varshney, A.; Kirmizialtin, S.; Sanbonmatsu, K.Y.; Dinman, J.D. Tracking Fluctuation Hotspots on the Yeast Ribosome through the Elongation Cycle. Nucleic Acids Res. 2017, 45, 4958–4971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakauskaite, R.; Dinman, J.D. RRNA Mutants in the Yeast Peptidyltransferase Center Reveal Allosteric Information Networks and Mechanisms of Drug Resistance. Nucleic Acids Res. 2008, 36, 1497–1507. [Google Scholar] [CrossRef] [Green Version]
- Bowen, A.M.; Musalgaonkar, S.; Moomau, C.A.; Gulay, S.P.; Mirvis, M.; Dinman, J.D. Ribosomal Protein US19 Mutants Reveal Its Role in Coordinating Ribosome Structure and Function. Transl. 2015, 3, e1117703. [Google Scholar] [CrossRef] [PubMed]
- Kisly, I.; Gulay, S.P.; Mäeorg, U.; Dinman, J.D.; Remme, J.; Tamm, T. The Functional Role of EL19 and EB12 Intersubunit Bridge in the Eukaryotic Ribosome. J. Mol. Biol. 2016, 428, 2203–2216. [Google Scholar] [CrossRef] [Green Version]
- Meskauskas, A.; Russ, J.R.; Dinman, J.D. Structure/Function Analysis of Yeast Ribosomal Protein L2. Nucleic Acids Res. 2008, 36, 1826–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaher, H.S.; Green, R. Fidelity at the Molecular Level: Lessons from Protein Synthesis. Cell 2009, 136, 746–762. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N.; Arenz, S.; Beckmann, R. Translation Regulation via Nascent Polypeptide-Mediated Ribosome Stalling. Curr. Opin. Struct. Biol. 2016, 37, 123–133. [Google Scholar] [CrossRef]
- Pechmann, S.; Willmund, F.; Frydman, J. The Ribosome as a Hub for Protein Quality Control. Mol. Cell 2013, 49, 411–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juszkiewicz, S.; Chandrasekaran, V.; Lin, Z.; Kraatz, S.; Ramakrishnan, V.; Hegde, R.S. ZNF598 Is a Quality Control Sensor of Collided Ribosomes. Mol. Cell 2018, 72, 469–481.e7. [Google Scholar] [CrossRef] [Green Version]
- Korostelev, A.; Ermolenko, D.N.; Noller, H.F. Structural Dynamics of the Ribosome. Curr. Opin. Chem. Biol. 2008, 12, 674–683. [Google Scholar] [CrossRef] [Green Version]
- Paci, M.; Fox, G.E. Major Centers of Motion in the Large Ribosomal RNAs. Nucleic Acids Res. 2015, 43, 4640–4649. [Google Scholar] [CrossRef] [Green Version]
- Paci, M.; Fox, G.E. Centers of Motion Associated with EF-Tu Binding to the Ribosome. RNA Biol. 2016, 13, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuth, N.R.; Barna, M. The Discovery of Ribosome Heterogeneity and Its Implications for Gene Regulation and Organismal Life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinman, J.D. Pathways to Specialized Ribosomes: The Brussels Lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef] [Green Version]
- Forterre, P. The Universal Tree of Life: An Update. Front. Microbiol. 2015, 6, 717. [Google Scholar] [CrossRef] [Green Version]
- Moroz, L.L. On the Independent Origins of Complex Brains and Neurons. Brain Behav. Evol 2009, 74, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Moroz, L.L.; Romanova, D.Y.; Kohn, A.B. Neural versus Alternative Integrative Systems: Molecular Insights into Origins of Neurotransmitters. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20190762. [Google Scholar] [CrossRef]
