
The Communication between Ocular Surface and Nasal Epithelia in 3D Cell Culture Technology for Translational Research: A Narrative Review

 , ,
, ,  and
and 
Abstract
:1. Historical Introduction into the Ocular and Nasal Communication
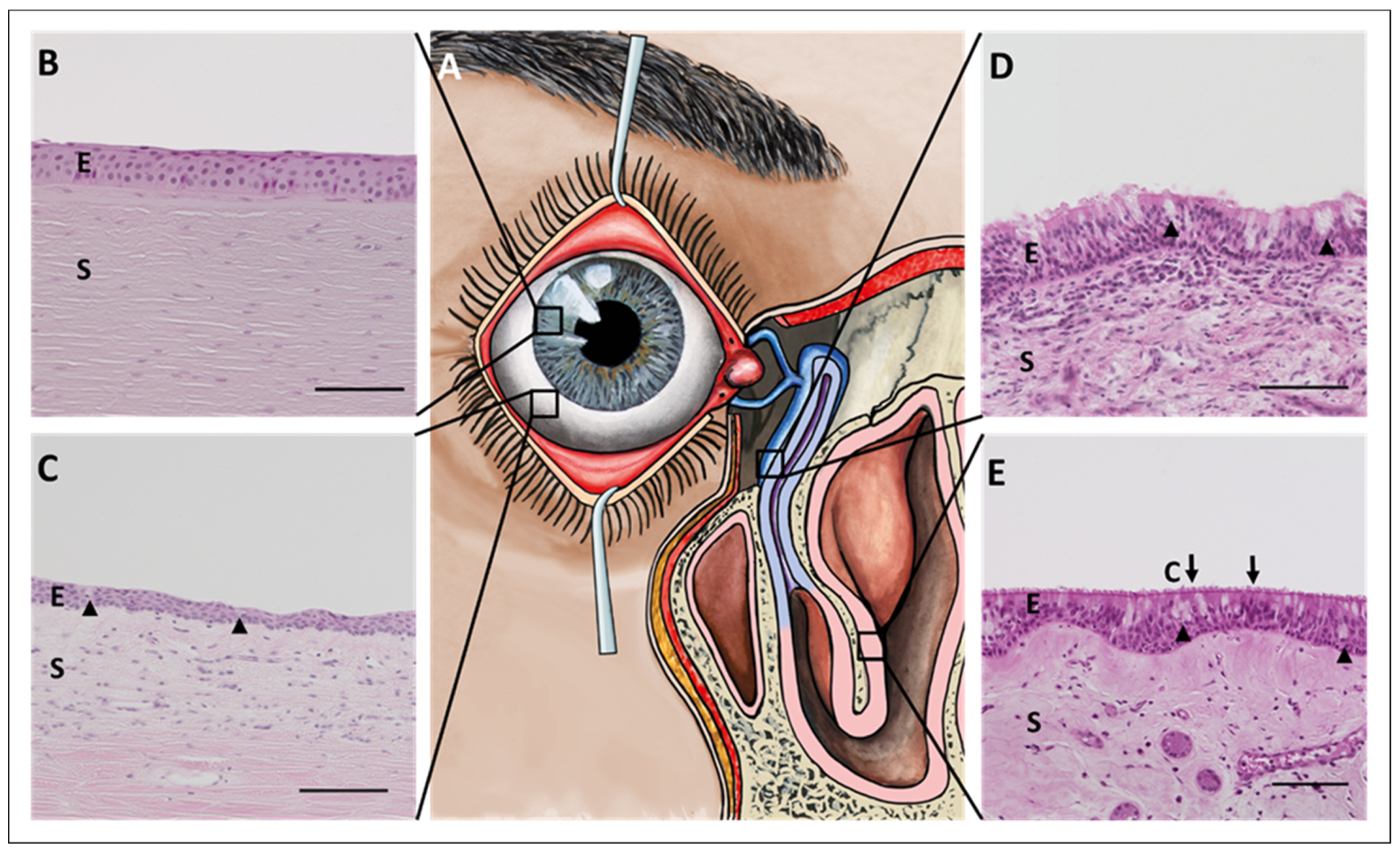
2. Morphology and Physiology of the Ocular Surface Epithelium, Nasolacrimal Ducts, and Nasal Epithelium
2.1. Cornea
- Corneal epithelium
- Bowman´s membrane
- Stroma (with Dua’s layer)
- Descemet´s membrane
- Corneal endothelium
2.2. Conjunctiva
- Bulbar conjunctiva
- Fornical conjunctiva
- Palpebral conjunctiva
2.3. Draining Tear Ducts
2.4. Nasal Cavity
- Cutaneous region
- Olfactory region
- Respiratory region
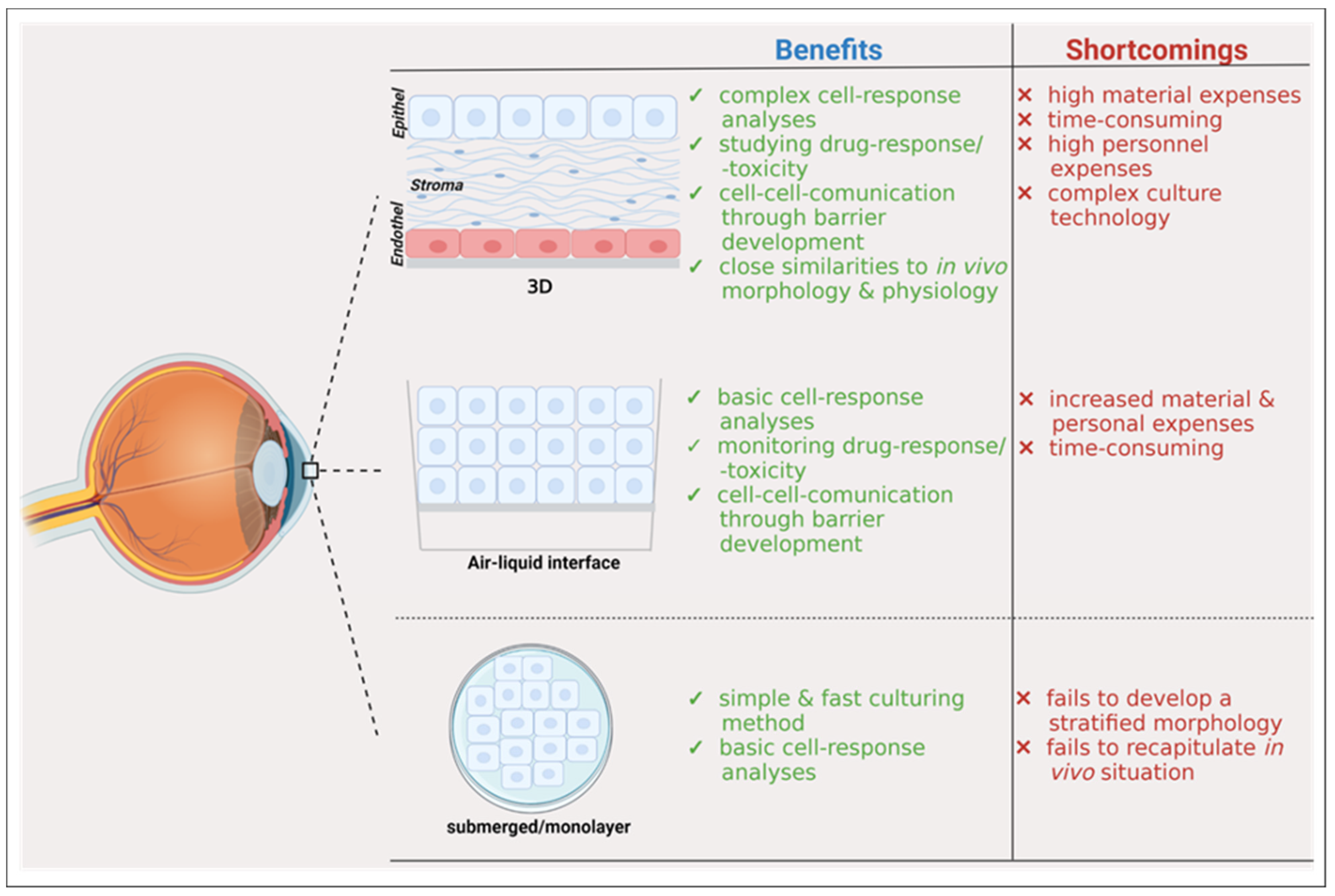
3. Development of 3D Cell Culture Models
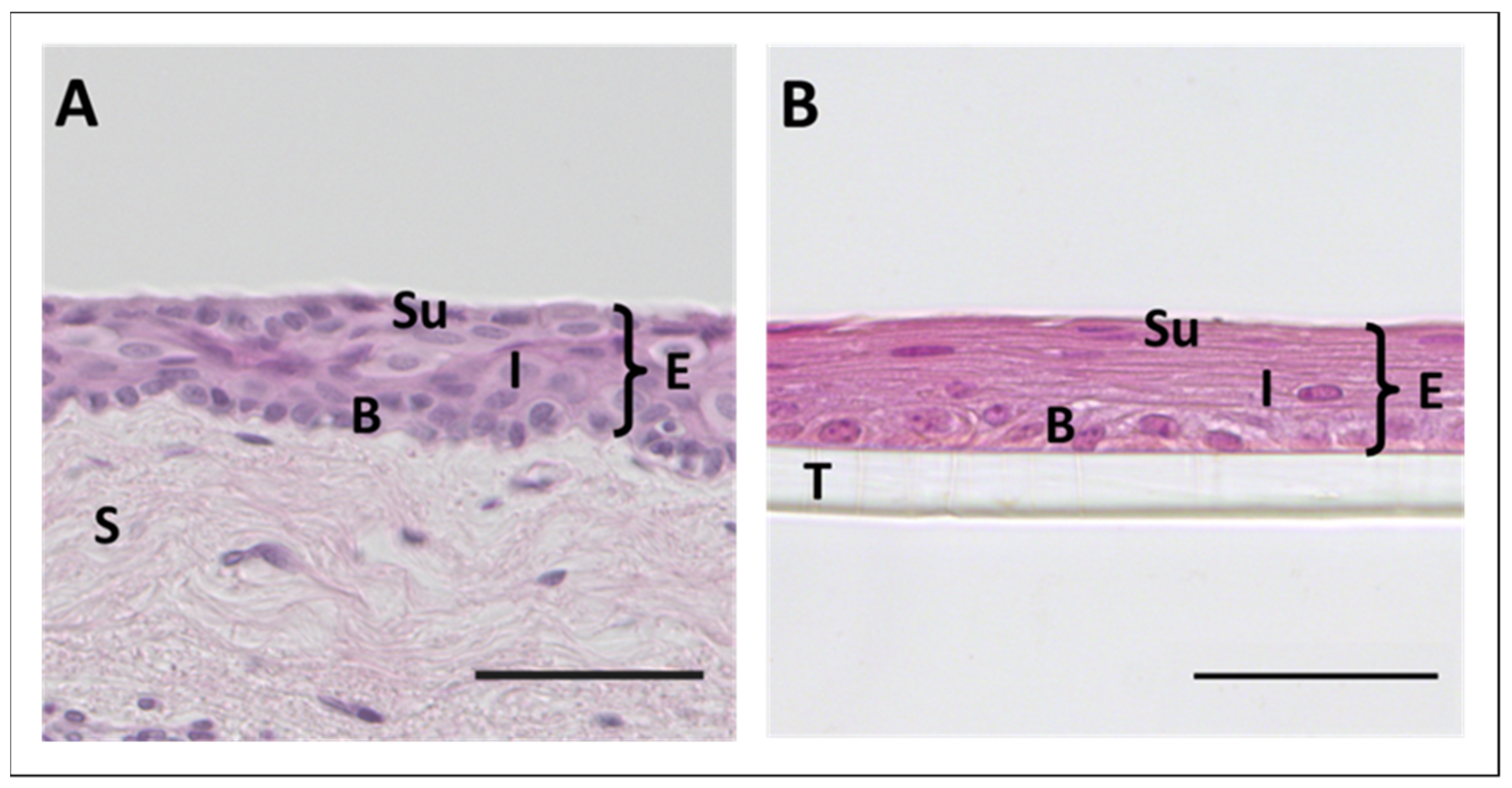
3.1. 3D Cell Culture of Corneal Epithelium
3.2. 3D Cell Culture of Epithelia with Goblet Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Tissue Source | Goblet Cells | Cell Culture Conditions | Ref. |
|---|---|---|---|---|
| 1 | Rabbit-derived conjunctiva | yes |
| [90] |
| 2 | Human-derived conjunctiva |
| [89] | |
| no |
| |||
| few |
| |||
| 3 | Bovine-derived conjunctiva | no |
| [91] |
| 4 | Rabbit-derived conjunctiva | yes |
| [92] |
| 5 | Human-derived conjunctiva | yes |
| [96] |
| 6 | Human-derived conjunctiva |
| [93] | |
| no |
| |||
| no-few |
| |||
| 7 | Human-derived conjunctiva |
| [94] | |
| Few-no |
| |||
|
3.3. Optical Coherence Tomography as a Useful Non-Invasive Technique for Ocular Surface Imaging and Disease Exploring
4. Clinically Relevant Molecular Applications of Conjunctival, Nasolacrimal, and Nasal 3D Cell Culture Models
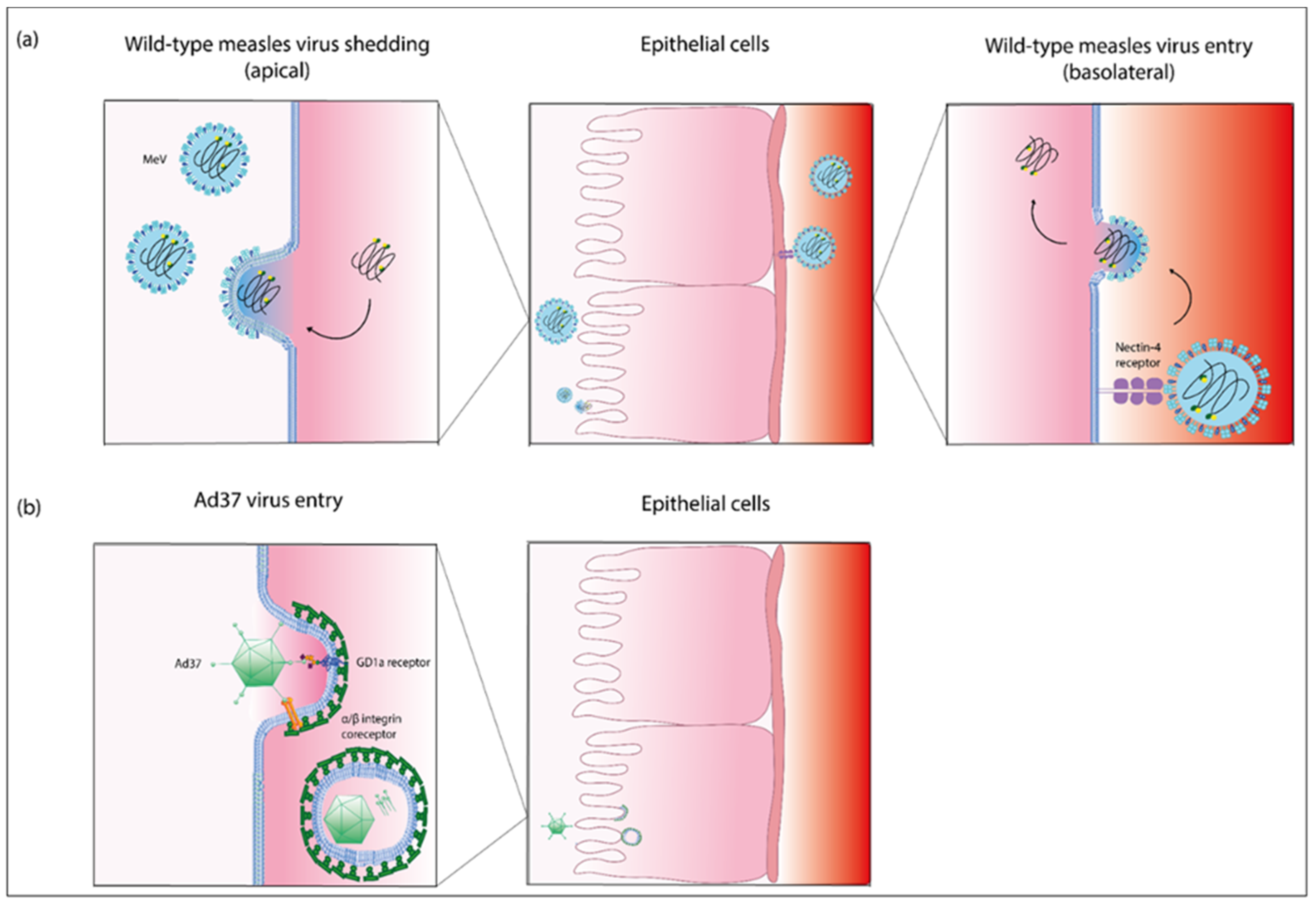
4.1. Pathogenesis of Measles Virus Infection in the Eye and the Nasolacrimal Tract
