
A Wheat TaTOE1-B1 Transcript TaTOE1-B1-3 Can Delay the Flowering Time of Transgenic Arabidopsis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analysis of TaTOE-B1
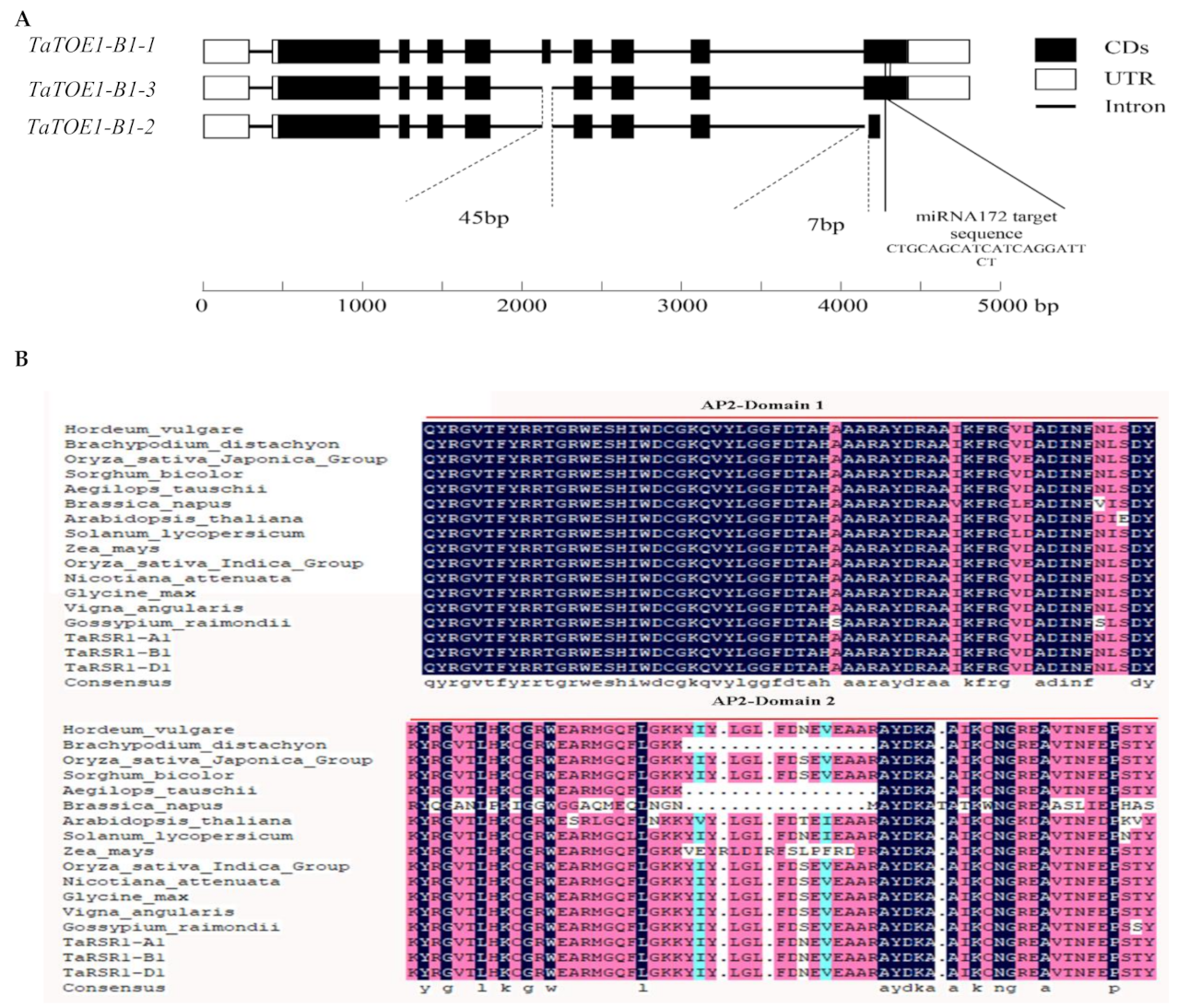
2.2. Cloning of TaTOE1-B1-1 and TaTOE1-B1-3
2.3. Overexpression of TaTOE1-B1-3 Results in Considerable Delayed Flowering Time in Transgenic Arabidopsis
2.4. Overexpression of TaTOE1-B1-3 Downregulates AtFT and AtSOC1 Expression in Transgenic Arabidopsis
2.5. TaTOE1-B1-1 and TaTOE1-B1-3 Are Localized in the Nucleus and Have Transcriptional Activation Activity
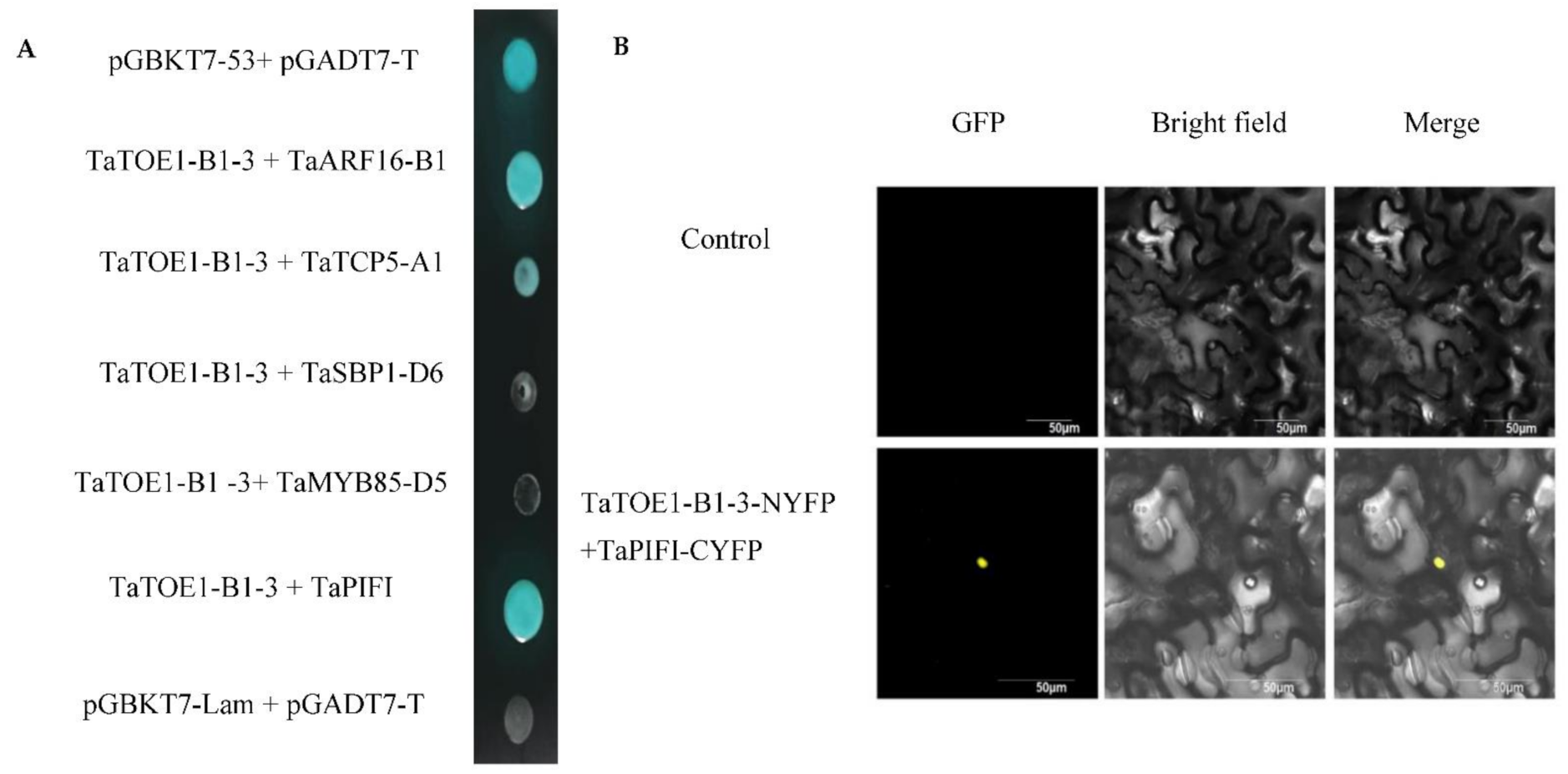
2.6. Identification of Interaction between Proteins TaTOE1-B1-3 and TaPIFI
3. Discussion
3.1. Differences in 15 Amino Acids May Be Important to the Functional Difference between TaTOE1-B1-1 and TaTOE1-B1-3
3.2. The Main Reason for Flowering Delay May Be Downregulated Expression of FT and SOC1
3.3. TaTOE1-B1-3 May Act as a Broad Floral Inhibitor in Plants
3.4. Protein Interaction between TaTOE1-B1-3 and TaPIFI May Play an Important Role in Flowering Inhibition
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Bioinformatics Analysis of Sequence
4.3. Isolation of TaTOE-B1-1 and TaTOE1-B1-3
4.4. Construction of TaTOE1-B1-1 and TaTOE1-B1-3 Transgenic Arabidopsis Lines
4.5. Phenotypic Identification of Transgenic Progeny
4.6. qRT-PCR Analysis
4.7. Subcellular Localization
4.8. Transcriptional Activator Activity Analysis
4.9. Prediction and Library Screening of Candidate TaTOE1-B1-3 Interaction Proteins
4.10. Point-to-Point Analysis in Y2H System
4.11. Bimolecular Fluorescence Complementation (BiFC) Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niu, X.M.; Xu, Y.C.; Li, Z.W.; Bian, Y.T.; Hou, X.H.; Chen, J.F.; Zou, Y.P.; Jiang, J.; Wu, X.; Ge, S.; et al. Transposable elements drive rapid phenotypic variation in Capsella rubella. Proc. Natl. Acad. Sci. USA 2019, 116, 6908–6913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.B.; Fu, X.D.; Yin, C.B.; Lu, F. Wheat speciation and adaptation: Perspectives from reticulate evolution. aBIOTECH 2021. (Preprint). [Google Scholar] [CrossRef]
- Laurent, C.; George, C. The quest for florigen: A review of recent progress. J. Exp. Bot. 2006, 57, 3395–3403. [Google Scholar]
