
Zapotin, a Polymethoxyflavone, with Potential Therapeutic Attributes




Abstract
:1. Introduction
2. Methodology
3. Natural Occurrence of Zapotin
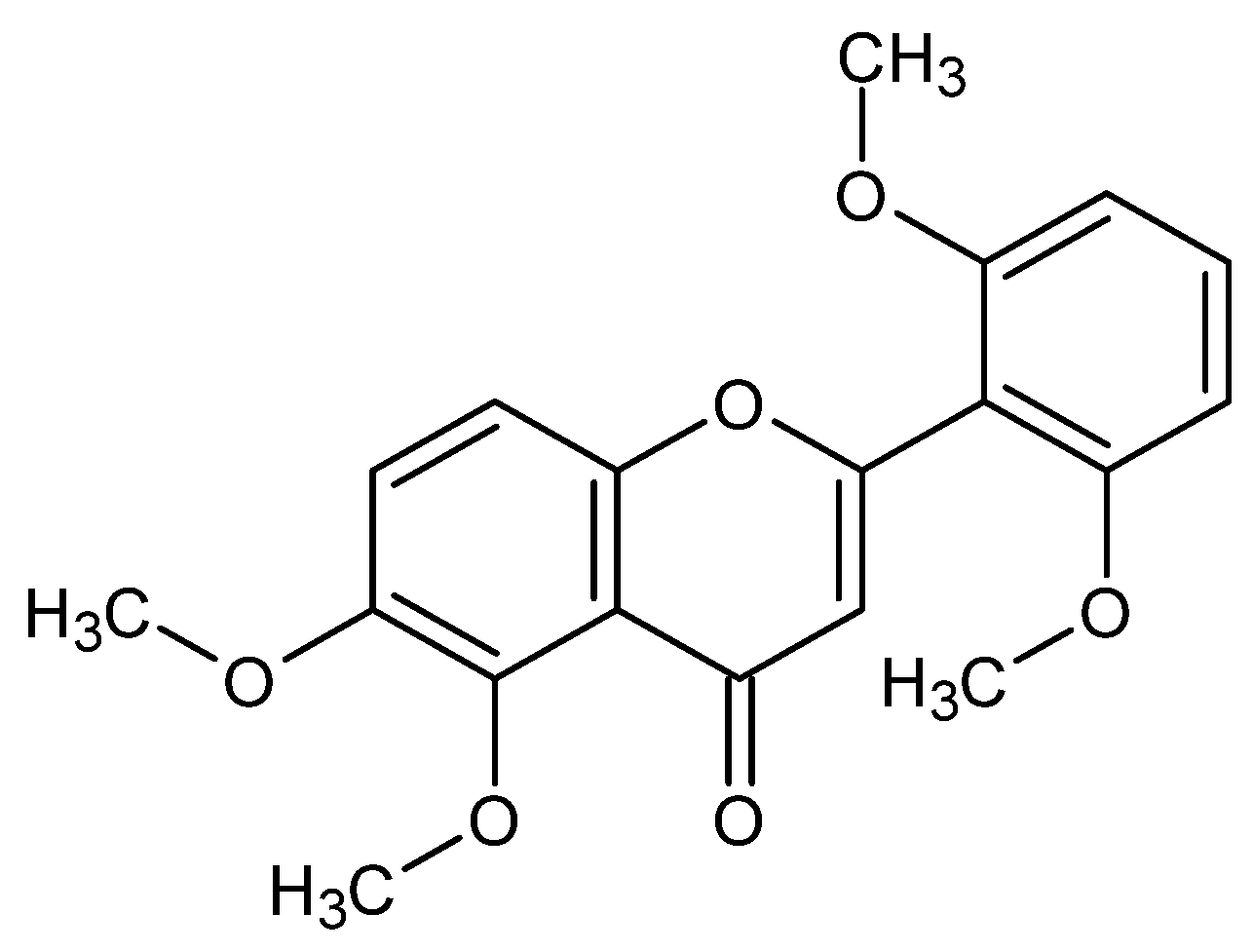
4. Structural Determination of Zapotin
5. Bio-Inspired Organic Synthesis of Zapotin
6. Spectroscopic Characterization of Zapotin
7. Techniques for the Analysis of Zapotin in Plant Material
8. Biological Activities of Zapotin
8.1. Antiviral Activity
8.2. Anticancer Activity
8.3. Antimutagenic Activity
8.4. Vasorelaxant Activity
8.5. Antimicrobial Activity
8.6. Antidepressant-like Activity
9. ADMET of Zapotin
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACF | aberrant crypt foci |
| AcOH | acetic acid |
| ADMET | absorption, distribution, metabolism, excretion, and toxicity |
| Al2O3 | aluminum oxide |
| BW | body weight |
| CC | column chromatography |
| CFU | colony-forming unit |
| CHCl3 | chloroform |
| Et2O | diethyl ether |
| EtOAc | ethyl acetate |
| F | film thickness |
| FST | forced swim test |
| HCl | hydrochloric acid |
| Hex | hexane |
| HPLC | high-performance liquid chromatography |
| HPLC-MS | high-performance liquid chromatography–mass spectrometry |
| HPMFs | hydroxypolymethoxyflavones |
| ID | inner diameter |
| IR | infrared radiation |
| L | length |
| LC-MS | liquid chromatography–mass spectrometry |
| LC-MS-MS | liquid chromatography–tandem mass spectrometry |
| LiHMDS | lithium hexamethyldisilazide |
| MeOH | methanol |
| MMOC | mouse mammary organ culture |
| mp | melting point |
| NMR | nuclear magnetic resonance spectroscopy |
| ODC | ornithine decarboxylase |
| PMFs | polymethoxyflavones |
| PKC | protein kinase C |
| Rf | retention factor |
| RP | reversed phase |
| SFC | supercritical fluid chromatography |
| t0.5 | time of half-life |
| THF | tetrahydrofurane |
| TPA | 12-O-tetradecanoylphorbol-13-acetate |
| UV | ultraviolet |
| VLC | vacuum liquid chromatography |
References
- Atanasov, A.G.; The International Natural Product Sciences Taskforce; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Rigano, F.; Arigò, A.; Dugo, P.; Mondello, L. Coumarins, psoralens and polymethoxyflavones in cold-pressed citrus essential oils: A Review. J. Essent. Oil Res. 2021, 33, 221–239. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Wei, C.-C.; Huang, J.; Pan, M.-H.; Shahidi, F.; Ho, C.-T. Anti-inflammatory effects of polymethoxyflavones from citrus peels: A review. J. Food Bioact. 2018, 3, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Pan, M.-H.; Lo, C.-Y.; Tan, D.; Wang, Y.; Shahidi, F.; Ho, C.-T. Chemistry and health effects of polymethoxyflavones and hydroxylated polymethoxyflavones. J. Funct. Foods 2009, 1, 2–12. [Google Scholar] [CrossRef]