4.2. Pathogenesis of Adenovirus Infection in the Eye and the Nasolacrimal Tract

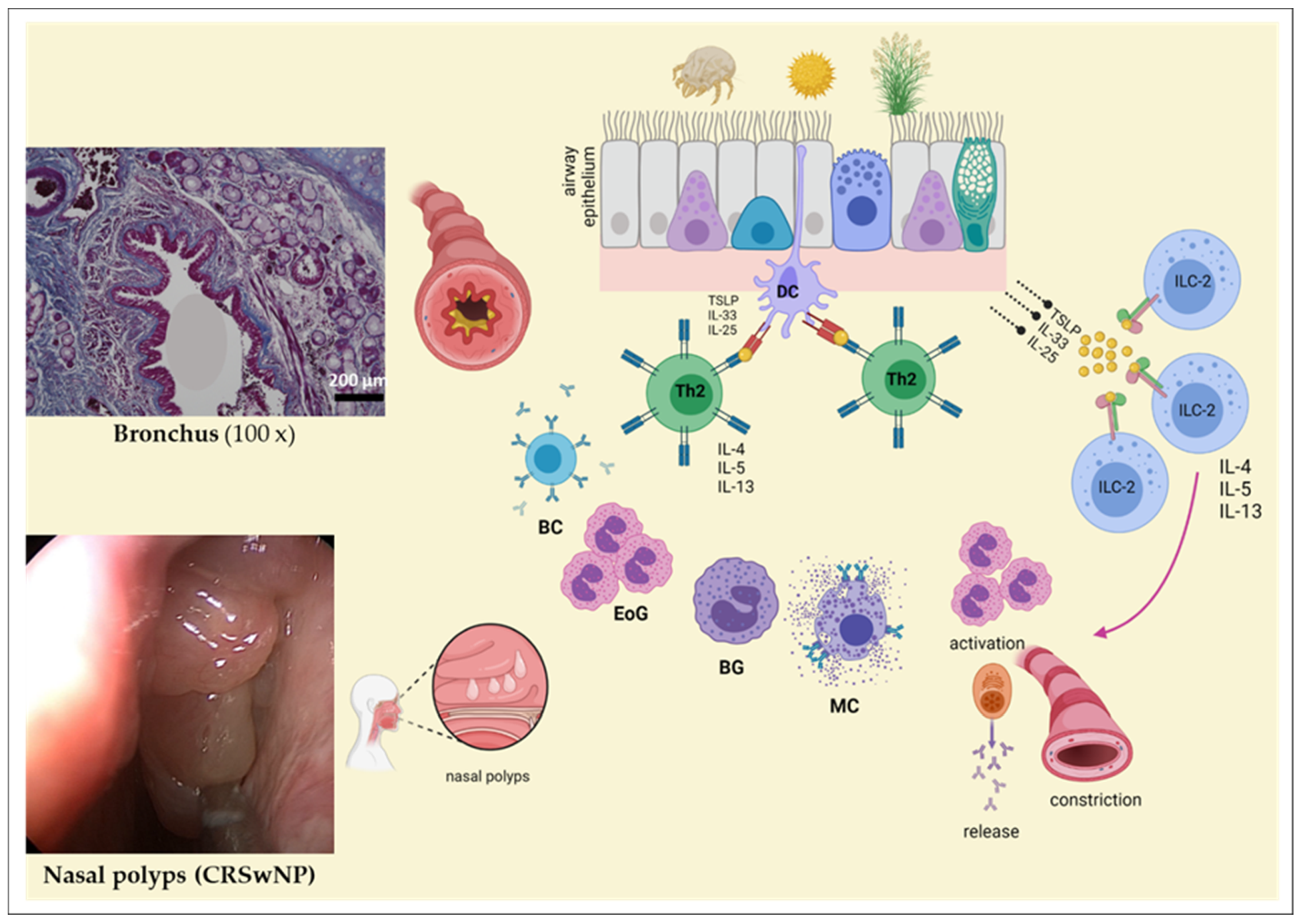
4.3. Allergen Exposure in Ocular and Nasolacrimal Systems

5. Conclusions and Experimental Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Medium | Composition | Ref. | |
|---|---|---|---|
| 1 | CjSC Medium |
| [90] |
| Goblet cell differentiation medium |
| ||
| 2 | CEC Medium |
| [89] |
| Differentiation medium |
| ||
| 3 | Complete growth medium |
| [91] |
| 4 | Growth medium |
| |
| 5 | Bronchial epithelial growth medium |
| [96] |
| Mixture Medium |
| ||
| 6 | Control medium (K) |
| [93] |
| XerumFree medium (XF) |
| ||
| Keratinocyte serum Free medium (SFM) |
| ||
| 7 | Epithelial cell culture medium |
| [94] |
References and Notes
- Available online: https://www.tagesspiegel.de/gesellschaft/panorama/emotionen-traenen-sind-echt-weinen-kann-man-lernen/1137512.html. (accessed on 10 September 2021).
- Messmer, E.M. Emotionale Tränen. Der Ophthalmol. 2009, 106, 593–602. [Google Scholar] [CrossRef]
- Paulsen, F. The human nasolacrimal ducts. Adv. Anat. Embryol. Cell Biol. 2003, 170, 1–106. [Google Scholar]
- Paulsen, F.; Garreis, F.; Schicht, M.; Brauer, L.; Ali, M.J.; Sel, S. Anatomy and physiology of the nasolacrimal ducts. HNO 2016, 64, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Leone, G. In Altero de Musculis Palpebrarum Atque Oculorum Motibus Deservientibus, Accurate Disseritur. Anatomici libri II, Ticini.
- De Ductu Salivali Novo, Saliva, Ductibus Oculorum Aquosis et Muore Oculi Aqueo. In Quibus Varia Oris Oculorium et Narium Vasa Discributu, Novique Salivae Lacymae et Muci Fontis Deteguntur et Novum Norilissimi Bilsii des Lymphae Notu et usu Comentrum Examinatur et Rehictur; Obervationes Anatomicae: Leyden, The Netherlands, 1662.