- Shaw, L.M.; Turner, A.S.; Laurie, D.A. The impact of photoperiod insensitive Ppd-D1a mutations on the photoperiod pathway across the three genomes of hexaploid wheat (Triticum aestivum). Plant J. 2012, 71, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Chong, K. Remembering winter through vernalization. Nat. Plants 2018, 4, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.J.; Hua, C.M.; Shen, L.S.; Yu, H. New insights into gibberellin signaling in regulating flowering in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, F.; Li, H.; Gul, A.; Subhani, A.; Ali, A.; Mujeeb, K.A.; Rasheed, A. Genome-Wide analyses reveal footprints of divergent selection and drought adaptive traits in synthetic-derived wheats. G3 Genes Genomes Genet. 2019, 9, 1957–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdennour, S.; Houcine, B.; Rhouma, S.; Sahbi, F.; Tahar, S. Stability and adaptability concepts of bread wheat (Triticum aestivum L.) in the northwest of Tunisia. Biol. Futura 2019, 70, 240–250. [Google Scholar] [CrossRef]
- Akram, S.; Arif, M.A.R.; Hameed, A. A GBS-based GWAS analysis of adaptability and yield traits in bread wheat (Triticum aestivum L.). J. Appl. Genet. 2021, 62, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Worland, A.J. The influence of flowering time genes on environmental adaptability in European wheats. Euphytica 1996, 89, 49–57. [Google Scholar] [CrossRef]
- Kamran, A.; Iqbal, M.; Spaner, D. Flowering time in wheat (Triticum aestivum L.): A key factor for global adaptability. Euphytica 2014, 197, 1–26. [Google Scholar] [CrossRef]
- Streck, N.A.; Weiss, A.; Baenziger, P.S. A generalized vernalization response function for winter wheat. Agron. J. 2003, 95, 155–159. [Google Scholar] [CrossRef]
- Yan, L.L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.L.; Loukoianov, A.; Blechl, A.; Tranquilli, G.; Ramakrishna, W.; SanMiguel, P.; Dubcovsky, J. The wheat VRN2 gene is a flowering repressor down-Regulated by vernalization. Science 2004, 303, 1640–1644. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Q.; Yu, M.; Fang, T.L.; Cao, S.H.; Brett, F.C.; Yan, L.L. Vernalization requirement duration in winter wheat is controlled by TaVRN-A1 at the protein level. Plant J. 2013, 76, 742–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kippes, N.; Debernardi, J.M.; Vasquez-Gross, H.A.; Akpinar, B.A.; Budak, H.; Kato, K.; Dubcovsky, J. Identification of the VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proc. Natl. Acad. Sci. USA 2015, 115, 5401–5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.Y.; Wang, X.; Wei, L.; Wang, J.X.; Yin, J. Characterization of Ppd-D1 alleles on the developmental traits and rhythmic expression of photoperiod genes in common wheat. J. Integr. Agric. 2016, 15, 502–511. [Google Scholar] [CrossRef]
- Errum, A.; Rehman, N.; Khan, M.R.; Ali, G.M. Genome-wide characterization and expression analysis of pseudo-response regulator gene family in wheat. Mol. Biol. Rep. 2021, 48, 2411–2427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.L.; Wang, L.; Zeng, L.P.; Zhang, C.; Ma, H. Arabidopsis TOE proteins convey a photoperiodic signal to antagonize CONSTANS and regulate flowering time. Genes Dev. 2015, 29, 975–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamuro, J.K.; Caster, B.; Villarroel, R.; Van Montagu, M.; Jofuku, K.D. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 7076–7081. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.C.; Feng, K.W.; Wang, M.X.; Wang, M.; Deng, P.C.; Song, W.N.; Nie, X.J. Genome-Wide identification, phylogeny and expression analysis of AP2/ERF transcription factors family in Brachypodium distachyon. BMC Genom. 2016, 17, 636. [Google Scholar] [CrossRef] [PubMed]
- Simons, J.K. Molecular characterization of the major wheat domestication gene Q. Genetics 2005, 172, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, J.R.; Finnegan, E.J.; Watanabe, N.; Trevaskis, B.; Swain, S.M. New alleles of the wheat domestication gene Q reveal multiple roles in growth and reproductive development. Development 2017, 144, 1959–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Liu, J.; Dong, H.X.; Sun, J.Q. Functional regulation of Q by microRNA172 and transcriptional co-repressor TOPLESS in controlling bread wheat spikelet density. Plant Biotechnol. J. 2017, 16, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.J.; Chen, Q.; Zheng, T.; Jiang, Y.F.; Qiao, Y.Y.; Guo, Z.R.; Qi, P.F. An overexpressed Q allele leads to increased spike density and improved processing qualityin common wheat (Triticum aestivum). G3 Genes Genomes Genet. 2018, 8, 771–778. [Google Scholar]
