
Porphyromonas gingivalis Components/Secretions Synergistically Enhance Pneumonia Caused by Streptococcus pneumoniae in Mice

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
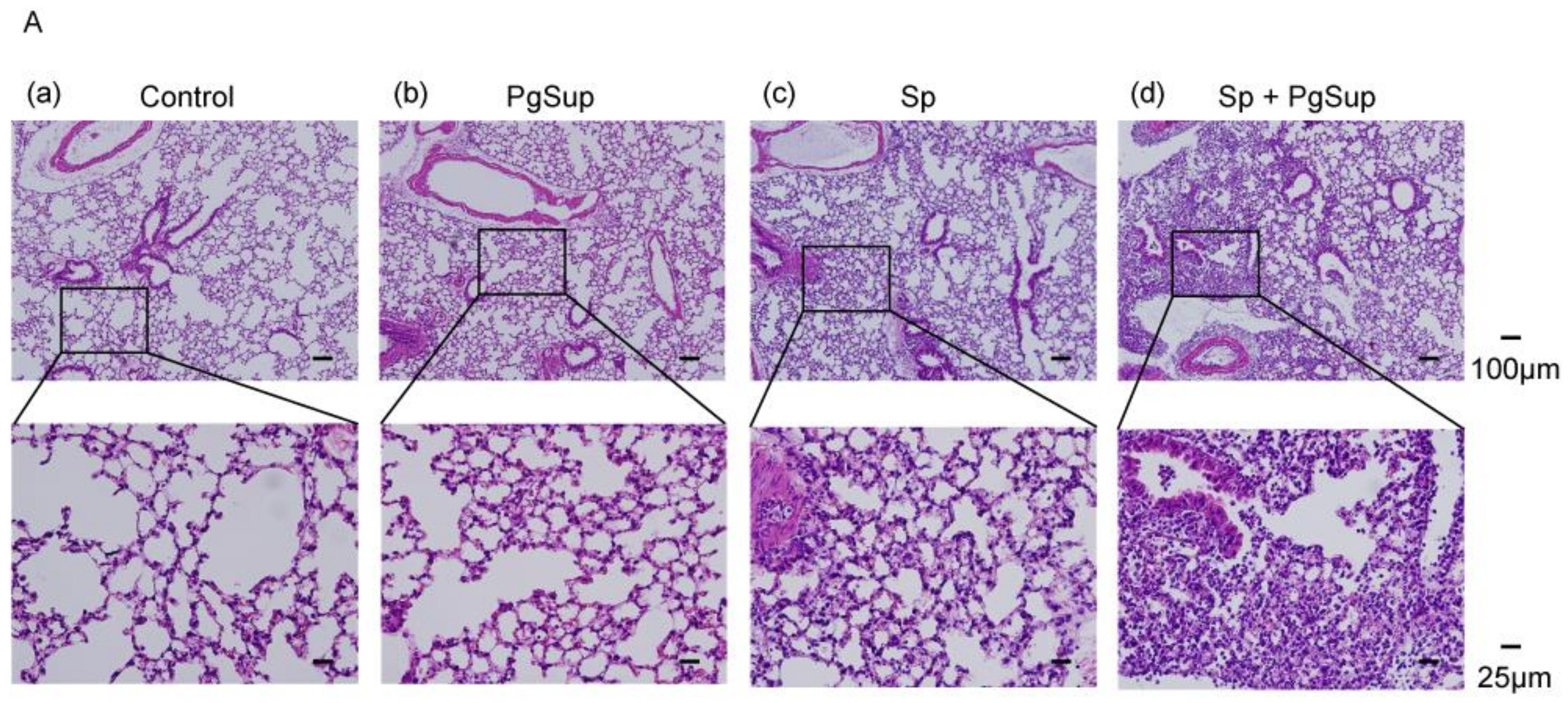
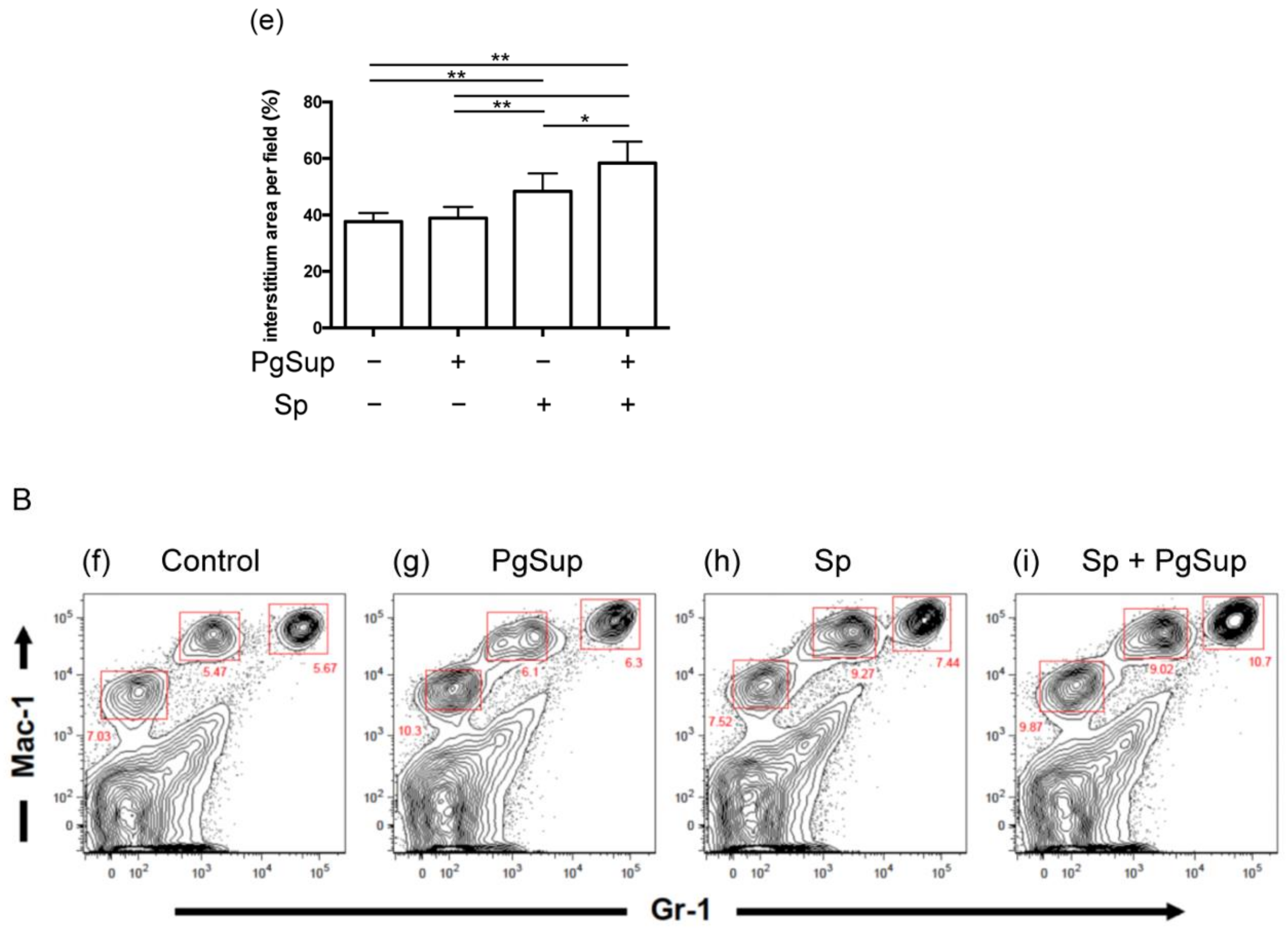
2.1. P. gingivalis Culture Supernatant (PgSup) Enhanced the Inflammation of S. pneumoniae (Sp) Infection, as Observed by Lung Histology and Flow Cytometric Analysis
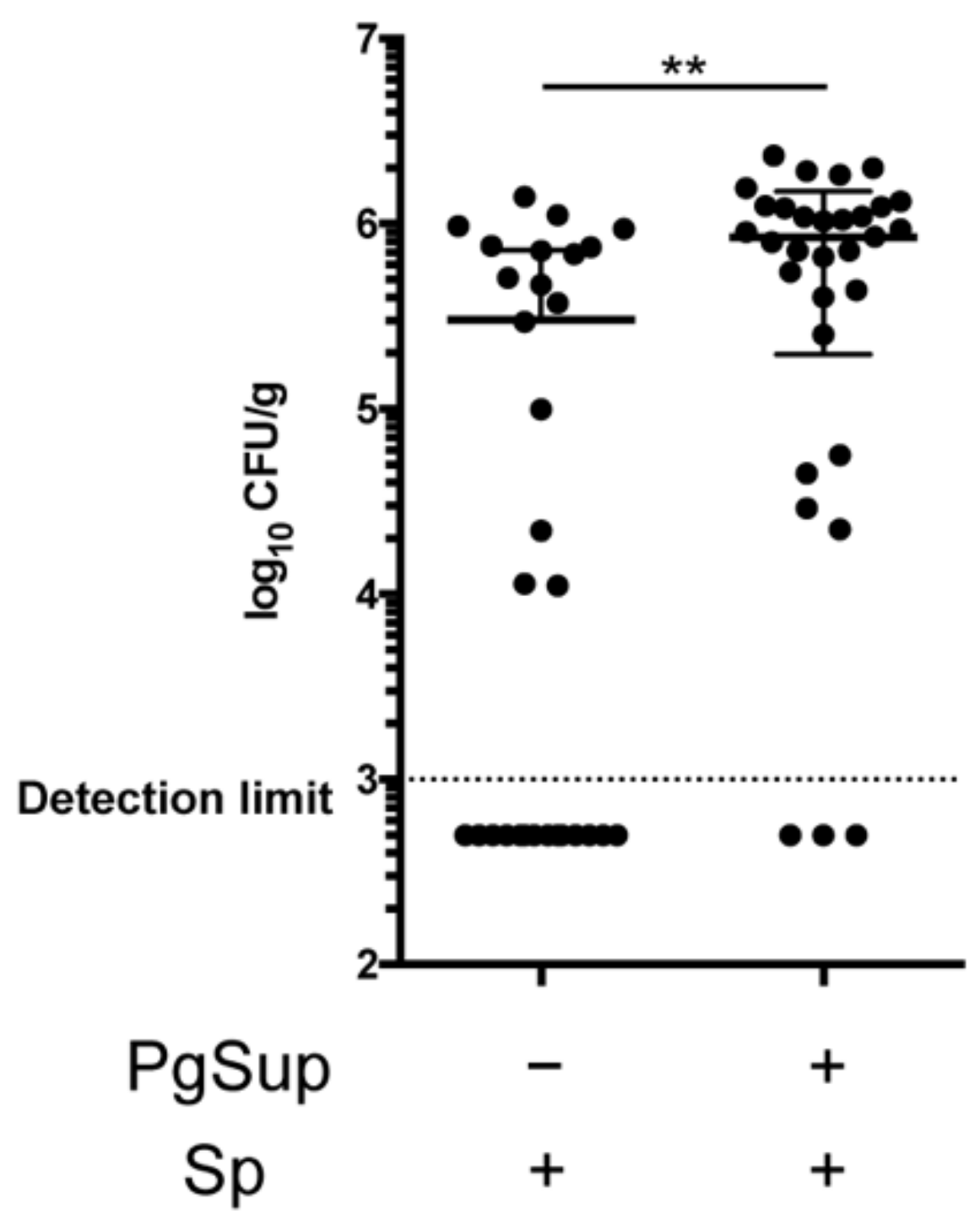
2.2. PgSup Increased the Number of S. pneumoniae in the Lung
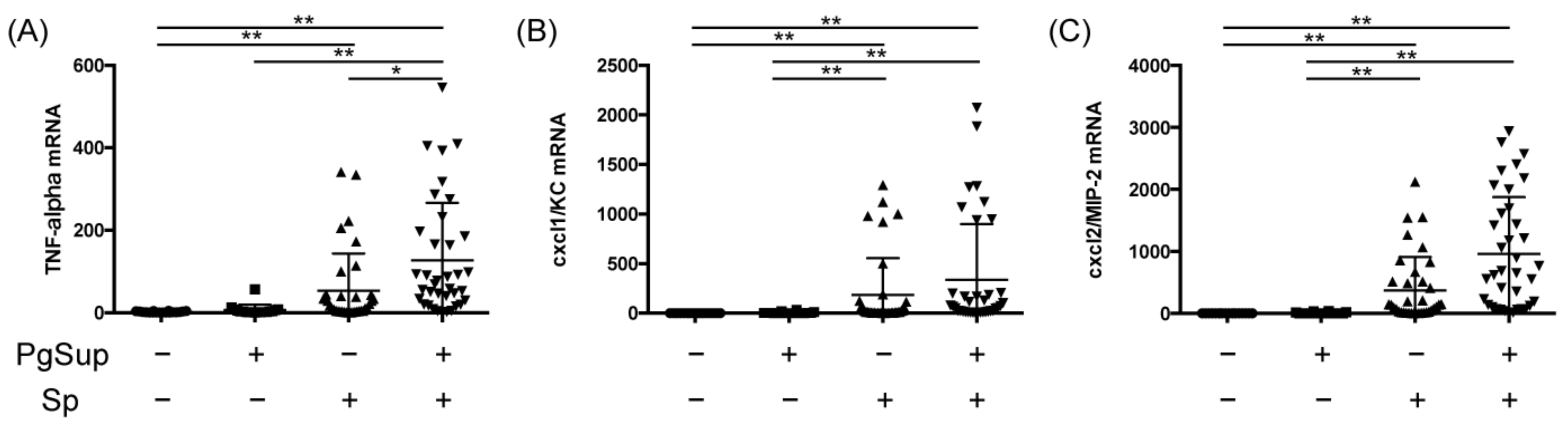
2.3. PgSup Enhanced Pro-Inflammatory Cytokine and Chemokine Gene Expression in the Lungs of Sp-Infected Mice
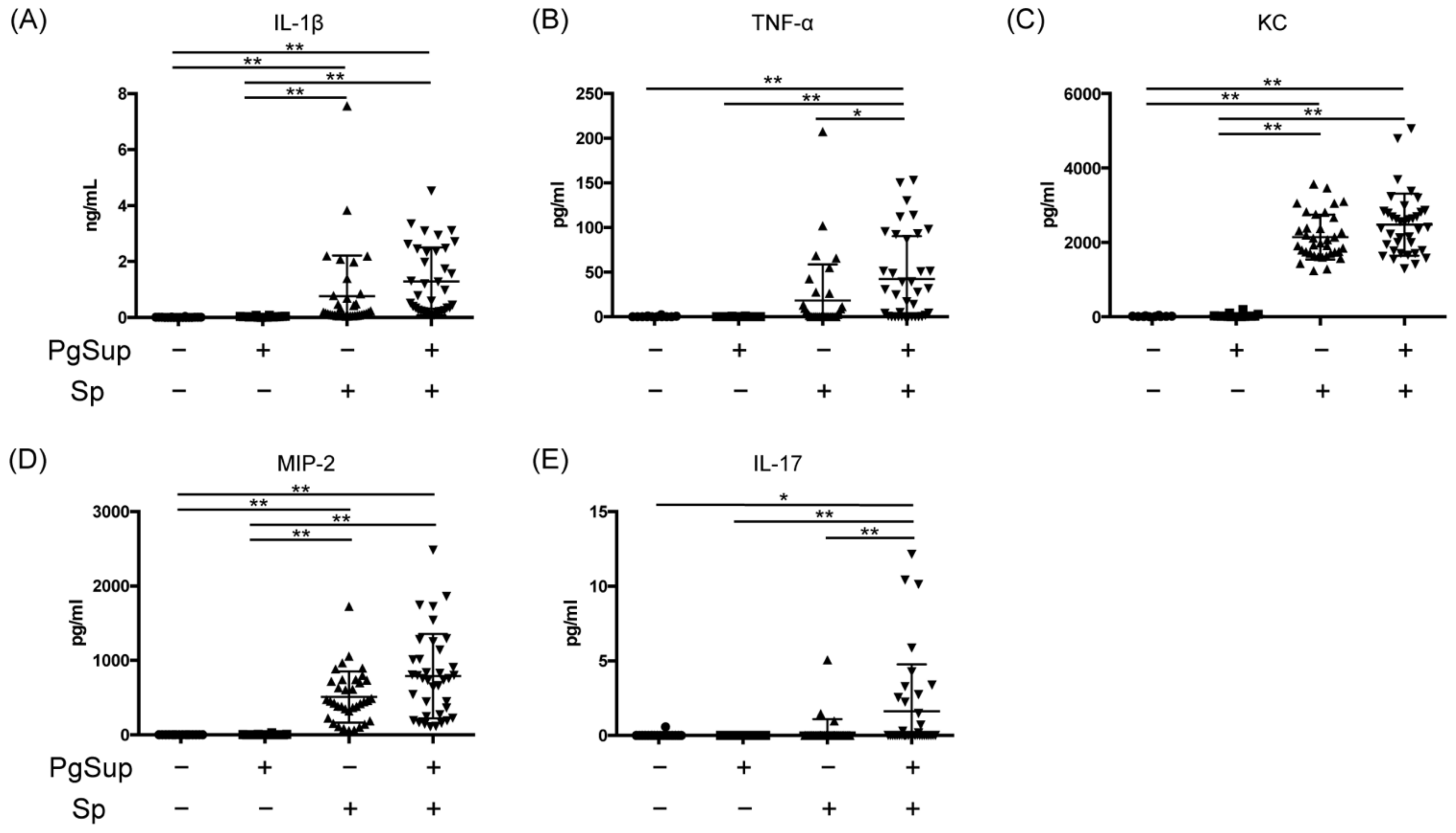
2.4. PgSup Enhanced the Production of Pro-Inflammatory Cytokines and Chemokines in the Lungs of Sp-Infected Mice
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Mice
4.3. Design of Animal Experiments
4.4. Histological Analysis
4.5. Cell Preparation and Flow Cytometric Analysis
4.6. Bacterial Numbers in the Lung
4.7. RNA Extraction and Quantitative Polymerase Chain Reaction (qPCR) Analysis
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, W.S.; Baudouin, S.V.; George, R.C.; Hill, A.T.; Jamieson, C.; Le Jeune, I.; Macfarlane, J.T.; Read, R.C.; Roberts, H.J.; Levy, M.L.; et al. BTS guidelines for the management of community acquired pneumonia in adults: Update 2009. Thorax 2009, 64, 1–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandell, L.A.; Niederman, M.S. Aspiration Pneumonia. N. Engl. J. Med. 2019, 380, 651–663. [Google Scholar] [CrossRef]
