
6-Benzylaminopurine Alleviates the Impact of Cu2+ Toxicity on Photosynthetic Performance of Ricinus communis L. Seedlings

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Effect of Cytokinins on Maintaining Tissue Water Status during Cu2+Toxicity
2.2. Effect of Cytokinins on Cu2+Induced Pigment Loss
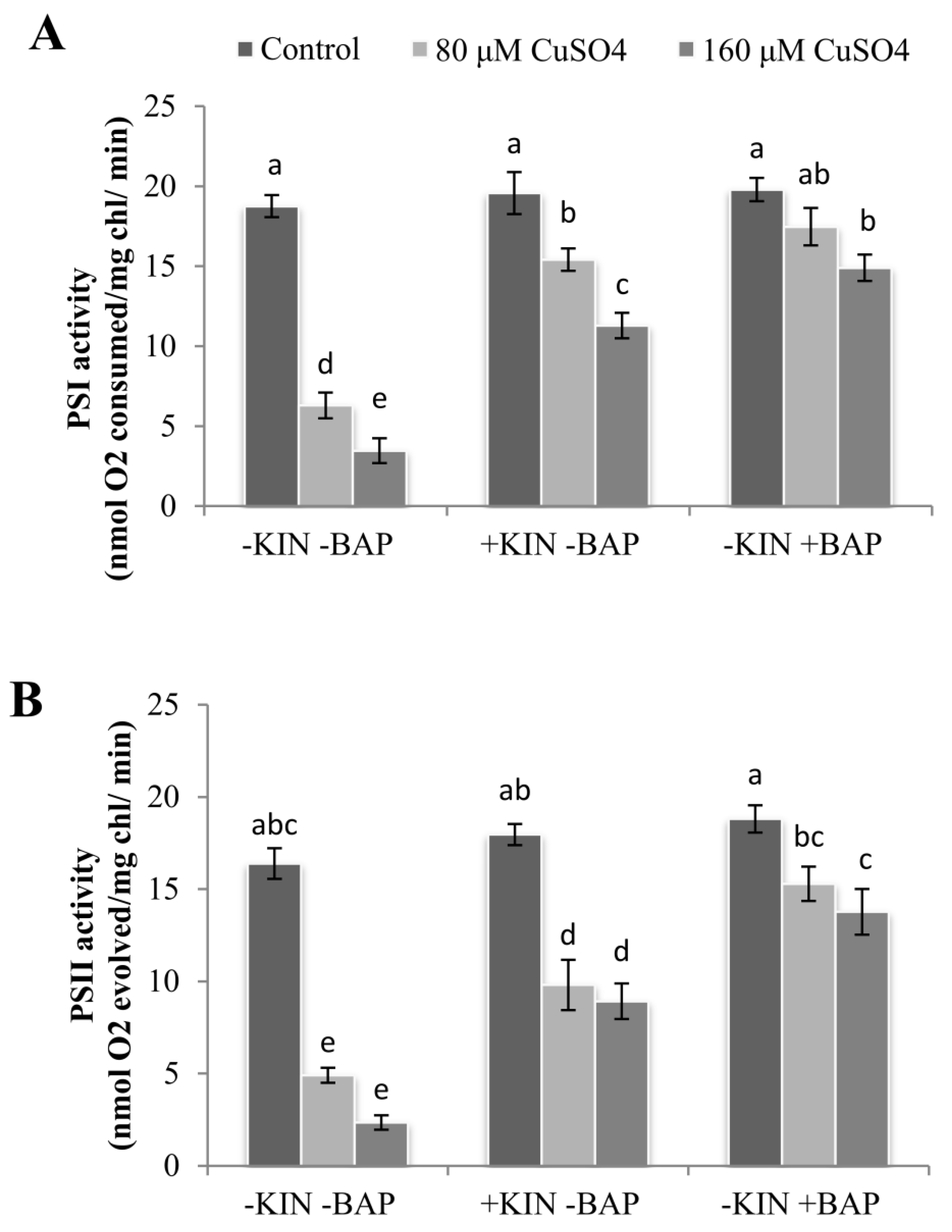
2.3. Effect of Cytokinins on Cu2+-Induced