
Ribosome Fate during Decoding of UGA-Sec Codons



{kind=link}
{kind=link}
Abstract
:1. Introduction
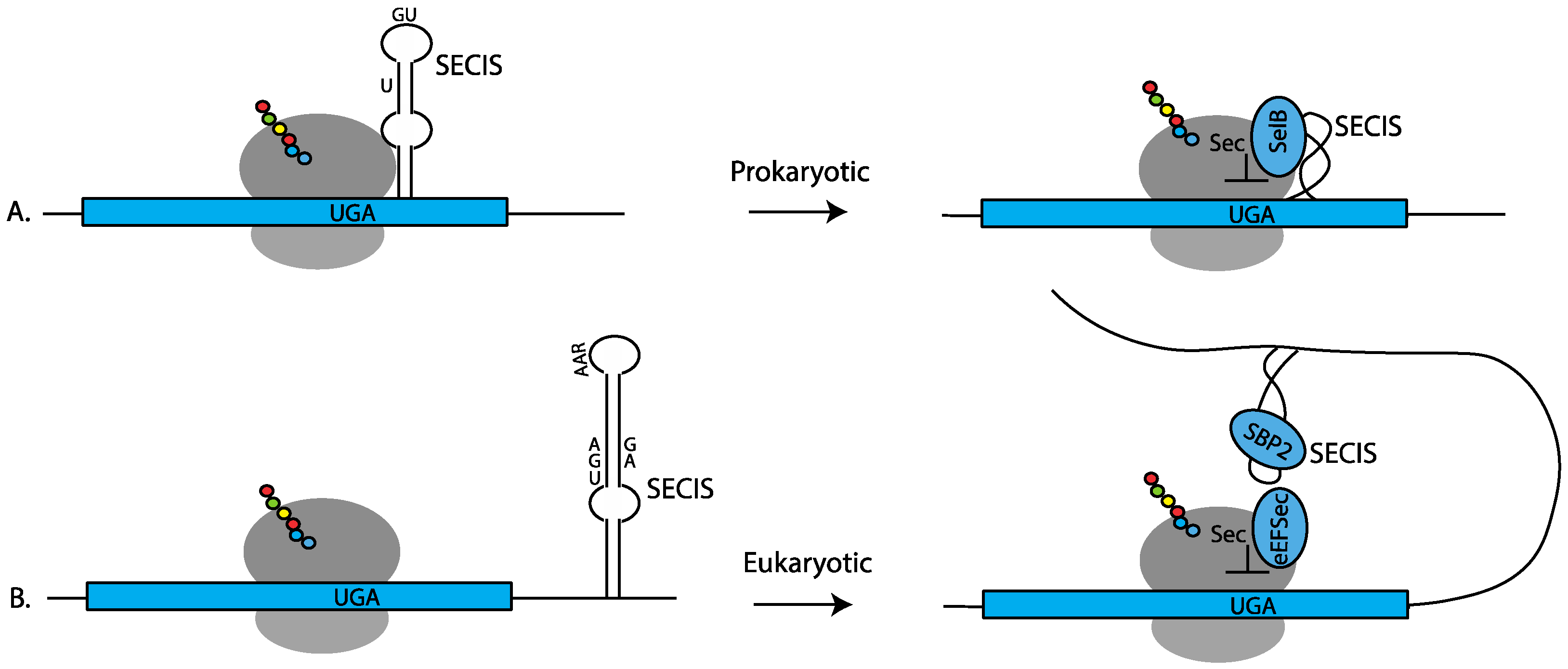
2. Co-Translational Sec-Incorporation
2.1. Cis-Acting Elements
2.1.1. Required Selenocysteine Insertion Sequences
2.1.2. Accessory Cis-Acting Selenocysteine Insertion Elements
2.2. Trans-Acting Factors
2.2.1. The Core SECIS-Binding Proteins
2.2.2. Accessory Trans-Acting Factors
2.2.3. Selenoprotein mRNA 5′ Cap Modifications and Recruitment of the SMN Complex
3. Competing Ribosome Fates at UGA-Sec Codons
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atkins, J.F.; Gesteland, R.F. Recoding: Expansion of decoding rules enriches gene expression. In Nucleic Acids and Molecular Biology; Springer: New York, NY, USA, 2010. [Google Scholar]
- Vindry, C.; Ohlmann, T.; Chavatte, L. Translation regulation of mammalian selenoproteins. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2480–2492. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.T.; Copeland, P.R. New directions for understanding the codon redefinition required for selenocysteine incorporation. Biol. Trace Elem. Res. 2019, 192, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigo, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Kryukov, G.V.; Gladyshev, V.N. The prokaryotic selenoproteome. EMBO Rep. 2004, 5, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Heider, J.; Baron, C.; Bock, A. Coding from a distance: Dissection of the mRNA determinants required for the incorporation of selenocysteine into protein. EMBO J. 1992, 11, 3759–3766. [Google Scholar] [CrossRef]
- Liu, Z.; Reches, M.; Groisman, I.; Engelberg-Kulka, H. The nature of the minimal ‘selenocysteine insertion sequence’ (secis) in Escherichia coli. Nucleic Acids Res. 1998, 26, 896–902. [Google Scholar] [CrossRef] [Green Version]
- Kromayer, M.; Wilting, R.; Tormay, P.; Bock, A. Domain structure of the prokaryotic selenocysteine-specific elongation factor Selb. J. Mol. Biol. 1996, 262, 413–420. [Google Scholar] [CrossRef]
- Berry, M.J.; Banu, L.; Chen, Y.Y.; Mandel, S.J.; Kieffer, J.D.; Harney, J.W.; Larsen, P.R. Recognition of uga as a selenocysteine codon in type i deiodinase requires sequences in the 3′ untranslated region. Nature 1991, 353, 273–276. [Google Scholar] [CrossRef]
- Berry, M.J.; Banu, L.; Harney, J.W.; Larsen, P.R. Functional characterization of the eukaryotic secis elements which direct selenocysteine insertion at uga codons. EMBO J. 1993, 12, 3315–3322. [Google Scholar] [CrossRef]
- Hill, K.E.; Lloyd, R.S.; Burk, R.F. Conserved nucleotide sequences in the open reading frame and 3′ untranslated region of selenoprotein p mRNA. Proc. Natl. Acad. Sci. USA 1993, 90, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Chu, F.F.; Newburger, P.E. Sequences in the 3′-untranslated region of the human cellular glutathione peroxidase gene are necessary and sufficient for selenocysteine incorporation at the uga codon. J. Biol. Chem. 1993, 268, 11463–11469. [Google Scholar] [CrossRef]