- Farooqui, H.; Jit, M.; Heymann, D.L.; Zodpey, S. Burden of Severe Pneumonia, Pneumococcal Pneumonia and Pneumonia Deaths in Indian States: Modelling Based Estimates. PLoS ONE 2015, 10, e0129191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.E.; Restrepo, M.I. New perspectives in aspiration community acquired Pneumonia. Expert Rev. Clin. Pharm. 2019, 12, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Hirsch, C.S. Aspiration pneumonia. Recognizing and managing a potentially growing disorder. Postgrad. Med. 2003, 113, 99–102, 105–106, 111–112. [Google Scholar] [CrossRef]
- Bartlett, J.G.; Gorbach, S.L. The triple threat of aspiration pneumonia. Chest 1975, 68, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Scannapieco, F.A.; Mylotte, J.M. Relationships Between Periodontal Disease and Bacterial Pneumonia. J. Periodontol. 1996, 67, 1114–1122. [Google Scholar] [CrossRef]
- Sjogren, P.; Nilsson, E.; Forsell, M.; Johansson, O.; Hoogstraate, J. A systematic review of the preventive effect of oral hygiene on pneumonia and respiratory tract infection in elderly people in hospitals and nursing homes: Effect estimates and methodological quality of randomized controlled trials. J. Am. Geriatr. Soc. 2008, 56, 2124–2130. [Google Scholar] [CrossRef]
- Azarpazhooh, A.; Leake, J.L. Systematic review of the association between respiratory diseases and oral health. J. Periodontol. 2006, 77, 1465–1482. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cao, Y.; Lin, J.; Ng, L.; Needleman, I.; Walsh, T.; Li, C. Oral care measures for preventing nursing home-acquired pneumonia. Cochrane Database Syst. Rev. 2018, 9, CD012416. [Google Scholar] [CrossRef]