- Craddock, T.J.A.; Tuszynski, J.A.; Hameroff, S. Cytoskeletal Signaling: Is Memory Encoded in Microtubule Lattices by CaMKII Phosphorylation? PLOS Comput. Biol. 2012, 8, e1002421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, G.; Vértes, P.E.; Towlson, E.K.; Chew, Y.L.; Walker, D.S.; Schafer, W.R.; Barabási, A.-L. Network Control Principles Predict Neuron Function in the Caenorhabditis Elegans Connectome. Nature 2017, 550, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Perkins, T.J.; Swain, P.S. Strategies for Cellular Decision-Making. Mol. Syst. Biol. 2009, 5, 326. [Google Scholar] [CrossRef]
- Tagkopoulos, I.; Liu, Y.-C.; Tavazoie, S. Predictive Behavior within Microbial Genetic Networks. Science 2008, 320, 1313–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seoane, L.F.; Solé, R.V. Information Theory, Predictability and the Emergence of Complex Life. R. Soc. Open Sci. 2018, 5, 172221. [Google Scholar] [CrossRef] [Green Version]
- McGregor, S.; Vasas, V.; Husbands, P.; Fernando, C. Evolution of Associative Learning in Chemical Networks. PLoS Comput. Biol. 2012, 8, e1002739. [Google Scholar] [CrossRef]
- Fernando, C.T.; Liekens, A.M.L.; Bingle, L.E.H.; Beck, C.; Lenser, T.; Stekel, D.J.; Rowe, J.E. Molecular Circuits for Associative Learning in Single-Celled Organisms. J. R. Soc. Interface 2009, 6, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.S.; Mumford, D. Hierarchical Bayesian Inference in the Visual Cortex. J. Opt. Soc. Am. A JOSAA 2003, 20, 1434–1448. [Google Scholar] [CrossRef]
- Da Costa, L.; Parr, T.; Sajid, N.; Veselic, S.; Neacsu, V.; Friston, K. Active Inference on Discrete State-Spaces: A Synthesis. J. Math. Psychol. 2020, 99, 102447. [Google Scholar] [CrossRef]
- Friston, K. A Theory of Cortical Responses. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 815–836. [Google Scholar] [CrossRef]
- George, D.; Hawkins, J. Towards a Mathematical Theory of Cortical Micro-Circuits. PLOS Comput. Biol. 2009, 5, e1000532. [Google Scholar] [CrossRef] [PubMed]
- Diaconescu, A.O.; Litvak, V.; Mathys, C.; Kasper, L.; Friston, K.J.; Stephan, K.E. A Computational Hierarchy in Human Cortex. arXiv 2017, arXiv:1709.02323. [Google Scholar]
- Houillon, A.; Bessière, P.; Droulez, J. The Probabilistic Cell: Implementation of a Probabilistic Inference by the Biochemical Mechanisms of Phototransduction. Acta Biotheor. 2010, 58, 103–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, C.R.; Garnier, S.; Beekman, M.; Latty, T. Information Integration and Multiattribute Decision Making in Non-Neuronal Organisms. Anim. Behav. 2015, 100, 44–50. [Google Scholar] [CrossRef]
- Reid, C.R.; MacDonald, H.; Mann, R.P.; Marshall, J.A.R.; Latty, T.; Garnier, S. Decision-Making without a Brain: How an Amoeboid Organism Solves the Two-Armed Bandit. J. R. Soc. Interface 2016, 13, 20160030. [Google Scholar] [CrossRef]
- Pawson, T. Dynamic Control of Signaling by Modular Adaptor Proteins. Curr. Opin. Cell Biol. 2007, 19, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Monod, J. Le Hasard et la Nécessité. Essai sur la philosophie naturelle de la biologie moderne; Média Diffusion: Roanne, France, 2014; ISBN 978-2-02-122439-9. [Google Scholar]
- Nussinov, R.; Tsai, C.-J.; Liu, J. Principles of Allosteric Interactions in Cell Signaling. J. Am. Chem. Soc. 2014, 136, 17692–17701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motlagh, H.N.; Wrabl, J.O.; Li, J.; Hilser, V.J. The Ensemble Nature of Allostery. Nature 2014, 508, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Nussinov, R. Amplification of Signaling via Cellular Allosteric Relay and Protein Disorder. Proc. Natl. Acad. Sci. USA 2009, 106, 6887–6888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Anslyn, E.V. Signal Amplification by Allosteric Catalysis. Angew. Chem. Int. Ed. Engl. 2006, 45, 1190–1196. [Google Scholar] [CrossRef]
