
CAF Proteins Help SOT1 Regulate the Stability of Chloroplast ndhA Transcripts

 , and
, and 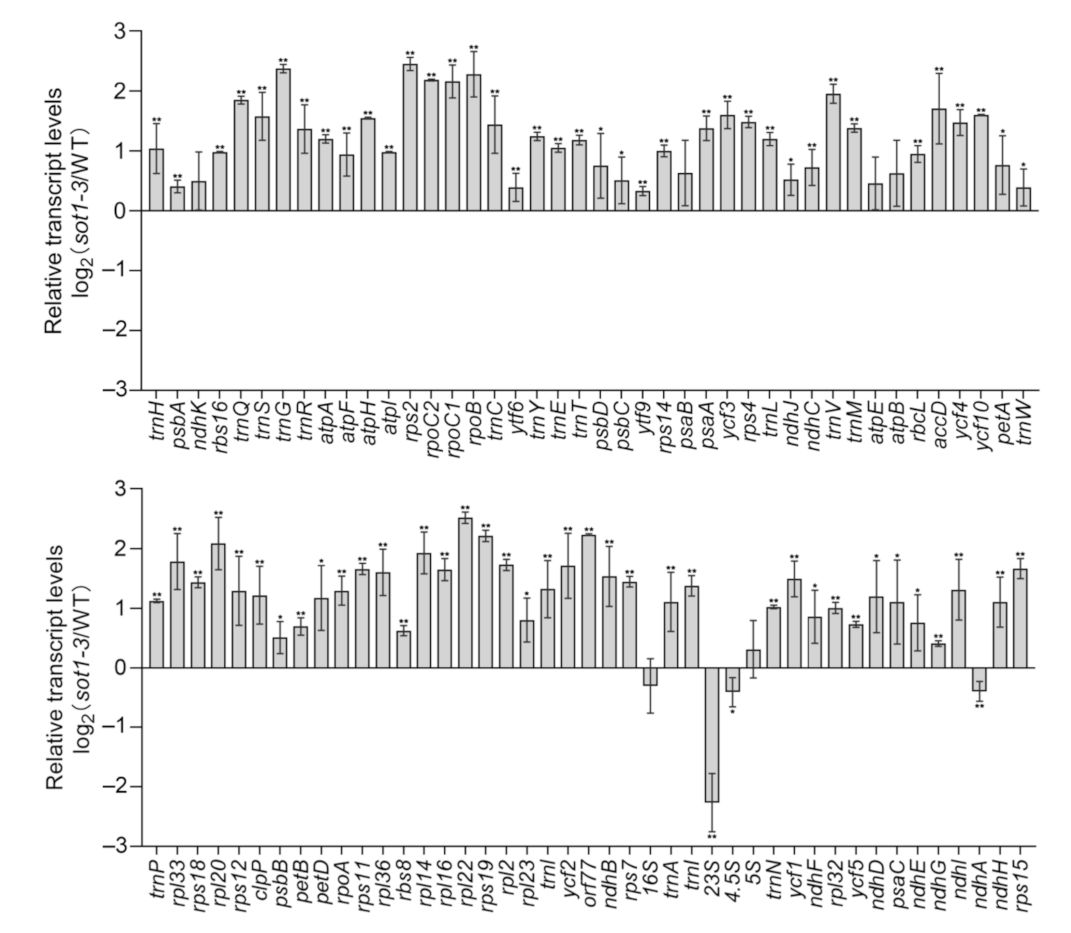{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SOT1 Post-Transcriptionally Regulates ndhA Expression
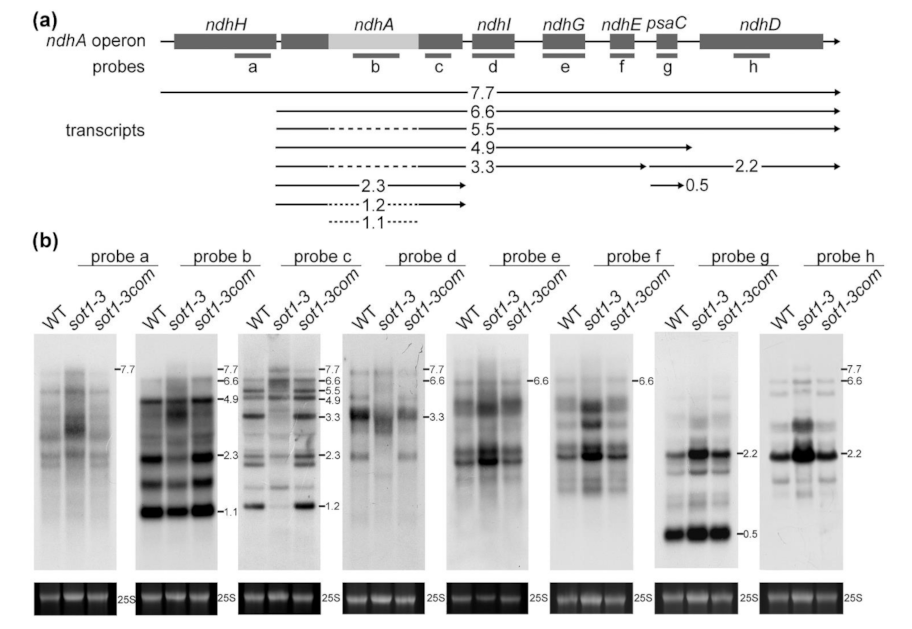
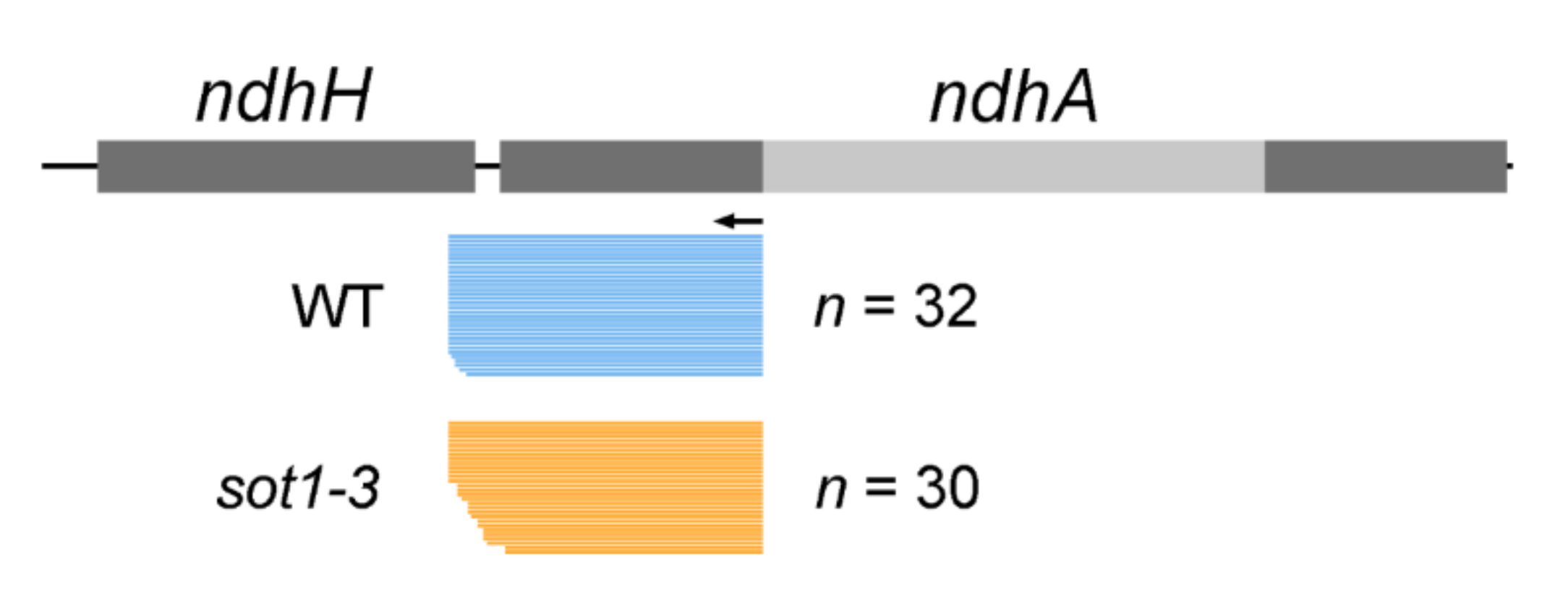
2.2. SOT1 Is Required for the Stabilization of the Processed 5′ Ends of ndhA Transcripts
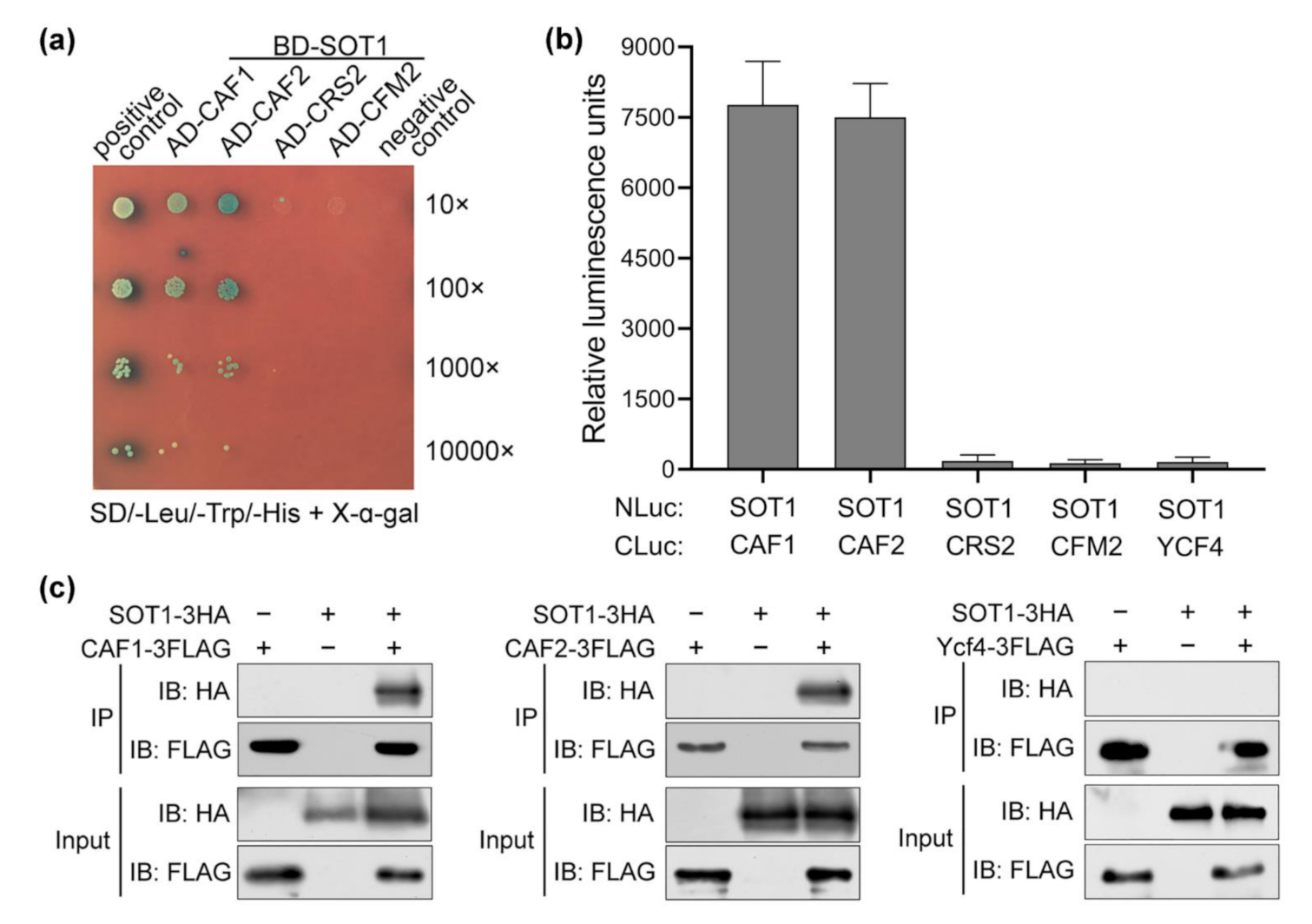
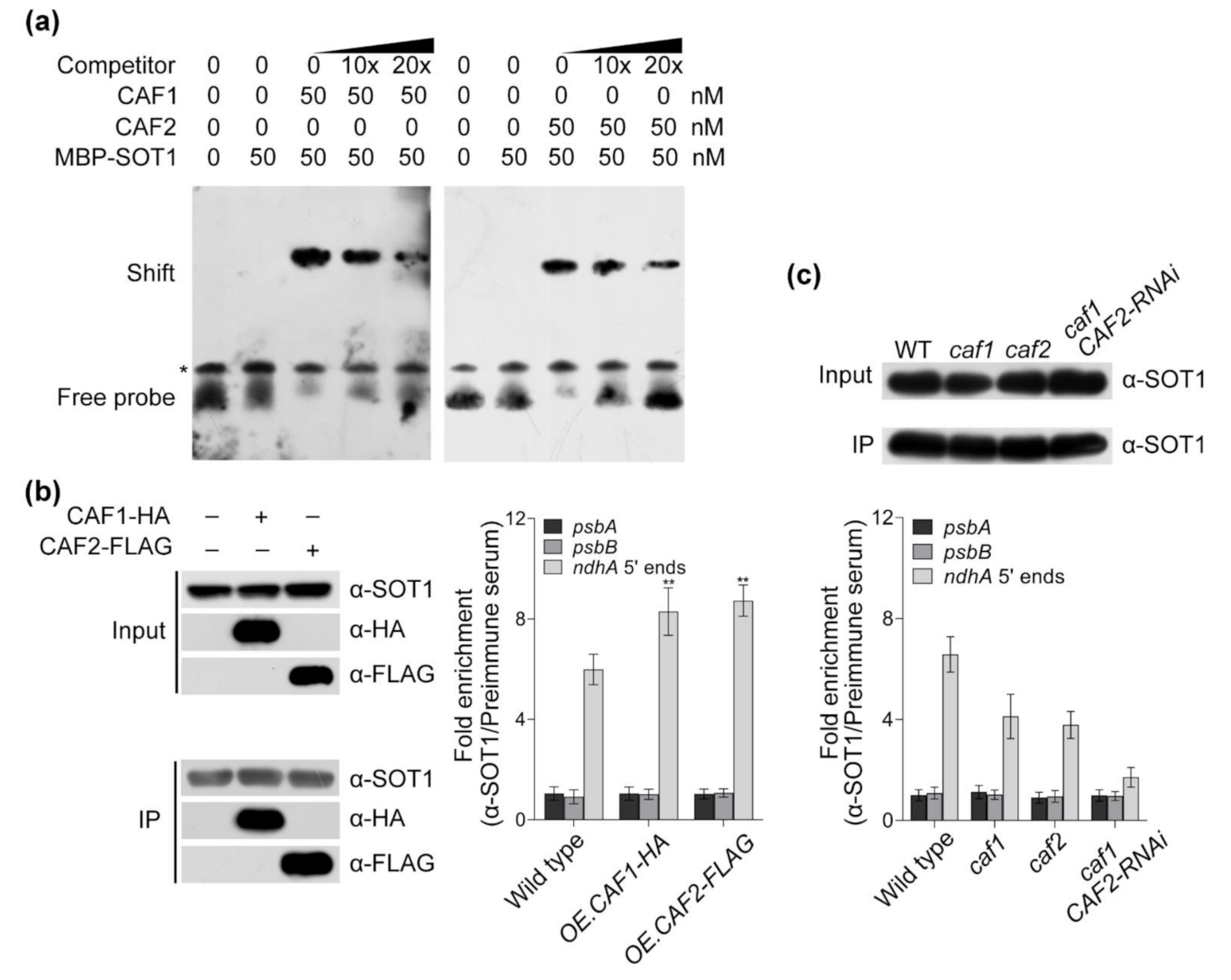
2.3. The SOT1-Mediated Stabilization of ndhA Transcripts Requires the Participation of Other Proteins
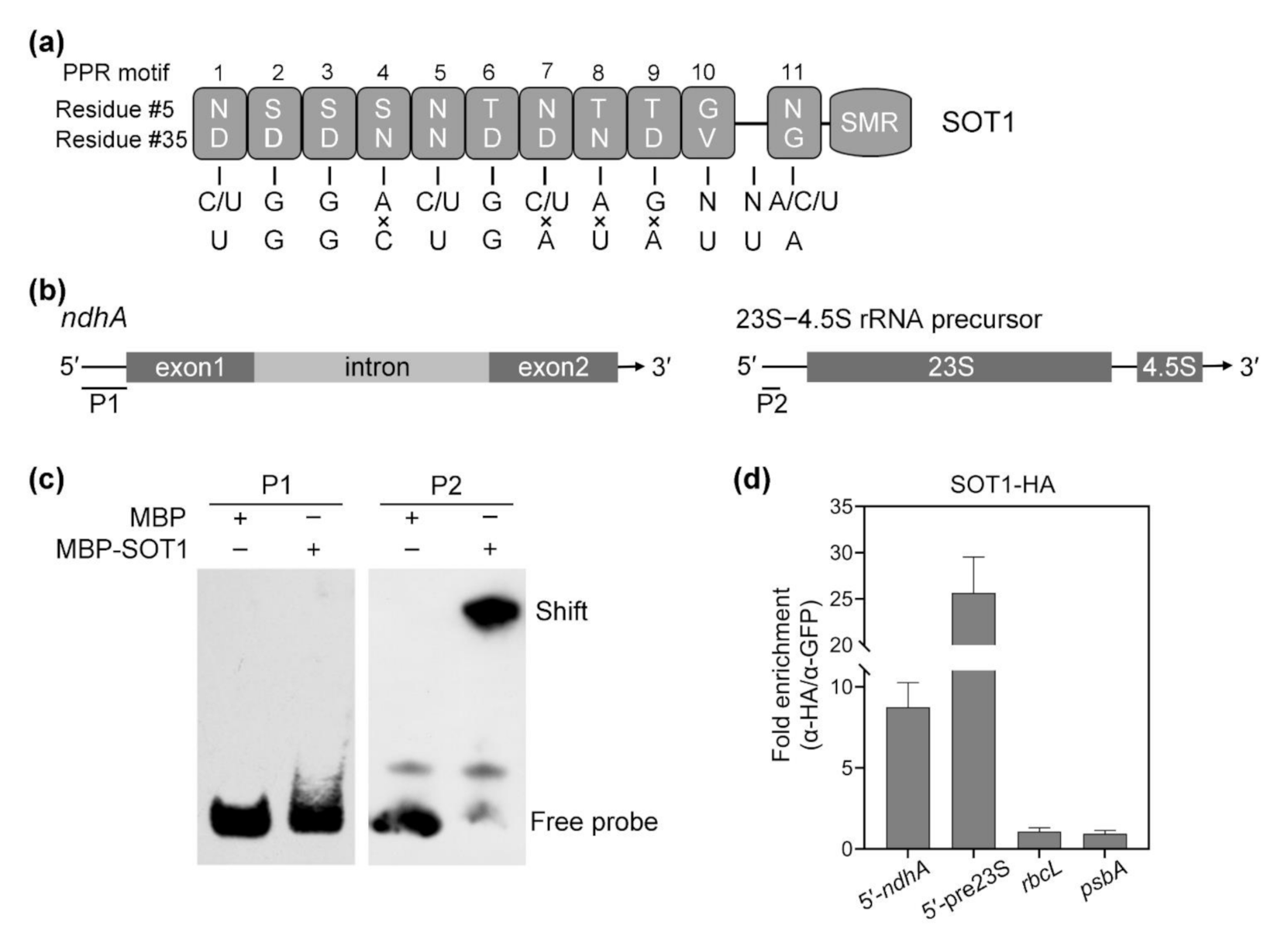
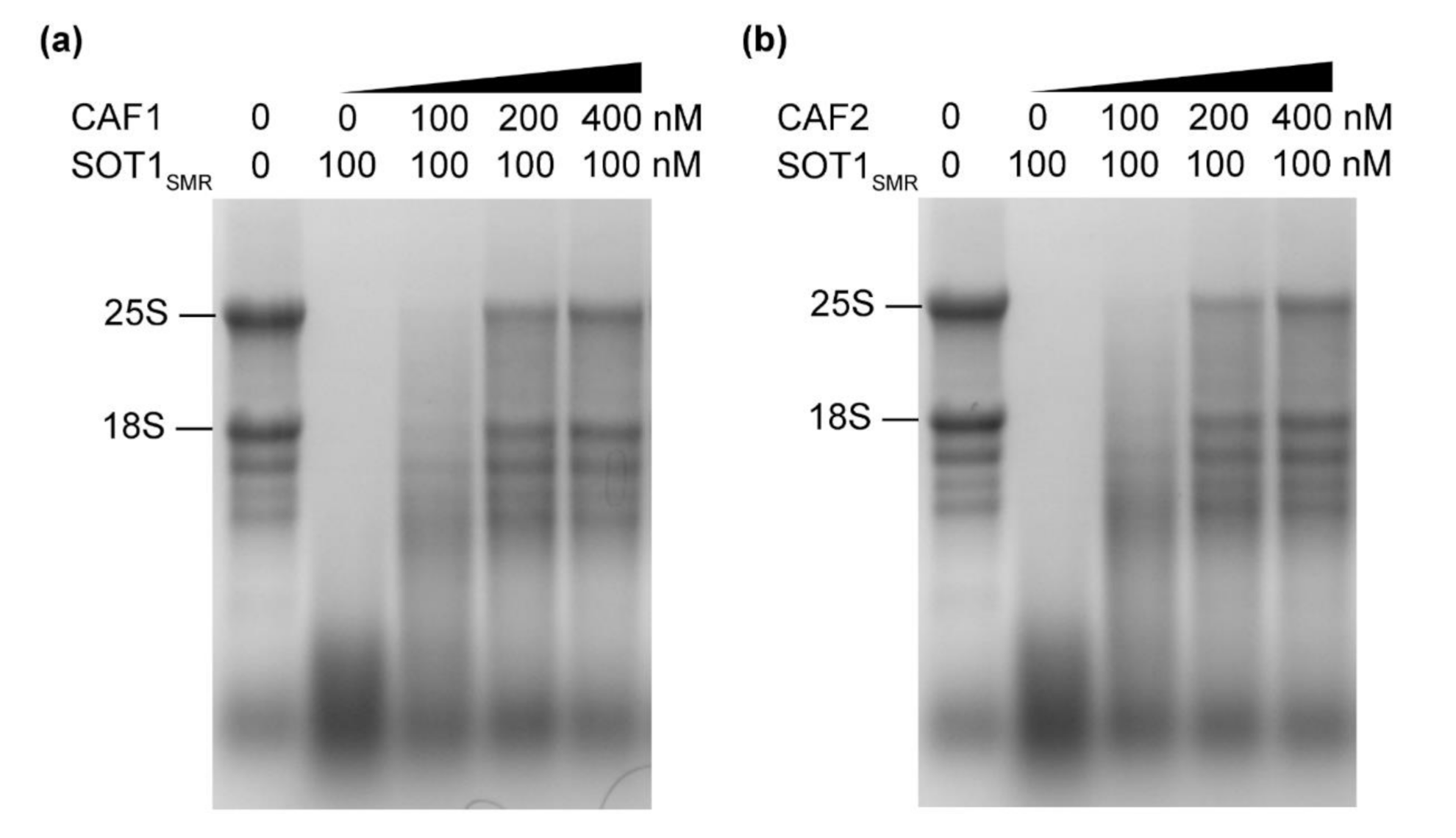
2.4. The CAF Proteins Activate SOT1 to Stabilize ndhA Transcripts by Promoting SOT1 Binding and Inhibiting its RNA Endonuclease Activity
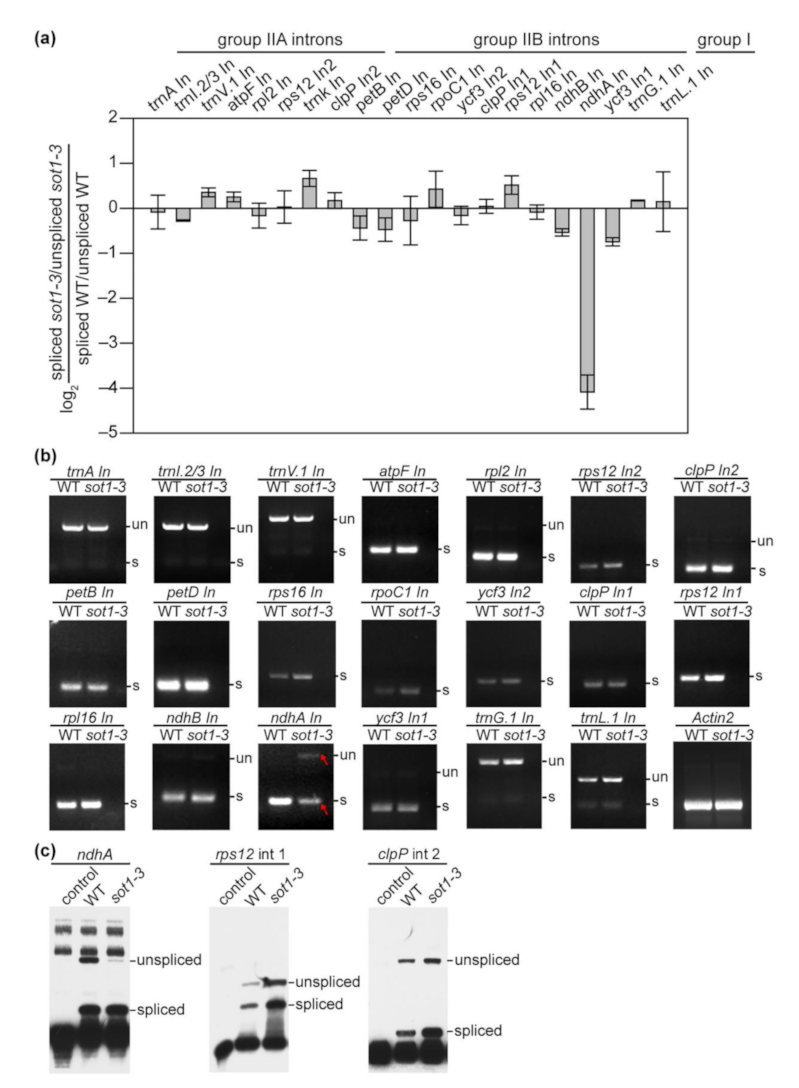
2.5. SOT1 Promotes the Splicing of ndhA Transcripts
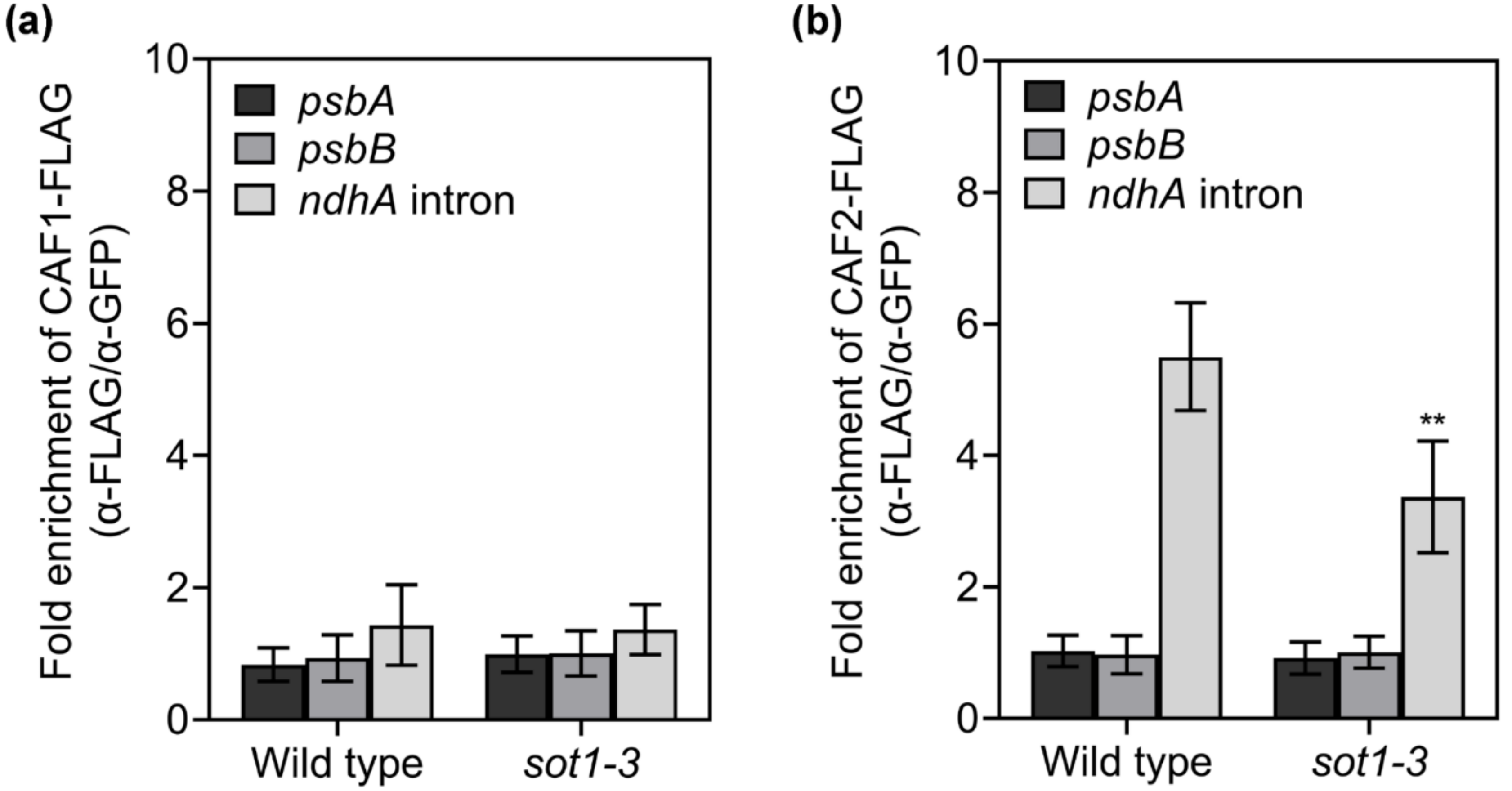
2.6. The Association of CAF2 with the ndhA Intron Is Decreased in sot1-3
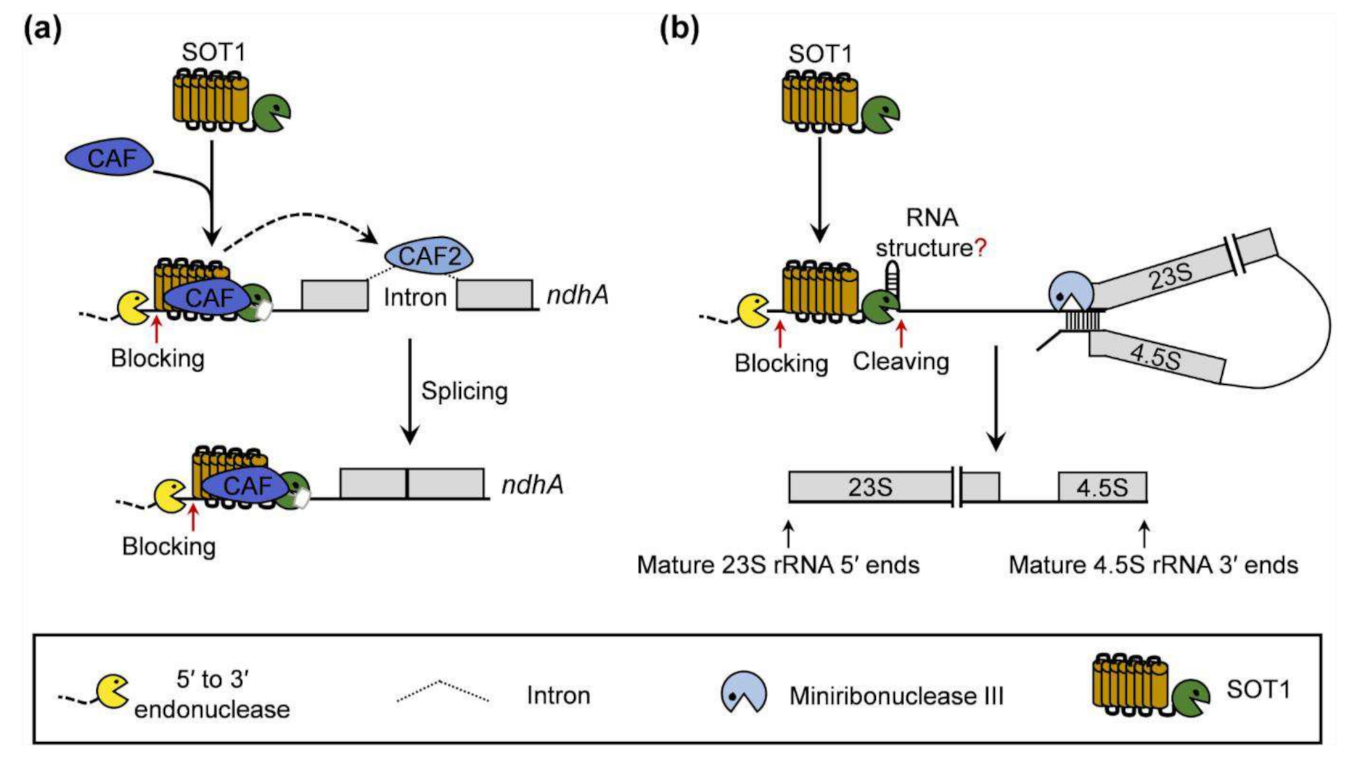
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Strand-specific RNA Sequencing
4.3. RNA Preparation, qPCR, and Gel Blot Analysis
4.4. Rapid Amplification of cDNA Ends
4.5. Yeast Two-Hybrid Assay
4.6. Luciferase Complementation Assays
4.7. Coimmunoprecipitation Assay
4.8. RNA Immunoprecipitation Analysis
4.9. Electrophoretic Mobility Shift Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leister, D.; Wang, L.; Kleine, T. Organellar gene expression and acclimation of plants to environmental stress. Front. Plant Sci. 2017, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, A.; Li, X.; Lu, C. The role of chloroplast gene expression in plant responses to environmental stress. Int. J. Mol. Sci. 2020, 21, 6082. [Google Scholar] [CrossRef] [PubMed]