- Lai, C.-S.; Li, S.; Chai, C.-Y.; Lo, C.-Y.; Ho, C.-T.; Wang, Y.-J.; Pan, M.-H. Inhibitory effect of citrus 5-hydroxy-3,6,7,8,3’,4’-hexamethoxyflavone on 12-O-tetradecanoylphorbol 13-acetate-induced skin inflammation and tumor promotion in mice. Carcinogenesis 2007, 28, 2581–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lambros, T.; Wang, Z.; Goodnow, R.; Ho, C.-T. Efficient and scalable method in isolation of polymethoxyflavones from orange peel extract by supercritical fluid chromatography. J. Chromatogr. B 2007, 846, 291–297. [Google Scholar] [CrossRef]
- Xu, Y.M.; del Ramirez-Ahumada, M.C.; Valeriote, F.A.; Gunatilaka, A.A.L. Solid tumor inhibitory and other constituents of Casimiroa tetrameria. Chin. J. Nat. Med. 2011, 9, 334–337. [Google Scholar] [CrossRef]
- Kincl, F.A.; Romo, J.; Rosenkranz, G.; Sondheimer, F. The constituents of Casimiroa edulis llave et lex. Part I. The seed. J. Chem. Soc. 1956, 804, 4163–4169. [Google Scholar] [CrossRef]
- Toton, E.; Romaniuk-Drapała, A.; Budzianowski, J.; Hofmann, J.; Rybczynska, M. Zapotin (5,6,2′,6′-tetramethoxyflavone) modulates the crosstalk between autophagy and apoptosis pathways in cancer cells with overexpressed constitutively active PKCϵ. Nutr. Cancer 2016, 68, 290–304. [Google Scholar] [CrossRef]
- Jakimiuk, K.; Wink, M.; Tomczyk, M. Flavonoids of the Caryophyllaceae. Phytochem. Rev. 2021, 20, 1–40. [Google Scholar] [CrossRef]
- FAO. Forest Resources Development Branch Some Medicinal Forest Plants of Africa and Latin America; Food and Agriculture Organization of the United Nations: Rome, Italy, 1986; pp. 63–68. [Google Scholar]
- Hartwich, C.; Bickern, W. Beitrag zur Kenntnis der Casimiroa edulis La Llave. Arch. Pharm. 1903, 241, 166–176. [Google Scholar] [CrossRef] [Green Version]
- El Sissi, H.I.; El Sissi, A.A.; Saleh, N.A.M. Local plants as potential sources of tannins. Plant Foods Hum. Nutr. 1965, 12, 390–396. [Google Scholar] [CrossRef]
- Zavaleta-Mancera, H.A. Anatomía de la semilla de Casimiroa edulis (Rutaceae), “zapote blanco”, durante su desarrollo. Bot. Sci. 2017, 51, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.F. The drug aspects of the white sapotes. Econ. Bot. 1962, 16, 288–294. [Google Scholar] [CrossRef]
- Yahia, E.M.; Gutierrez-Orozco, F. White sapote (Casimiroa edulis Llave & Lex). In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Yahia, E.M., Ed.; Woodhead Publishing Limited: Sawston, UK, 2011; Volume 4, pp. 474–482. [Google Scholar]