- Elliott, R.C.; Betzner, A.S.; Huttner, E.; Oakes, M.P.; Tucker, W.Q.; Gerentes, D.; Smyth, D.R. AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 1996, 8, 155–168. [Google Scholar]
- Zhao, L.F.; Xu, S.B.; Chai, T.Y.; Wang, T. OsAP2-1, an AP2-like gene from Oryza sativa, is required for flower development and male fertility. Sex Plant Reprod. 2006, 19, 197–206. [Google Scholar] [CrossRef]
- Krizek, B. AINTEGUMENTA and AINTEGUMENTA-LIKE6 act redundantly to regulate Arabidopsis floral growth and patterning. Plant Physiol. 2009, 150, 1916–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moose, S.P.; Sisco, P.H. Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity. Genes Dev. 1996, 10, 3018–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.H.; Helliwell, C.A. Regulation of flowering time and floral patterning by miR172. J. Exp. Bot. 2011, 62, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Zikhali, M.; Wingen, L.U.; Leverington-Waite, M.; Specel, S.; Griffiths, S. The identification of new candidate genes Triticum aestivum FLOWERING LOCUS T3-B1 (TaFT3-B1) and TARGET OF EAT1 (TaTOE1-B1) controlling the short-day photoperiod response in bread wheat. Plant Cell Environ. 2017, 40, 2678–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.H.; Seo, Y.H.; Seo, P.J.; Jose, L.R.; Yun, J.; Chua, N.H.; Park, C.M. The GIGANTEA-Regulated MicroRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.H.; Lee, S.M.; Yun, J.; Lee, M.Y.; Park, C.M. The miR172 target TOE3 represses AGAMOUS expression during Arabidopsis floral patterning. Plant Sci. 2014, 215–216, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.Z.; Xu, W.; Liu, G.Q.; Peng, X.Q.; Guo, T.C. Comprehensive analysis of the transcription of starch synthesis genes and the transcription factor RSR1 in wheat (Triticum aestivum) endosperm. Genome 2013, 56, 115–122. [Google Scholar] [CrossRef]
- Wang, M.J.; Ding, L.; Liu, X.H.; Liu, J.X. Two B-box domain proteins, BBX28 and BBX29, regulate flowering time at low ambient temperature in Arabidopsis. Plant Mol. Biol. 2021, 106, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Prem, N. Identifying Objectives for breeding improved vegetable varieties-hard but vital Choice. J. Food Nutr. Res. 2014, 2, 867–871. [Google Scholar]