- Pfalz, J.; Pfannschmidt, T. Essential nucleoid proteins in early chloroplast development. Trends Plant Sci. 2013, 18, 186–194. [Google Scholar] [CrossRef]
- Barkan, A. Expression of plastid genes: Organelle-specific elaborations on a prokaryotic scaffold. Plant Physiol. 2011, 155, 1520–1532. [Google Scholar] [CrossRef] [Green Version]
- Lurin, C.; Andres, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyere, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B.; et al. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Yin, P.; Li, Q.; Yan, C.; Liu, Y.; Liu, J.; Yu, F.; Wang, Z.; Long, J.; He, J.; Wang, H.W.; et al. Structural basis for the modular recognition of single-stranded RNA by PPR proteins. Nature 2013, 504, 168–171. [Google Scholar] [CrossRef]
- Shen, C.; Zhang, D.; Guan, Z.; Liu, Y.; Yang, Z.; Yang, Y.; Wang, X.; Wang, Q.; Zhang, Q.; Fan, S.; et al. Structural basis for specific single-stranded RNA recognition by designer pentatricopeptide repeat proteins. Nat. Commun. 2016, 7, 11285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, N.; Hattori, M.; Andres, C.; Iida, K.; Lurin, C.; Schmitz-Linneweber, C.; Sugita, M.; Small, I. On the expansion of the pentatricopeptide repeat gene family in plants. Mol. Biol. Evol. 2008, 25, 1120–1128. [Google Scholar] [CrossRef]
- Fujii, S.; Small, I. The evolution of RNA editing and pentatricopeptide repeat genes. New Phytol. 2011, 191, 37–47. [Google Scholar] [CrossRef]
