
Aptamer-Aptamer Chimera for Targeted Delivery and ATP-Responsive Release of Doxorubicin into Cancer Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussions
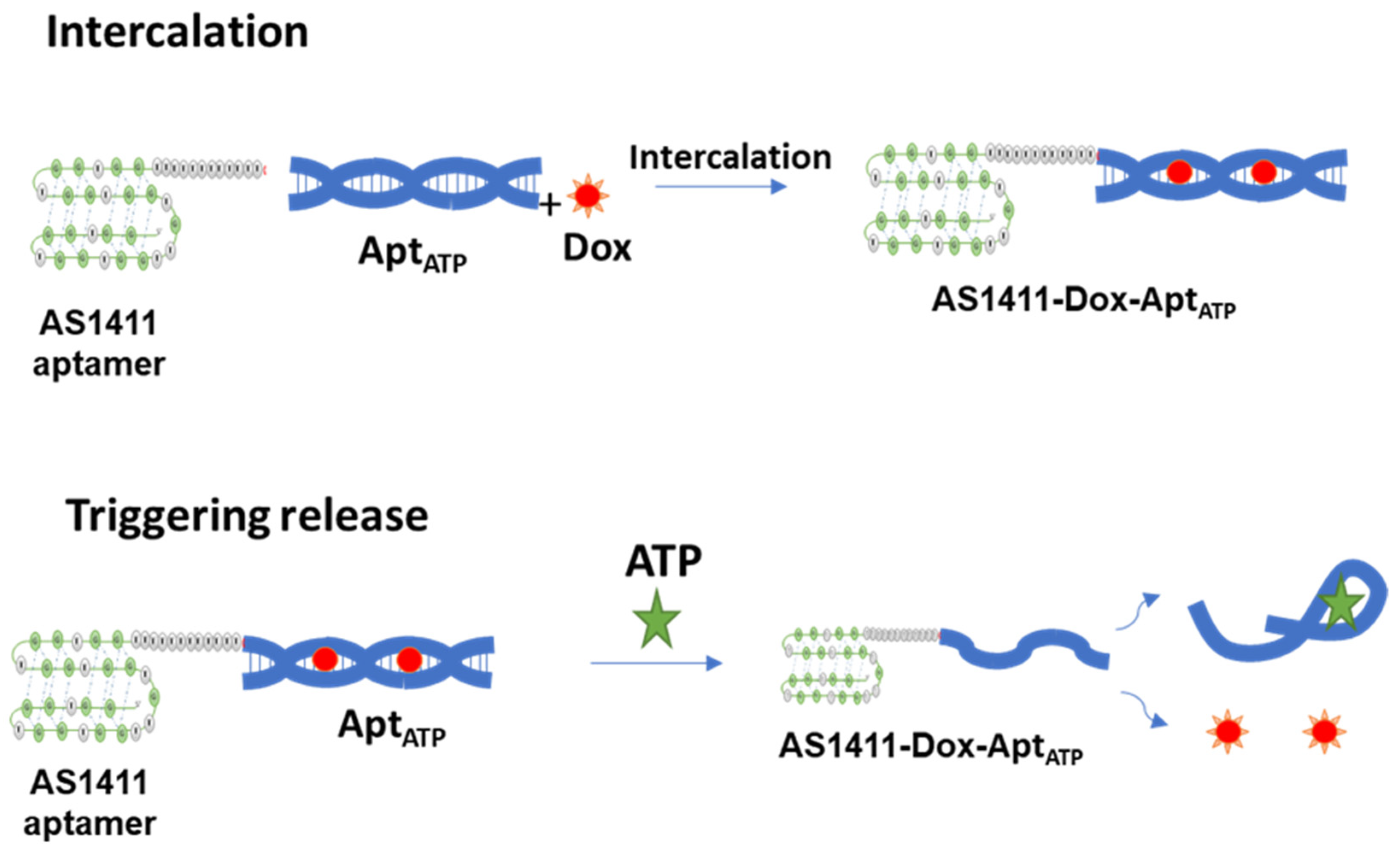
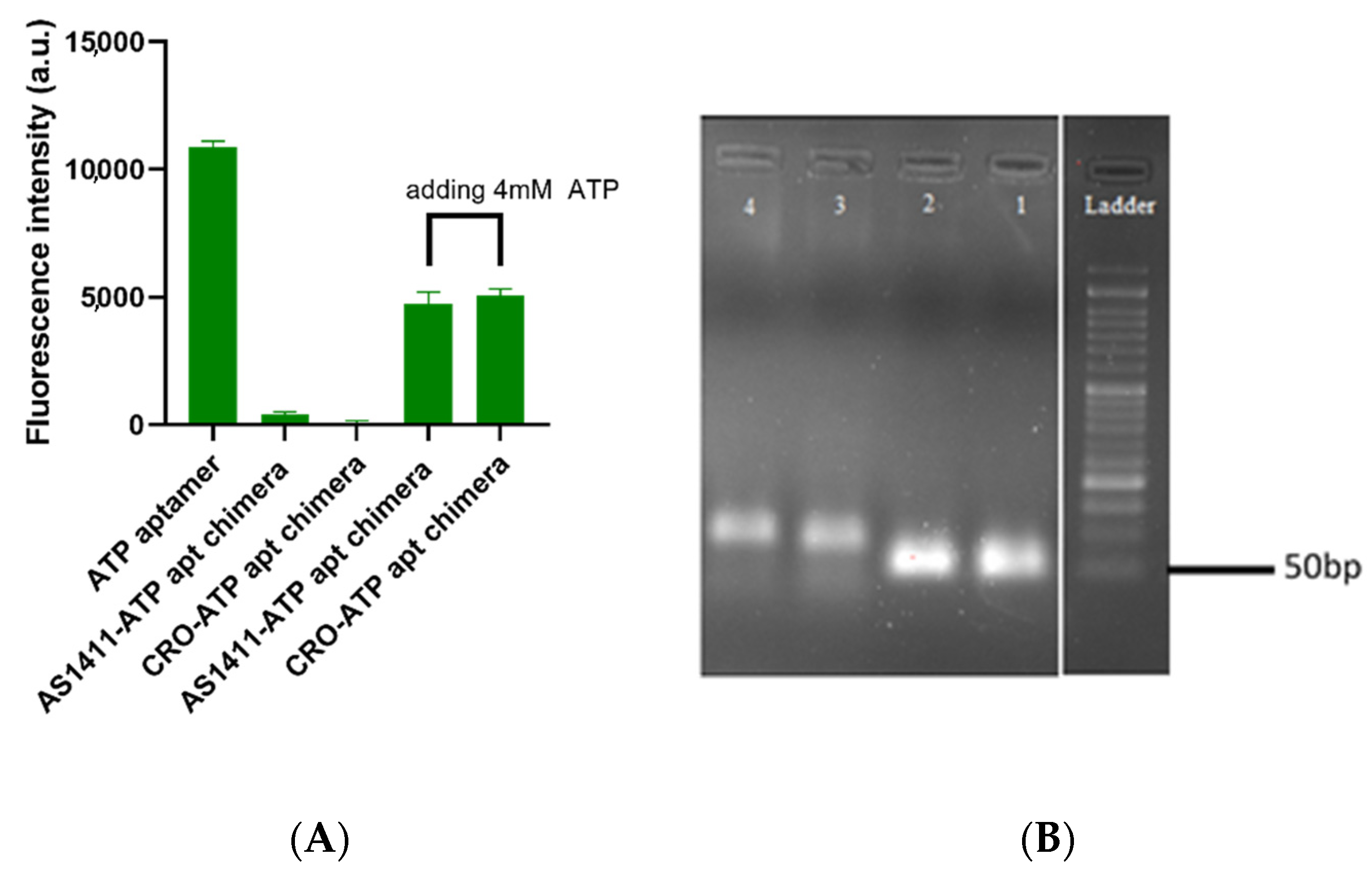
2.1. The Successful Generation of AS1411–ATPapt Chimera Loaded with Doxorubicin
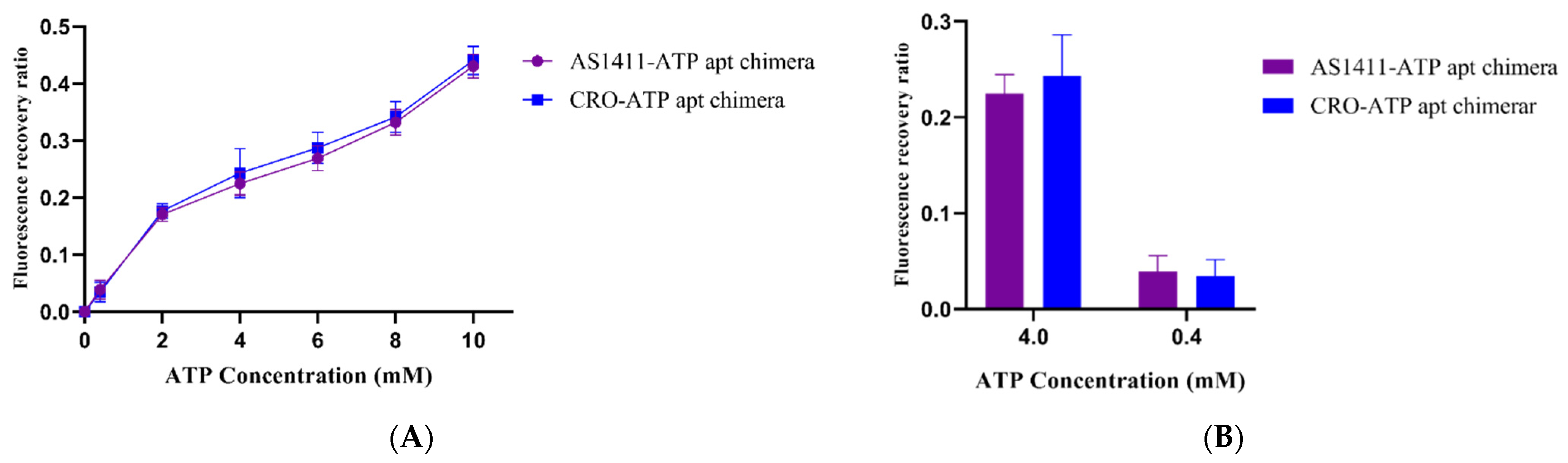
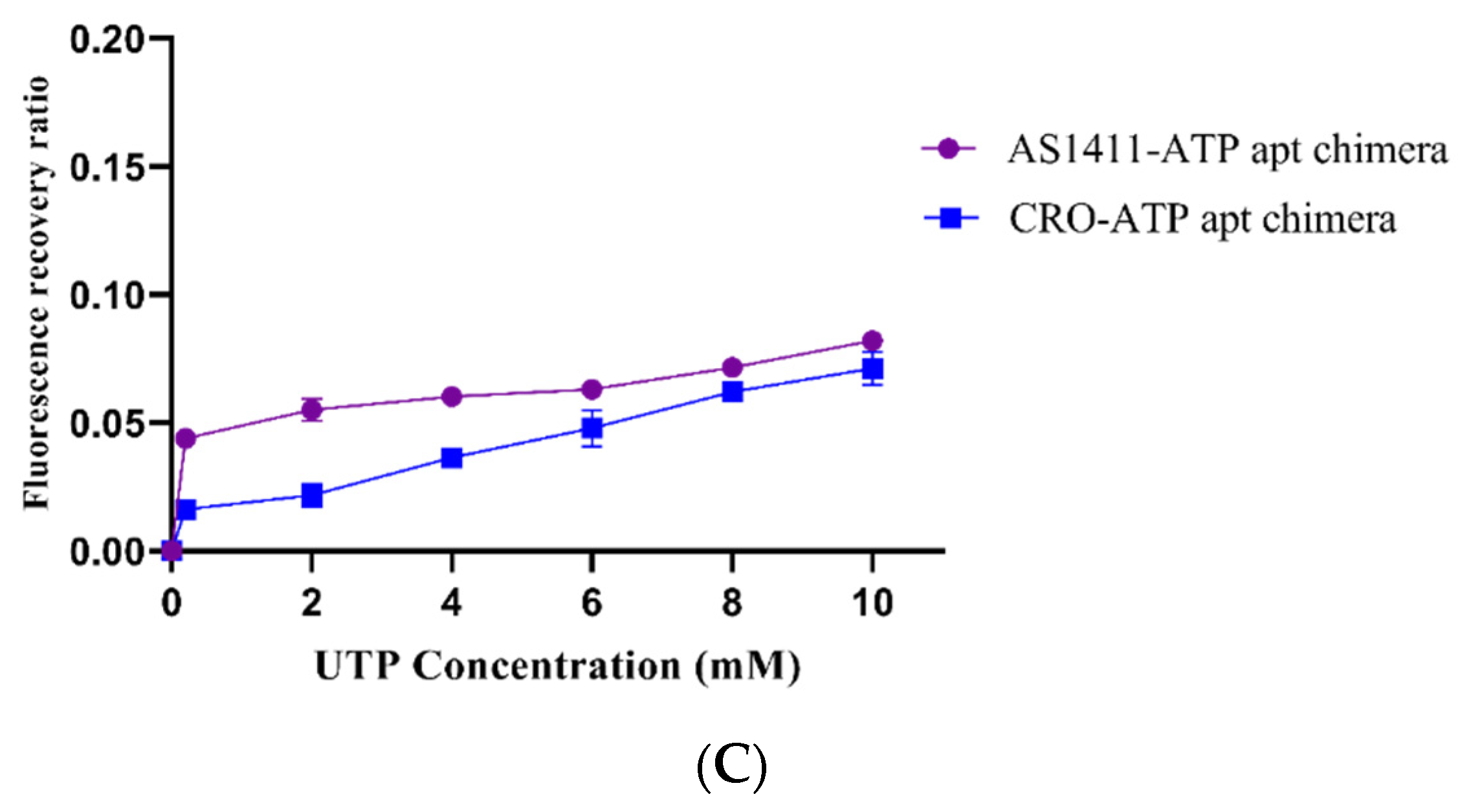
2.2. ATP-Responsiveness for Doxorubicin Release from the Generated Chimeras
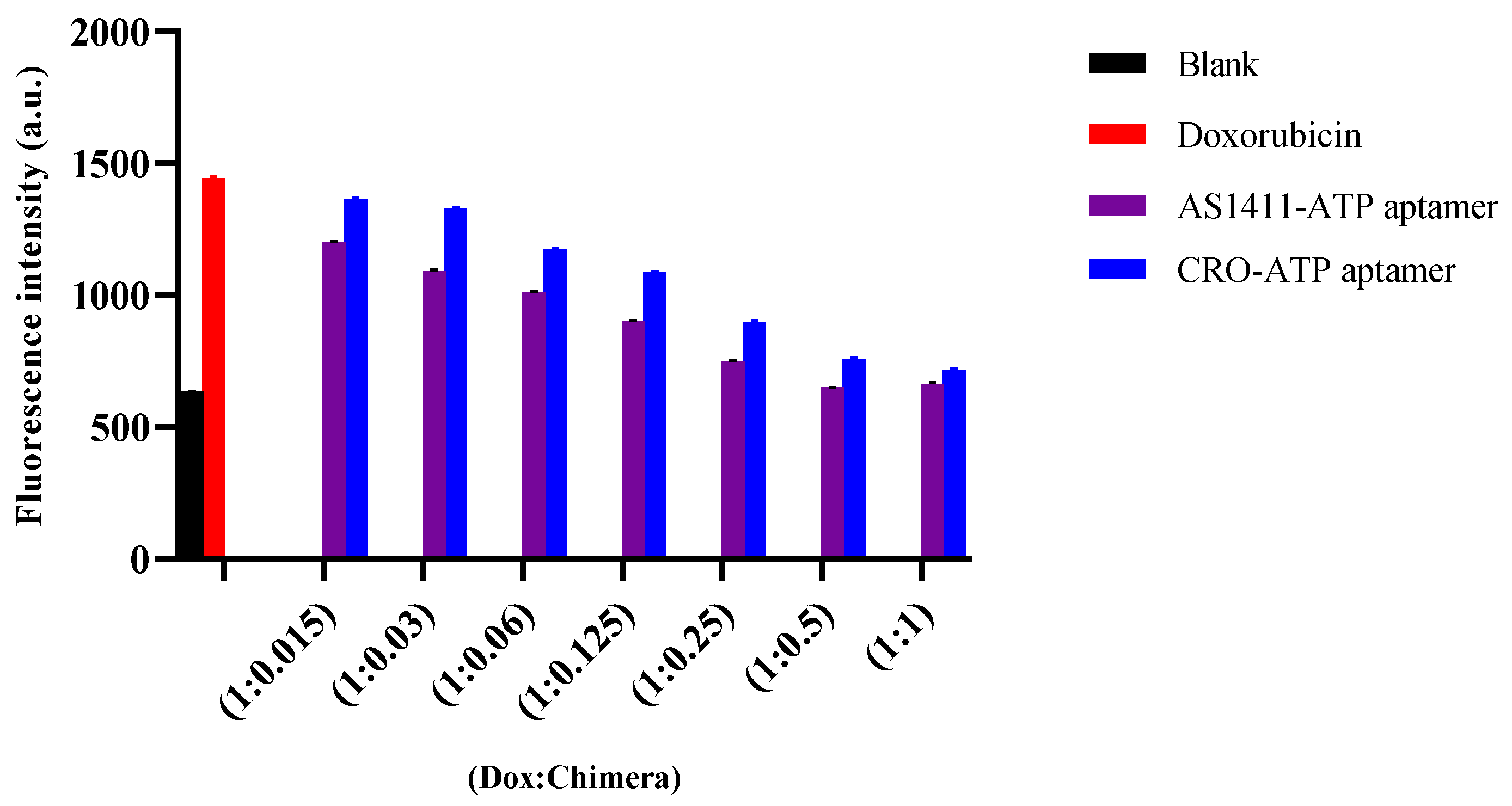
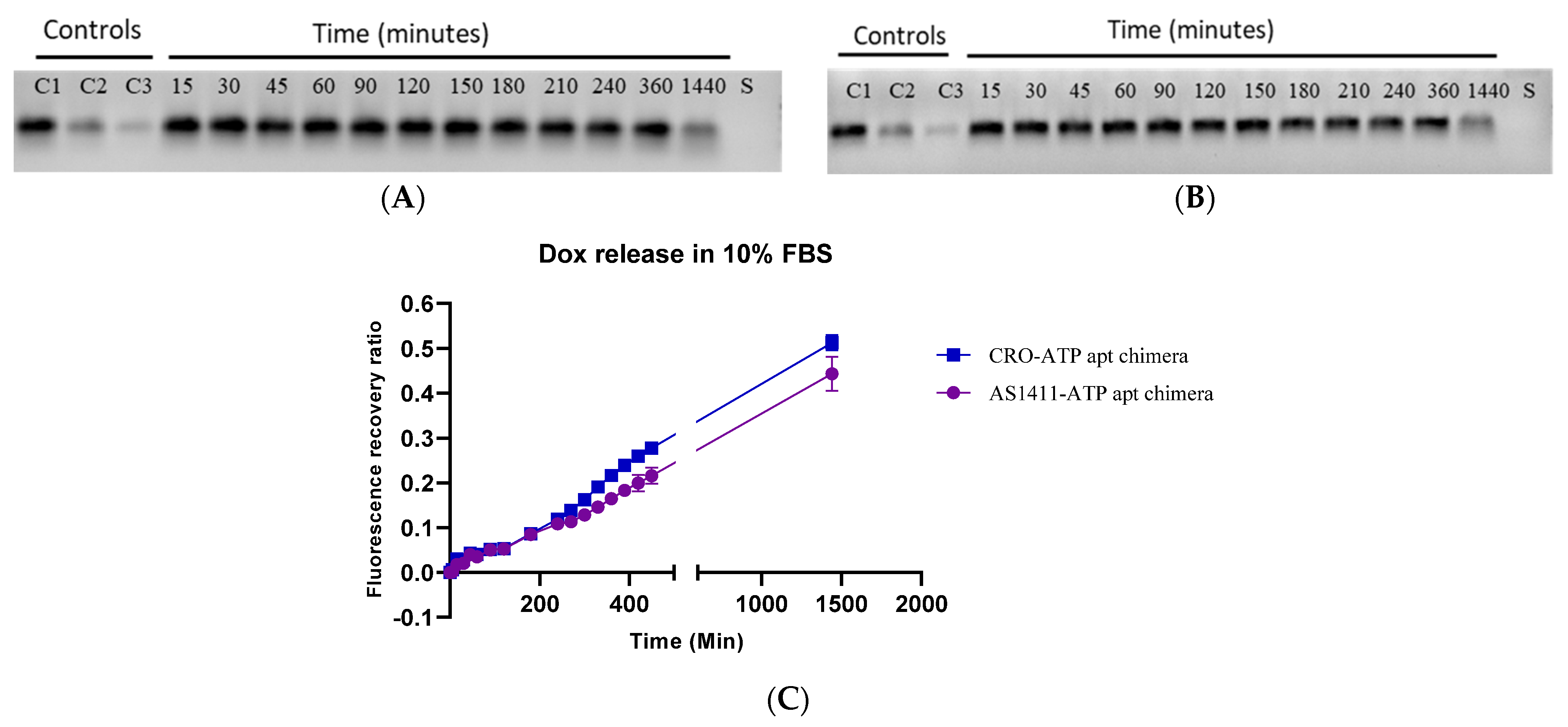
2.3. The Stability of Doxorubicin-Loaded AS1411–ATPapt Chimera in Serum
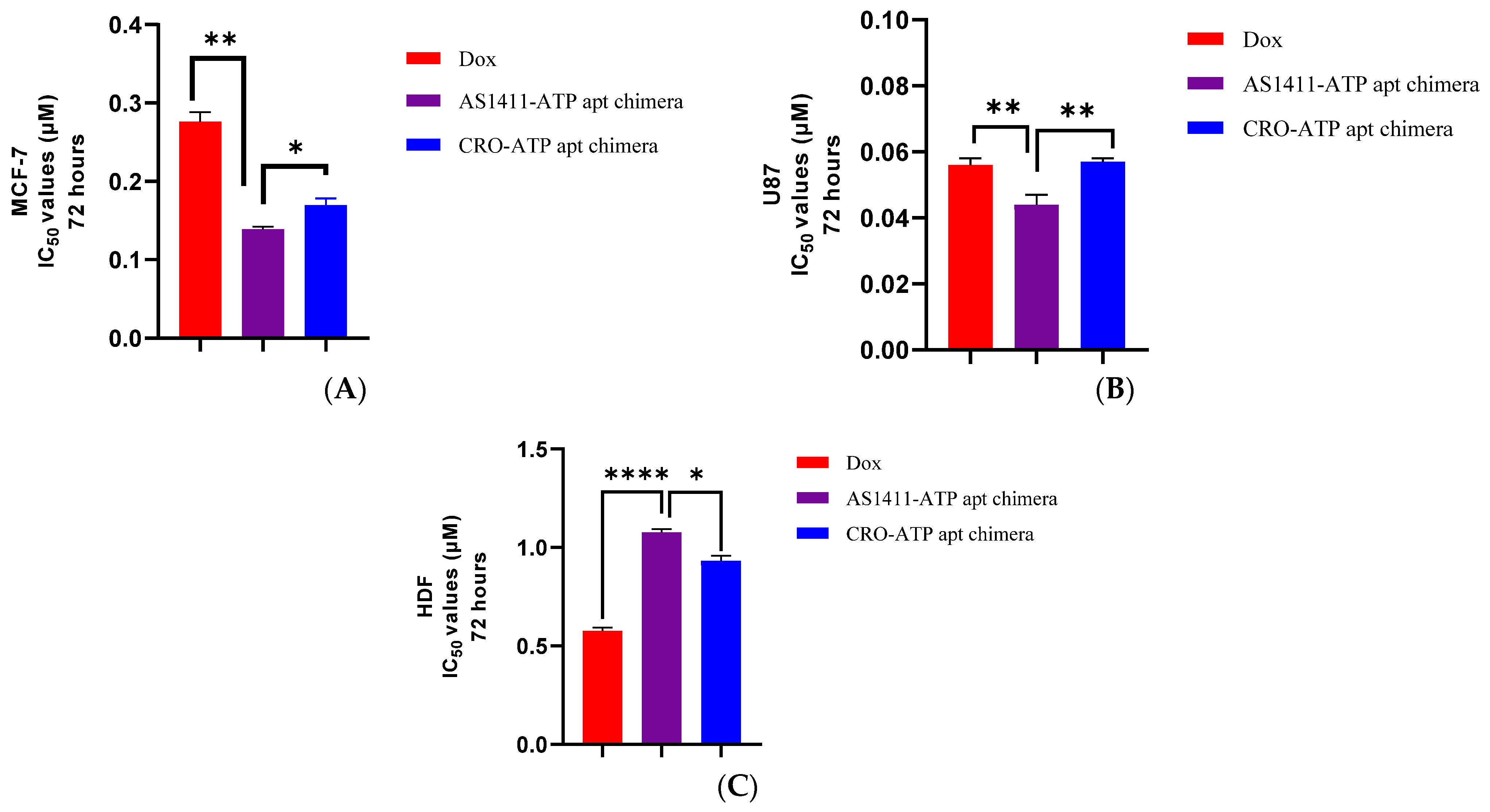
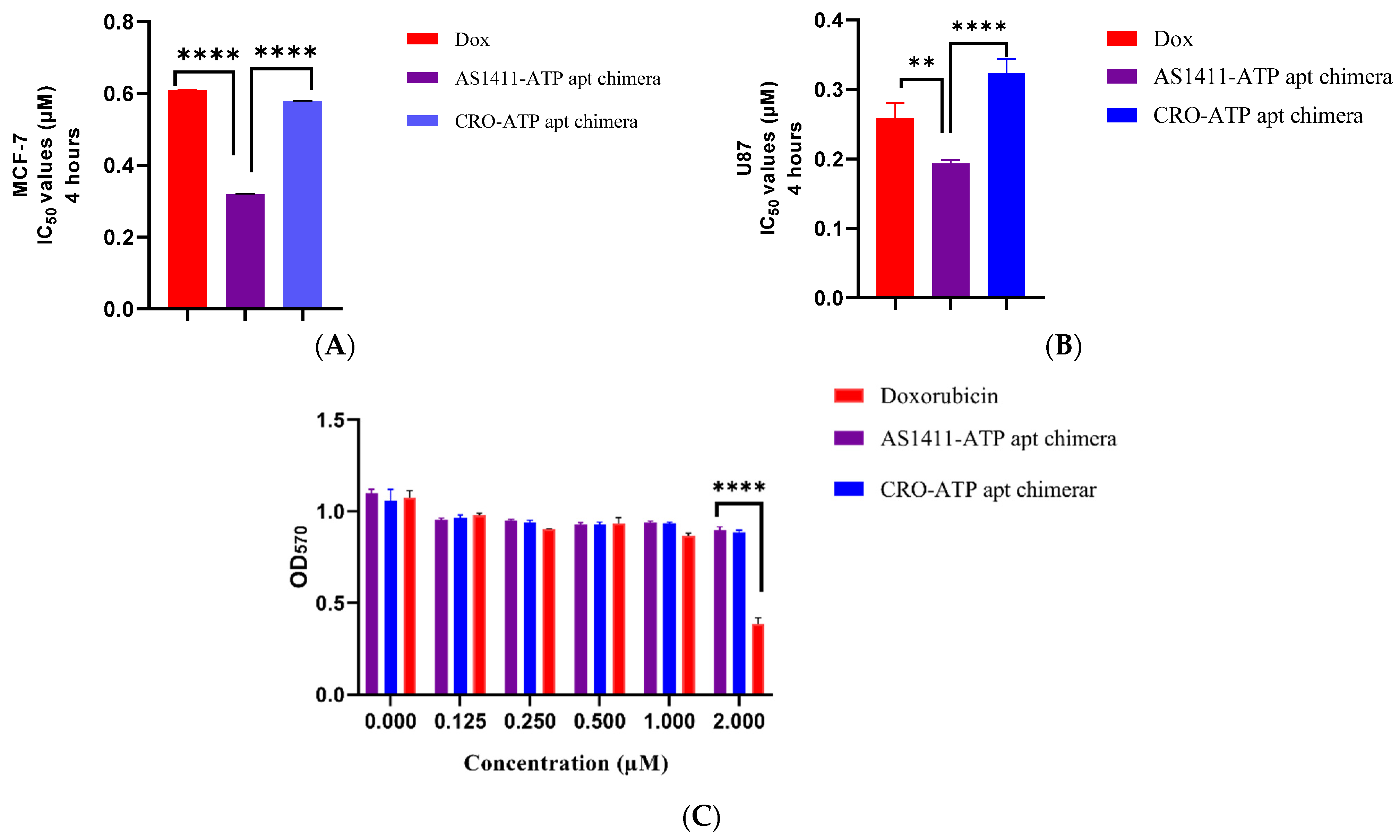
2.4. Cytotoxicity of AS1411–ATPapt Chimera in Cancer Cells
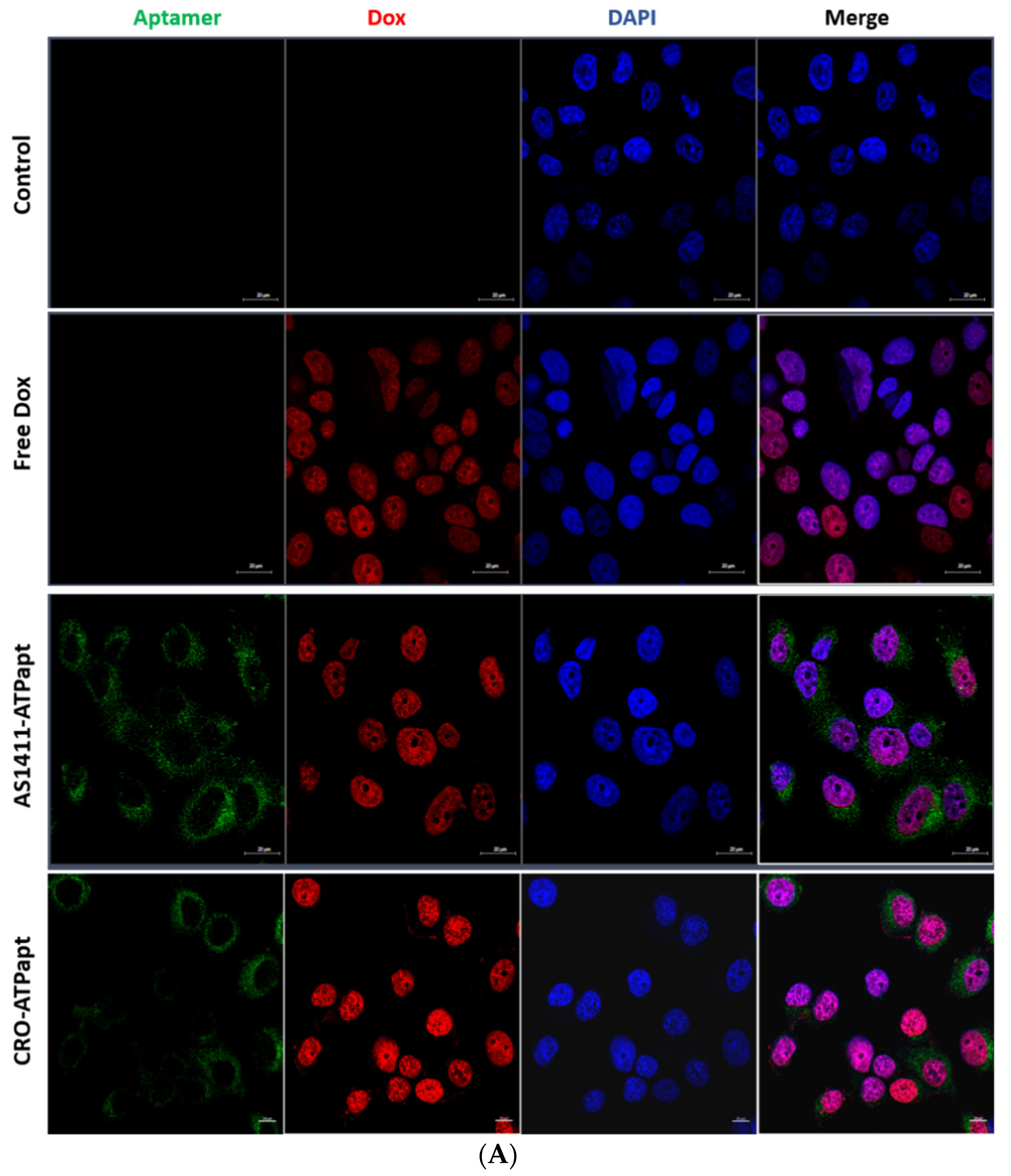
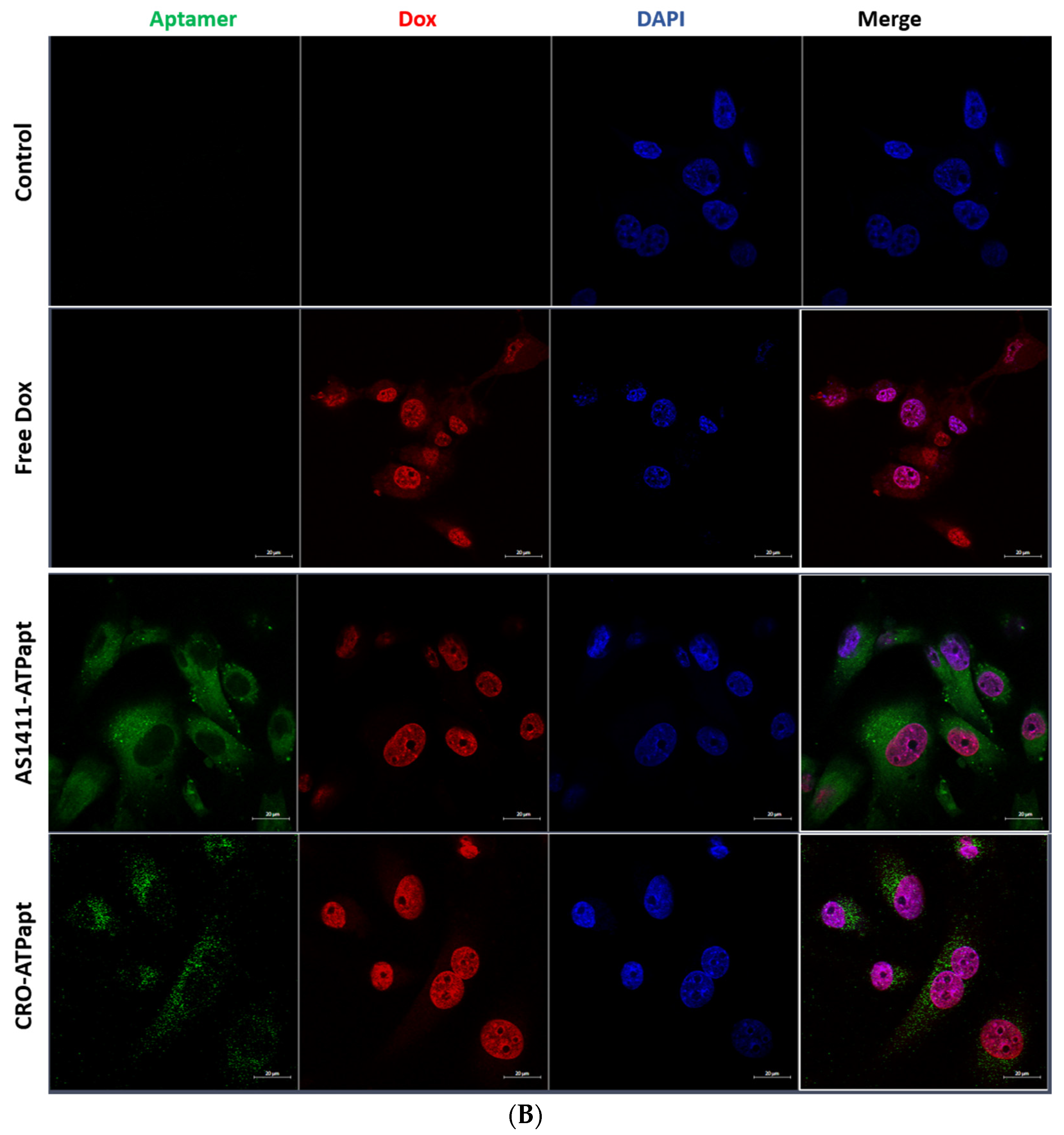
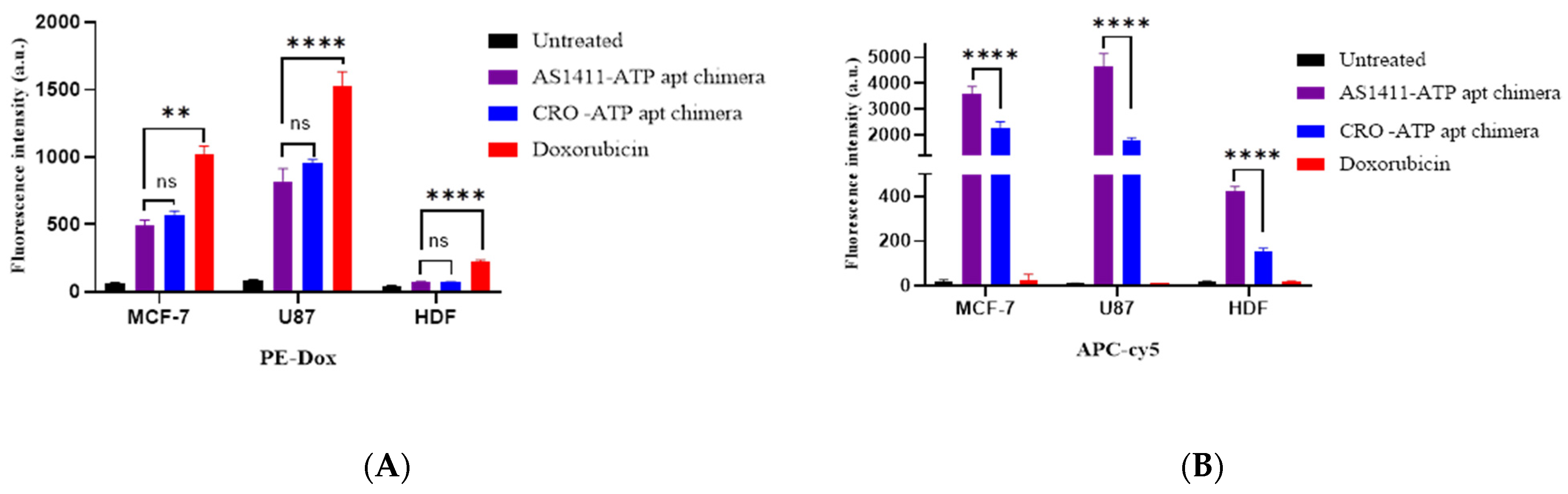
2.5. The AS1411–ATPapt Chimera Is Efficiently Taken up by Cells
3. Materials and Methods
3.1. Chemicals, Reagents, and Cell Lines