- Tun, K.N.W.; Aminah, N.S.; Kristanti, A.N.; Aung, H.T.; Takaya, Y. Natural products isolated from Casimiroa. Open Chem. 2020, 18, 778–797. [Google Scholar] [CrossRef]
- Power, F.B.; Callan, T. CCXXVI.—The constituents of the seeds of Casimiroa edulis. J. Chem. Soc. Trans. 1911, 99, 1993–2010. [Google Scholar] [CrossRef]
- Heneka, B.; Rimpler, H.; Ankli, A.; Sticher, O.; Gibbons, S.; Heinrich, M. A furanocoumarin and polymethoxylated flavonoids from the Yucatec Mayan plant Casimiroa tetrameria. Phytochemistry 2005, 66, 649–652. [Google Scholar] [CrossRef]
- Heinrich, M.; Heneka, B.; Rimpler, H.; Ankli, A.; Sticher, O.; Kostiza, T. Spasmolytic and antidiarrhoeal properties of the Yucatec Mayan medicinal plant Casimiroa tetrameria. J. Pharm. Pharmacol. 2010, 57, 1081–1085. [Google Scholar] [CrossRef]
- Iriarte, J.; Kincl, F.A.; Rosenkranz, G.; Sondheimer, F. The constituents of Casimiroa edulis Lluve et Lex. Part II.* The bark. J. Chem. Soc. 1956, 6, 951–952. [Google Scholar]
- Garratt, P.; Scheinmann, F.; Sondheimer, F. Constituents of Casimiroa edulis Llaveet Lex.—VIII. Tetrahedron 1967, 23, 2413–2416. [Google Scholar] [CrossRef]
- Dreyer, D.; Bertelli, D. The structure of zapotin. Tetrahedron 1967, 23, 4607–4612. [Google Scholar] [CrossRef]
- Miller, S.L.; Haber, W.A.; Setzer, W.N. Chemical composition of the leaf essential oil of Casimiroa edulis La Llave & Lex. (Rutaceae) from Monteverde, Costa Rica. Nat. Prod. Commun. 2009, 4, 425–426. [Google Scholar] [CrossRef] [Green Version]
- Mahjoub, J.M.S. UV Study on the Major Flavonoid from Zingiber officinale (Ginger). Master’s Thesis, Sudan University of Science and Technology, Khartoum, Sudan, 2015. [Google Scholar]
- Verma, A.K.; Pratap, R. The biological potential of flavones. Nat. Prod. Rep. 2010, 27, 1571–1593. [Google Scholar] [CrossRef] [PubMed]
- Awaad, A.; El-Sayed, N.; Maitland, D.; Mabry, T. Phenolic Antioxidants from Casimiroa edulis. Leaves. Pharm. Biol. 2006, 44, 258–262. [Google Scholar] [CrossRef]
- Nagai, H.; Tanaka, T.; Goto, T.; Kusudo, T.; Takahashi, N.; Kawada, T. Phenolic compounds from leaves of Casimiroa edulis showed adipogenesis activity. Biosci. Biotechnol. Biochem. 2014, 78, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Awaad, A.S.; Al-Jaber, N.A.; Soliman, G.A.; Al-Outhman, M.R.; Zain, M.E.; Moses, J.E.; El-Meligy, R.M. New Biological Activities of Casimiroa edulis Leaf Extract and Isolated Compounds. Phytother. Res. 2011, 26, 452–457. [Google Scholar] [CrossRef]