- Turanov, A.A.; Lobanov, A.V.; Fomenko, D.E.; Morrison, H.G.; Sogin, M.L.; Klobutcher, L.A.; Hatfield, D.L.; Gladyshev, V.N. Genetic code supports targeted insertion of two amino acids by one codon. Science 2009, 323, 259–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turanov, A.A.; Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Uga codon position-dependent incorporation of selenocysteine into mammalian selenoproteins. Nucleic. Acids. Res. 2013, 41, 6952–6959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, R.; Carbon, P.; Krol, A. An essential non-watson-crick base pair motif in 3’utr to mediate selenoprotein translation. RNA 1998, 4, 74–84. [Google Scholar]
- Walczak, R.; Westhof, E.; Carbon, P.; Krol, A. A novel rna structural motif in the selenocysteine insertion element of eukaryotic selenoprotein mrnas. RNA 1996, 2, 367–379. [Google Scholar] [PubMed]
- Buettner, C.; Harney, J.W.; Berry, M.J. The Caenorhabditis elegans homologue of thioredoxin reductase contains a selenocysteine insertion sequence (secis) element that differs from mammalian secis elements but directs selenocysteine incorporation. J. Biol. Chem. 1999, 274, 21598–21602. [Google Scholar] [CrossRef] [Green Version]
- Grundner-Culemann, E.; Martin, G.W., 3rd; Harney, J.W.; Berry, M.J. Two distinct secis structures capable of directing selenocysteine incorporation in eukaryotes. RNA 1999, 5, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M. Secisearch3 and seblastian: In-silico tools to predict secis elements and selenoproteins. Methods Mol. Biol. 2018, 1661, 3–16. [Google Scholar]
- Fixsen, S.M.; Howard, M.T. Processive selenocysteine incorporation during synthesis of eukaryotic selenoproteins. J. Mol. Biol. 2010, 399, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.P.; Sturts, R.; Vetick, M.; Copeland, P.R. Processive incorporation of multiple selenocysteine residues is driven by a novel feature of the selenocysteine insertion sequence. J. Biol. Chem. 2018, 293, 19377–19386. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Shetty, S.; Baird, L.; Wu, S.; Loughran, G.; Copeland, P.R.; Atkins, J.F.; Howard, M.T. Multiple RNA structures affect translation initiation and uga redefinition efficiency during synthesis of selenoprotein p. Nucleic Acids Res. 2017, 45, 13004–13015. [Google Scholar] [CrossRef] [Green Version]
- Stoytcheva, Z.; Tujebajeva, R.M.; Harney, J.W.; Berry, M.J. Efficient incorporation of multiple selenocysteines involves an inefficient decoding step serving as a potential translational checkpoint and ribosome bottleneck. Mol. Cell Biol. 2006, 26, 9177–9184. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Mariotti, M.; Santesmasses, D.; Hill, K.E.; Baclaocos, J.; Aparicio-Prat, E.; Li, S.; Mackrill, J.; Wu, Y.; Howard, M.T.; et al. Human selenoprotein p and s variant mrnas with different numbers of secis elements and inferences from mutant mice of the roles of multiple secis elements. Open Biol. 2016, 6, 160241. [Google Scholar] [CrossRef]
- Feng, Y.X.; Yuan, H.; Rein, A.; Levin, J.G. Bipartite signal for read-through suppression in murine leukemia virus mRNA: An eight-nucleotide purine-rich sequence immediately downstream of the gag termination codon followed by an RNA pseudoknot. J. Virol. 1992, 66, 5127–5132. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Rice, C.M. The signal for translational readthrough of a uga codon in sindbis virus rna involves a single cytidine residue immediately downstream of the termination codon. J. Virol. 1993, 67, 5062–5067. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Phillips-Jones, M.K.; Watson, F.J.; Hill, L.S. Codon context effects on nonsense suppression in human cells. Biochem. Soc. Trans. 1993, 21, 846–851. [Google Scholar] [CrossRef] [Green Version]
