
A Meta-Analysis of Human Transcriptomics Data in the Context of Peritoneal Dialysis Identifies Novel Receptor-Ligand Interactions as Potential Therapeutic Targets


Abstract
:1. Introduction
2. Results and Discussion
2.1. Transcriptomics Studies Included in Our Meta-Analysis
2.2. Differentially Expressed Genes
2.3. Functional Analysis—Gene Set Enrichment Analysis
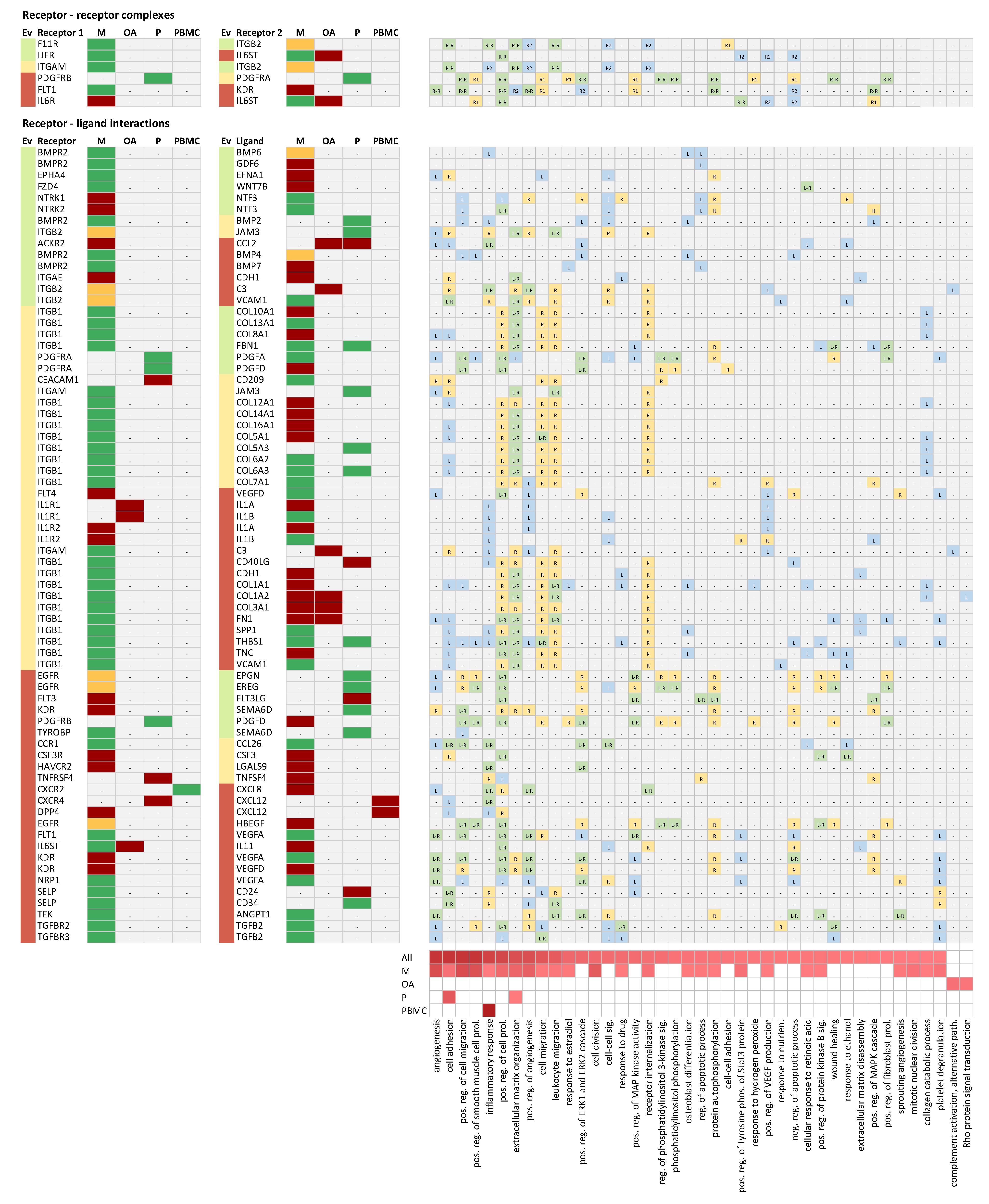
2.4. Receptor-Ligand Interaction Analysis
2.4.1. Receptor-Receptor Complex between Leukemia Inhibitory Factor Receptor (LIFR) and IL6ST
2.4.2. Receptor-Ligand Interaction between the Atypical Chemokine Receptor 2 (ACKR2) and C-C motif Chemokine Ligand 2 (CCL2)
2.4.3. Receptor-Ligand Interactions between Bone Morphogenic Protein Receptor Type 2 (BMPR2) and Growth Differentiation Factor 6 (GDF6) as Well as Bone Morphogenic Protein 6 (BMP6)
2.4.4. Receptor-Ligand Interactions between the Epidermal Growth Factor Receptor (EGFR) and Epiregulin (EREG) as Well as the Epithelial Mitogen (EPGN)
2.4.5. Receptor-Ligand Interaction between Frizzled Class Receptor 4 (FZD4) and Wnt Family Member 7B (WNT7B)
2.4.6. Receptor-Ligand Interactions between KDR and Semaphorin 6D (SEMAD6) as Well as TYROBP and SEMAD6
2.4.7. Receptor-Receptor Complex between F11R and ITGB2
2.4.8. Receptor-Ligand Interaction between EPH Receptor A4 (EPHA4) and Ephrin A1 (EFNA1)
2.4.9. Receptor-Ligand Interactions between NTRK1 and NTF3 as Well as NTRK2 and NTF3
2.5. Study Limitations and Planned Next Steps
3. Materials and Methods
3.1. Defining the Set of Eligible Transcriptomics Studies in the Context of Peritoneal Dialysis for the Current Meta-Analysis
3.2. Consolidating the Sets of Differentially Expressed Genes (DEGs)
3.3. Functional Gene Set Enrichment Analysis
3.4. Receptor-Ligand Interactome Analysis
3.5. Statistical Analysis and Data Manipulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Witowski, J.; López-Cabrera, M. Peritoneal Dialysis and Its Local and Systemic Complications: From the Bench to the Clinic. Front. Physiol. 2020, 11, 188. [Google Scholar] [CrossRef]
- Bartosova, M.; Schmitt, C.P. Biocompatible Peritoneal Dialysis: The Target Is Still Way Off. Front. Physiol. 2018, 9, 1853. [Google Scholar] [CrossRef]
- Herzog, R.; Sacnun, J.M.; González-Mateo, G.; Bartosova, M.; Bialas, K.; Wagner, A.; Unterwurzacher, M.; Sobieszek, I.J.; Daniel-Fischer, L.; Rusai, K.; et al. Lithium preserves peritoneal membrane integrity by suppressing mesothelial cell αB-crystallin. Sci. Transl. Med. 2021, 13, eaaz9705. [Google Scholar] [CrossRef]
- Büchel, J.; Bartosova, M.; Eich, G.; Wittenberger, T.; Klein-Hitpass, L.; Steppan, S.; Hackert, T.; Schaefer, F.; Passlick-Deetjen, J.; Schmitt, C.P. Interference of peritoneal dialysis fluids with cell cycle mechanisms. Perit. Dial. Int. 2015, 35, 259–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, R.; Kuster, L.; Becker, J.; Gluexam, T.; Pils, D.; Spittler, A.; Bhasin, M.K.; Alper, S.L.; Vychytil, A.; Aufricht, C.; et al. Functional and Transcriptomic Characterization of Peritoneal Immune-Modulation by Addition of Alanyl-Glutamine to Dialysis Fluid. Sci. Rep. 2017, 7, 6229. [Google Scholar] [CrossRef] [Green Version]