- Hilser, V.J.; Wrabl, J.O.; Motlagh, H.N. Structural and Energetic Basis of Allostery. Annu. Rev. Biophys. 2012, 41, 585–609. [Google Scholar] [CrossRef] [Green Version]
- Hilser, V.J.; Thompson, E.B. Intrinsic Disorder as a Mechanism to Optimize Allosteric Coupling in Proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 8311–8315. [Google Scholar] [CrossRef] [Green Version]
- Gunasekaran, K.; Ma, B.; Nussinov, R. Is Allostery an Intrinsic Property of All Dynamic Proteins? Proteins 2004, 57, 433–443. [Google Scholar] [CrossRef]
- Bhat, A.S.; Dustin Schaeffer, R.; Kinch, L.; Medvedev, K.E.; Grishin, N.V. Recent Advances Suggest Increased Influence of Selective Pressure in Allostery. Curr. Opin. Struct. Biol. 2020, 62, 183–188. [Google Scholar] [CrossRef]
- Zhang, Y.; Doruker, P.; Kaynak, B.; Zhang, S.; Krieger, J.; Li, H.; Bahar, I. Intrinsic Dynamics Is Evolutionarily Optimized to Enable Allosteric Behavior. Curr. Opin. Struct. Biol. 2020, 62, 14–21. [Google Scholar] [CrossRef]
- Lockless, S.W.; Ranganathan, R. Evolutionarily Conserved Pathways of Energetic Connectivity in Protein Families. Science 1999, 286, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Süel, G.M.; Lockless, S.W.; Wall, M.A.; Ranganathan, R. Evolutionarily Conserved Networks of Residues Mediate Allosteric Communication in Proteins. Nat. Struct. Biol. 2003, 10, 59–69. [Google Scholar] [CrossRef]
- Goodey, N.M.; Benkovic, S.J. Allosteric Regulation and Catalysis Emerge via a Common Route. Nat. Chem. Biol. 2008, 4, 474–482. [Google Scholar] [CrossRef]
- Abrusán, G.; Marsh, J.A. Ligand-Binding-Site Structure Shapes Allosteric Signal Transduction and the Evolution of Allostery in Protein Complexes. Mol. Biol. Evol 2019, 36, 1711–1727. [Google Scholar] [CrossRef]
- Leander, M.; Yuan, Y.; Meger, A.; Cui, Q.; Raman, S. Functional Plasticity and Evolutionary Adaptation of Allosteric Regulation. Proc. Natl. Acad. Sci. USA 2020, 117, 25445–25454. [Google Scholar] [CrossRef] [PubMed]
- Timsit, Y.; Allemand, F.; Chiaruttini, C.; Springer, M. Coexistence of Two Protein Folding States in the Crystal Structure of Ribosomal Protein L20. EMBO Rep. 2006, 7, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Timsit, Y.; Acosta, Z.; Allemand, F.; Chiaruttini, C.; Springer, M. The Role of Disordered Ribosomal Protein Extensions in the Early Steps of Eubacterial 50 S Ribosomal Subunit Assembly. Int. J. Mol. Sci. 2009, 10, 817–834. [Google Scholar] [CrossRef] [Green Version]
- Babu, Y.S.; Bugg, C.E.; Cook, W.J. Structure of Calmodulin Refined at 2.2 A Resolution. J. Mol. Biol. 1988, 204, 191–204. [Google Scholar] [CrossRef]
- Simonson, T.; Aleksandrov, A.; Satpati, P. Electrostatic Free Energies in Translational GTPases: Classic Allostery and the Rest. Biochim. Biophys. Acta 2015, 1850, 1006–1016. [Google Scholar] [CrossRef]
- Sato, T.; Ohnuki, J.; Takano, M. Dielectric Allostery of Protein: Response of Myosin to ATP Binding. J. Phys. Chem. B 2016, 120, 13047–13055. [Google Scholar] [CrossRef]
- Sato, T.; Ohnuki, J.; Takano, M. Long-Range Coupling between ATP-Binding and Lever-Arm Regions in Myosin via Dielectric Allostery. J. Chem. Phys. 2017, 147, 215101. [Google Scholar] [CrossRef]
- Preto, J.; Pettini, M.; Tuszynski, J.A. Possible Role of Electrodynamic Interactions in Long-Distance Biomolecular Recognition. Phys. Rev. E Stat. Nonlin Soft Matter Phys. 2015, 91, 052710. [Google Scholar] [CrossRef] [Green Version]