3.2. Design and Formation of Aptamer–Aptamer Chimeras
3.3. Evaluate ATP Responsiveness of the Aptamer–Aptamer Chimeras
3.4. Doxorubicin Loading into the Aptamer–Aptamer Chimeras
3.5. Triggering the Release of Doxorubicin from the Aptamer–Aptamer Chimeras
3.6. In Vitro Serum Stability Assay of Chimeras
3.7. Cell Viability (MTT) Assay
3.8. Cellular Uptake Assesment of Free Doxorubicin and Doxorubicin-Loaded Chimeras Using FACS
3.9. Confocal Laser Microscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Lazaro, I.; Mooney, D.J. Obstacles and opportunities in a forward vision for cancer nanomedicine. Nat. Mater. 2021, 20, 1469–1479. [Google Scholar] [CrossRef]
- Rahman, M.; Zaki Ahmad, M.; Kazmi, I.; Akhter, S.; Afzal, M.; Gupta, G.; Ranjan Sinha, V. Emergence of nanomedicine as cancer targeted magic bullets: Recent development and need to address the toxicity apprehension. Curr. Drug Discov. Technol. 2012, 9, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Alshaer, W.; Hillaireau, H.; Fattal, E. Aptamer-guided nanomedicines for anticancer drug delivery. Adv. Drug Deliv. Rev. 2018, 134, 122–137. [Google Scholar] [CrossRef]