- Satheesh, N. Review on distribution, nutritional and medicinal values of Casimiroa edulis Llave—An underutilized fruit in Ethiopia. Am. Eurasian J. Agric. Environ. Sci. 2015, 15, 1574–1583. [Google Scholar] [CrossRef]
- Ramaa, C.S.; Tilekar, K.N.; Patil, V.M. Liver cancer: Different approaches for targeting. Int. J. Pharmtech. Res. 2010, 2, 834–842. [Google Scholar]
- Machado, A.P.D.F.; Geraldi, M.V.; Nascimento, R.D.P.D.; Moya, A.M.T.M.; Vezza, T.; Diez-Echave, P.; Gálvez, J.J.; Cazarin, C.B.B.; Júnior, M.R.M. Polyphenols from food by-products: An alternative or complementary therapy to IBD conventional treatments. Food Res. Int. 2021, 140, 110018. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.N.; Wall, M.E.; Wani, M.C.; Taylor, H.L. Plant Antitumor Agents, 21. Flavones, coumarins, and an alkaloid from Sargentia greggii. J. Nat. Prod. 1985, 48, 952–956. [Google Scholar] [CrossRef]
- Froldi, G.; Bertin, R.; Secchi, E.; Zagotto, G.; Martínez-Vázquez, M.; García-Argáez, A. Vasorelaxation by extracts of Casimiroa spp. in rat resistance vessels and pharmacological study of cellular mechanisms. J. Ethnopharmacol. 2011, 134, 637–643. [Google Scholar] [CrossRef]
- Bertin, R.; Argaez, A.N.G.; Martínez-Vázquez, M.; Froldi, G. Age-dependent vasorelaxation of Casimiroa edulis and Casimiroa pubescens extracts in rat caudal artery in vitro. J. Ethnopharmacol. 2011, 137, 934–936. [Google Scholar] [CrossRef]
- Oyebode, O.T.; Owumi, S.E.; Oyelere, A.K.; Olorunsogo, O.O. Calliandra portoricensis Benth exhibits anticancer effects via alteration of Bax/Bcl-2 ratio and growth arrest in prostate LNCaP cells. J. Ethnopharmacol. 2019, 233, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Agunu, A.; Abdurahman, E.; Shok, M.; Yusuf, S.A. Analgesic activity of the roots and leaves extracts of Calliandra portoricensis. Fitoterapia 2005, 76, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, M.; Zampini, I.; Alberto, M.; Cuello, S.; Torres, S.; Pérez, J.; Quispe, C.; Hirschmann, G.S.; Isla, M. Polyphenols rich fraction from Geoffroea decorticans fruits flour affects key enzymes involved in metabolic syndrome, oxidative stress and inflammatory process. Food Chem. 2016, 190, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Isla, M.I.; Ezquer, M.E.; Leal, M.; Moreno, M.A.; Zampini, I.C. Flower beverages of native medicinal plants from Argentina (Acacia caven, Geoffroea decorticans and Larrea divaricata) as antioxidant and anti-inflammatory. J. Ethnopharmacol. 2021, 281, 114490. [Google Scholar] [CrossRef]