- Zaza, G.; Pontrelli, P.; Pertosa, G.; Granata, S.; Rossini, M.; Porreca, S.; Staal, F.J.T.; Gesualdo, L.; Grandaliano, G.; Schena, F.P. Dialysis-related systemic microinflammation is associated with specific genomic patterns. Nephrol. Dial. Transplant. 2008, 23, 1673–1681. [Google Scholar] [CrossRef] [Green Version]
- Reimold, F.R.; Braun, N.; Zsengellér, Z.K.; Stillman, I.E.; Karumanchi, S.A.; Toka, H.R.; Latus, J.; Fritz, P.; Biegger, D.; Segerer, S.; et al. Transcriptional patterns in peritoneal tissue of encapsulating peritoneal sclerosis, a complication of chronic peritoneal dialysis. PLoS ONE 2013, 8, e56389. [Google Scholar] [CrossRef] [Green Version]
- Scherer, A.; Günther, O.P.; Balshaw, R.F.; Hollander, Z.; Wilson-McManus, J.; Ng, R.; McMaster, W.R.; McManus, B.M.; Keown, P.A. Alteration of human blood cell transcriptome in uremia. BMC Med. Genom. 2013, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Kokoroishi, K.; Nakashima, A.; Doi, S.; Ueno, T.; Doi, T.; Yokoyama, Y.; Honda, K.; Kanawa, M.; Kato, Y.; Kohno, N.; et al. High glucose promotes TGF-β1 production by inducing FOS expression in human peritoneal mesothelial cells. Clin. Exp. Nephrol. 2016, 20, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Carpio, V.; Sandoval, P.; Aguilera, A.; Albar-Vizcaíno, P.; Perez-Lozano, M.L.; González-Mateo, G.T.; Acuña-Ruiz, A.; García-Cantalejo, J.; Botías, P.; Bajo, M.A.; et al. Genomic reprograming analysis of the Mesothelial to Mesenchymal Transition identifies biomarkers in peritoneal dialysis patients. Sci. Rep. 2017, 7, 44941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosova, M.; Schaefer, B.; Bermejo, J.L.; Tarantino, S.; Lasitschka, F.; Macher-Goeppinger, S.; Sinn, P.; Warady, B.A.; Zaloszyc, A.; Parapatics, K.; et al. Complement Activation in Peritoneal Dialysis-Induced Arteriolopathy. J. Am. Soc. Nephrol. 2018, 29, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Ryu, H.-M.; Suh, J.; Lee, K.-J.; Choi, S.-Y.; Choi, S.; Kim, Y.-L.; Huh, J.Y.; Ha, H. Network-based integrated analysis of omics data reveal novel players of TGF-β1-induced EMT in human peritoneal mesothelial cells. Sci. Rep. 2019, 9, 1497. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Feng, S.; Dairi, G.; Guan, Q.; Chafeeva, I.; Wang, H.; Liggins, R.; da Roza, G.; Kizhakkedathu, J.N.; Du, C. Transcriptome analysis of signaling pathways of human peritoneal mesothelial cells in response to different osmotic agents in a peritoneal dialysis solution. BMC Nephrol. 2019, 20, 181. [Google Scholar] [CrossRef] [PubMed]