- Descriptio Anatomica Oculi Humani. Göttingen, Germany, 1755.
- Die bedeutende Publikation von Johann Gottfried Zinn (1727–1759)“Descriptio anatomica oculi humani” (1755). Klin. Monbl. Augenheilkund. 2021, 238, 1018–1020. [CrossRef] [PubMed]
- Ali, M.J.; Zetzsche, M.; Scholz, M.; Hahn, D.; Gaffling, S.; Heichel, J.; Hammer, C.M.; Brauer, L.; Paulsen, F. New insights into the lacrimal pump. Ocul. Surf. 2020, 18, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.D.P.; Argueso, P.; Georgiev, G.A.; Holopainen, J.M.; Laurie, G.W.; Millar, T.J.; Papas, E.B.; Rolland, J.P.; Schmidt, T.A.; Stahl, U.; et al. TFOS DEWS II Tear Film Report. Ocul. Surf. 2017, 15, 366–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Pagano, G.; Pallardo, F.V.; Lyakhovich, A.; Tiano, L.; Trifuoggi, M. Mitigating the pro-oxidant state and melanogenesis of Retinitis pigmentosa: By counteracting mitochondrial dysfunction. Cell Mol. Life Sci. 2021, 78, 7491–7503. [Google Scholar] [CrossRef] [PubMed]
- Bielmeier, C.B.; Roth, S.; Schmitt, S.I.; Boneva, S.K.; Schlecht, A.; Vallon, M.; Tamm, E.R.; Ergun, S.; Neueder, A.; Braunger, B.M. Transcriptional Profiling Identifies Upregulation of Neuroprotective Pathways in Retinitis Pigmentosa. Int. J. Mol. Sci. 2021, 22, 6307. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; Abdalla, E.M.; Scimone, C.; Alibrandi, S.; Rinaldi, C.; Nabil, K.M.; D’Angelo, R.; Sidoti, A. Impairments of Photoreceptor Outer Segments Renewal and Phototransduction Due to a Peripherin Rare Haplotype Variant: Insights from Molecular Modeling. Int. J. Mol. Sci. 2021, 22, 3484. [Google Scholar] [CrossRef] [PubMed]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract. Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Asbell, P.; Brocks, D. Cornea Overview. 2010.