- Okuda, M.; Kaneko, Y.; Ichinohe, T.; Ishihara, K.; Okuda, K. Reduction of potential respiratory pathogens by oral hygienic treatment in patients undergoing endotracheal anesthesia. J. Anesth. 2003, 17, 84–91. [Google Scholar] [CrossRef]
- Bogaert, D.; de Groot, R.; Hermans, P.W. Streptococcus pneumoniae colonisation: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Parker, D.; Prince, A. Innate immunity in the respiratory epithelium. Am. J. Respir. Cell Mol. Biol. 2011, 45, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Mengesha, B.G.; Conti, H.R. The Role of IL-17 in Protection against Mucosal Candida Infections. J. Fungi 2017, 3, 52. [Google Scholar] [CrossRef]
- Liu, R.; Lauridsen, H.M.; Amezquita, R.A.; Pierce, R.W.; Jane-Wit, D.; Fang, C.; Pellowe, A.S.; Kirkiles-Smith, N.C.; Gonzalez, A.L.; Pober, J.S. IL-17 Promotes Neutrophil-Mediated Immunity by Activating Microvascular Pericytes and Not Endothelium. J. Immunol. 2016, 197, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Riviere, G.R.; Smith, K.S.; Tzagaroulaki, E.; Kay, S.L.; Zhu, X.; DeRouen, T.A.; Adams, D.F. Periodontal status and detection frequency of bacteria at sites of periodontal health and gingivitis. J. Periodontol. 1996, 67, 109–115. [Google Scholar] [CrossRef]