- Parikova, A.; Hruba, P.; Krejcik, Z.; Stranecky, V.; Franekova, J.; Krediet, R.T.; Viklicky, O. Peritoneal dialysis induces alterations in the transcriptome of peritoneal cells before detectible peritoneal functional changes. Am. J. Physiol. Ren. Physiol. 2020, 318, F229–F237. [Google Scholar] [CrossRef]
- Strippoli, R.; Sandoval, P.; Moreno-Vicente, R.; Rossi, L.; Battistelli, C.; Terri, M.; Pascual-Antón, L.; Loureiro, M.; Matteini, F.; Calvo, E.; et al. Caveolin1 and YAP drive mechanically induced mesothelial to mesenchymal transition and fibrosis. Cell Death Dis. 2020, 11, 647. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Vento-Tormo, M.; Teichmann, S.A.; Vento-Tormo, R. CellPhoneDB: Inferring cell-cell communication from combined expression of multi-subunit ligand-receptor complexes. Nat. Protoc. 2020, 15, 1484–1506. [Google Scholar] [CrossRef]
- Taniguchi, T.; Miyagawa, T.; Tamaki, Z.; Nakamura, K.; Yamashita, T.; Saigusa, R.; Takahashi, T.; Toyama, T.; Ichimura, Y.; Yoshizaki, A.; et al. A possible implication of reduced levels of LIF, LIFR, and gp130 in vasculopathy related to systemic sclerosis. Arch. Dermatol. Res. 2017, 309, 833–842. [Google Scholar] [CrossRef]
- Korotaeva, A.A.; Samoilova, E.V.; Chepurnova, D.A.; Zhitareva, I.V.; Shuvalova, Y.A.; Prokazova, N. V Soluble glycoprotein 130 is inversely related to severity of coronary atherosclerosis. Biomarkers 2018, 23, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Moreno Velásquez, I.; Golabkesh, Z.; Källberg, H.; Leander, K.; de Faire, U.; Gigante, B. Circulating levels of interleukin 6 soluble receptor and its natural antagonist, sgp130, and the risk of myocardial infarction. Atherosclerosis 2015, 240, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Zolk, O.; Ng, L.L.; O’Brien, R.J.; Weyand, M.; Eschenhagen, T. Augmented expression of cardiotrophin-1 in failing human hearts is accompanied by diminished glycoprotein 130 receptor protein abundance. Circulation 2002, 106, 1442–1446. [Google Scholar] [CrossRef] [Green Version]
- Lux, M.; Blaut, A.; Eltrich, N.; Bideak, A.; Müller, M.B.; Hoppe, J.M.; Gröne, H.-J.; Locati, M.; Vielhauer, V. The Atypical Chemokine Receptor 2 Limits Progressive Fibrosis after Acute Ischemic Kidney Injury. Am. J. Pathol. 2019, 189, 231–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bideak, A.; Blaut, A.; Hoppe, J.M.; Müller, M.B.; Federico, G.; Eltrich, N.; Gröne, H.-J.; Locati, M.; Vielhauer, V. The atypical chemokine receptor 2 limits renal inflammation and fibrosis in murine progressive immune complex glomerulonephritis. Kidney Int. 2018, 93, 826–841. [Google Scholar] [CrossRef] [Green Version]
- Urrego-Callejas, T.; Álvarez, S.S.; Arias, L.F.; Reyes, B.O.; Vanegas-García, A.L.; González, L.A.; Muñoz-Vahos, C.H.; Vásquez, G.; Quintana, L.F.; Gómez-Puerta, J.A. Urinary levels of ceruloplasmin and monocyte chemoattractant protein-1 correlate with extra-capillary proliferation and chronic damage in patients with lupus nephritis. Clin. Rheumatol. 2021, 40, 1853–1859. [Google Scholar] [CrossRef]