- Makwana, V.; Karanjia, J.; Haselhorst, T.; Anoopkumar-Dukie, S.; Rudrawar, S. Liposomal doxorubicin as targeted delivery platform: Current trends in surface functionalization. Int. J. Pharm. 2021, 593, 120117. [Google Scholar] [CrossRef]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; del Pilar Rodriguez-Torres, M.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Xiang, J. Aptamer-Functionalized Nanoparticles in Targeted Delivery and Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 9123. [Google Scholar] [CrossRef] [PubMed]
- Odeh, F.; Nsairat, H.; Alshaer, W.; Ismail, M.A.; Esawi, E.; Qaqish, B.; Al Bawab, A.; Ismail, S.I. Aptamers Chemistry: Chemical Modifications and Conjugation Strategies. Molecules 2019, 25, 3. [Google Scholar] [CrossRef] [Green Version]
- Gray, B.P.; Song, X.; Hsu, D.; Kratschmer, C.; Levy, M.; Barry, A.; Sullenger, B. An Aptamer for Broad Cancer Targeting and Therapy. Cancers 2020, 12, 3217. [Google Scholar] [CrossRef]
- Vindigni, G.; Raniolo, S.; Iacovelli, F.; Unida, V.; Stolfi, C.; Desideri, A.; Biocca, S. AS1411 Aptamer Linked to DNA Nanostructures Diverts Its Traffic Inside Cancer Cells and Improves Its Therapeutic Efficacy. Pharmaceutics 2021, 13, 1671. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yan, Z.; Jin, K.; Pang, Q.; Jiang, T.; Lu, H.; Liu, X.; Pang, Z.; Yu, L.; Jiang, X. Precise glioblastoma targeting by AS1411 aptamer-functionalized poly (l-γ-glutamylglutamine)-paclitaxel nanoconjugates. J. Colloid Interface Sci. 2017, 490, 783–796. [Google Scholar] [CrossRef]
- Soundararajan, S.; Chen, W.; Spicer, E.K.; Courtenay-Luck, N.; Fernandes, D.J. The nucleolin targeting aptamer AS1411 destabilizes Bcl-2 messenger RNA in human breast cancer cells. Cancer Res. 2008, 68, 2358–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Hou, J.; Liu, X.; Guo, Y.; Wu, Y.; Zhang, L.; Yang, Z. Nucleolin-targeting liposomes guided by aptamer AS1411 for the delivery of siRNA for the treatment of malignant melanomas. Biomaterials 2014, 35, 3840–3850. [Google Scholar] [CrossRef]