- Yoshioka, H.; Yoshimura, A.; Kaneko, T.; Golenbock, D.T.; Hara, Y. Analysis of the activity to induce toll-like receptor (TLR)2- and TLR4-mediated stimulation of supragingival plaque. J. Periodontol. 2008, 79, 920–928. [Google Scholar] [CrossRef]
- Nayak, B.N.; Kaur, G.; Buttar, H.S. TNF-alpha modulation by natural bioactive molecules in mouse RAW 264.7 macrophage cells. J. Complementary Integr. Med. 2016, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sawant, K.V.; Sepuru, K.M.; Lowry, E.; Penaranda, B.; Frevert, C.W.; Garofalo, R.P.; Rajarathnam, K. Neutrophil recruitment by chemokines Cxcl1/KC and Cxcl2/MIP2: Role of Cxcr2 activation and glycosaminoglycan interactions. J. Leukoc. Biol. 2021, 109, 777–791. [Google Scholar] [CrossRef]
- De Filippo, K.; Dudeck, A.; Hasenberg, M.; Nye, E.; van Rooijen, N.; Hartmann, K.; Gunzer, M.; Roers, A.; Hogg, N. Mast cell and macrophage chemokines CXCL1/CXCL2 control the early stage of neutrophil recruitment during tissue inflammation. Blood 2013, 121, 4930–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Cao, X. Interleukin-17 and its expanding biological functions. Cell Mol. Immunol. 2010, 7, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Stark, M.A.; Huo, Y.; Burcin, T.L.; Morris, M.A.; Olson, T.S.; Ley, K. Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity 2005, 22, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemec, A.; Pavlica, Z.; Nemec-Svete, A.; Erzen, D.; Milutinovic, A.; Petelin, M. Aerosolized clindamycin is superior to aerosolized dexamethasone or clindamycin-dexamethasone combination in the treatment of severe Porphyromonas gingivalis aspiration pneumonia in an experimental murine model. Exp. Lung. Res. 2012, 38, 9–18. [Google Scholar] [CrossRef]