- de Oliveira Júnior, W.V.; Turani, S.D.; Marinho, M.A.S.; Pinto, S.W.L.; Otoni, A.; Figueiredo, R.C.; Rios, D.R.A. CA-125 and CCL2 may indicate inflammation in peritoneal dialysis patients. J. Bras. Nefrol. 2021. [Google Scholar] [CrossRef]
- Dituri, F.; Cossu, C.; Mancarella, S.; Giannelli, G. The Interactivity between TGFβ and BMP Signaling in Organogenesis, Fibrosis, and Cancer. Cells 2019, 8, 1130. [Google Scholar] [CrossRef] [Green Version]
- Martens, H.; Hennies, I.; Getwan, M.; Christians, A.; Weiss, A.-C.; Brand, F.; Gjerstad, A.C.; Christians, A.; Gucev, Z.; Geffers, R.; et al. Rare heterozygous GDF6 variants in patients with renal anomalies. Eur. J. Hum. Genet. 2020, 28, 1681–1693. [Google Scholar] [CrossRef] [PubMed]
- Dorman, L.J.; Tucci, M.; Benghuzzi, H. In vitro effects of bmp-2, bmp-7, and bmp-13 on proliferation and differentation of mouse mesenchymal stem cells. Biomed. Sci. Instrum. 2012, 48, 81–87. [Google Scholar]
- Skibba, M.; Qian, Y.; Bao, Y.; Lan, J.; Peng, K.; Zhao, Y.; Zhong, P.; Hu, J.; Li, X.; Liang, G. New EGFR inhibitor, 453, prevents renal fibrosis in angiotensin II-stimulated mice. Eur. J. Pharmacol. 2016, 789, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Y.; Han, Y.; Xia, W.; Zhang, L.; Xu, S.; Ju, H.; Zhang, X.; Ren, G.; Liu, L.; et al. EREG-driven oncogenesis of Head and Neck Squamous Cell Carcinoma exhibits higher sensitivity to Erlotinib therapy. Theranostics 2020, 10, 10589–10605. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Hsieh, P.-L.; Chou, C.-L.; Yang, C.-C.; Lee, S.-W.; Tian, Y.-F.; Shiue, Y.-L.; Li, W.-S. High EREG Expression Is Predictive of Better Outcomes in Rectal Cancer Patients Receiving Neoadjuvant Concurrent Chemoradiotherapy. Oncology 2020, 98, 549–557. [Google Scholar] [CrossRef]
- Liu, S.; Ye, D.; Xu, D.; Liao, Y.; Zhang, L.; Liu, L.; Yu, W.; Wang, Y.; He, Y.; Hu, J.; et al. Autocrine epiregulin activates EGFR pathway for lung metastasis via EMT in salivary adenoid cystic carcinoma. Oncotarget 2016, 7, 25251–25263. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Xie, K.-L.; Wu, H.-L.; Wu, K. Functional suppression of Epiregulin impairs angiogenesis and aggravates left ventricular remodeling by disrupting the extracellular-signal-regulated kinase1/2 signaling pathway in rats after acute myocardial infarction. J. Cell. Physiol. 2019, 234, 18653–18665. [Google Scholar] [CrossRef]
- Singh, B.; Carpenter, G.; Coffey, R.J. EGF receptor ligands: Recent advances. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Kochupurakkal, B.S.; Harari, D.; Di-Segni, A.; Maik-Rachline, G.; Lyass, L.; Gur, G.; Kerber, G.; Citri, A.; Lavi, S.; Eilam, R.; et al. Epigen, the last ligand of ErbB receptors, reveals intricate relationships between affinity and mitogenicity. J. Biol. Chem. 2005, 280, 8503–8512. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M. Multi-layered prevention and treatment of chronic inflammation, organ fibrosis and cancer associated with canonical WNT/β-catenin signaling activation (Review). Int. J. Mol. Med. 2018, 42, 713–725. [Google Scholar] [CrossRef]