- Strawa, J.; Szoka, Ł.; Tomczyk, M. Preliminary studies of the chemical composition and cytotoxic activity of Hottonia palustris herb. Planta Med. 2019, 85, 1533. [Google Scholar] [CrossRef]
- Ramesh, S.; Rajan, R.; Santhanam, R. Freshwater Phytopharmaceutical Compounds, 1st ed.; CRC Press: Boca Raton, FL, USA, 2013; p. 201. [Google Scholar]
- Shastri, S.L.; Krishna, V.; Ravi Kumar, S.; Pradeepa, K. Phytochemical analysis, antibacterial property and molecular docking studies of Mammea suriga Kosterm. World J. Pharm. Res. 2016, 4, 331–340. [Google Scholar]
- Bhat, P.; Hegde, G.R.; Hegde, G.; Mulgund, G.S. Ethnomedicinal plants to cure skin diseases—An account of the traditional knowledge in the coastal parts of Central Western Ghats, Karnataka, India. J. Ethnopharmacol. 2014, 151, 493–502. [Google Scholar] [CrossRef]
- El-Seideek, L.; Zaied, S.F.; Hassan, M.I.; Elgammal, M.H. Antimicrobial, biochemical, organoleptic and stability properties of cookies fortified by pomegranate juice during storage. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 288–299. [Google Scholar]
- Arun, N.; Road, R.; Pradesh, U. Punica granatum: A review on pharmacological and therapeutic properties. Int. J. Pharm. Sci. Res. 2012, 3, 1240–1245. [Google Scholar]
- Budzianowski, J.; Morozowska, M.; Wesołowska, M. Lipophilic flavones of Primula veris L. from field cultivation and in vitro cultures. Phytochemistry 2005, 66, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Tarapatskyy, M.; Gumienna, A.; Sowa, P.; Kapusta, I.; Puchalski, C. Bioactive phenolic compounds from Primula veris L.: Influence of the extraction conditions and purification. Molecules 2021, 26, 997. [Google Scholar] [CrossRef] [PubMed]
- Ayers, S.; Zink, D.L.; Mohn, K.; Powell, J.S.; Brown, C.M.; Murphy, T.; Brand, R.; Pretorius, S.; Stevenson, D.; Thompson, D.; et al. Flavones from Struthiola argentea with anthelmintic activity in vitro. Phytochemistry 2008, 69, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Yugandhar, P.; Savithramma, N. Spectroscopic and chromatographic exploration of different phytochemical and mineral contents from Syzygium alternifolim (Wt.) Walp. an endemic, endangered medicinal tree taxon. J. Appl. Pharm. Sci. 2017, 7, 073–085. [Google Scholar] [CrossRef] [Green Version]
- European Pharmacopoeia, 10th ed.; European Directorate for the Quality of Medicines and HealthCare of the Council of Europe (EDQM): Strasbourg, France, 2019; p. 1588.