- Gutmann, B.; Royan, S.; Schallenberg-Rudinger, M.; Lenz, H.; Castleden, I.R.; McDowell, R.; Vacher, M.A.; Tonti-Filippini, J.; Bond, C.S.; Knoop, V.; et al. The expansion and diversification of pentatricopeptide repeat RNA-editing factors in plants. Mol. Plant 2020, 13, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Melonek, J.; Boykin, L.M.; Small, I.; Howell, K.A. PPR-SMRs: Ancient proteins with enigmatic functions. RNA Biol. 2013, 10, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lu, C. The enigmatic roles of PPR-SMR proteins in plants. Adv. Sci. 2019, 6, 1900361. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Huang, J.; Chory, J. Unraveling the linkage between retrograde signaling and RNA metabolism in plants. Trends Plant Sci. 2020, 25, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Steiner, S.; Schroter, Y.; Pfalz, J.; Pfannschmidt, T. Identification of essential subunits in the plastid-encoded RNA polymerase complex reveals building blocks for proper plastid development. Plant Physiol. 2011, 157, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Liu, H.; Zhai, G.; Wang, L.; Shao, J.; Tao, Y. OspTAC2 encodes a pentatricopeptide repeat protein and regulates rice chloroplast development. J. Genet. Genomics 2016, 43, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, F.; Rodermel, S. An Arabidopsis pentatricopeptide repeat protein, SUPPRESSOR OF VARIEGATION7, is required for FtsH-mediated chloroplast biogenesis. Plant Physiol. 2010, 154, 1588–1601. [Google Scholar] [CrossRef] [Green Version]
- Zoschke, R.; Kroeger, T.; Belcher, S.; Schottler, M.A.; Barkan, A.; Schmitz-Linneweber, C. The pentatricopeptide repeat-SMR protein ATP4 promotes translation of the chloroplast atpB/E mRNA. Plant J. 2012, 72, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, R.; Qu, Y.; Zubo, Y.O.; Borner, T.; Schmitz-Linneweber, C. Mutation of the pentatricopeptide repeat-SMR protein SVR7 impairs