- Olmi, S.; Gori, M.; Donato, I.; Pettini, M. Collective Behavior of Oscillating Electric Dipoles. Sci. Rep. 2018, 8, 15748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, W.; Lang, E.J.; Bai, Y.; Fan, Y.; Parker, E.J. Diverse Allosteric Componentry and Mechanisms Control Entry into Aromatic Metabolite Biosynthesis. Curr. Opin. Struct. Biol. 2020, 65, 159–167. [Google Scholar] [CrossRef]
- Webby, C.J.; Jiao, W.; Hutton, R.D.; Blackmore, N.J.; Baker, H.M.; Baker, E.N.; Jameson, G.B.; Parker, E.J. Synergistic Allostery, a Sophisticated Regulatory Network for the Control of Aromatic Amino Acid Biosynthesis in Mycobacterium Tuberculosis. J. Biol. Chem. 2010, 285, 30567–30576. [Google Scholar] [CrossRef] [Green Version]
- Jiao, W.; Hutton, R.D.; Cross, P.J.; Jameson, G.B.; Parker, E.J. Dynamic Cross-Talk among Remote Binding Sites: The Molecular Basis for Unusual Synergistic Allostery. J. Mol. Biol. 2012, 415, 716–726. [Google Scholar] [CrossRef]
- Blackmore, N.J.; Reichau, S.; Jiao, W.; Hutton, R.D.; Baker, E.N.; Jameson, G.B.; Parker, E.J. Three Sites and You Are out: Ternary Synergistic Allostery Controls Aromatic Amino Acid Biosynthesis in Mycobacterium Tuberculosis. J. Mol. Biol. 2013, 425, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Fan, Y.; Blackmore, N.J.; Parker, E.J. A Single Amino Acid Substitution Uncouples Catalysis and Allostery in an Essential Biosynthetic Enzyme in Mycobacterium Tuberculosis. J. Biol. Chem. 2020, 295, 6252–6262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Ren, Z.; Zeng, X.; Bandara, S.; Yang, X. Structural Basis of Molecular Logic OR in a Dual-Sensor Histidine Kinase. Proc. Natl Acad Sci. USA 2019, 116, 19973–19982. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Lummis, S.C.R. The Roles of Aromatic Residues in the Glycine Receptor Transmembrane Domain. BMC Neurosci. 2018, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Malvankar, N.S.; Vargas, M.; Nevin, K.; Tremblay, P.-L.; Evans-Lutterodt, K.; Nykypanchuk, D.; Martz, E.; Tuominen, M.T.; Lovley, D.R. Structural Basis for Metallic-Like Conductivity in Microbial Nanowires. mBio 2015, 6, e00084-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boon, E.M.; Barton, J.K. Charge Transport in DNA. Curr. Opin. Struct. Biol. 2002, 12, 320–329. [Google Scholar] [CrossRef]
- Rudrauf, D.; Lutz, A.; Cosmelli, D.; Lachaux, J.-P.; Le Van Quyen, M. From Autopoiesis to Neurophenomenology: Francisco Varela’s Exploration of the Biophysics of Being. Biol. Res. 2003, 36, 27–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, F. Autopoiesis and a Biology of Intentionality. In Proceedings of the Workshop “Autopoiesis and Perception”, Dublin Ireland, 25–26 August 1992; pp. 4–14. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timsit, Y.; Grégoire, S.-P. Towards the Idea of Molecular Brains. Int. J. Mol. Sci. 2021, 22, 11868. https://doi.org/10.3390/ijms222111868
Timsit Y, Grégoire S-P. Towards the Idea of Molecular Brains. International Journal of Molecular Sciences. 2021; 22(21):11868. https://doi.org/10.3390/ijms222111868
Chicago/Turabian StyleTimsit, Youri, and Sergeant-Perthuis Grégoire. 2021. "Towards the Idea of Molecular Brains" International Journal of Molecular Sciences 22, no. 21: 11868. https://doi.org/10.3390/ijms222111868
APA StyleTimsit, Y., & Grégoire, S. -P. (2021). Towards the Idea of Molecular Brains. International Journal of Molecular Sciences, 22(21), 11868. https://doi.org/10.3390/ijms222111868