- Pérez-Lozano, M.L.; Sandoval, P.; Rynne-Vidal, A.; Aguilera, A.; Jiménez-Heffernan, J.A.; Albar-Vizcaíno, P.; Majano, P.L.; Sánchez-Tomero, J.A.; Selgas, R.; López-Cabrera, M. Functional relevance of the switch of VEGF receptors/co-receptors during peritoneal dialysis-induced mesothelial to mesenchymal transition. PLoS ONE 2013, 8, e60776. [Google Scholar] [CrossRef] [Green Version]
- Marquez-Exposito, L.; Lavoz, C.; Rodrigues-Diez, R.R.; Rayego-Mateos, S.; Orejudo, M.; Cantero-Navarro, E.; Ortiz, A.; Egido, J.; Selgas, R.; Mezzano, S.; et al. Gremlin Regulates Tubular Epithelial to Mesenchymal Transition via VEGFR2: Potential Role in Renal Fibrosis. Front. Pharmacol. 2018, 9, 1195. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Ma, X.; Xie, X.; Wu, H.; Wang, L.; Feng, Y.; Yu, Z.; Liu, C.; Qi, J.; Zhu, Q. FN-EDA mediates angiogenesis of hepatic fibrosis via integrin-VEGFR2 in a CD63 synergetic manner. Cell Death Discov. 2020, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, T.; Zhang, H.; Kumanogoh, A.; Takegahara, N.; Yabuki, M.; Harada, K.; Hori, M.; Kikutani, H. Guidance of myocardial patterning in cardiac development by Sema6D reverse signalling. Nat. Cell Biol. 2004, 6, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Nakanishi, Y.; Kioi, Y.; Okuzaki, D.; Kimura, T.; Takamatsu, H.; Koyama, S.; Nojima, S.; Nishide, M.; Hayama, Y.; et al. Semaphorin 6D reverse signaling controls macrophage lipid metabolism and anti-inflammatory polarization. Nat. Immunol. 2018, 19, 561–570. [Google Scholar] [CrossRef]
- Takegahara, N.; Takamatsu, H.; Toyofuku, T.; Tsujimura, T.; Okuno, T.; Yukawa, K.; Mizui, M.; Yamamoto, M.; Prasad, D.V.R.; Suzuki, K.; et al. Plexin-A1 and its interaction with DAP12 in immune responses and bone homeostasis. Nat. Cell Biol. 2006, 8, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, H.; Ishii, K.; Horiuchi, H.; Arai, H.; Kawamoto, T.; Okawa, K.; Iwamatsu, A.; Kita, T. Cutting edge: Combined treatment of TNF-alpha and IFN-gamma causes redistribution of junctional adhesion molecule in human endothelial cells. J. Immunol. 1999, 163, 553–557. [Google Scholar] [PubMed]
- Azari, B.M.; Marmur, J.D.; Salifu, M.O.; Cavusoglu, E.; Ehrlich, Y.H.; Kornecki, E.; Babinska, A. Silencing of the F11R gene reveals a role for F11R/JAM-A in the migration of inflamed vascular smooth muscle cells and in atherosclerosis. Atherosclerosis 2010, 212, 197–205. [Google Scholar] [CrossRef]
- Schenkel, A.R.; Mamdouh, Z.; Muller, W.A. Locomotion of monocytes on endothelium is a critical step during extravasation. Nat. Immunol. 2004, 5, 393–400. [Google Scholar] [CrossRef]