accumulation and translation of chloroplast ATP synthase subunits in Arabidopsis thaliana. J. Plant Res. 2013, 126, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Ruwe, H.; Miranda, R.G.; Zoschke, R.; Hase, N.; Schmitz-Linneweber, C.; Barkan, A. Unexpected functional versatility of the pentatricopeptide repeat proteins PGR3, PPR5 and PPR10. Nucleic Acids Res. 2018, 46, 10448–10459. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Y.; Fang, Q.; Zhu, Y.; Zhang, Y.; Liu, X.; Lin, Y.; Barkan, A.; Zhou, F. The PPR-SMR protein ATP4 is required for editing the chloroplast rps8 mRNA in rice and maize. Plant Physiol. 2020, 184, 2011–2021. [Google Scholar] [CrossRef]
- Zhou, W.; Lu, Q.; Li, Q.; Wang, L.; Ding, S.; Zhang, A.; Wen, X.; Zhang, L.; Lu, C. PPR-SMR protein SOT1 has RNA endonuclease activity. Proc. Natl. Acad. Sci. USA 2017, 114, E1554–E1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Liu, S.; Ruwe, H.; Zhang, D.; Melonek, J.; Zhu, Y.; Hu, X.; Gusewski, S.; Yin, P.; Small, I.D.; et al. SOT1, a pentatricopeptide repeat protein with a small MutS-related domain, is required for correct processing of plastid 23S-4.5S rRNA precursors in Arabidopsis thaliana. Plant J. 2016, 85, 607–621. [Google Scholar]
- Zoschke, R.; Watkins, K.P.; Miranda, R.G.; Barkan, A. The PPR-SMR protein PPR53 enhances the stability and translation of specific chloroplast RNAs in maize. Plant J. 2016, 85, 594–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.S.; Chen, J.J.W.; Chiu, C.C.; Hsiao, H.C.W.; Yang, C.J.; Jin, X.H.; Leebens-Mack, J.; de Pamphilis, C.W.; Huang, Y.T.; Yang, L.H.; et al. Concomitant loss of NDH complex-related genes within chloroplast and nuclear genomes in some orchids. Plant J. 2017, 90, 994–1006. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W.; Shikanai, T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu. Rev. Plant Biol. 2016, 67, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Peltier, G.; Aro, E.M.; Shikanai, T. NDH-1 and NDH-2 plastoquinone reductases in oxygenic photosynthesis. Annu. Rev. Plant Biol. 2016, 67, 55–80. [Google Scholar] [CrossRef] [PubMed]
- Maria del Campo, E.; Sabater, B.; Martin, M. Characterization of the 5’- and 3’-ends of mRNAs of ndhH, ndhA and ndhI genes of the plastid ndhH-D operon. Biochimie 2006, 88, 347–357. [Google Scholar]
- Stern, D.B.; Goldschmidt-Clermont, M.; Hanson, M.R. Chloroplast RNA metabolism. Annu. Rev. Plant Biol. 2010, 61, 125–155. [Google Scholar] [CrossRef]
- Barkan, A.; Rojas, M.; Fujii, S.; Yap, A.; Chong, Y.S.; Bond, C.S.; Small, I. A combinatorial amino acid code for RNA recognition by pentatricopeptide repeat proteins. PLoS Genet. 2012, 8, e1002910. [Google Scholar] [CrossRef]
- Yagi, Y.; Hayashi, S.; Kobayashi, K.; Hirayama, T.; Nakamura, T. Elucidation of the RNA recognition code for pentatricopeptide repeat proteins involved in organelle RNA editing in plants. PLoS ONE 2013, 8, e57286. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yao, Y.; Hong, S.; Yang, Y.; Shen, C.; Zhang, Q.; Zhang, D.; Zou, T.; Yin, P. Delineation of pentatricopeptide repeat codes for target RNA prediction. Nucleic Acids Res. 2019, 47, 3728–3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryamanesh, N.; Ruwe, H.; Vincis Pereira Sanglard, L.; Eshraghi, L.; Bussell, J.D.; Howell, K.A.; Colas des Francs-Small, C.; Small, I.D. The pentatricopeptide repeat protein EMB2654 is essential for trans-splicing of a chloroplast small ribosomal subunit transcript. Plant Physiol. 