
Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality


Abstract
:1. Introduction
2. Results
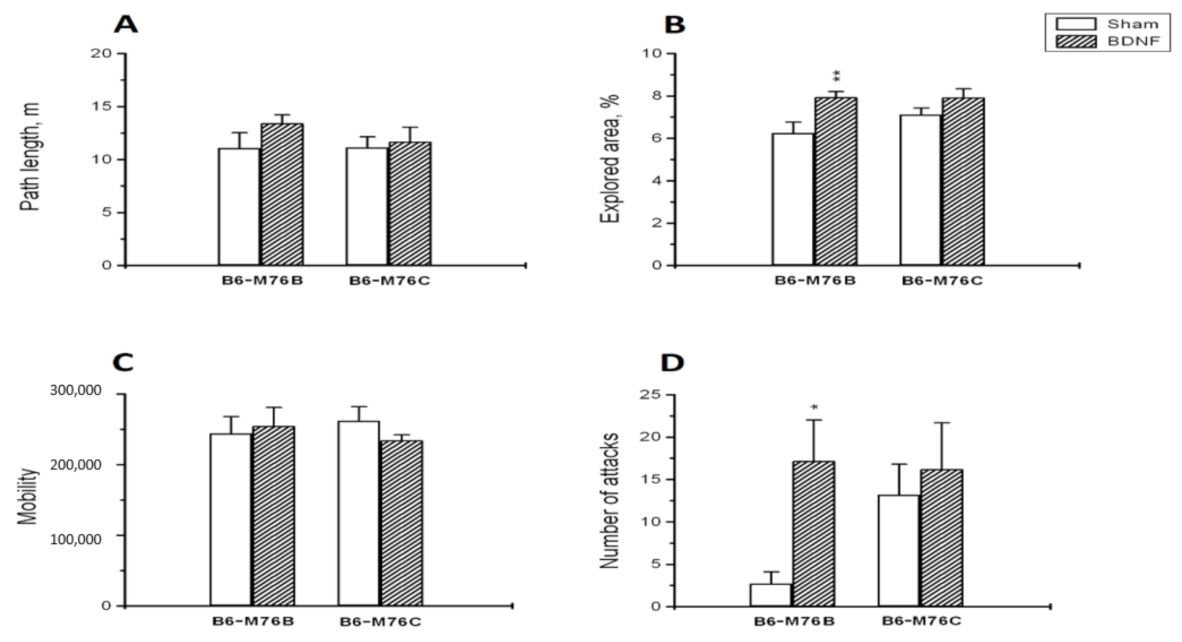
2.1. Behavioral Studies
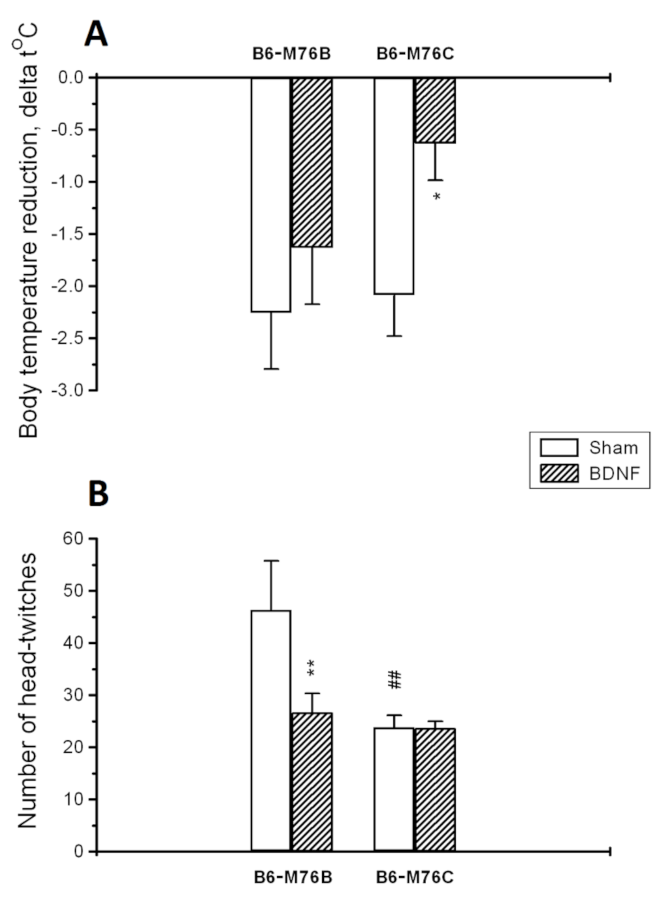
2.2. 5-HT1A and 5-HT2A Receptors Functional Responses
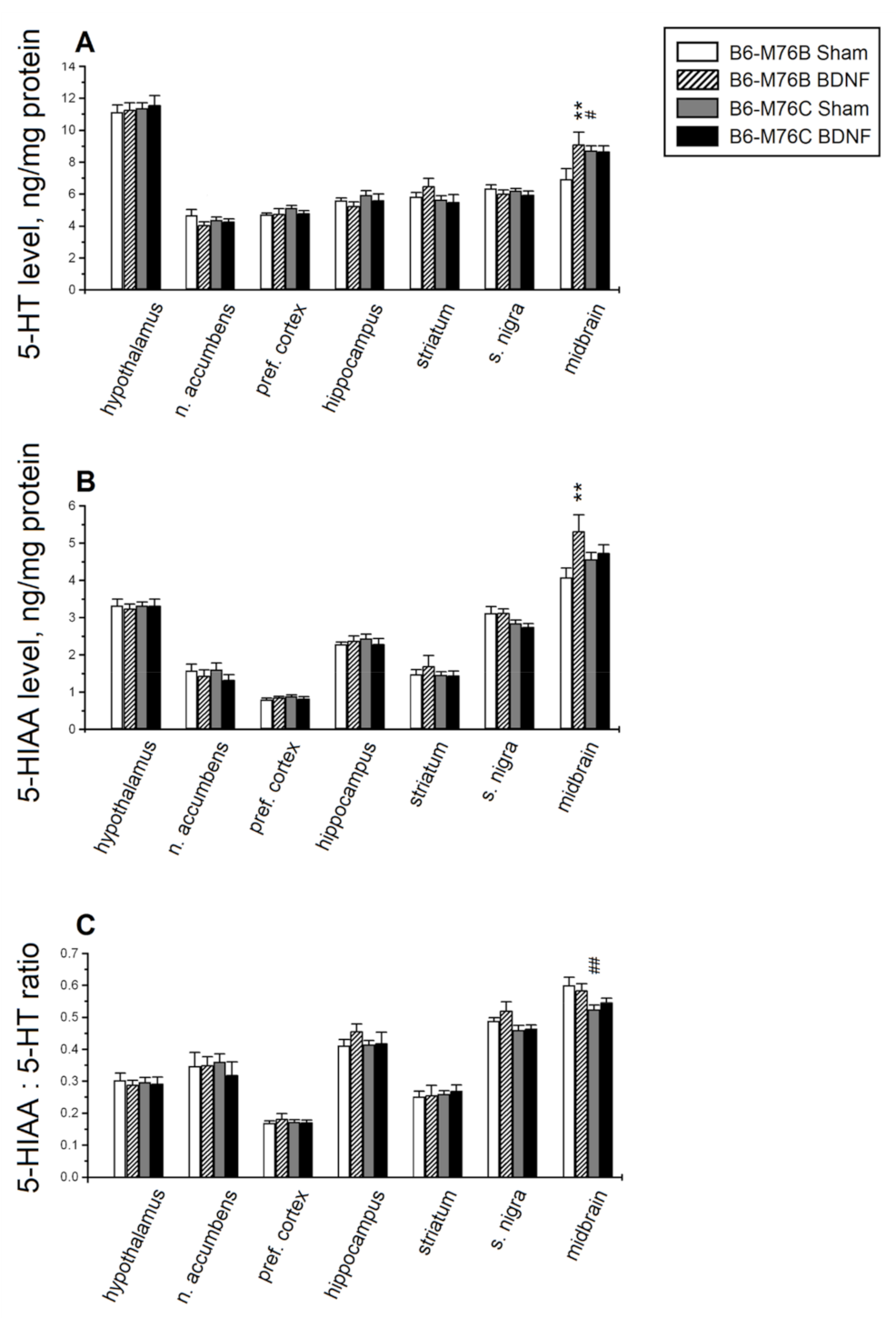
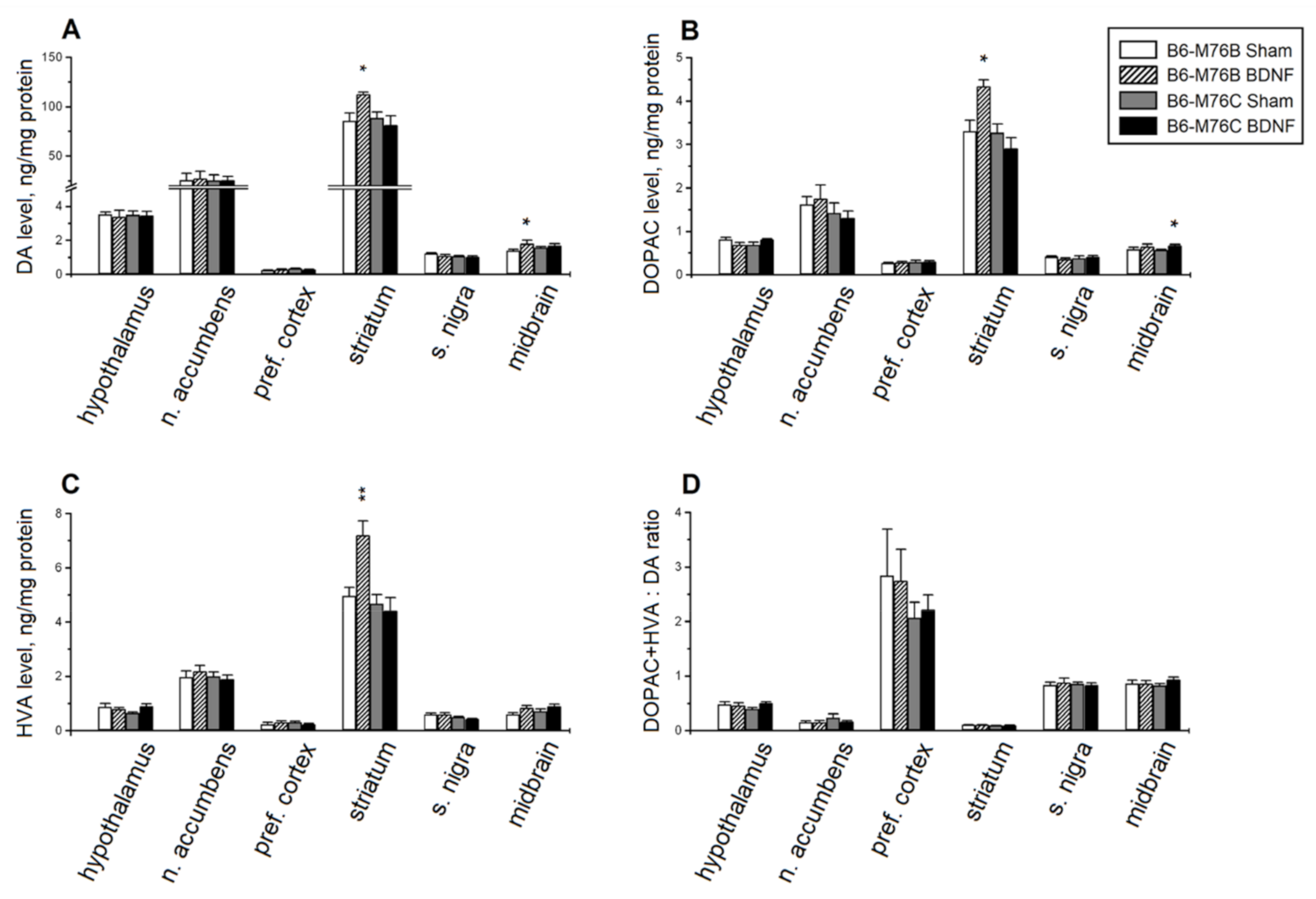
2.3. HPLC Analysis
3. Discussion
4. Materials and Methods
4.1. Breeding of Lines
4.2. Animals
4.3. Drugs
4.4. Behavioral Assay
4.5. Open-Field Test
4.6. Forced Swim Test
4.7. Intermale Aggression Test
4.8. 5-HT1A Receptor Functional Response
4.9. 5-HT2A Receptor Functional Response
4.10. High-Performance Liquid Chromatography (HPLC)
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brunoni, A.R.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef]
- Hellweg, R.; Ziegenhorn, A.; Heuser, I.; Deuschle, M. Serum Concentrations of Nerve Growth Factor and Brain-Derived Neurotrophic Factor in Depressed Patients before and after Antidepressant Treatment. Pharmacopsychiatry 2008, 41, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Tokumura, M.; Abe, K. Effects of rolipram, a phosphodiesterase 4 inhibitor, in combination with imipramine on depressive behavior, CRE-binding activity and BDNF level in learned helplessness rats. Eur. J. Pharmacol. 2004, 498, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Kim, Y.-K. Plasma Brain-Derived Neurotrophic Factor as a Peripheral Marker for the Action Mechanism of Antidepressants. Neuropsychobiology 2008, 57, 194–199. [Google Scholar] [CrossRef]
- Rogóz, Z.; Legutko, B. Combined treatment with imipramine and metyrapone induces hippocampal and cortical brain-derived neurotrophic factor gene expression in rats. Pharmacol. Rep. 2005, 57, 840–844. [Google Scholar] [PubMed]
- Sen, S.; Duman, R.; Sanacora, G. Serum Brain-Derived Neurotrophic Factor, Depression, and Antidepressant Medications: Meta-Analyses and Implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Duman, R.S. Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.D.; Banasr, M.; Duman, R.S. Future antidepressant targets: Neurotrophic factors and related signaling cascades. Drug Discov. Today Ther. Strat. 2008, 5, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Siuciak, J.A.; Boylan, C.; Fritsche, M.; Altar, C.A.; Lindsay, R.M. BDNF increases monoaminergic activity in rat brain following intracerebroventricular or intraparenchymal administration. Brain Res. 1996, 710, 11–20. [Google Scholar] [CrossRef]