- Hamrah, P.; Sahin, A. Limbus and corneal epithelium. In Ocular Surface Disease: Cornea, Conjunctiva and Tear Film; Elsevier: Amsterdam, The Netherlands, 2013; pp. 29–33. [Google Scholar]
- Michelacci, Y.M. Collagens and proteoglycans of the corneal extracellular matrix. Braz. J. Med. Biol. Res. 2003, 36, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Hay, E.D. Development of the vertebrate cornea. Int. Rev. Cytol. 1980, 63, 263–322. [Google Scholar] [PubMed]
- Joyce, N.C. Proliferative capacity of corneal endothelial cells. Exp. Eye Res. 2012, 95, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, W.M.; McLaren, J.W. Clinical responses of the corneal endothelium. Exp. Eye Res. 2004, 78, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Gipson, I.K. Distribution of mucins at the ocular surface. Exp. Eye Res. 2004, 78, 379–388. [Google Scholar] [CrossRef]
- Jumblatt, M.M.; McKenzie, R.W.; Jumblatt, J.E. MUC5AC mucin is a component of the human precorneal tear film. Investig. Ophthalmol. Vis. Sci. 1999, 40, 43–49. [Google Scholar]
- McKenzie, R.W.; Jumblatt, J.E.; Jumblatt, M.M. Quantification of MUC2 and MUC5AC transcripts in human conjunctiva. Investig. Ophthalmol. Vis. Sci. 2000, 41, 703–708. [Google Scholar]
- Paulsen, F.P.; Berry, M.S. Mucins and TFF peptides of the tear film and lacrimal apparatus. Prog. Histochem. Cytochem. 2006, 41, 1–53. [Google Scholar] [CrossRef] [PubMed]
- Knop, E.; Knop, N. [Eye-associated lymphoid tissue (EALT) is continuously spread throughout the ocular surface from the lacrimal gland to the lacrimal drainage system]. Ophthalmologe 2003, 100, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Steven, P.; Gebert, A. Conjunctiva-associated lymphoid tissue-current knowledge, animal models and experimental prospects. Ophthalmic. Res. 2009, 42, 2–8. [Google Scholar] [CrossRef]
- Gipson, I.K. Goblet cells of the conjunctiva: A review of recent findings. Prog. Retin. Eye Res. 2016, 54, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubota, K.; Tseng, S.C.; Nordlund, M.L. Anatomy and physiology of the ocular surface. In Ocular Surface Disease Medical and Surgical Management; Springer: Berlin/Heidelberg, Germany, 2002; pp. 3–15. [Google Scholar]
- Forrester, J.V.; Dick, A.D.; McMenamin, P.G.; Roberts, F.; Pearlman, E. Anatomy of the eye and orbit. Eye Basic Sci. Pract. 2002, 2, 66–90. [Google Scholar]
- Obata, H. Anatomy and histopathology of the human lacrimal gland. Cornea 2006, 25, S82–S89. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, F.P.; Hinz, M.; Schaudig, U.; Thale, A.B.; Hoffmann, W. TFF peptides in the human efferent tear ducts. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3359–3364. [Google Scholar]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, T.; Zhao, J. Uniform distribution of epithelial stem cells in the bulbar conjunctiva. Investig. Ophthalmol. Vis. Sci. 2005, 46, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Stewart, R.M.; Sheridan, C.M.; Hiscott, P.S.; Czanner, G.; Kaye, S.B. Human conjunctival stem cells are predominantly located in the medial canthal and inferior forniceal areas. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2021–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, F.; Thale, A.; Kohla, G.; Schauer, R.; Rochels, R.; Parwaresch, R.; Tillmann, B. Functional anatomy of human lacrimal duct epithelium. Anat. Embryol. 1998, 198, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, F.P.; Corfield, A.P.; Hinz, M.; Hoffmann, W.; Schaudig, U.; Thale, A.B.; Berry, M. Characterization of mucins in human lacrimal sac and nasolacrimal duct. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1807–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, F.P.; Foge, M.; Thale, A.B.; Tillmann, B.N.; Mentlein, R. Animal model for the absorption of lipophilic substances from tear fluid by the epithelium of the nasolacrimal ducts. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3137–3143. [Google Scholar]
- Paulsen, F.P.; Schaudig, U.; Thale, A.B. Drainage of tears: Impact on the ocular surface and lacrimal system. Ocul. Surf. 2003, 1, 180–191. [Google Scholar] [CrossRef]
- Paulsen, F.P.; Paulsen, J.I.; Thale, A.B.; Tillmann, B.N. Mucosa-associated lymphoid tissue in human efferent tear ducts. Virchows Arch. 2000, 437, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, F.P.; Schaudig, U.; Maune, S.; Thale, A.B. Loss of tear duct-associated lymphoid tissue in association with the scarring of symptomatic dacryostenosis. Ophthalmology 2003, 110, 85–92. [Google Scholar] [CrossRef]
- Paulsen, F.P.; Pufe, T.; Schaudig, U.; Held-Feindt, J.; Lehmann, J.; Schroder, J.M.; Tillmann, B.N. Detection of natural peptide antibiotics in human nasolacrimal ducts. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2157–2163. [Google Scholar]
- Paulsen, F.P.; Thale, A.B.; Hallmann, U.J.; Schaudig, U.; Tillmann, B.N. The cavernous body of the human efferent tear ducts: Function in tear outflow mechanism. Investig. Ophthalmol. Vis. Sci. 2000, 41, 965–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayub, M.; Thale, A.B.; Hedderich, J.; Tillmann, B.N.; Paulsen, F.P. The cavernous body of the human efferent tear ducts contributes to regulation of tear outflow. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4900–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkema, J.R.; Carey, S.A.; Wagner, J.G. The nose revisited: A brief review of the comparative structure, function, and toxicologic pathology of the nasal epithelium. Toxicol. Pathol. 2006, 34, 252–269. [Google Scholar] [CrossRef]
- Mygind, N.; Dahl, R. Anatomy, physiology and function of the nasal cavities in health and disease. Adv. Drug Deliv. Rev. 1998, 29, 3–12. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Peiser, C.; Dinh, Q.T.; Matthias, J.; Eynott, P.R.; Heppt, W.; Carlstedt, I.; Witt, C.; Fischer, A.; Chung, K.F. Distribution of respiratory mucin proteins in human nasal mucosa. Laryngoscope 2003, 113, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V.; Dickey, B.F. Airway mucus function and dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos Silva, E.; Ulrich, M.; Döring, G.; Botzenhart, K.; Gött, P. Trefoil factor family domain peptides in the human respiratory tract. J. Pathol. 2000, 190, 133–142. [Google Scholar] [CrossRef]
- Ali, M.S.; Pearson, J.P. Upper airway mucin gene expression: A review. Laryngoscope 2007, 117, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Tos, M. Distribution of mucus producing elements in the respiratory tract. Differences between upper and lower airway. Eur. J. Respir. Dis. Suppl. 1983, 128, 269–279. [Google Scholar] [PubMed]
- Manafi, N.; Shokri, F.; Achberger, K.; Hirayama, M.; Mohammadi, M.H.; Noorizadeh, F.; Hong, J.; Liebau, S.; Tsuji, T.; Quinn, P.M.J.; et al. Organoids and organ chips in ophthalmology. Ocul. Surf. 2021, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Blaiss, M.S.; Hammerby, E.; Robinson, S.; Kennedy-Martin, T.; Buchs, S. The burden of allergic rhinitis and allergic rhinoconjunctivitis on adolescents: A literature review. Ann. Allergy Asthma Immunol. 2018, 121, 43–52.e43. [Google Scholar] [CrossRef] [Green Version]
- Shiju, T.M.; Carlos de Oliveira, R.; Wilson, S.E. 3D in vitro corneal models: A review of current technologies. Exp. Eye Res. 2020, 200, 108213. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Duval, K.; Grover, H.; Han, L.H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Naumova, E.A.; Bellm, A.; Behrendt, A.K.; Giachero, F.; Bahlmann, N.; Zhang, W.; Wirth, S.; Ehrhardt, A.; Arnold, W.H.; et al. The mystery behind the nostrils-technical clues for successful nasal epithelial cell cultivation. Ann. Anat. 2021, 238, 151748. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Naumova, E.A.; Bellm, A.; Behrendt, A.K.; Giachero, F.; Bahlmann, N.; Zhang, W.; Wirth, S.; Paulsen, F.; Arnold, W.H.; et al. From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection. Viruses 2021, 13, 387. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Naumova, E.A.; Paulsen, F.; Zhang, W.; Gopon, F.; Theis, C.; Lutz, S.; Ehrke-Schulz, E.; Arnold, W.H.; Wirth, S.; et al. House Dust Mite Exposure Causes Increased Susceptibility of Nasal Epithelial Cells to Adenovirus Infection. Viruses 2020, 12, 1151. [Google Scholar] [CrossRef] [PubMed]