- Mottagui-Tabar, S.; Bjornsson, A.; Isaksson, L.A. The second to last amino acid in the nascent peptide as a codon context determinant. EMBO J. 1994, 13, 249–257. [Google Scholar] [CrossRef]
- Mottagui-Tabar, S.; Tuite, M.F.; Isaksson, L.A. The influence of 5′ codon context on translation termination in Saccharomyces cerevisiae. Eur. J. Biochem. 1998, 257, 249–254. [Google Scholar] [CrossRef]
- Wills, N.M.; Gesteland, R.F.; Atkins, J.F. Evidence that a downstream pseudoknot is required for translational read-through of the moloney murine leukemia virus gag stop codon. Proc. Natl. Acad. Sci. USA 1991, 88, 6991–6995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundner-Culemann, E.; Martin, G.W., 3rd; Tujebajeva, R.; Harney, J.W.; Berry, M.J. Interplay between termination and translation machinery in eukaryotic selenoprotein synthesis. J. Mol. Biol. 2001, 310, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Copeland, P.R. Functional analysis of the interplay between translation termination, selenocysteine codon context, and selenocysteine insertion sequence-binding protein 2. J. Biol. Chem. 2007, 282, 36797–36807. [Google Scholar] [CrossRef] [Green Version]
- McCaughan, K.K.; Brown, C.M.; Dalphin, M.E.; Berry, M.J.; Tate, W.P. Translational termination efficiency in mammals is influenced by the base following the stop codon. Proc. Natl. Acad. Sci. USA 1995, 92, 5431–5435. [Google Scholar] [CrossRef] [Green Version]
- Howard, M.T.; Aggarwal, G.; Anderson, C.B.; Khatri, S.; Flanigan, K.M.; Atkins, J.F. Recoding elements located adjacent to a subset of eukaryal selenocysteine-specifying uga codons. EMBO J. 2005, 24, 1596–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, M.T.; Moyle, M.W.; Aggarwal, G.; Carlson, B.A.; Anderson, C.B. A recoding element that stimulates decoding of uga codons by sec trna[ser]sec. RNA 2007, 13, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Maiti, B.; Arbogast, S.; Allamand, V.; Moyle, M.W.; Anderson, C.B.; Richard, P.; Guicheney, P.; Ferreiro, A.; Flanigan, K.M.; Howard, M.T. A mutation in the sepn1 selenocysteine redefinition element (SRE) reduces selenocysteine incorporation and leads to Sepn1-related myopathy. Hum. Mutat. 2009, 30, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.S.; Bejerano, G.; Siepel, A.; Rosenbloom, K.; Lindblad-Toh, K.; Lander, E.S.; Kent, J.; Miller, W.; Haussler, D. Identification and classification of conserved rna secondary structures in the human genome. PLoS Comput. Biol. 2006, 2, e33. [Google Scholar] [CrossRef]
- Bubenik, J.L.; Miniard, A.C.; Driscoll, D.M. Alternative transcripts and 3′utr elements govern the incorporation of selenocysteine into selenoprotein s. PLoS ONE 2013, 8, e62102. [Google Scholar] [CrossRef] [Green Version]
- Cockman, E.M.; Narayan, V.; Willard, B.; Shetty, S.P.; Copeland, P.R.; Driscoll, D.M. Identification of the selenoprotein s positive uga recoding (spur) element and its position-dependent activity. RNA Biol. 2019, 16, 1682–1696. [Google Scholar] [CrossRef]
- Copeland, P.R.; Fletcher, J.E.; Carlson, B.A.; Hatfield, D.L.; Driscoll, D.M. A novel RNA binding protein, SBP2, is required for the translation of mammalian selenoprotein mrnas. EMBO J. 2000, 19, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Fagegaltier, D.; Hubert, N.; Yamada, K.; Mizutani, T.; Carbon, P.; Krol, A. Characterization of mselb, a novel mammalian elongation factor for selenoprotein translation. EMBO J. 2000, 19, 4796–4805. [Google Scholar] [CrossRef] [Green Version]