- Klein, A.; Santini, M.; Aznar, S.; Knudsen, G.M.; Rios, M. Changes in 5-HT2A-mediated behavior and 5-HT2A- and 5-HT1A receptor binding and expression in conditional brain-derived neurotrophic factor knock-out mice. Neuroscience 2010, 169, 1007–1016. [Google Scholar] [CrossRef] [Green Version]
- Balu, D.; Hoshaw, B.A.; Malberg, J.E.; Rosenzweig-Lipson, S.; Schechter, L.E.; Lucki, I. Differential regulation of central BDNF protein levels by antidepressant and non-antidepressant drug treatments. Brain Res. 2008, 1211, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrén, E. Neurotrophic effects of antidepressant drugs. Curr. Opin. Pharmacol. 2004, 4, 58–64. [Google Scholar] [CrossRef]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef] [PubMed]
- Hyman, C.; Hofer, M.; Barde, Y.; Juhasz, M.; Yancopoulos, G.D.; Squinto, S.P.; Lindsay, R.M. BDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigra. Nat. Cell Biol. 1991, 350, 230–232. [Google Scholar] [CrossRef]
- Spina, M.B.; Squinto, S.P.; Miller, J.; Lindsay, R.M.; Hyman, C. Brain-Derived Neurotrophic Factor Protects Dopamine Neurons Against 6-Hydroxydopamine and N-Methyl-4-Phenylpyridinium Ion Toxicity: Involvement of the Glutathione System. J. Neurochem. 1992, 59, 99–106. [Google Scholar] [CrossRef]
- Goggi, J.; Pullar, I.A.; Carney, S.L.; Bradford, H.F. Modulation of neurotransmitter release induced by brain-derived neurotrophic factor in rat brain striatal slices in vitro. Brain Res. 2002, 941, 34–42. [Google Scholar] [CrossRef]
- Adachi, N.; Yoshimura, A.; Chiba, S.; Ogawa, S.; Kunugi, H. Rotigotine, a dopamine receptor agonist, increased BDNF protein levels in the rat cortex and hippocampus. Neurosci. Lett. 2018, 662, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Dellu-Hagedorn, F.; Fitoussi, A.; De Deurwaerdère, P. Correlative analysis of dopaminergic and serotonergic metabolism across the brain to study monoaminergic function and interaction. J. Neurosci. Methods 2017, 280, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Di Matteo, V.; Di Giovanni, G. Serotonin–dopamine interaction: An overview. Prog. Brain Res. 2008, 172, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Naumenko, V.S.; Kondaurova, E.M.; Bazovkina, D.V.; Tsybko, A.S.; Il’chibaeva, T.V.; Popova, N.K. On the role of 5-HT(1A) receptor gene in behavioral effect of brain-derived neurotrophic factor. J. Neurosci. Res. 2014, 92, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.; Kondaurova, E.; Bazovkina, D.; Tsybko, A.; Tikhonova, M.; Kulikov, A.; Popova, N. Effect of brain-derived neurotrophic factor on behavior and key members of the brain serotonin system in genetically predisposed to behavioral