- Chioni, A.M.; Bajwa, R.T.; Grose, R. 3D Organotypic Culture Model to Study Components of ERK Signaling. Methods Mol. Biol. 2017, 1487, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Chioni, A.M.; Grose, R. Organotypic modelling as a means of investigating epithelial-stromal interactions during tumourigenesis. Fibrogenesis Tissue Repair 2008, 1, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, T.B.; Ford, A.; Wang, S.; Cairns, D.M.; Parker, R.N.; Deardorff, P.M.; Ghezzi, C.E.; Kaplan, D.L. Assembly and Application of a Three-Dimensional Human Corneal Tissue Model. Curr. Protoc. Toxicol. 2019, 81, e84. [Google Scholar] [CrossRef]
- McKay, T.B.; Karamichos, D.; Hutcheon, A.E.K.; Guo, X.; Zieske, J.D. Corneal Epithelial-Stromal Fibroblast Constructs to Study Cell-Cell Communication in Vitro. Bioengineering 2019, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espana, E.M.; Birk, D.E. Composition, structure and function of the corneal stroma. Exp. Eye Res. 2020, 198, 108137. [Google Scholar] [CrossRef]
- Meek, K.M.; Knupp, C. Corneal structure and transparency. Prog. Retin. Eye Res. 2015, 49, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Petrovski, G.; Albert, R.; Alkanaan, A.; Kirat, O.; Khan, A.D.; Almubrad, T. Ultrastructure and 3D transmission electron tomography of collagen fibrils and proteoglycans of swollen human corneal stroma. Histol. Histopathol. 2019, 34, 91–102. [Google Scholar] [CrossRef]
- Alkanaan, A.; Barsotti, R.; Kirat, O.; Khan, A.; Almubrad, T.; Akhtar, S. Collagen fibrils and proteoglycans of peripheral and central stroma of the keratoconus cornea-Ultrastructure and 3D transmission electron tomography. Sci. Rep. 2019, 9, 19963. [Google Scholar] [CrossRef]
- Meek, K.M.; Leonard, D.W. Ultrastructure of the corneal stroma: A comparative study. Biophys. J. 1993, 64, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Alkatan, H.; Kirat, O.; Almubrad, T. Ultrastructural and three-dimensional study of post-LASIK ectasia cornea. Microsc. Res. Tech. 2014, 77, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Alkatan, H.M.; Kirat, O.; Khan, A.A.; Almubrad, T. Collagen Fibrils and Proteoglycans of Macular Dystrophy Cornea: Ultrastructure and 3D Transmission Electron Tomography. Microsc. Microanal. 2015, 21, 666–679. [Google Scholar] [CrossRef]
- Annadanam, A.; Stoeger, C.G.; Galloway, J.D.; Hikes, M.T.; Jun, A.S. Optical Coherence Tomography Assessment of the Cornea During Corneal Swelling: Should the Term "Descemet Membrane Folds" Be Reconsidered? Cornea 2019, 38, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.S.; Brown, D.M.; Conn, R.E.; McNabb, R.P.; Pardue, M.T.; Kuo, A.N. Topography and pachymetry maps for mouse corneas using optical coherence tomography. Exp. Eye Res. 2020, 190, 107868. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Nioi, M.; d’Aloja, E.; Loy, F.; Fossarello, M. The architecture of corneal stromal striae on optical coherence tomography and histology in an animal model and in humans. Sci. Rep. 2020, 10, 19861. [Google Scholar] [CrossRef] [PubMed]
- Kaluzhny, Y.; Kinuthia, M.W.; Lapointe, A.M.; Truong, T.; Klausner, M.; Hayden, P. Oxidative stress in corneal injuries of different origin: Utilization of 3D human corneal epithelial tissue model. Exp. Eye Res. 2020, 190, 107867. [Google Scholar] [CrossRef]
- Kaluzhny, Y.; Kinuthia, M.W.; Truong, T.; Lapointe, A.M.; Hayden, P.; Klausner, M. New Human Organotypic Corneal Tissue Model for Ophthalmic Drug Delivery Studies. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2880–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, I.; Capallere, C.; Arcioni, M.; Brulas, M.; Plaza, C.; Meyrignac, C.; Bauza, E.; Botto, J.M. Establishment and performance assessment of an in-house 3D Reconstructed Human Cornea-Like Epithelium (RhCE) as a screening tool for the identification of liquid chemicals with potential eye hazard. Toxicol. In Vitro 2019, 61, 104604. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.C.G.; Chialchia, A.R.; de Castro, E.G.; MRL, E.S.; Arantes, D.A.C.; Batista, A.C.; Kitten, G.T.; Valadares, M.C. A new corneal epithelial biomimetic 3D model for in vitro eye toxicity assessment: Development, characterization and applicability. Toxicol. In Vitro 2020, 62, 104666. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Foster, J.W.; Wahlin, K.; Adams, S.M.; Birk, D.E.; Zack, D.J.; Chakravarti, S. Cornea organoids from human induced pluripotent stem cells. Sci. Rep. 2017, 7, 41286. [Google Scholar] [CrossRef]
- Van Meenen, J.; Ni Dhubhghaill, S.; Van den Bogerd, B.; Koppen, C. An Overview of Advanced In Vitro Corneal Models: Implications for Pharmacological Testing. Tissue Eng. Part B Rev. 2021. [Google Scholar] [CrossRef]
- Rotzer, V.; Melega, F.; Garreis, F.; Paulsen, F.; Waschke, J. E-Cadherin Is Important for Meibomian Gland Function as Revealed by a New Human ex Vivo Slice Culture Model. Am. J. Pathol. 2019, 189, 1559–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampel, U.; Garreis, F.; Burgemeister, F.; Essel, N.; Paulsen, F. Effect of intermittent shear stress on corneal epithelial cells using an in vitro flow culture model. Ocul. Surf. 2018, 16, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Ronkko, S.; Vellonen, K.S.; Jarvinen, K.; Toropainen, E.; Urtti, A. Human corneal cell culture models for drug toxicity studies. Drug Deliv. Transl. Res. 2016, 6, 660–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Chun, Y.S.; Lee, S.H.; Mun, S.K.; Jung, H.S.; Lee, S.H.; Son, Y.; Kim, J.C. Ocular surface reconstruction with autologous nasal mucosa in cicatricial ocular surface disease. Am. J. Ophthalmol. 2010, 149, 45–53.e42. [Google Scholar] [CrossRef] [PubMed]
- Wenkel, H.; Rummelt, V.; Naumann, G.O. Long term results after autologous nasal mucosal transplantation in severe mucus deficiency syndromes. Br. J. Ophthalmol. 2000, 84, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielory, L.; Delgado, L.; Katelaris, C.H.; Leonardi, A.; Rosario, N.; Vichyanoud, P. ICON: Diagnosis and management of allergic conjunctivitis. Ann. Allergy Asthma Immunol. 2020, 124, 118–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Henao, C.M.; Herrera-Morales, C.I.; Ramirez-Giraldo, R.; Cardona-Villa, R. Quality of life and clinical characterization of patients with vernal keratoconjunctivitis in a pediatric population in Colombia. Allergol. Immunopathol. 2018, 46, 370–377. [Google Scholar] [CrossRef]
- Mikhail, E.; Azizoglu, S.; Gokhale, M.; Suphioglu, C. Questionnaires Assessing the Quality of Life of Ocular Allergy Patients. J. Allergy Clin. Immunol. Pract. 2020, 8, 2945–2952. [Google Scholar] [CrossRef]
- Witt, J.; Dietrich, J.; Mertsch, S.; Schrader, S.; Spaniol, K.; Geerling, G. Decellularized porcine conjunctiva as an alternative substrate for tissue-engineered epithelialized conjunctiva. Ocul. Surf. 2020, 18, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Deng, X.; Wang, P.; Yu, C.; Kiratitanaporn, W.; Wu, X.; Schimelman, J.; Tang, M.; Balayan, A.; Yao, E. Rapid bioprinting of conjunctival stem cell micro-constructs for subconjunctival ocular injection. Biomaterials 2021, 267, 120462. [Google Scholar] [CrossRef] [PubMed]
- Paladino, G.; Marino, C.; Mule, S.L.T.; Civiale, C.; Rusciano, D.; Enea, V. Cytokeratin expression in primary epithelial cell culture from bovine conjunctiva. Tissue Cell 2004, 36, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Zhu, M.; Chen, J.; Shao, C.; Yan, C.; Wang, Z.; Fan, X.; Gu, P.; Fu, Y. Reconstruction of conjunctival epithelium-like tissue using a temperature-responsive culture dish. Mol. Vis. 2015, 21, 1113. [Google Scholar] [PubMed]