- Tujebajeva, R.M.; Copeland, P.R.; Xu, X.M.; Carlson, B.A.; Harney, J.W.; Driscoll, D.M.; Hatfield, D.L.; Berry, M.J. Decoding apparatus for eukaryotic selenocysteine insertion. EMBO Rep. 2000, 1, 158–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilgenfeld, R.; Bock, A.; Wilting, R. Structural model for the selenocysteine-specific elongation factor selb. Biochimie 1996, 78, 971–978. [Google Scholar] [CrossRef]
- Dobosz-Bartoszek, M.; Pinkerton, M.H.; Otwinowski, Z.; Chakravarthy, S.; Soll, D.; Copeland, P.R.; Simonovic, M. Crystal structures of the human elongation factor EEFSec suggest a non-canonical mechanism for selenocysteine incorporation. Nat. Commun. 2016, 7, 12941. [Google Scholar] [CrossRef] [PubMed]
- Allmang, C.; Carbon, P.; Krol, A. The SBP2 and 15.5 kd/snu13p proteins share the same RNA binding domain: Identification of SBP2 amino acids important to secisRNA binding. RNA 2002, 8, 1308–1318. [Google Scholar] [CrossRef] [Green Version]
- Caban, K.; Kinzy, S.A.; Copeland, P.R. The l7ae rna binding motif is a multifunctional domain required for the ribosome-dependent sec incorporation activity of sec insertion sequence binding protein 2. Mol. Cell Biol. 2007, 27, 6350–6360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.E.; Copeland, P.R.; Driscoll, D.M.; Krol, A. The selenocysteine incorporation machinery: Interactions between the secis RNA and the secis-binding protein SBP2. RNA 2001, 7, 1442–1453. [Google Scholar]
- Zavacki, A.M.; Mansell, J.B.; Chung, M.; Klimovitsky, B.; Harney, J.W.; Berry, M.J. Coupled tRNA(sec)-dependent assembly of the selenocysteine decoding apparatus. Mol. Cell 2003, 11, 773–781. [Google Scholar] [CrossRef]
- Fradejas-Villar, N.; Seeher, S.; Anderson, C.B.; Doengi, M.; Carlson, B.A.; Hatfield, D.L.; Schweizer, U.; Howard, M.T. The RNA-binding protein SECISBP2 differentially modulates uga codon reassignment and RNA decay. Nucleic Acids Res. 2017, 45, 4094–4107. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.; Copeland, P.R. The selenocysteine-specific elongation factor contains unique sequences that are required for both nuclear export and selenocysteine incorporation. PLoS ONE 2016, 11, e0165642. [Google Scholar] [CrossRef]
- Donovan, J.; Copeland, P.R. Selenocysteine Insertion Sequence Binding Protein 2L is implicated as a novel post-transcriptional regulator of selenoprotein expression. PLoS ONE 2012, 7, e35581. [Google Scholar] [CrossRef] [Green Version]
- Copeland, P.R.; Stepanik, V.A.; Driscoll, D.M. Insight into mammalian selenocysteine insertion: Domain structure and ribosome binding properties of sec insertion sequence binding protein 2. Mol. Cell Biol. 2001, 21, 1491–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kossinova, O.; Malygin, A.; Krol, A.; Karpova, G. The SBP2 protein central to selenoprotein synthesis contacts the human ribosome at expansion segment 7l of the 28s rRNA. RNA 2014, 20, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Shen, Q.; Newburger, P.E. Recognition and binding of the human selenocysteine insertion sequence by nucleolin. J. Cell Biochem. 2000, 77, 507–516. [Google Scholar] [CrossRef]