disorders mouse strains. Neuroscience 2012, 214, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, E.A.; Bazovkina, D.V.; Akulov, A.E.; Tsybko, A.S.; Fursenko, D.V.; Kulikov, A.V.; Naumenko, V.S.; Ponimaskin, E.; Kondaurova, E.M. Alterations in pharmacological and behavioural responses in recombinant mouse line with an increased predisposition to catalepsy: Role of the 5-HT1A receptor. Br. J. Pharmacol. 2016, 173, 2147–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondaurova, E.M.; Rodnyy, A.Y.; Ilchibaeva, T.V.; Tsybko, A.S.; Eremin, D.V.; Antonov, Y.V.; Popova, N.K.; Naumenko, V.S. Genetic Background Underlying 5-HT1A Receptor Functioning Affects the Response to Fluoxetine. Int. J. Mol. Sci. 2020, 21, 8784. [Google Scholar] [CrossRef]
- De Boer, S.F.; Koolhaas, J.M. 5-HT1A and 5-HT1B receptor agonists and aggression: A pharmacological challenge of the serotonin deficiency hypothesis. Eur. J. Pharmacol. 2005, 526, 125–139. [Google Scholar] [CrossRef]
- Albert, P.; Lemonde, S. 5-HT1A Receptors, Gene Repression, and Depression: Guilt by Association. Neuroscientist 2004, 10, 575–593. [Google Scholar] [CrossRef] [PubMed]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.; Beck, S.G.; et al. 5-HT1A Autoreceptor Levels Determine Vulnerability to Stress and Response to Antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Boldrini, M.; Underwood, M.; Mann, J.J.; Arango, V. Serotonin-1A autoreceptor binding in the dorsal raphe nucleus of depressed suicides. J. Psychiatr. Res. 2008, 42, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Stockmeier, C.A.; Shapiro, L.A.; Dilley, G.E.; Kolli, T.N.; Friedman, L.; Rajkowska, G. Increase in Serotonin-1A Autoreceptors in the Midbrain of Suicide Victims with Major Depression—Postmortem Evidence for Decreased Serotonin Activity. J. Neurosci. 1998, 18, 7394–7401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blier, P.; Ward, N.M. Is there a role for 5-HT1A agonists in the treatment of depression? Biol. Psychiatry 2003, 53, 193–203. [Google Scholar] [CrossRef]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Govindarajan, A.; Rao, B.S.; Nair, D.; Trinh, M.; Mawjee, N.; Tonegawa, S.; Chattarji, S. Transgenic brain-derived neurotrophic factor expression causes both anxiogenic and antidepressant effects. Proc. Natl. Acad. Sci. USA 2006, 103, 13208–13213. [Google Scholar] [CrossRef] [Green Version]
- Rios, M.; Fan, G.; Fekete, C.; Kelly, J.; Bates, B.; Kuehn, R.; Lechan, R.M.; Jaenisch, R. Conditional deletion of brain-derived neurotrophic factor in the postnatal brain leads to obesity and hyperactivity. Mol. Endocrinol. 2001, 15, 1748–1757. [Google Scholar] [CrossRef]