- Bertolin, M.; Breda, C.; Ferrari, S.; Van Acker, S.I.; Zakaria, N.; Di Iorio, E.; Migliorati, A.; Ponzin, D.; Ferrari, B.; Lužnik, Z. Optimized protocol for regeneration of the conjunctival epithelium using the cell suspension technique. Cornea 2019, 38, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Posadas, L.; Soriano-Romani, L.; Lopez-Garcia, A.; Diebold, Y. An engineered human conjunctival-like tissue to study ocular surface inflammatory diseases. PLoS ONE 2017, 12, e0171099. [Google Scholar] [CrossRef] [PubMed]
- Rosellini, A.; Papini, S.; Giannarini, C.; Nardi, M.; Revoltella, R.P. Human conjunctival epithelial precursor cells and their progeny in 3D organotypic culture. Int. J. Dev. Biol. 2002, 51, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.-H.; Lee, J.H.; Yoon, J.-H.; Lee, H.K.; Seo, K.Y. Multi-layered culture of primary human conjunctival epithelial cells producing MUC5AC. Exp. Eye Res. 2007, 85, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Keegan, D.E.; Brewington, J.J. Nasal Epithelial Cell-Based Models for Individualized Study in Cystic Fibrosis. Int. J. Mol. Sci. 2021, 22, 4448. [Google Scholar] [CrossRef] [PubMed]
- Nioi, M.; Napoli, P.E.; Demontis, R.; Locci, E.; Fossarello, M.; d’Aloja, E. Morphological analysis of corneal findings modifications after death: A preliminary OCT study on an animal model. Exp. Eye Res. 2018, 169, 20–27. [Google Scholar] [CrossRef]
- Huang, D.; Swanson, E.A.; Lin, C.P.; Schuman, J.S.; Stinson, W.G.; Chang, W.; Hee, M.R.; Flotte, T.; Gregory, K.; Puliafito, C.A.; et al. Optical coherence tomography. Science 1991, 254, 1178–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, P.E.; Nioi, M.; Mangoni, L.; Gentile, P.; Braghiroli, M.; d’Aloja, E.; Fossarello, M. Fourier-Domain OCT Imaging of the Ocular Surface and Tear Film Dynamics: A Review of the State of the Art and an Integrative Model of the Tear Behavior During the Inter-Blink Period and Visual Fixation. J. Clin. Med. 2020, 9, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nioi, M.; Napoli, P.E.; Mayerson, S.M.; Fossarello, M.; d’Aloja, E. Optical coherence tomography in forensic sciences: A review of the literature. Forensic Sci. Med. Pathol. 2019, 15, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Podoleanu, A.G. Optical coherence tomography. J. Microsc. 2012, 247, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, J.; Swanson, E. The Development, Commercialization, and Impact of Optical Coherence Tomography. Investig. Ophthalmol. Vis. Sci. 2016, 57, OCT1–OCT13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, P.E.; Nioi, M.; d’Aloja, E.; Fossarello, M. Post-Mortem Corneal Thickness Measurements with a Portable Optical Coherence Tomography System: A Reliability Study. Sci. Rep. 2016, 6, 30428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, P.E.; Nioi, M.; Gabiati, L.; Laurenzo, M.; De-Giorgio, F.; Scorcia, V.; Grassi, S.; d’Aloja, E.; Fossarello, M. Repeatability and reproducibility of post-mortem central corneal thickness measurements using a portable optical coherence tomography system in humans: A prospective multicenter study. Sci. Rep. 2020, 10, 14508. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Coronella, F.; Satta, G.M.; Zucca, I.A.; Fossarello, M. A novel OCT technique to measure in vivo the corneal adhesiveness for sodium carboxymethylcellulose in humans and its validity in the diagnosis of dry eye. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3179–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messner, A.; Fischak, C.; Pfister, M.; Schutzenberger, K.; Garreis, F.; Paulsen, F.; Stegmann, H.; Aranha Dos Santos, V.; Garhofer, G.; Schmetterer, L.; et al. Characterization of dry eye disease in a mouse model by optical coherence tomography and fluorescein staining. Biomed. Opt. Express 2019, 10, 4884–4895. [Google Scholar] [CrossRef]
- Meyer, J.H.; Larsen, P.P.; Strack, C.; Harmening, W.M.; Krohne, T.U.; Holz, F.G.; Schmitz-Valckenberg, S. Optical coherence tomography angiography (OCT-A) in an animal model of laser-induced choroidal neovascularization. Exp. Eye Res. 2019, 184, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, M.; Hara, A.; Ishiguro, S.I. Optical Coherence Tomography of Animal Models of Retinitis Pigmentosa: From Animal Studies to Clinical Applications. Biomed. Res. Int. 2019, 2019, 8276140. [Google Scholar] [CrossRef] [Green Version]
- Walther, J.; Gaertner, M.; Cimalla, P.; Burkhardt, A.; Kirsten, L.; Meissner, S.; Koch, E. Optical coherence tomography in biomedical research. Anal. Bioanal. Chem. 2011, 400, 2721–2743. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.A.; Moss, W.J.; Takeda, M.; de Swart, R.L.; Thompson, K.M.; Goodson, J.L. Measles. Nat. Rev. Dis. Primers 2016, 2, 16049. [Google Scholar] [CrossRef]
- Cox, R.M.; Plemper, R.K. Structure and organization of paramyxovirus particles. Curr. Opin. Virol. 2017, 24, 105–114. [Google Scholar] [CrossRef]
- Griffin, D.E.; Lin, W.H.; Pan, C.H. Measles virus, immune control, and persistence. FEMS Microbiol. Rev. 2012, 36, 649–662. [Google Scholar] [CrossRef]
- Navaratnarajah, C.K.; Generous, A.R.; Yousaf, I.; Cattaneo, R. Receptor-mediated cell entry of paramyxoviruses: Mechanisms, and consequences for tropism and pathogenesis. J. Biol. Chem. 2020, 295, 2771–2786. [Google Scholar] [CrossRef] [Green Version]
- Leonard, V.H.; Sinn, P.L.; Hodge, G.; Miest, T.; Devaux, P.; Oezguen, N.; Braun, W.; McCray, P.B., Jr.; McChesney, M.B.; Cattaneo, R. Measles virus blind to its epithelial cell receptor remains virulent in rhesus monkeys but cannot cross the airway epithelium and is not shed. J. Clin. Investig. 2008, 118, 2448–2458. [Google Scholar] [CrossRef] [Green Version]
- Moss, W.J. Measles. Lancet 2017, 390, 2490–2502. [Google Scholar] [CrossRef]
- de Swart, R.L. Measles studies in the macaque model. Curr. Top. Microbiol. Immunol. 2009, 330, 55–72. [Google Scholar] [CrossRef]
- Lin, W.W.; Tsay, A.J.; Lalime, E.N.; Pekosz, A.; Griffin, D.E. Primary differentiated respiratory epithelial cells respond to apical measles virus infection by shedding multinucleated giant cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2013264118. [Google Scholar] [CrossRef] [PubMed]
- Lightwood, R.; Nolan, R. Epithelial giant cells in measles as an acid in diagnosis. J. Pediatr. 1970, 77, 59–64. [Google Scholar] [CrossRef]
- Nommensen, F.E.; Dekkers, N.W. Detection of measles antigen in conjunctival epithelial lesions staining by lissamine green during measles virus infection. J. Med. Virol. 1981, 7, 157–162. [Google Scholar] [CrossRef]
- Ludlow, M.; McQuaid, S.; Milner, D.; de Swart, R.L.; Duprex, W.P. Pathological consequences of systemic measles virus infection. J. Pathol. 2015, 235, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Chodosh, J.; Chintakuntlawar, A.V.; Robinson, C.M. Human Eye Infections. Encycl. Virol. 2008, 491–497. [Google Scholar] [CrossRef]
- Dekkers, N. The cornea in measles. Doc. Ophthalmol. 2005, 52, v-121. [Google Scholar]
- Belser, J.A.; Rota, P.A.; Tumpey, T.M. Ocular tropism of respiratory viruses. Microbiol. Mol. Biol. Rev. 2013, 77, 144–156. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, K.; Kobayashi, A.; Higashide, T.; Shirao, Y.; Sakurai, M.; Shirota, Y.; Kagaya, M. Detection of measles virus genomic RNA in tear samples from a patient with measles keratitis. Cornea 2002, 21, 610–612. [Google Scholar] [CrossRef]
- Ludlow, M.; Rennick, L.J.; Sarlang, S.; Skibinski, G.; McQuaid, S.; Moore, T.; de Swart, R.L.; Duprex, W.P. Wild-type measles virus infection of primary epithelial cells occurs via the basolateral surface without syncytium formation or release of infectious virus. J. Gen. Virol. 2010, 91, 971–979. [Google Scholar] [CrossRef]
- Papp, K. Experiments proving that the route of infection in measles is the contamination of the conjunctival mucosa. Rev. Immunol. Ther. Antimicrob. 1956, 20, 27–36. [Google Scholar] [PubMed]
- Kempe, C.H.; Fulginiti, V.A. The pathogenesis of measles virus infection. Arch. Virol. 1965, 16, 103–128. [Google Scholar] [CrossRef] [PubMed]
- Laksono, B.M.; De Vries, R.D.; McQuaid, S.; Duprex, W.P.; De Swart, R.L. Measles Virus Host Invasion and Pathogenesis. Viruses 2016, 8, 210. [Google Scholar] [CrossRef] [Green Version]