- Sondheimer, F.; Meisels, A. Constituents of Casimiroa edulis Llave et Lex.—VI: 2′,5,6-trimethoxyflavone, 2′,5,6,7-tetramethoxyflavone (zapotin) and 5-hydroxy-2′,6,7-trimethoxyflavone (zapotinin). Tetrahedron 1960, 9, 139–144. [Google Scholar] [CrossRef]
- Farkas, L.; Nógrádi, M. Bemerkungen zur Konstitution des Zapotins und Zapotinins, I. Synthese einiger in 2′-Stellung substituierter Flavonoide. Eur. J. Inorg. Chem. 1965, 98, 164–170. [Google Scholar] [CrossRef]
- Pai, B.; Subramaniam, P.; Subramanyam, V. 2′,5,6,7-tetramethoxyflavone and the structure of zapotin. Tetrahedron 1965, 21, 3573–3574. [Google Scholar] [CrossRef]
- Farkas, L.; Gottsegen, A.; Nogradi, M. On the structure of zapotin and zapotinin. II.: The synthesis of zapotin. Tetrahedron Lett. 1968, 9, 3993–3996. [Google Scholar] [CrossRef]
- Murillo, G.; Hirschelman, W.H.; Ito, A.; Moriarty, R.M.; Kinghorn, A.D.; Pezzuto, J.M.; Mehta, R.G. Zapotin, a phytochemical present in a Mexican fruit, prevents colon carcinogenesis. Nutr. Cancer 2007, 57, 28–37. [Google Scholar] [CrossRef]
- Maiti, A.; Cuendet, M.; Kondratyuk, T.; Croy, V.L.; Pezzuto, J.M.; Cushman, M. Synthesis and cancer chemopreventive activity of zapotin, a natural product from Casimiroa edulis. J. Med. Chem. 2007, 50, 350–355. [Google Scholar] [CrossRef] [Green Version]
- Yandamuri, N.; Nagabathula, S.; Kurra, S.; Batthula, S.; Allada, N.; Bandam, P. Comparative study of new trends in HPLC. Int. J. Pharm. Sci. Rev. Res. 2013, 23, 52–57. [Google Scholar]
- Ito, A.; Shamon, L.A.; Yu, B.; Mata-Greenwood, E.; Lee, S.K.; van Breemen, R.B.; Mehta, R.G.; Farnsworth, N.R.; Fong, H.H.S.; Pezzuto, J.M.; et al. Antimutagenic constituents of Casimiroa edulis with potential cancer chemopreventive activity. J. Agric. Food Chem. 1998, 46, 3509–3516. [Google Scholar] [CrossRef]
- Argaez, A.N.G.; González-Lugo, N.M.; Parra-Delgado, H.; Martínez-Vázquez, M. Casimiroin, zapoterin, zapotin and 5,6,2′,3′,4′-pentamethoxyflavone from Casimiroa pubescens. Biochem. Syst. Ecol. 2005, 33, 441–443. [Google Scholar] [CrossRef]
- Ubaldo-Suárez, D.; Reyes, R.E.; De La Rosa-Sierra, R.; Martínez-Vázquez, M. Antidepressant-like effect of Casimiroa pubescens root extracts. Nat. Prod. Res. 2018, 33, 2526–2530. [Google Scholar] [CrossRef]
- Bailly, C. The subgroup of 2′-hydroxy-flavonoids: Molecular diversity, mechanism of action, and anticancer properties. Bioorg. Med. Chem. 2021, 32, 116001. [Google Scholar] [CrossRef]
- Juszczak, A.M.; Zovko-Končić, M.; Tomczyk, M. Recent trends in the application of chromatographic techniques in the analysis of luteolin and its derivatives. Biomolecules 2019, 9, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, V.; Jaiswal, N.; Srivastava, M. A review on medicinal properties of Centella asiatica. Asian J. Pharm. Clin. Res. 2017, 10, 69. [Google Scholar] [CrossRef] [Green Version]
- Esposito, F.; Zinzula, L.; Maxia, A.; Tramontano, E.; Sanna, C. Inhibition of HIV-1 reverse transcriptase associated activities by the hydroalcoholic extract of Casimiroa edulis seeds. Nat. Prod. Res. 2011, 25, 1067–1073. [Google Scholar] [CrossRef]
- Mata-Greenwood, E.; Ito, A.; Westenburg, H.; Cui, B.; Mehta, R.G.; Kinghorn, A.D.; Pezzuto, J.M. Discovery of novel inducers of cellular differentiation using HL-60 promyelocytic cells. Anticancer Res. 2001, 21, 1763–1770. [Google Scholar]