- Shirayama, Y.; Chen, A.C.; Nakagawa, S.; Russell, D.S.; Duman, R.S. Brain-derived neurotrophic factor produces anti-depressant effects in behavioral models of depression. J. Neurosci. 2002, 22, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.; Unger, T.; Byrnes, J.; Rios, M. Examination of behavioral deficits triggered by targeting BDNF in fetal or postnatal brains of mice. Neuroscience 2006, 142, 49–58. [Google Scholar] [CrossRef]
- Chourbaji, S.; Hellweg, R.; Brandis, D.; Zörner, B.; Zacher, C.; Lang, U.E.; Henn, F.A.; Hörtnagl, H.; Gass, P. Mice with reduced brain-derived neurotrophic factor expression show decreased choline acetyltransferase activity, but regular brain monoamine levels and unaltered emotional behavior. Brain Res. Mol. Brain Res. 2004, 121, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ito, W.; Chehab, M.; Thakur, S.; Li, J.; Morozov, A. BDNF-restricted knockout mice as an animal model for aggression. Genes Brain Behav. 2011, 10, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Lyons, W.E.; Mamounas, L.A.; Ricaurte, G.A.; Coppola, V.; Reid, S.W.; Bora, S.H.; Wihler, C.; Koliatsos, V.E.; Tessarollo, L. Brain-derived neurotrophic factor-deficient mice develop aggressiveness and hyperphagia in conjunction with brain serotonergic abnormalities. Proc. Natl. Acad. Sci. USA 1999, 96, 15239–15244. [Google Scholar] [CrossRef] [Green Version]
- Deltheil, T.; Guiard, B.; Cerdan, J.; David, D.; Tanaka, K.; Repérant, C.; Guilloux, J.-P.; Coudoré, F.; Hen, R.; Gardier, A. Behavioral and serotonergic consequences of decreasing or increasing hippocampus brain-derived neurotrophic factor protein levels in mice. Neuropharmacology 2008, 55, 1006–1014. [Google Scholar] [CrossRef]
- Papaleo, F.; Silverman, J.L.; Aney, J.; Tian, Q.; Barkan, C.L.; Chadman, K.K.; Crawley, J.N. Working memory deficits, increased anxiety-like traits, and seizure susceptibility in BDNF overexpressing mice. Learn. Mem. 2011, 18, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A. Hippocampal BDNF overexpression or microR124a silencing reduces anxiety- and autism-like behaviors in rats. Behav. Brain Res. 2017, 326, 281–290. [Google Scholar] [CrossRef]
- Bell, R.; Hobson, H. 5-HT1A receptor influences on rodent social and agonistic behavior: A review and empirical study. Neurosci. Biobehav. Rev. 1994, 18, 325–338. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kozhemyakina, R.V.; Plyusnina, I.F.; Kulikov, A.V.; Popova, N.K. Serotonin 5-HT1A receptor in infancy-onset aggression: Comparison with genetically defined aggression in adult rats. Behav. Brain Res. 2013, 243, 97–101. [Google Scholar] [CrossRef]