- Syed, N.H.; Kalyna, M.; Marquez, Y.; Barta, A.; Brown, J.W. Alternative splicing in plants-coming of age. Trends Plant Sci. 2012, 17, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, R.; Crespi, M.; Bennett, M.J. One Gene, Many Proteins: Mapping cell-specific alternative splicing in plants. Dev. Cell 2016, 39, 383–385. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.X.; Zhang, D.D.; Li, Z.W.; Liang, H.L.; Deng, R.F.; Su, X.G.; Jiang, Y.M.; Duan, X.W. Alternative splicing of MaMYB16L regulates starch degradation in banana fruit during ripening. J. Integr. Plant Biol. 2021, 63, 1341–1352. [Google Scholar] [CrossRef]
- Simpson, C.G.; Hedley, P.E.; Watters, J.A.; Clark, G.P.; McQuade, C.; Machray, G.C.; Brown, J.W. Requirements for mini-exon inclusion in potato invertase mRNAs provides evidence for exon-scanning interactions in plants. Cold Spring Harb. Lab. Press 2000, 6, 422–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabala, G.; Vodkin, L. Novel exon combinations generated by alternative splicing of gene fragments mobilized by a CACTA transposon in Glycine max. BMC Plant Biol. 2007, 7, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qüesta, J.I.; Song, J.; Geraldo, N.; An, H.; Dean, C. Arabidopsis transcriptional repressor VAL1 triggers Polycomb silencing at FLC during vernalization. Science 2016, 353, 485–488. [Google Scholar] [CrossRef]
- Shim, J.S.; Kubota, A.; Imaizumi, T. Circadian clock and photoperiodic flowering in Arabidopsis: CONSTANS is a hub for signal integration. Plant Physiol. 2017, 173, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.; Susila, H.; Nasim, Z.; Jung, J.Y.; Ahn, J.H. Arabidopsis ABF3 and ABF4 transcription factors act with the NF-YC complex to regulate SOC1 expression and mediate drought-accelerated flowering. Mol. Plant 2019, 12, 489–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Portis, A.R., Jr. A novel nucleus-encoded Chloroplast protein, PIFI, is involved in NAD(P)H dehydrogenase complex-mediated Chlororespiratory Electron Transport in Arabidopsis. Plant Physiol. 2007, 144, 1742–1752. [Google Scholar] [CrossRef] [Green Version]