- Ludlow, M.; de Vries, R.; Lemon, K.; McQuaid, S.; Millar, E.; Van Amerongen, G.; Yüksel, S.; Verburgh, R.J.; Osterhaus, A.; De Swart, R.; et al. Infection of lymphoid tissues in the macaque upper respiratory tract contributes to the emergence of transmissible measles virus. J. Gen. Virol. 2013, 94, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, M.; Lemon, K.; de Vries, R.; McQuaid, S.; Millar, E.L.; Van Amerongen, G.; Yüksel, S.; Verburgh, R.J.; Osterhaus, A.; De Swart, R.L.; et al. Measles Virus Infection of Epithelial Cells in the Macaque Upper Respiratory Tract Is Mediated by Subepithelial Immune Cells. J. Virol. 2013, 87, 4033–4042. [Google Scholar] [CrossRef] [Green Version]
- Tompkins, V. A characteristic cell in nasal secretions during prodromal measles. J. Am. Med. Assoc. 1955, 157, 711. [Google Scholar] [CrossRef] [PubMed]
- Hippee, C.E.; Singh, B.K.; Thurman, A.L.; Cooney, A.L.; Pezzulo, A.A.; Cattaneo, R.; Sinn, P.L. Measles virus exits human airway epithelia within dislodged metabolically active infectious centers. PLoS Pathog. 2021, 17, e1009458. [Google Scholar] [CrossRef] [PubMed]
- Lion, T. Adenovirus Infections in Immunocompetent and Immunocompromised Patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaiya, J.; Saha, A.; Ismail, A.; Zhou, X.; Su, T.; Chodosh, J. Adenovirus and the Cornea: More Than Meets the Eye. Viruses 2021, 13, 293. [Google Scholar] [CrossRef]
- Ariga, T.; Shimada, Y.; Shiratori, K.; Ohgami, K.; Yamazaki, S.; Tagawa, Y.; Kikuchi, M.; Miyakita, Y.; Fujita, K.; Ishiko, H.; et al. Five New Genome Types of Adenovirus Type 37 Caused Epidemic Keratoconjunctivitis in Sapporo, Japan, for More Than 10 Years. J. Clin. Microbiol. 2005, 43, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lu, X.; Sun, Y.; Lin, C.; Li, F.; Yang, Y.; Liang, Z.; Jia, L.; Chen, L.; Jiang, B.; et al. A swimming pool-associated outbreak of pharyngoconjunctival fever caused by human adenovirus type 4 in Beijing, China. Int. J. Infect. Dis. 2018, 75, 89–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, S.-C.; Shen, S.-C.; Chang, S.-W.; Huang, S.C.M.; Hsiao, C.-H. Corneal Superinfection in Acute Viral Conjunctivitis in Young Children. J. Pediatr. Ophthalmol. Strabismus 2008, 45, 374–376. [Google Scholar] [CrossRef]
- Rhodes, G.C.; Blinkhorn, S.A.; Yong, L.C. Cardiovascular lesions in experimental acute and chronic renal failure in the rat. Exp. Pathol. 1987, 31, 221–229. [Google Scholar] [CrossRef]
- Aydin, M.; Schellhorn, S.; Wirth, S.; Zhang, W.; Ehrhardt, A. Human Species D Adenoviruses Isolated from Diarrheal Feces Show Low Infection Rates in Primary Nasal Epithelial Cells. Children 2021, 8, 563. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.C.; Storm, R.J.; Bauer, J.; Johansson, S.M.; Lookene, A.; Angstrom, J.; Hedenstrom, M.; Eriksson, T.L.; Frangsmyr, L.; Rinaldi, S.; et al. The GD1a glycan is a cellular receptor for adenoviruses causing epidemic keratoconjunctivitis. Nat. Med. 2011, 17, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.; Trauger, S.A.; Pache, L.; Mullen, T.M.; von Seggern, D.J.; Siuzdak, G.; Nemerow, G.R. Membrane cofactor protein is a receptor for adenoviruses associated with epidemic keratoconjunctivitis. J. Virol. 2004, 78, 3897–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, N.; Frangsmyr, L.; Imhof, S.; Caraballo, R.; Elofsson, M.; Arnberg, N. Sialic Acid-Containing Glycans as Cellular Receptors for Ocular Human Adenoviruses: Implications for Tropism and Treatment. Viruses 2019, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Li, Z.Y.; Liu, Y.; Persson, J.; Beyer, I.; Moller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2011, 17, 96–104. [Google Scholar] [CrossRef]
- Roelvink, P.W.; Lizonova, A.; Lee, J.G.; Li, Y.; Bergelson, J.M.; Finberg, R.W.; Brough, D.E.; Kovesdi, I.; Wickham, T.J. The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J. Virol. 1998, 72, 7909–7915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Storm, R.J.; Persson, B.D.; Skalman, L.N.; Frangsmyr, L.; Lindstrom, M.; Rankin, G.; Lundmark, R.; Domellof, F.P.; Arnberg, N. Human Adenovirus Type 37 Uses alphaVbeta1 and alpha3beta1 Integrins for Infection of Human Corneal Cells. J. Virol. 2017, 91, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramke, M.; Lam, E.; Meyer, M.; Knipper, A.; Heim, A. Porcine corneal cell culture models for studying epidemic keratoconjunctivitis. Mol. Vis. 2013, 19, 614–622. [Google Scholar] [PubMed]
- Rajaiya, J.; Zhou, X.; Barequet, I.; Gilmore, M.S.; Chodosh, J. Novel model of innate immunity in corneal infection. In Vitro Cell Dev. Biol. Anim. 2015, 51, 827–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of growth and differentiation in a telomerase-immortalized human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Lin, L.T.; Richardson, C.D. The Host Cell Receptors for Measles Virus and Their Interaction with the Viral Hemagglutinin (H) Protein. Viruses 2016, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M. Measles virus breaks through epithelial cell barriers to achieve transmission. J. Clin. Investig. 2008, 118, 2386–2389. [Google Scholar] [CrossRef] [PubMed]
- Nemerow, G.R.; Stewart, P.L. Role of alpha(v) integrins in adenovirus cell entry and gene delivery. Microbiol. Mol. Biol. Rev. 1999, 63, 725–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aneeza, W.H.; Husain, S.; Rahman, R.A.; Van Dort, D.; Abdullah, A.; Gendeh, B.S. Efficacy of mometasone furoate and fluticasone furoate on persistent allergic rhinoconjunctivitis. Allergy Rhinol. 2013, 4, e120–e126. [Google Scholar] [CrossRef]
- Ciprandi, G.; Leonardi, S.; Zicari, A.M.; Tosca, M.A.; Marseglia, G. Allergic rhinoconjunctivitis: Pathophysiological mechanism and new therapeutic approach. Acta Biomed. 2020, 91, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Origlieri, C.; Bielory, L. Intranasal corticosteroids and allergic rhinoconjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Bielory, L. Allergic conjunctivitis and the impact of allergic rhinitis. Curr. Allergy Asthma Rep. 2010, 10, 122–134. [Google Scholar] [CrossRef]
- Rosario, N.; Bielory, L. Epidemiology of allergic conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 471–476. [Google Scholar] [CrossRef]
- Dhami, S.; Nurmatov, U.; Arasi, S.; Khan, T.; Asaria, M.; Zaman, H.; Agarwal, A.; Netuveli, G.; Roberts, G.; Pfaar, O.; et al. Allergen immunotherapy for allergic rhinoconjunctivitis: A systematic review and meta-analysis. Allergy 2017, 72, 1597–1631. [Google Scholar] [CrossRef] [Green Version]
- Bousquet, J.; Anto, J.M.; Bachert, C.; Baiardini, I.; Bosnic-Anticevich, S.; Walter Canonica, G.; Melen, E.; Palomares, O.; Scadding, G.K.; Togias, A.; et al. Allergic rhinitis. Nat. Rev. Dis. Primers 2020, 6, 95. [Google Scholar] [CrossRef]
- Kakli, H.A.; Riley, T.D. Allergic Rhinitis. Prim. Care 2016, 43, 465–475. [Google Scholar] [CrossRef]
- Bousquet, J.; Khaltaev, N.; Cruz, A.A.; Denburg, J.; Fokkens, W.J.; Togias, A.; Zuberbier, T.; Baena-Cagnani, C.E.; Canonica, G.W.; van Weel, C.; et al. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in collaboration with the World Health Organization, GA(2)LEN and AllerGen). Allergy 2008, 63 (Suppl. S86), 8–160. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, N.T.; McGrath, K.G. Allergic and nonallergic rhinitis. Allergy Asthma Proc. 2019, 40, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Skoner, D.P. Allergic rhinitis: Definition, epidemiology, pathophysiology, detection, and diagnosis. J. Allergy Clin. Immunol. 2001, 108, S2–S8. [Google Scholar] [CrossRef] [PubMed]
- Schuler Iv, C.F.; Montejo, J.M. Allergic Rhinitis in Children and Adolescents. Pediatr. Clin. N. Am. 2019, 66, 981–993. [Google Scholar] [CrossRef]
- Bousquet, J.; Schunemann, H.J.; Togias, A.; Bachert, C.; Erhola, M.; Hellings, P.W.; Klimek, L.; Pfaar, O.; Wallace, D.; Ansotegui, I.; et al. Next-generation Allergic Rhinitis and Its Impact on Asthma (ARIA) guidelines for allergic rhinitis based on Grading of Recommendations Assessment, Development and Evaluation (GRADE) and real-world evidence. J. Allergy Clin. Immunol. 2020, 145, 70–80.e73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozek, J.L.; Bousquet, J.; Agache, I.; Agarwal, A.; Bachert, C.; Bosnic-Anticevich, S.; Brignardello-Petersen, R.; Canonica, G.W.; Casale, T.; Chavannes, N.H.; et al. Allergic Rhinitis and its Impact on Asthma (ARIA) guidelines-2016 revision. J. Allergy Clin. Immunol. 2017, 140, 950–958. [Google Scholar] [CrossRef] [Green Version]