- Olivier, B.; Mos, J.; Van Oorschot, R.; Hen, R. Serotonin Receptors and Animal Models of Aggressive Behavior. Pharmacopsychiatry 1995, 28 (Suppl. 2), 80–90. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S.; Plyusnina, I.Z.; Kulikov, A.V. Reduction in 5-HT1A receptor density, 5-HT1A mRNA expression, and functional correlates for 5-HT1A receptors in genetically defined aggressive rats. J. Neurosci. Res. 2005, 80, 286–292. [Google Scholar] [CrossRef]
- Popova, N.; Naumenko, V.; Cybko, A.; Bazovkina, D. Receptor-genes cross-talk: Effect of chronic 5-HT1A agonist 8-hydroxy-2-(di-n-propylamino) tetralin treatment on the expression of key genes in brain serotonin system and on behavior. Neuroscience 2010, 169, 229–235. [Google Scholar] [CrossRef]
- Blier, P.; Seletti, B.; Gilbert, F.; Young, S.N.; Benkelfat, C. Serotonin 1A Receptor Activation and Hypothermia in Humans Lack of Evidence for a Presynaptic Mediation. Neuropsychopharmacology 2002, 27, 301–308. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Bazovkina, D.V.; Kondaurova, E.M.; Zubkov, E.A.; Kulikov, A.V. The role of 5-HT2A receptor and 5-HT2A/5-HT1A receptor interaction in the suppression of catalepsy. Genes Brain Behav. 2010, 9, 519–524. [Google Scholar]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. 5-HT1A receptor as a key player in the brain 5-HT system. Rev. Neurosci. 2013, 24, 191–204. [Google Scholar] [CrossRef]
- Trajkovska, V.; Santini, M.; Marcussen, A.; Thomsen, M.; Hansen, H.; Mikkelsen, J.; Arneberg, L.; Kokaia, M.; Knudsen, G.M.; Aznar, S. BDNF downregulates 5-HT2A receptor protein levels in hippocampal cultures. Neurochem. Int. 2009, 55, 697–702. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Bazovkina, D.V.; Kondaurova, E.M. On the Functional Cross-Talk between Brain 5-HT1A and 5-HT2A Receptors. IP Pavlov J. High. Nerv. Act. 2015, 65, 240–247. [Google Scholar]
- Onn, S.-P.; West, A.R.; Grace, A.A. Dopamine-mediated regulation of striatal neuronal and network interactions. Trends Neurosci. 2000, 23, S48–S56. [Google Scholar] [CrossRef]
- Bailey, M.R.; Goldman, O.; Bello, E.P.; Chohan, M.O.; Jeong, N.; Winiger, V.; Chun, E.; Schipani, E.; Kalmbach, A.; Cheer, J.; et al. An Interaction between Serotonin Receptor Signaling and Dopamine Enhances Goal-Directed Vigor and Persistence in Mice. J. Neurosci. 2018, 38, 2149–2162. [Google Scholar] [CrossRef] [PubMed]