- Malik, M.T.; O’Toole, M.G.; Casson, L.K.; Thomas, S.D.; Bardi, G.T.; Reyes-Reyes, E.; Ng, C.K.; Kang, K.A.; Bates, P.J. AS1411-conjugated gold nanospheres and their potential for breast cancer therapy. Oncotarget 2015, 6, 22270–22281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, C.L.; Catuogno, S.; Condorelli, G.; Ungaro, P.; De Franciscis, V. Aptamer Chimeras for Therapeutic Delivery: The Challenging Perspectives. Genes 2018, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Zhao, S.; Yu, X.; Huang, S.; Liu, H.Y. Simultaneous targeting of CD44 and EpCAM with a bispecific aptamer effectively inhibits intraperitoneal ovarian cancer growth. Theranostics 2017, 7, 1373–1388. [Google Scholar] [CrossRef]
- Macdonald, J.; Henri, J.; Goodman, L.; Xiang, D.; Duan, W.; Shigdar, S. Development of a Bifunctional Aptamer Targeting the Transferrin Receptor and Epithelial Cell Adhesion Molecule (EpCAM) for the Treatment of Brain Cancer Metastases. ACS Chem. Neurosci. 2017, 8, 777–784. [Google Scholar] [CrossRef]
- Dhanasekaran, S.; Chopra, S. Getting a Handle on Smart Drug Delivery Systems—A Comprehensive View of Therapeutic Targeting Strategies. Smart Drug Deliv. Syst. 2016, 1, 31–62. [Google Scholar]
- Chandrasekaran, A.R. Nuclease Resist. Dna Nanostructures. Nat. Rev. Chem. 2021, 5, 225–239. [Google Scholar] [CrossRef]
- Reyes-Reyes, E.M.; Šalipur, F.R.; Shams, M.; Forsthoefel, M.K.; Bates, P.J. Mechanistic studies of anticancer aptamer AS1411 reveal a novel role for nucleolin in regulating Rac1 activation. Mol. Oncol. 2015, 9, 1392–1405. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Reyes, E.M.; Teng, Y.; Bates, P.J. A new paradigm for aptamer therapeutic AS1411 action: Uptake by macropinocytosis and its stimulation by a nucleolin-dependent mechanism. Cancer Res. 2010, 70, 8617–8629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.; Rossi, J.J. Aptamers: Uptake mechanisms and intracellular applications. Adv. Drug Deliv. Rev. 2018, 134, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.T.; Wu, C.Y.; Chung, C.Y.; Hwu, Y.; Cheng, S.H.; Mou, C.Y.; Lo, L.W. Probing