- Miniard, A.C.; Middleton, L.M.; Budiman, M.E.; Gerber, C.A.; Driscoll, D.M. Nucleolin binds to a subset of selenoprotein mRNAs and regulates their expression. Nucleic Acids Res. 2010, 38, 4807–4820. [Google Scholar] [CrossRef] [Green Version]
- Bifano, A.L.; Atassi, T.; Ferrara, T.; Driscoll, D.M. Identification of nucleotides and amino acids that mediate the interaction between ribosomal protein L30 and the secis element. BMC Mol. Biol. 2013, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Halic, M.; Becker, T.; Frank, J.; Spahn, C.M.; Beckmann, R. Localization and dynamic behavior of ribosomal protein L30e. Nat. Struct. Mol. Biol. 2005, 12, 467–468. [Google Scholar] [CrossRef]
- Macias, S.; Bragulat, M.; Tardiff, D.F.; Vilardell, J. L30 binds the nascent rpl30 transcript to repress u2 snrnp recruitment. Mol. Cell 2008, 30, 732–742. [Google Scholar] [CrossRef]
- Chavatte, L.; Brown, B.A.; Driscoll, D.M. Ribosomal protein L30 is a component of the uga-selenocysteine recoding machinery in eukaryotes. Nat. Struct. Mol. Biol. 2005, 12, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Budiman, M.E.; Bubenik, J.L.; Driscoll, D.M. Identification of a signature motif for the eIF4a3-secis interaction. Nucleic Acids Res. 2011, 39, 7730–7739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraiuolo, M.A.; Lee, C.-S.; Ler, L.W.; Hsu, J.L.; Costa-Mattioli, M.; Luo, M.J.; Reed, R.; Sonenberg, N. A nuclear translation-like factor eIF4aiii is recruited to the mRNA during splicing and functions in nonsense-mediated decay. Proc. Natl. Acad. Sci. USA 2004, 101, 4118–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, I.M.; Gatfield, D.; St Johnston, D.; Izaurralde, E. An eif4aiii-containing complex required for mRNA localization and nonsense-mediated mRNA decay. Nature 2004, 427, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, T.; Tange, T.O.; Sonenberg, N.; Moore, M.J. Eif4aiii binds spliced mRNA in the exon junction complex and is essential for nonsense-mediated decay. Nat. Struct. Mol. Biol. 2004, 11, 346–351. [Google Scholar] [CrossRef]
- Sonenberg, N. EIF4e, the mRNA cap-binding protein: From basic discovery to translational research. Biochem. Cell Biol. 2008, 86, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Gribling-Burrer, A.S.; Leichter, M.; Wurth, L.; Huttin, A.; Schlotter, F.; Troffer-Charlier, N.; Cura, V.; Barkats, M.; Cavarelli, J.; Massenet, S.; et al. Secis-binding protein 2 interacts with the smn complex and the methylosome for selenoprotein mrnp assembly and translation. Nucleic Acids Res. 2017, 45, 5399–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurth, L.; Gribling-Burrer, A.S.; Verheggen, C.; Leichter, M.; Takeuchi, A.; Baudrey, S.; Martin, F.; Krol, A.; Bertrand, E.; Allmang, C. Hypermethylated-capped selenoprotein mRNAs in mammals. Nucleic Acids Res. 2014, 42, 8663–8677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.K.; Tisdale, S.; Lotti, F.; Pellizzoni, L. Smn control of rnp assembly: From post-transcriptional gene regulation to motor neuron disease. Semin. Cell Dev. Biol. 2014, 32, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.P.; Kiledjian, N.T.; Copeland, P.R. The polypyrimidine tract binding protein, PTBP1, regulates selenium homeostasis via the selenoprotein p 3’ untranslated region. bioRxiv 2020. [Google Scholar] [CrossRef]