- Kanade, G.G.; Nathani, R.; Huma, L.; Mehta, R.G.; Murillo, G. Zapotin mediates its action by inducing P53 in wild-type P53 positive human lung cancer cells. In Proceeding of the AACR 101st Annual Meeting, Washington, DC, USA, 17–21 April 2010; p. 5694. [Google Scholar] [CrossRef]
- Cuendet, M.; Oteham, C.P.; Maiti, A.; A Craig, B.; Cushman, M.; Moon, R.C.; Pezzuto, J.M. Zapotin prevents mouse skin tumorigenesis during the stages of initiation and promotion. Anticancer Res. 2009, 28, 3705–3709. [Google Scholar]
- Toton, E.; Lisiak, N.; Rubis, B.; Budzianowski, J.; Gruber, P.; Hofmann, J.; Rybczynska, M. The tetramethoxyflavone zapotin selectively activates protein kinase C epsilon, leading to its down-modulation accompanied by Bcl-2, c-Jun and c-Fos decrease. Eur. J. Pharmacol. 2012, 682, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J. Studies of Metabolism and Disposition of Natural Products Using Mass Spectrometry. Ph.D. Thesis, University of Illinois, Chicago, IL, USA, 2010. [Google Scholar]


{kind=link}
| Species | Family | Location | Parts of Plant | Traditional Uses | References |
|---|---|---|---|---|---|
| Casimiroa edulis | Rutaceae | not given | bark, seeds, leaves | hypertension, anxiety, insomnia, painkiller | [8,11,13,17,21,22,23,24,25,26,27,28,29,30,31,32] |
| Casimiroa greggii | Rutaceae | Mexico | root | not found | [33] |
| Casimiroa pubescens | Rutaceae | Mexico | seeds | antihypertensive | [34,35] |
| Casimiroa tetrameria | Rutaceae | Mexico | leaves | gastrointestinal problems | [19,20] |
| Calliandra portoricensis | Fabaceae | Nigeria | root bark | lumbago, pain, gonorrhea, anticonvulsant, antimicrobial, analgesic, antidepressant | [36,37] |
| Geoffroea decorticans | Fabaceae | Argentina | fruits | dysentery, expectorant agent, flatulence | [38,39] |
| Hottonia palustris | Primulaceae | Poland | herb | heart problems | [40,41] |
| Mammea suriga | Calophyllaceae | India | stem bark | eczema | [42,43] |
| Punica granatum | Punicaceae | Egypt | peel extract | inflammation, rheumatism, sore throat, snakebite, diabetes, burns | [44,45] |
| Primula veris | Primulaceae | Poland | leaves a | respiratory, cardiac, nervous system conditions, expectorant, sedative | [46,47] |
| Struthiola argentea | Thymelaeaceae | South Africa | herb | not found | [48] |
| Syzygium alternifolium | Myrtaceae | India | fruit | gastrointestinal problems, painkiller | [49] |
| Plant | Part of the Plant | Extract | Separation Conditions | References |
|---|---|---|---|---|
| Casimiroa edulis | seeds, trunk, and root bark | EtOH |
| [8,21] |
| MeOH |
| [58] | ||
| Casimiroa pubescens | seeds | EtOAc |
| [59,60] |
| Casimiroa tetrameria | leaves | EtOH |
| [19] |
| Primula veris | leaves | CHCl3 |
| [45,61] |
| Struthiola argentea | herb | MeOH |
| [48] |
| Plant | Part of the Plant | Extract | Column | Mobile Phase | Conditions | References |
|---|---|---|---|---|---|---|
| Casimiroa pubescens | seeds | MeOH | Restek Pinnacle C18 | 5% AcOH and H2O:MeOH | 30–90% B: 0–50 min, 90% B: 50–55 min | [34] |
| Casimiroa portoricensis | root bark | MeOH | not given | not given | not given | [36] |
| Struthiola argentea | herb | Hex/CH2Cl2 | Zorbax RX-C8, Zorbax RX-C18 | 0.1% THF and H2O:ACN | 25–100% | [48] |