- Devroye, C.; Cathala, A.; Piazza, P.V.; Spampinato, U. The central serotonin2B receptor as a new pharmacological target for the treatment of dopamine-related neuropsychiatric disorders: Rationale and current status of research. Pharmacol. Ther. 2018, 181, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Flik, G.; Folgering, J.H.A.; Cremers, T.I.H.F.; Westerink, B.H.C.; Dremencov, E. Interaction Between Brain Histamine and Serotonin, Norepinephrine, and Dopamine Systems: In Vivo Microdialysis and Electrophysiology Study. J. Mol. Neurosci. 2015, 56, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Kondaurova, E.M.; Bazovkina, D.V.; Kulikov, A.V.; Popova, N.K. Selective breeding for catalepsy changes the distribution of microsatellite D13Mit76 alleles linked to the 5-HT1A serotonin receptor gene in mice. Genes Brain Behav. 2006, 5, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, A.V.; Bazovkina, D.V.; Kondaurova, E.M.; Popova, N.K. Genetic structure of hereditary catalepsy in mice. Genes Brain Behav. 2008, 7, 506–512. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Bazovkina, D.V.; Moisan, M.-P.; Mormede, P. The mapping of the gene of susceptibility to catalepsy in mice using polymorphic microsatellite markers. Dokl. Biol. Sci. 2003, 393, 531–534. [Google Scholar] [CrossRef]
- Slotnick, B.M.; Leonard, C.M. A Stereotaxic Atlas of the Albino Mouse Forebrain; US Department of Health, Education and Welfare: Rockville, MD, USA, 1975. [Google Scholar]
- Naumenko, V.S.; Kondaurova, E.; Popova, N.K. On the role of brain 5-HT7 receptor in the mechanism of hypothermia: Comparison with hypothermia mediated via 5-HT1A and 5-HT3 receptor. Neuropharmacology 2011, 61, 1360–1365. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Tikhonova, M.A.; Kulikov, V.A. Automated measurement of spatial preference in the open field test with transmitted lighting. J. Neurosci. Methods 2008, 170, 345–351. [Google Scholar] [CrossRef]
- Khotskin, N.V.; Plyusnina, A.V.; Kulikova, E.A.; Bazhenova, E.Y.; Fursenko, D.V.; Sorokin, I.E.; Kolotygin, I.; Mormede, P.; Terenina, E.E.; Shevelev, O.B.; et al. On association of the lethal yellow (A(Y)) mutation in the agouti gene with the alterations in mouse brain and behavior. Behav. Brain Res. 2019, 359, 446–456. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Morozova, M.V.; Kulikov, V.A.; Kirichuk, V.S.; Popova, N.K. Automated analysis of antidepressants’ effect in the forced swim test. J. Neurosci. Methods 2010, 191, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Osipova, D.V.; Kulikov, A.V.; Popova, N.K. C1473G polymorphism in mouse tph2 gene is linked to tryptophan hydroxylase-2 activity in the brain, intermale aggression, and depressive-like behavior in the forced swim test. J. Neurosci. Res. 2009, 87, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Kulikov, A.V. Genetic analysis of ‘spontaneous’ intermale aggression in mice. Aggr. Behav. 