2017, 173, 1164–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakura, Y.; Barkan, A. A CRM domain protein functions dually in group I and group II intron splicing in land plant chloroplasts. Plant Cell 2007, 19, 3864–3875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkan, A.; Klipcan, L.; Ostersetzer, O.; Kawamura, T.; Asakura, Y.; Watkins, K.P. The CRM domain: An RNA binding module derived from an ancient ribosome-associated protein. RNA 2007, 13, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostheimer, G.J.; Rojas, M.; Hadjivassiliou, H.; Barkan, A. Formation of the CRS2-CAF2 group II intron splicing complex is mediated by a 22-amino acid motif in the COOH-terminal region of CAF2. J. Biol. Chem. 2006, 281, 4732–4738. [Google Scholar] [CrossRef] [Green Version]
- Ostheimer, G.J.; Williams-Carrier, R.; Belcher, S.; Osborne, E.; Gierke, J.; Barkan, A. Group II intron splicing factors derived by diversification of an ancient RNA-binding domain. EMBO J. 2003, 22, 3919–3929. [Google Scholar] [CrossRef] [Green Version]
- Asakura, Y.; Barkan, A. Arabidopsis orthologs of maize chloroplast splicing factors promote splicing of orthologous and species-specific group II introns. Plant Physiol. 2006, 142, 1656–1663. [Google Scholar] [CrossRef] [Green Version]
- De Longevialle, A.F.; Small, I.D.; Lurin, C. Nuclearly encoded splicing factors implicated in RNA splicing in higher plant organelles. Mol. Plant 2010, 3, 691–705. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Linneweber, C.; Williams-Carrier, R.; Barkan, A. RNA immunoprecipitation and microarray analysis show a chloroplast Pentatricopeptide repeat protein to be associated with the 5’ region of mRNAs whose translation it activates. Plant Cell 2005, 17, 2791–2804. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhang, Q.; Guan, Z.; Wang, Q.; Li, L.; Ruan, F.; Lin, R.; Zou, T.; Yin, P. MORF9 increases the RNA-binding activity of PLS-type pentatricopeptide repeat protein in plastid RNA editing. Nat. Plants 2017, 3, 17037. [Google Scholar] [CrossRef]
- Zhao, C.; Pyle, A.M. Structural insights into the mechanism of group II intron splicing. Trends Biochem. Sci. 2017, 42, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Meteignier, L.V.; Ghandour, R.; Meierhoff, K.; Zimmerman, A.; Chicher, J.; Baumberger, N.; Alioua, A.; Meurer, J.; Zoschke, R.; Hammani, K. The Arabidopsis mTERF-repeat MDA1 protein plays a dual function in transcription and stabilization of specific chloroplast transcripts within the psbE and ndhH operons. New Phytol. 2020, 227, 1376–1391. [Google Scholar] [CrossRef] [PubMed]
- Hotto, A.M.; Castandet, B.; Gilet, L.; Higdon, A.; Condon, C.; Stern, D.B. Arabidopsis chloroplast mini-ribonuclease III participates in rRNA maturation and intron recycling. Plant Cell 2015, 27, 724–740. [Google Scholar] [CrossRef]
- Castandet, B.; Hotto, A.M.; Strickler, S.R.; Stern, D.B. ChloroSeq, an optimized chloroplast RNA-seq bioinformatic pipeline, reveals remodeling of the organellar transcriptome under heat stress. G3 2016, 6, 2817–2827. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Zhang, Y.; Hu, Z.; Huang, X.; Zhang, B.; Lu, Q.; Wen, X.; Wang, Y.; Lu, C. mTERF5 acts as a transcriptional pausing factor to positively regulate transcription of chloroplast psbEFLJ. Mol. Plant 2019, 12, 1259–1277. [Google Scholar] [CrossRef]