the dynamics of doxorubicin-DNA intercalation during the initial activation of apoptosis by fluorescence lifetime imaging microscopy (FLIM). PLoS ONE 2012, 7, e44947. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNA Strand | Sequence |
|---|---|
| ATP aptamer | 5′-ACC TGG GGG AGT ATT GCG GAG GAA GGT-3′ |
| AS1411–cDNA for ATP aptamer | 5′ -GGTGGTGGTGGTTGTGGTGGTGGTGGTTTTTTTTTTTT ACCTTCCTCCGCAATACTCCCCCAGGT-3′ |
| CRO–cDNA for ATP aptamer | 5′ -CCTCCTCCTCCTTCTCCTCCTCCTCCTTTTTTTTTTTTT ACCTTCCTCCGCAATACTCCCCCA GGT -3′ |
| AS1411–cDNA for ATP aptamer—Iowa Black RQ | 5′-GGTGGTGGTGGTTGTGGTGGTGGTGGTTTTTTTTTTTT ACCTTCCTCCGCAATACTCCCCC AGGT-Iowa Black RQ quencher 3′ |
| Cy5–ATP aptamer | 5′-cy5-ACC TGG GGG AGT ATT GCG GAG GAA GGT-3′ |
| CRO–cDNA for ATP aptamer— Iowa Black RQ | 5′-CCTCCTCCTCCTTCTCCTCCTCCTCCTTTTTTTTTTTTT ACCTTCCTCCGCAATACTCCCCCAGGT Iowa Black RQ quencher -3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esawi, E.; Alshaer, W.; Mahmoud, I.S.; Alqudah, D.A.; Azab, B.; Awidi, A. Aptamer-Aptamer Chimera for Targeted Delivery and ATP-Responsive Release of Doxorubicin into Cancer Cells. Int. J. Mol. Sci. 2021, 22, 12940. https://doi.org/10.3390/ijms222312940
Esawi E, Alshaer W, Mahmoud IS, Alqudah DA, Azab B, Awidi A. Aptamer-Aptamer Chimera for Targeted Delivery and ATP-Responsive Release of Doxorubicin into Cancer Cells. International Journal of Molecular Sciences. 2021; 22(23):12940. https://doi.org/10.3390/ijms222312940
Chicago/Turabian StyleEsawi, Ezaldeen, Walhan Alshaer, Ismail Sami Mahmoud, Dana A. Alqudah, Bilal Azab, and Abdalla Awidi. 2021. "Aptamer-Aptamer Chimera for Targeted Delivery and ATP-Responsive Release of Doxorubicin into Cancer Cells" International Journal of Molecular Sciences 22, no. 23: 12940. https://doi.org/10.3390/ijms222312940
APA StyleEsawi, E., Alshaer, W., Mahmoud, I. S., Alqudah, D. A., Azab, B., & Awidi, A. (2021). Aptamer-Aptamer Chimera for Targeted Delivery and ATP-Responsive Release of Doxorubicin into Cancer Cells. International Journal of Molecular Sciences, 22(23), 12940. https://doi.org/10.3390/ijms222312940