1986, 12, 425–431. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Rezvani, A.H.; Knapp, D.J.; Crews, F.T.; Janowsky, D.S. Further selection of rat lines differing in 5-HT-1A receptor sensitivity: Behavioral and functional correlates. Psychiatr. Genet. 1996, 6, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Green, A.R.; Heal, D.J. The effects of drugs on serotonin-mediated behavioural models. In Neuropharmacology of Serotonin; Green, A.R., Ed.; Oxford University: Oxford, UK, 1985; pp. 326–365. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Two-Way ANOVA | ||
|---|---|---|---|
| Effect of Genotype | Effect of Treatment | Effect of Interaction | |
| 5-HT | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| N. accumbens | F1.29 < 1 | F1.29 = 2.02, p > 0.05 | F1.29 = 1.11, p > 0.05 |
| Pref. cortex | F1.29 = 1.31, p > 0.05 | F1.29 < 1 | F1.29 < 1 |
| Hippocampus | F1.29 = 1.22, p > 0.05 | F1.29 < 1 | F1.29 < 1 |
| Striatum | F1.29 = 2.39, p > 0.05 | F1.29 < 1 | F1.29 = 1.25, p >0.05 |
| S. nigra | F1.29 < 1 | F1.29 = 2.03, p > 0.05 | F1.29 < 1 |
| Midbrain | F1.28 = 1.87, p > 0.05 | F1.28 = 4.59 * | F1.28 = 4.97 * |
| 5-HIAA | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| N. accumbens | F1.29 < 1 | F1.29 = 1.41, p > 0.05 | F1.29 < 1 |
| Pref. cortex | F1.29 < 1 | F1.29 < 1 | F1.29 = 1.23, p > 0.05 |
| Hippocampus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| Striatum | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| S. nigra | F1.29 = 7.17 * | F1.29 < 1 | F1.29 < 1 |
| Midbrain | F1.28 < 1 | F1.28 = 6.86 * | F1.28 = 3.82, p = 0.06 |
| 5-HIAA/5-HT | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| N. accumbens | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| Pref. cortex | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| Hippocampus | F1.29 < 1 | F1.29 = 1.04, p > 0.05 | F1.29 < 1 |
| Striatum | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| S. nigra | F1.29 = 6.63 * | F1.29 = 1.22, p > 0.05 | F1.29 < 1 |
| Midbrain | F1.28 = 10.31 ** | F1.28 < 1 | F1.29 = 1.18, p > 0.05 |
| Structure | Two-Way ANOVA | ||
|---|---|---|---|
| Effect of Genotype | Effect of Treatment | Effect of Interaction | |
| DA | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| N. accumbens | F1.28 < 1 | F1.28 < 1 | F1.28 < 1 |