- Pace, C.C.; McCullough, G.H. The association between oral microorgansims and aspiration pneumonia in the institutionalized elderly: Review and recommendations. Dysphagia 2010, 25, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Jung, Y.J.; Miller, D.P.; Perpich, J.D.; Fitzsimonds, Z.R.; Shen, D.; Ohshima, J.; Lamont, R.J. Porphyromonas gingivalis Tyrosine Phosphatase Php1 Promotes Community Development and Pathogenicity. mBio 2019, 10, e02004-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enersen, M.; Nakano, K.; Amano, A. Porphyromonas gingivalis fimbriae. J. Oral Microbiol. 2013, 5, 20265. [Google Scholar] [CrossRef]
- Benedyk, M.; Mydel, P.M.; Delaleu, N.; Plaza, K.; Gawron, K.; Milewska, A.; Maresz, K.; Koziel, J.; Pyrc, K.; Potempa, J. Gingipains: Critical Factors in the Development of Aspiration Pneumonia Caused by Porphyromonas gingivalis. J. Innate Immun. 2016, 8, 185–198. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yoneda, M.; Okamoto, K.; Maeda, K.; Yamamoto, K. Purification and characterization of a novel arginine-specific cysteine proteinase (argingipain) involved in the pathogenesis of periodontal disease from the culture supernatant of Porphyromonas gingivalis. J. Biol. Chem. 1994, 269, 21371–21378. [Google Scholar] [CrossRef]