- Losson, R.; Lacroute, F. Interference of nonsense mutations with eukaryotic messenger RNA stability. Proc. Natl. Acad. Sci. USA 1979, 76, 5134–5137. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Popp, M.W.; Maquat, L.E. Quality and quantity control of gene expression by nonsense-mediated mRNA decay. Nat. Rev. Mol. Cell Biol. 2019, 20, 406–420. [Google Scholar] [CrossRef]
- Novoselov, S.V.; Lobanov, A.V.; Hua, D.; Kasaikina, M.V.; Hatfield, D.L.; Gladyshev, V.N. A highly efficient form of the selenocysteine insertion sequence element in protozoan parasites and its use in mammalian cells. Proc. Natl. Acad. Sci. USA 2007, 104, 7857–7862. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.P.; Copeland, P.R. Selenocysteine incorporation: A trump card in the game of mRNA decay. Biochimie 2015, 114, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, S.P.; Copeland, P.R. The selenium transport protein, selenoprotein p, requires coding sequence determinants to promote efficient selenocysteine incorporation. J. Mol. Biol. 2018, 430, 5217–5232. [Google Scholar] [CrossRef]
- Doma, M.K.; Parker, R. Endonucleolytic cleavage of eukaryotic mRNAs with stalls in translation elongation. Nature 2006, 440, 561–564. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.; Cluet, D.; Ricci, E.P. Ribosome dynamics and mRNA turnover, a complex relationship under constant cellular scrutiny. Wiley Interdiscip. Rev. RNA 2021, 12, e1658. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Grosely, R.; Prabhakar, A.; Lapointe, C.P.; Wang, J.; Puglisi, J.D. How messenger RNA and nascent chain sequences regulate translation elongation. Annu. Rev. Biochem. 2018, 87, 421–449. [Google Scholar] [CrossRef]
- Brandman, O.; Stewart-Ornstein, J.; Wong, D.; Larson, A.; Williams, C.C.; Li, G.W.; Zhou, S.; King, D.; Shen, P.S.; Weibezahn, J.; et al. A ribosome-bound quality control complex triggers degradation of nascent peptides and signals translation stress. Cell 2012, 151, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Ikeuchi, K.; Saeki, Y.; Iwasaki, S.; Schmidt, C.; Udagawa, T.; Sato, F.; Tsuchiya, H.; Becker, T.; Tanaka, K.; et al. Ubiquitination of stalled ribosome triggers ribosome-associated quality control. Nat. Commun. 2017, 8, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeuchi, K.; Tesina, P.; Matsuo, Y.; Sugiyama, T.; Cheng, J.; Saeki, Y.; Tanaka, K.; Becker, T.; Beckmann, R.; Inada, T. Collided ribosomes form a unique structural interface to induce hel2-driven quality control pathways. EMBO J. 2019, 38, e100276. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copeland, P.R.; Howard, M.T. Ribosome Fate during Decoding of UGA-Sec Codons. Int. J. Mol. Sci. 2021, 22, 13204. https://doi.org/10.3390/ijms222413204
Copeland PR, Howard MT. Ribosome Fate during Decoding of UGA-Sec Codons. International Journal of Molecular Sciences. 2021; 22(24):13204. https://doi.org/10.3390/ijms222413204
Chicago/Turabian StyleCopeland, Paul R., and Michael T. Howard. 2021. "Ribosome Fate during Decoding of UGA-Sec Codons" International Journal of Molecular Sciences 22, no. 24: 13204. https://doi.org/10.3390/ijms222413204
APA StyleCopeland, P. R., & Howard, M. T. (2021). Ribosome Fate during Decoding of UGA-Sec Codons. International Journal of Molecular Sciences, 22(24), 13204. https://doi.org/10.3390/ijms222413204