- Saggu, G.; Okubo, K.; Chen, Y.; Vattepu, R.; Tsuboi, N.; Rosetti, F.; Cullere, X.; Washburn, N.; Tahir, S.; Rosado, A.M.; et al. Cis interaction between sialylated FcγRIIA and the αI-domain of Mac-1 limits antibody-mediated neutrophil recruitment. Nat. Commun. 2018, 9, 5058. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, G.; Weber, K.S.C.; Zernecke, A.; Schröder, A.; Weber, C. JAM-1 is a ligand of the beta(2) integrin LFA-1 involved in transendothelial migration of leukocytes. Nat. Immunol. 2002, 3, 151–158. [Google Scholar] [CrossRef]
- Babinska, A.; Clement, C.C.; Przygodzki, T.; Talar, M.; Li, Y.; Braun, M.; Wzorek, J.; Swiatkowska, M.; Ehrlich, Y.H.; Kornecki, E.; et al. A peptide antagonist of F11R/JAM-A reduces plaque formation and prolongs survival in an animal model of atherosclerosis. Atherosclerosis 2019, 284, 92–101. [Google Scholar] [CrossRef]
- Li, J.; Dong, W.; Gao, X.; Chen, W.; Sun, C.; Li, J.; Gao, S.; Zhang, Y.; He, J.; Lu, D.; et al. EphA4 is highly expressed in the atria of heart and its deletion leads to atrial hypertrophy and electrocardiographic abnormalities in rats. Life Sci. 2021, 278, 119595. [Google Scholar] [CrossRef]
- Hao, Y.; Li, G. Role of EFNA1 in tumorigenesis and prospects for cancer therapy. Biomed. Pharmacother. 2020, 130, 110567. [Google Scholar] [CrossRef]
- Cocco, E.; Scaltriti, M.; Drilon, A. NTRK fusion-positive cancers and TRK inhibitor therapy. Nat. Rev. Clin. Oncol. 2018, 15, 731–747. [Google Scholar] [CrossRef] [PubMed]
- Liot, G.; Gabriel, C.; Cacquevel, M.; Ali, C.; MacKenzie, E.T.; Buisson, A.; Vivien, D. Neurotrophin-3-induced PI-3 kinase/Akt signaling rescues cortical neurons from apoptosis. Exp. Neurol. 2004, 187, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Ren, Z.; Yang, X.; Yang, R.; Chen, Y.; Liu, Z.; Dai, Z.; Zhang, Y.; He, Y.; Zhang, C.; et al. Nerve growth factor (NGF)-TrkA axis in head and neck squamous cell carcinoma triggers EMT and confers resistance to the EGFR inhibitor erlotinib. Cancer Lett. 2020, 472, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Lee, J.-J.; Kim, M.S.; Son, B.H.; Cho, Y.K.; Kim, H.-P. DNA methylation-dependent regulation of TrkA, TrkB, and TrkC genes in human hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2011, 406, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; De Vitis, C.; Noto, A.; Fattore, L.; Mariotta, S.; Cherubini, E.; Roscilli, G.; Liguori, G.; Scognamiglio, G.; Rocco, G.; et al. TrkB is responsible for EMT transition in malignant pleural effusions derived cultures from adenocarcinoma of the lung. Cell Cycle 2013, 12, 1696–1703. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, R.; Baker, P.J.; Kent, K.C.; Ye, Y.; Han, J.J.; Tejada, R.; Silane, M.; Upmacis, R.; Deeb, R.; Chen, Y.; et al. Decreased neurotrophin TrkB receptor expression reduces lesion size in the apolipoprotein E-null mutant mouse. Circulation 2005, 112, 3644–3653. [Google Scholar] [CrossRef]