| Pref. cortex | F1.29 = 1.63, p > 0.05 | F1.29 < 1 | F1.29 = 1.61, p > 0.05 |
| Striatum | F1.28 = 3.27, p = 0.08 | F1.28 = 1.61, p > 0.05 | F1.28 = 4.61 * |
| S. nigra | F1.29 = 3.49, p = 0.07 | F1.29 = 2.02, p > 0.05 | F1.29 < 1 |
| Midbrain | F1.28 < 1 | F1.28 = 4.72 * | F1.28 = 1.14, p > 0.05 |
| DOPAC | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 = 5.66 * |
| N. accumbens | F1.28 = 1.78, p > 0.05 | F1.28 < 1 | F1.28 < 1 |
| Pref. cortex | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| Striatum | F1.28 = 8.89 ** | F1.28 = 1.91, p > 0.05 | F1.28 = 8.05 ** |
| S. nigra | F1.29 < 1 | F1.29 < 1 | F1.29 = 1.22, p > 0.05 |
| Midbrain | F1.28 < 1 | F1.28 = 5.01 * | F1.28 < 1 |
| HVA | |||
| Hypothalamus | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| N. accumbens | F1.28 <1 | F1.28 < 1 | F1.28 < 1 |
| Pref. cortex | F1.29 < 1 | F1.29 < 1 | F1.29 = 2.05, p > 0.05 |
| Striatum | F1.28 = 11.94 ** | F1.28 = 4.93 * | F1.28 = 7.93 ** |
| S. nigra | F1.29 = 9.85 ** | F1.29 < 1 | F1.29 < 1 |
| Midbrain | F1.28 = 1.15, p > 0.05 | F1.28 = 6.52 * | F1.28 < 1 |
| DOPAC + HVA/DA | |||
| Hypothalamus | F1.29 < 1 | F1.29 = 1.32, p > 0.05 | F1.29 = 2.87, p > 0.05 |
| N. accumbens | F1.28 < 1 | F1.28 < 1 | F1.28 < 1 |
| Pref. cortex | F1.29 = 1.74, p > 0.05 | F1.29 < 1 | F1.29 < 1 |
| Striatum | F1.28 = 5.20 * | F1.28 < 1 | F1.28 < 1 |
| S. nigra | F1.29 < 1 | F1.29 < 1 | F1.29 < 1 |
| Midbrain | F1.28 < 1 | F1.28 = 1.22, p > 0.05 | F1.28 = 1.22, p > 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazovkina, D.; Naumenko, V.; Bazhenova, E.; Kondaurova, E. Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality. Int. J. Mol. Sci. 2021, 22, 11987. https://doi.org/10.3390/ijms222111987
Bazovkina D, Naumenko V, Bazhenova E, Kondaurova E. Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality. International Journal of Molecular Sciences. 2021; 22(21):11987. https://doi.org/10.3390/ijms222111987
Chicago/Turabian StyleBazovkina, Darya, Vladimir Naumenko, Ekaterina Bazhenova, and Elena Kondaurova. 2021. "Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality" International Journal of Molecular Sciences 22, no. 21: 11987. https://doi.org/10.3390/ijms222111987
APA StyleBazovkina, D., Naumenko, V., Bazhenova, E., & Kondaurova, E. (2021). Effect of Central Administration of Brain-Derived Neurotrophic Factor (BDNF) on Behavior and Brain Monoamine Metabolism in New Recombinant Mouse Lines Differing by 5-HT1A Receptor Functionality. International Journal of Molecular Sciences, 22(21), 11987. https://doi.org/10.3390/ijms222111987