- Abe, N.; Kadowaki, T.; Okamoto, K.; Nakayama, K.; Ohishi, M.; Yamamoto, K. Biochemical and functional properties of lysine-specific cysteine proteinase (Lys-gingipain) as a virulence factor of Porphyromonas gingivalis in periodontal disease. J. Biochem. 1998, 123, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Lantz, M.S.; Allen, R.D.; Duck, L.W.; Blume, J.L.; Switalski, L.M.; Hook, M. Identification of Porphyromonas gingivalis components that mediate its interactions with fibronectin. J. Bacteriol. 1991, 173, 4263–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, A.; Naito, M.; Imamura, T.; Potempa, J.; Travis, J.; Nakayama, K.; Takada, H. Dual regulation of interleukin-8 production in human oral epithelial cells upon stimulation with gingipains from Porphyromonas gingivalis. J. Med. Microbiol. 2008, 57, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Takii, R.; Kadowaki, T.; Baba, A.; Tsukuba, T.; Yamamoto, K. A functional virulence complex composed of gingipains, adhesins, and lipopolysaccharide shows high affinity to host cells and matrix proteins and escapes recognition by host immune systems. Infect. Immun. 2005, 73, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Belibasakis, G.N. Doxycycline inhibits TREM-1 induction by Porphyromonas gingivalis. FEMS Immunol. Med. Microbiol. 2012, 66, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Haleem, K.S.; Ali, Y.M.; Yesilkaya, H.; Kohler, T.; Hammerschmidt, S.; Andrew, P.W.; Schwaeble, W.J.; Lynch, N.J. The Pneumococcal Surface Proteins PspA and PspC Sequester Host C4-Binding Protein To Inactivate Complement C4b on the Bacterial Surface. Infect. Immun. 2019, 87, e00742-18. [Google Scholar] [CrossRef] [Green Version]