- Chen, H.; Zou, Y.; Shang, Y.; Lin, H.; Wang, Y.; Cai, R.; Tang, X.; Zhou, J.M. Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 2008, 146, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, Z.; Zhang, Y.; Zhou, W.; Zhang, A.; Lu, C. Pentatricopeptide repeat protein PHOTOSYSTEM I BIOGENESIS FACTOR2 is required for splicing of ycf3. J. Integr. Plant Biol. 2020, 62, 1741–1761. [Google Scholar] [CrossRef] [PubMed]
- Pfalz, J.; Bayraktar, O.A.; Prikryl, J.; Barkan, A. Site-specific binding of a PPR protein defines and stabilizes 5’ and 3’ mRNA termini in chloroplasts. EMBO J. 2009, 28, 2042–2052. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Luo, W.; Zhou, W.; Yin, X.; Wang, X.; Li, X.; Jiang, C.; Zhang, Q.; Kang, X.; Zhang, A.; et al. CAF Proteins Help SOT1 Regulate the Stability of Chloroplast ndhA Transcripts. Int. J. Mol. Sci. 2021, 22, 12639. https://doi.org/10.3390/ijms222312639
Li X, Luo W, Zhou W, Yin X, Wang X, Li X, Jiang C, Zhang Q, Kang X, Zhang A, et al. CAF Proteins Help SOT1 Regulate the Stability of Chloroplast ndhA Transcripts. International Journal of Molecular Sciences. 2021; 22(23):12639. https://doi.org/10.3390/ijms222312639
Chicago/Turabian StyleLi, Xiuming, Wenzhen Luo, Wen Zhou, Xiaopeng Yin, Xuemei Wang, Xiujin Li, Chenchen Jiang, Qingqing Zhang, Xiaojing Kang, Aihong Zhang, and et al. 2021. "CAF Proteins Help SOT1 Regulate the Stability of Chloroplast ndhA Transcripts" International Journal of Molecular Sciences 22, no. 23: 12639. https://doi.org/10.3390/ijms222312639
APA StyleLi, X., Luo, W., Zhou, W., Yin, X., Wang, X., Li, X., Jiang, C., Zhang, Q., Kang, X., Zhang, A., Zhang, Y., & Lu, C. (2021). CAF Proteins Help SOT1 Regulate the Stability of Chloroplast ndhA Transcripts. International Journal of Molecular Sciences, 22(23), 12639. https://doi.org/10.3390/ijms222312639