- Kawaguchi-Manabe, H.; Ieda, M.; Kimura, K.; Manabe, T.; Miyatake, S.; Kanazawa, H.; Kawakami, T.; Ogawa, S.; Suematsu, M.; Fukuda, K. A novel cardiac hypertrophic factor, neurotrophin-3, is paradoxically downregulated in cardiac hypertrophy. Life Sci. 2007, 81, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Study Acronym | Platform | Sample Type | Group 1 | Group 2 | References |
|---|---|---|---|---|---|
| Zaza2008 | Affymetrix U133A | patient PBMCs | PD | CKD5 (predialysis) | [6] |
| Reimold2013 | Affymetrix U113 Plus PM | patient mesothelial cells | PD | Uremic | [7] |
| Scherer2013 | Affymetrix U133 Plus 2.0 | patient PBMCs | PD | CKD5 (predialysis) | [8] |
| Büchel2015 | Affymetrix U133 Plus 2.0 | primary mesothelial cells | PD fluids | control medium | [4] |
| Kokoroishi2016 | Affymetrix U133 Plus 2.0 | primary mesothelial cells | high glucose | normal glucose | [9] |
| Herzog2017 | Illumina HiSeq 2000 | patient peritoneal cells | PD without AlaGln | PD with AlaGln | [5] |
| Ruiz-Carpio2017 | Agilent Whole Human Genome Microarrays Kit 4 × 44 K | patient mesothelial cells | epithelioid | non-epithelioid | [10] |
| Bartosova2018 | Illumina Human Sentrix Beads | patient omental arterioles | PD | CKD5 (predialysis) | [11] |
| Han2019 | Illumina HumanRef-8 v2.0 | primary peritoneal cells | TGFB1 stimulated | wt | [12] |
| Liu2019 | Agilent SurePrint G3 Human GE 8 × 60 K Microarray kit | mesothelial cell culture | without HPG | with HPG | [13] |
| Parikova2020 | Illumina Human HT-12 v4 Expression BeadChips | patient peritoneal cells | long-term PD | short-term PD | [14] |
| Strippoli2020 | GRCm38.76 | primary mesothelial cells | stretched | non-stretched | [15] |
| GO Biological Process | All | M | OA | P | PBMC |
|---|---|---|---|---|---|
| angiogenesis | 64 (4.59) | 56 (3.53) | 2 | 8 | - |
| cell adhesion | 110 (4.59) | 88 (1.84) | - | 23 (2.97) | 3 |
| positive regulation of cell migration | 56 (4.59) | 49 (3.53) | - | 9 | - |
| positive regulation of smooth muscle cell proliferation | 28 (4.59) | 23 (3.32) | 3 | 4 | - |
| inflammatory response | 92 (3.99) | 71 (1.33) | - | 14 | 7 (5.66) |
| positive regulation of cell proliferation | 107 (3.84) | 90 (2.2) | 9 | 12 | 1 |
| extracellular matrix organization | 55 (3.46) | 47 (2.39) | - | 12 (1.4) | - |
| positive regulation of angiogenesis | 38 (3.46) | 33 (2.66) | 2 | 3 | 1 |
| cell migration | 49 (3.1) | 39 (1.47) | - | 10 | - |
| leukocyte migration | 38 (2.85) | 31 (1.62) | 4 | 4 | 1 |
| response to estradiol | 30 (2.28) | 25 (1.54) | 1 | 3 | 1 |
| positive regulation of ERK1 and ERK2 cascade | 47 (2.25) | 37 | 2 | 9 | - |
| cell division | 79 (2.2) | 76 (2.9) | 1 | 2 | - |
| cell-cell signaling | 61 (2.04) | 52 | 2 | 8 | 1 |
| response to drug | 70 (2.04) | 60 (1.41) | 4 | 8 | 1 |
| positive regulation of MAP kinase activity | 22 (2.03) | 17 | 1 | 4 | - |
| receptor internalization | 18 (1.96) | 17 (1.83) | - | - | 1 |
| regulation of phosphatidylinositol 3-kinase signaling | 26 (1.93) | 20 | - | 7 | - |
| phosphatidylinositol phosphorylation | 29 (1.76) | 25 | - | 4 | - |