- Ishizuka, S.; Yamaya, M.; Suzuki, T.; Takahashi, H.; Ida, S.; Sasaki, T.; Inoue, D.; Sekizawa, K.; Nishimura, H.; Sasaki, H. Effects of rhinovirus infection on the adherence of Streptococcus pneumoniae to cultured human airway epithelial cells. J. Infect. Dis. 2003, 188, 1928–1939. [Google Scholar]
- Kamio, N.; Hayata, M.; Tamura, M.; Tanaka, H.; Imai, K. Porphyromonas gingivalis enhances pneumococcal adhesion to human alveolar epithelial cells by increasing expression of host platelet-activating factor receptor. FEBS Lett. 2021, 595, 1604–1612. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Nishiyama, S.; Nishikawa, K.; Kadowaki, T.; Yamamoto, K.; Noguchi, T.; Yoshimura, F. A novel type of two-component regulatory system affecting gingipains in Porphyromonas gingivalis. Microbiol. Immunol. 2003, 47, 849–858. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Whole Cells (Cells) | Neutrophils (Cells) | Macrophages (Cells) | Neutrophils (%) | Macrophages (%) |
|---|---|---|---|---|---|
| Control | 6.15 ± 0.78 × 106 | 3.51 ± 1.07 × 105 b | 0.85 ± 0.04 × 106 d | 5.62 ± 1.16 | 14.06 ± 1.71 |
| PgSup | 5.90 ± 0.67 × 106 a | 3.99 ± 1.65 × 105 c | 0.90 ± 0.08 ×106 e,f | 6.67 ± 2.14 | 15.31 ± 1.05 |
| Sp | 7.98 ± 1.11 × 106 a | 5.67 ± 2.67 × 105 | 1.52 ± 0.29 × 106 e | 7.38 ± 3.94 | 18.95 ± 1.99 |