- Rosati, M.G.; Peters, A.T. Relationships among allergic rhinitis, asthma, and chronic rhinosinusitis. Am. J. Rhinol. Allergy 2016, 30, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Tantilipikorn, P.; Sompornrattanaphan, M.; Suwanwech, T.; Ngaotepprutaram, P. Chronic Rhinosinusitis and Allergy: Increased Allergen Sensitization Versus Real Allergic Rhinitis Multimorbidity: A Systematic Review. Curr. Allergy Asthma Rep. 2020, 20, 19. [Google Scholar] [CrossRef]
- Leonardi, A.; Bogacka, E.; Fauquert, J.L.; Kowalski, M.L.; Groblewska, A.; Jedrzejczak-Czechowicz, M.; Doan, S.; Marmouz, F.; Demoly, P.; Delgado, L. Ocular allergy: Recognizing and diagnosing hypersensitivity disorders of the ocular surface. Allergy 2012, 67, 1327–1337. [Google Scholar] [CrossRef]
- Miraglia Del Giudice, M.; Allegorico, A.; Marseglia, G.L.; Martelli, A.; Calvani, M.; Cardinale, F.; Duse, M.; Chiappini, E.; Manti, S.; Cravidi, C.; et al. Allergic rhinoconjunctivitis. Acta Biomed. 2020, 91, e2020007. [Google Scholar] [CrossRef]
- Patel, D.S.; Arunakirinathan, M.; Stuart, A.; Angunawela, R. Allergic eye disease. BMJ 2017, 359, j4706. [Google Scholar] [CrossRef] [PubMed]
- Jalbert, I.; Golebiowski, B. Environmental aeroallergens and allergic rhino-conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 476–481. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef] [PubMed]
- Gans, M.D.; Gavrilova, T. Understanding the immunology of asthma: Pathophysiology, biomarkers, and treatments for asthma endotypes. Paediatr. Respir. Rev. 2020, 36, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Aron, J.L.; Akbari, O. Regulatory T cells and type 2 innate lymphoid cell-dependent asthma. Allergy 2017, 72, 1148–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasha, M.A.; Patel, G.; Hopp, R.; Yang, Q. Role of innate lymphoid cells in allergic diseases. Allergy Asthma Proc. 2019, 40, 138–145. [Google Scholar] [CrossRef]
- Kabata, H.; Moro, K.; Koyasu, S. The group 2 innate lymphoid cell (ILC2) regulatory network and its underlying mechanisms. Immunol. Rev. 2018, 286, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Massoud, A.H.; Charbonnier, L.M.; Lopez, D.; Pellegrini, M.; Phipatanakul, W.; Chatila, T.A. An asthma-associated IL4R variant exacerbates airway inflammation by promoting conversion of regulatory T cells to TH17-like cells. Nat. Med. 2016, 22, 1013–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seumois, G.; Ramirez-Suastegui, C.; Schmiedel, B.J.; Liang, S.; Peters, B.; Sette, A.; Vijayanand, P. Single-cell transcriptomic analysis of allergen-specific T cells in allergy and asthma. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Choy, D.F.; Hart, K.M.; Borthwick, L.A.; Shikotra, A.; Nagarkar, D.R.; Siddiqui, S.; Jia, G.; Ohri, C.M.; Doran, E.; Vannella, K.M.; et al. TH2 and TH17 inflammatory pathways are reciprocally regulated in asthma. Sci. Transl. Med. 2015, 7, 301ra129. [Google Scholar] [CrossRef] [Green Version]
- Dykewicz, M.S.; Rodrigues, J.M.; Slavin, R.G. Allergic fungal rhinosinusitis. J. Allergy Clin. Immunol. 2018, 142, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltzer, E.O.; Blaiss, M.S.; Derebery, M.J.; Mahr, T.A.; Gordon, B.R.; Sheth, K.K.; Simmons, A.L.; Wingertzahn, M.A.; Boyle, J.M. Burden of allergic rhinitis: Results from the Pediatric Allergies in America survey. J. Allergy Clin. Immunol. 2009, 124, S43–S70. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. Barrier Epithelial Cells and the Control of Type 2 Immunity. Immunity 2015, 43, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holgate, S.T. Innate and adaptive immune responses in asthma. Nat. Med. 2012, 18, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Wang, C.; Zhang, L. Recent developments and highlights in allergic rhinitis. Allergy 2019, 74, 2320–2328. [Google Scholar] [CrossRef] [Green Version]
- Asher, M.I.; Montefort, S.; Bjorksten, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H.; Group, I.P.T.S. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC Phases One and Three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, B.B.; Salvi, S.; Chhatwal, J.; Jain, K.C.; Kumar, L.; Joshi, M.K.; Pandramajal, S.B.; Awasthi, S.; Bhave, S.; et al. Allergic rhinitis, rhinoconjunctivitis, and eczema: Prevalence and associated factors in children. Clin. Respir. J. 2018, 12, 547–556. [Google Scholar] [CrossRef]
- Ison, M.G.; Hayden, R.T. Adenovirus. Microbiol. Spectr. 2016, 4, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Wold, W.S.; Toth, K. Adenovirus vectors for gene therapy, vaccination and cancer gene therapy. Curr. Gene Ther. 2013, 13, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Aref, S.; Bailey, K.; Fielding, A. Measles to the Rescue: A Review of Oncolytic Measles Virus. Viruses 2016, 8, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, S.; Yadava, P.K. Measles virus: Background and oncolytic virotherapy. Biochem. Biophys. Rep. 2018, 13, 58–62. [Google Scholar] [CrossRef]
- Huang, H.; Liu, Y.; Liao, W.; Cao, Y.; Liu, Q.; Guo, Y.; Lu, Y.; Xie, Z. Oncolytic adenovirus programmed by synthetic gene circuit for cancer immunotherapy. Nat. Commun. 2019, 10, 4801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leber, M.F.; Neault, S.; Jirovec, E.; Barkley, R.; Said, A.; Bell, J.C.; Ungerechts, G. Engineering and combining oncolytic measles virus for cancer therapy. Cytokine Growth Factor Rev. 2020, 56, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lobaina Mato, Y. Nasal route for vaccine and drug delivery: Features and current opportunities. Int. J. Pharm. 2019, 572, 118813. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, J.V., Jr.; Golshahi, L.; Rahman, N.; Li, L. Evaluation of Intranasal Vaccine Delivery Using Anatomical Replicas of Infant Nasal Airways. Pharm. Res. 2021, 38, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Yuki, Y.; Uchida, Y.; Sawada, S.I.; Nakahashi-Ouchida, R.; Sugiura, K.; Mori, H.; Yamanoue, T.; Machita, T.; Honma, A.; Kurokawa, S.; et al. Characterization and Specification of a Trivalent Protein-Based Pneumococcal Vaccine Formulation Using an Adjuvant-Free Nanogel Nasal Delivery System. Mol. Pharm. 2021, 18, 1582–1592. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydin, M.; Dietrich, J.; Witt, J.; Finkbeiner, M.S.C.; Park, J.J.-H.; Wirth, S.; Engeland, C.E.; Paulsen, F.; Ehrhardt, A. The Communication between Ocular Surface and Nasal Epithelia in 3D Cell Culture Technology for Translational Research: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 12994. https://doi.org/10.3390/ijms222312994
Aydin M, Dietrich J, Witt J, Finkbeiner MSC, Park JJ-H, Wirth S, Engeland CE, Paulsen F, Ehrhardt A. The Communication between Ocular Surface and Nasal Epithelia in 3D Cell Culture Technology for Translational Research: A Narrative Review. International Journal of Molecular Sciences. 2021; 22(23):12994. https://doi.org/10.3390/ijms222312994
Chicago/Turabian StyleAydin, Malik, Jana Dietrich, Joana Witt, Maximiliane S. C. Finkbeiner, Jonas J.-H. Park, Stefan Wirth, Christine E. Engeland, Friedrich Paulsen, and Anja Ehrhardt. 2021. "The Communication between Ocular Surface and Nasal Epithelia in 3D Cell Culture Technology for Translational Research: A Narrative Review" International Journal of Molecular Sciences 22, no. 23: 12994. https://doi.org/10.3390/ijms222312994
APA StyleAydin, M., Dietrich, J., Witt, J., Finkbeiner, M. S. C., Park, J. J. -H., Wirth, S., Engeland, C. E., Paulsen, F., & Ehrhardt, A. (2021). The Communication between Ocular Surface and Nasal Epithelia in 3D Cell Culture Technology for Translational Research: A Narrative Review. International Journal of Molecular Sciences, 22(23), 12994. https://doi.org/10.3390/ijms222312994