| Plant | Part of the Plant | Extract | Column a | Conditions | References |
|---|---|---|---|---|---|
| Casimiroa pubescens | root | not given | not given (0.25 × 30.0 × 0.25) | 30 to 310 °C; 8 °C/min; 6 min at 310 °C | [60] |
| Syzygium alternifolim | fruit | MeOH | VF-5MS (0.25 × 30.0 × 0.25) | 3 min at 70 °C; 10 °C/min to 240 °C, then 5 °C/min to 300 °C, 9 min at 300 °C | [49] |
| Mammea suriga | stem bark | petroleum ether | RESTEK Rtx-5 (0.25 × 30.0 × 0.25) | 3 min at 70 °C; 10 °C/min to 240 °C, then 5 °C/min to 300 °C, 9 min at 300 °C | [42] |
| Punica granatum | peel | not given | Agilent HP-5ms (0.25 × 30.0 × 0.25) | 2 min at 60 °C; 10 °C/min to 280 °C | [44] |
| Activity | Experimental Model | Exposure/ Incubation | Concentration | Efficacy | References |
|---|---|---|---|---|---|
| Anti-viral | RDDP5 assay | not given | 70% ethanolic extracts from C. edulis |
| [64] |
| Anti-cancer | K562 cell line | not given | 70% ethanolic extracts from C. edulis |
| [64] |
| HL-60 cell line | not given | not given |
| [65] | |
| LNCaP, DU-145, lung adenocarcinoma, healthy VERO cell lines | 24 h incubation with extracts, 72 h at 37 °C with medium, 2–4 h at 37 °C with MTS | MeOH fraction of C. portoricensis |
| [36] | |
| A549 cell line | not given | Zapotin, MeOH extract of C. portoricensis |
| [66] | |
| disk diffusion assay for cytotoxicity, Colon38, L1210, MCF-7, U251N, PANC-1, H-116 cell lines | not given | 1 μg/disk |
| [7] | |
| female CD-1 mice, 4 weeks old | 15 weeks | 1, 5, and 10 μmol/mouse |
| [67] | |
| HeLaPKCεA/E subline |
|
|
| [9,68] | |
| SW480, SW620,HT-29 cell lines, female CF-1 mice |
| 5.0 or 10.0 mg/kg BW in the induction of ACF, 1.0 µM zapotin in flow cytometry |
| [55] | |
| ODC, HepG2 and HL-60 cell lines |
| 12 μM zapotin in the quantification of apoptosis |
| [56] | |
| Anti- mutagenic | Aroclor 1254-pretreated rats | not given | not given |
| [58] |
| Vasorelaxant | adhering tissue and arterial rings from 6-month-old rats | 45 min before viability | Casimiroa spp. extracts, 20 μg/mL per se |
| [34] |
| Antimicrobial | suspensions of microorganisms containing 106 CFU/mL | 72 h at 37 °C | water juice peel from Punica granatum |
| [44] |
| Anti- depressant-like | mice | first dose 60 min before testing, second dose 24, 7, and 1 h before testing | hexane (HCP), ethyl acetate (ECP), and methanol (MCP) extracts from C. pubescens |
| [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strawa, J.W.; Jakimiuk, K.; Tomczyk, M. Zapotin, a Polymethoxyflavone, with Potential Therapeutic Attributes. Int. J. Mol. Sci. 2021, 22, 13227. https://doi.org/10.3390/ijms222413227
Strawa JW, Jakimiuk K, Tomczyk M. Zapotin, a Polymethoxyflavone, with Potential Therapeutic Attributes. International Journal of Molecular Sciences. 2021; 22(24):13227. https://doi.org/10.3390/ijms222413227
Chicago/Turabian StyleStrawa, Jakub W., Katarzyna Jakimiuk, and Michał Tomczyk. 2021. "Zapotin, a Polymethoxyflavone, with Potential Therapeutic Attributes" International Journal of Molecular Sciences 22, no. 24: 13227. https://doi.org/10.3390/ijms222413227
APA StyleStrawa, J. W., Jakimiuk, K., & Tomczyk, M. (2021). Zapotin, a Polymethoxyflavone, with Potential Therapeutic Attributes. International Journal of Molecular Sciences, 22(24), 13227. https://doi.org/10.3390/ijms222413227