| Sp + PgSup | 7.78 ± 1.61 × 106 | 8.09 ± 1.65 × 105 b,c | 1.32 ± 0.17 × 106 d,f | 10.41 ± 0.58 | 17.19 ± 1.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okabe, T.; Kamiya, Y.; Kikuchi, T.; Goto, H.; Umemura, M.; Suzuki, Y.; Sugita, Y.; Naiki, Y.; Hasegawa, Y.; Hayashi, J.-i.; et al. Porphyromonas gingivalis Components/Secretions Synergistically Enhance Pneumonia Caused by Streptococcus pneumoniae in Mice. Int. J. Mol. Sci. 2021, 22, 12704. https://doi.org/10.3390/ijms222312704
Okabe T, Kamiya Y, Kikuchi T, Goto H, Umemura M, Suzuki Y, Sugita Y, Naiki Y, Hasegawa Y, Hayashi J-i, et al. Porphyromonas gingivalis Components/Secretions Synergistically Enhance Pneumonia Caused by Streptococcus pneumoniae in Mice. International Journal of Molecular Sciences. 2021; 22(23):12704. https://doi.org/10.3390/ijms222312704
Chicago/Turabian StyleOkabe, Teppei, Yosuke Kamiya, Takeshi Kikuchi, Hisashi Goto, Masayuki Umemura, Yuki Suzuki, Yoshihiko Sugita, Yoshikazu Naiki, Yoshiaki Hasegawa, Jun-ichiro Hayashi, and et al. 2021. "Porphyromonas gingivalis Components/Secretions Synergistically Enhance Pneumonia Caused by Streptococcus pneumoniae in Mice" International Journal of Molecular Sciences 22, no. 23: 12704. https://doi.org/10.3390/ijms222312704
APA StyleOkabe, T., Kamiya, Y., Kikuchi, T., Goto, H., Umemura, M., Suzuki, Y., Sugita, Y., Naiki, Y., Hasegawa, Y., Hayashi, J. -i., Kawamura, S., Sawada, N., Takayanagi, Y., Fujimura, T., Higuchi, N., & Mitani, A. (2021). Porphyromonas gingivalis Components/Secretions Synergistically Enhance Pneumonia Caused by Streptococcus pneumoniae in Mice. International Journal of Molecular Sciences, 22(23), 12704. https://doi.org/10.3390/ijms222312704