- Dal Bosco, C.; Lezhneva, L.; Biehl, A.; Leister, D.; Strotmann, H.; Wanner, G.; Meurer, J. Inactivation of the Chloroplast ATP synthase γ subunit results in high non-photochemical fluorescence quenching and altered nuclear gene expression in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 1060–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Henriques, R.; Lin, S.S. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protocols 2006, 1, 641–646. [Google Scholar] [CrossRef]
- Tomasz, C.; Mark, S.; Thomas, A.; Michael, K.U.; Wolf-Rüd, S. Genome-Wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar]
- Li, Z.; Ma, S.C.; Liu, D.; Zhang, L.L.; Du, X.J.; Xia, Y.; Song, Q.L.; Li, Y.; Zhang, Y.M.; Li, Z.L.; et al. Morphological and proteomic analysis of young spikes reveals new insights into the formation of multiple-pistil wheat. Plant Sci. 2020, 296, 110503. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Song, T.Q.; Zhou, J.F.; Cheng, J.; Li, R.B.; Yu, M.; Zhang, Y.R.; Yu, Y.; Zhang, B.; Zhang, X.K. A transcription factor TaTCP20 regulates the expression of Ppd-D1b in common wheat. Plant Biotechnol. Rep. 2021, 15, 359–367. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, T.; Yu, Y.; Zhang, M.; Zhou, H.; Zhang, S.; Yu, M.; Zhou, J.; Cheng, J.; Xiang, J.; Yang, S.; et al. A Wheat TaTOE1-B1 Transcript TaTOE1-B1-3 Can Delay the Flowering Time of Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 12645. https://doi.org/10.3390/ijms222312645
Song T, Yu Y, Zhang M, Zhou H, Zhang S, Yu M, Zhou J, Cheng J, Xiang J, Yang S, et al. A Wheat TaTOE1-B1 Transcript TaTOE1-B1-3 Can Delay the Flowering Time of Transgenic Arabidopsis. International Journal of Molecular Sciences. 2021; 22(23):12645. https://doi.org/10.3390/ijms222312645
Chicago/Turabian StyleSong, Tianqi, Yang Yu, Mingfei Zhang, Hongwei Zhou, Shuangxing Zhang, Ming Yu, Jianfei Zhou, Jie Cheng, Jishan Xiang, Songjie Yang, and et al. 2021. "A Wheat TaTOE1-B1 Transcript TaTOE1-B1-3 Can Delay the Flowering Time of Transgenic Arabidopsis" International Journal of Molecular Sciences 22, no. 23: 12645. https://doi.org/10.3390/ijms222312645
APA StyleSong, T., Yu, Y., Zhang, M., Zhou, H., Zhang, S., Yu, M., Zhou, J., Cheng, J., Xiang, J., Yang, S., & Zhang, X. (2021). A Wheat TaTOE1-B1 Transcript TaTOE1-B1-3 Can Delay the Flowering Time of Transgenic Arabidopsis. International Journal of Molecular Sciences, 22(23), 12645. https://doi.org/10.3390/ijms222312645