| osteoblast differentiation | 31 (1.76) | 27 (1.46) | 1 | 4 | - |
| regulation of apoptotic process | 52 (1.76) | 46 (1.5) | 1 | 4 | 1 |
| protein autophosphorylation | 44 (1.67) | 39 (1.31) | - | 5 | - |
| cell-cell adhesion | 62 (1.65) | 54 | 6 | 2 | - |
| positive regulation of tyrosine phosphorylation of Stat3 protein | 16 (1.64) | 15 (1.84) | 1 | - | - |
| response to hydrogen peroxide | 19 (1.59) | 16 | - | 3 | - |
| positive regulation of vascular endothelial growth factor production | 13 (1.57) | 11 (1.33) | 2 | - | - |
| response to nutrient | 24 (1.57) | 21 | - | 3 | - |
| negative regulation of apoptotic process | 93 (1.56) | 77 | 9 | 10 | 1 |
| cellular response to retinoic acid | 23 (1.55) | 20 (1.31) | 2 | 1 | - |
| positive regulation of protein kinase B signaling | 26 (1.55) | 23 (1.38) | - | 3 | - |
| wound healing | 25 (1.51) | 19 | 2 | 5 | 1 |
| response to ethanol | 30 (1.47) | 27 | 2 | 2 | 1 |
| extracellular matrix disassembly | 24 (1.46) | 22 | 3 | - | - |
| positive regulation of MAPK cascade | 25 (1.46) | 21 | 1 | 3 | - |
| positive regulation of fibroblast proliferation | 19 (1.38) | 13 | 3 | 4 | - |
| sprouting angiogenesis | 12 (1.36) | 12 (1.51) | - | 2 | - |
| mitotic nuclear division | 56 (1.35) | 54 (1.76) | 1 | 1 | - |
| collagen catabolic process | 21 (1.33) | 19 (1.34) | 1 | 2 | - |
| platelet degranulation | 29 (1.33) | 28 (1.76) | 4 | - | - |
| complement activation, alternative pathway | 7 | 5 | 3 (1.85) | - | - |
| Rho protein signal transduction | 15 | 11 | 4 (1.67) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evgeniou, M.; Sacnun, J.M.; Kratochwill, K.; Perco, P. A Meta-Analysis of Human Transcriptomics Data in the Context of Peritoneal Dialysis Identifies Novel Receptor-Ligand Interactions as Potential Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 13277. https://doi.org/10.3390/ijms222413277
Evgeniou M, Sacnun JM, Kratochwill K, Perco P. A Meta-Analysis of Human Transcriptomics Data in the Context of Peritoneal Dialysis Identifies Novel Receptor-Ligand Interactions as Potential Therapeutic Targets. International Journal of Molecular Sciences. 2021; 22(24):13277. https://doi.org/10.3390/ijms222413277
Chicago/Turabian StyleEvgeniou, Michail, Juan Manuel Sacnun, Klaus Kratochwill, and Paul Perco. 2021. "A Meta-Analysis of Human Transcriptomics Data in the Context of Peritoneal Dialysis Identifies Novel Receptor-Ligand Interactions as Potential Therapeutic Targets" International Journal of Molecular Sciences 22, no. 24: 13277. https://doi.org/10.3390/ijms222413277
APA StyleEvgeniou, M., Sacnun, J. M., Kratochwill, K., & Perco, P. (2021). A Meta-Analysis of Human Transcriptomics Data in the Context of Peritoneal Dialysis Identifies Novel Receptor-Ligand Interactions as Potential Therapeutic Targets. International Journal of Molecular Sciences, 22(24), 13277. https://doi.org/10.3390/ijms222413277