Reduction in Photosystem Activities
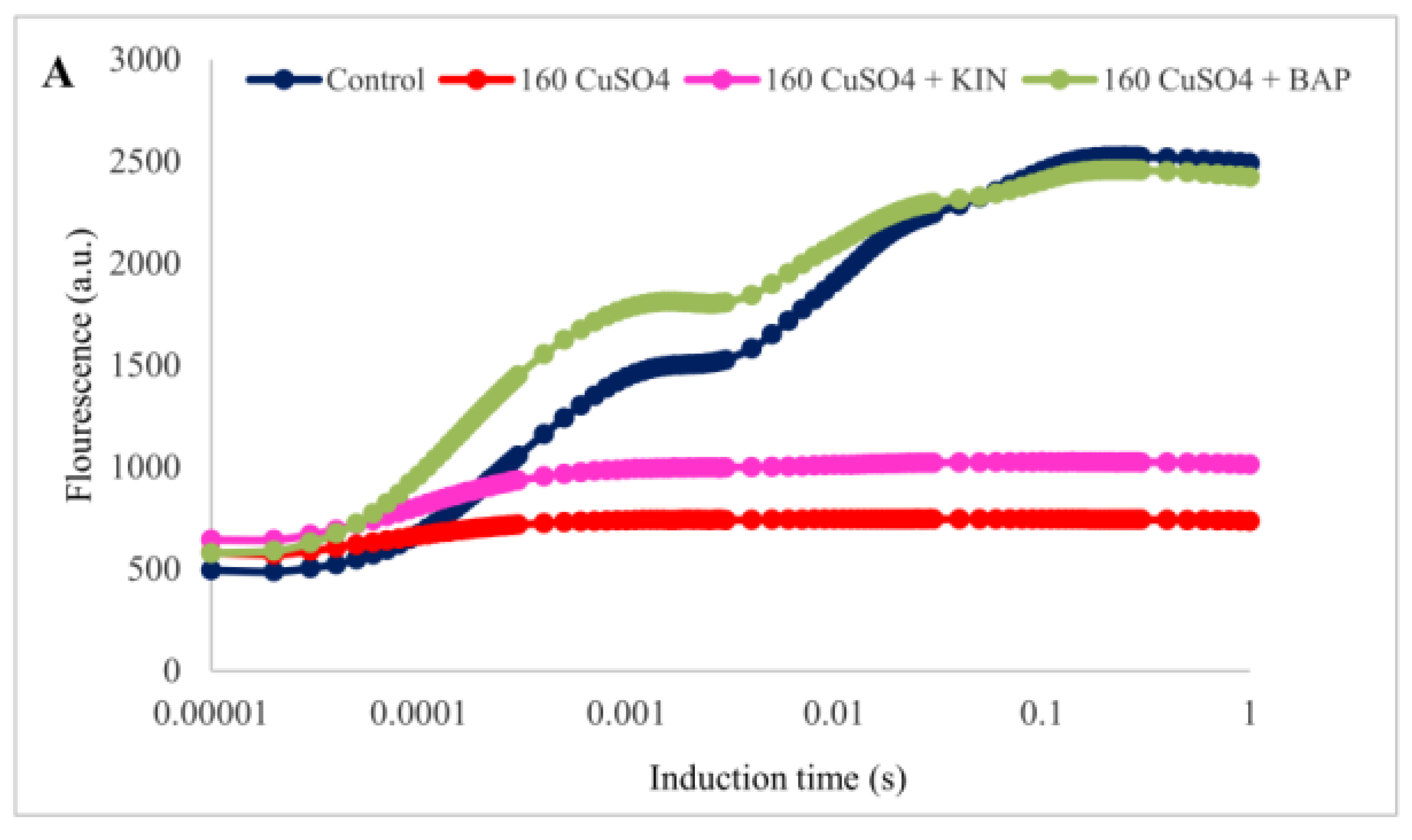
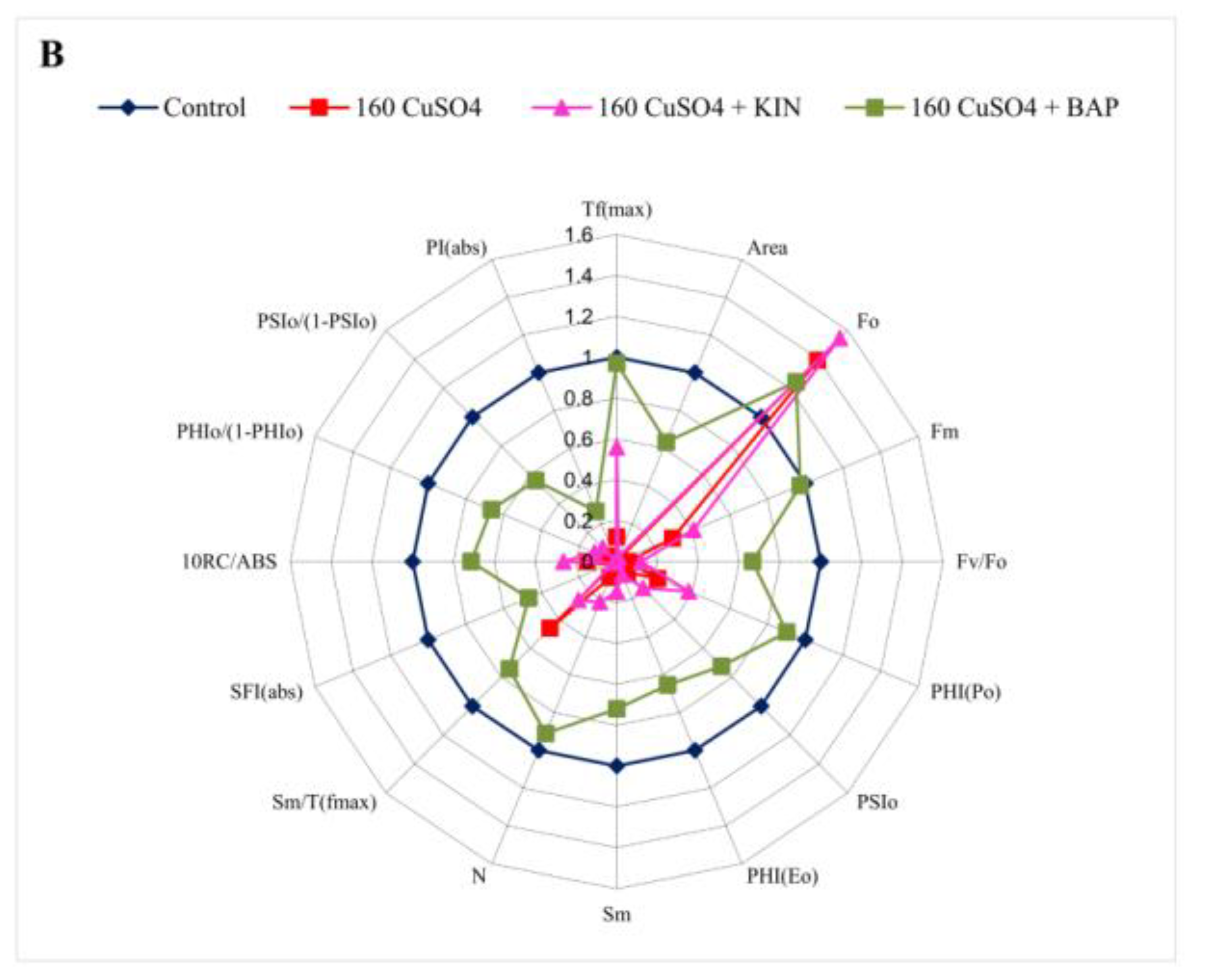
2.4. Modulation of Chlorophyll Fluorescence Parameters by Cytokinin Application
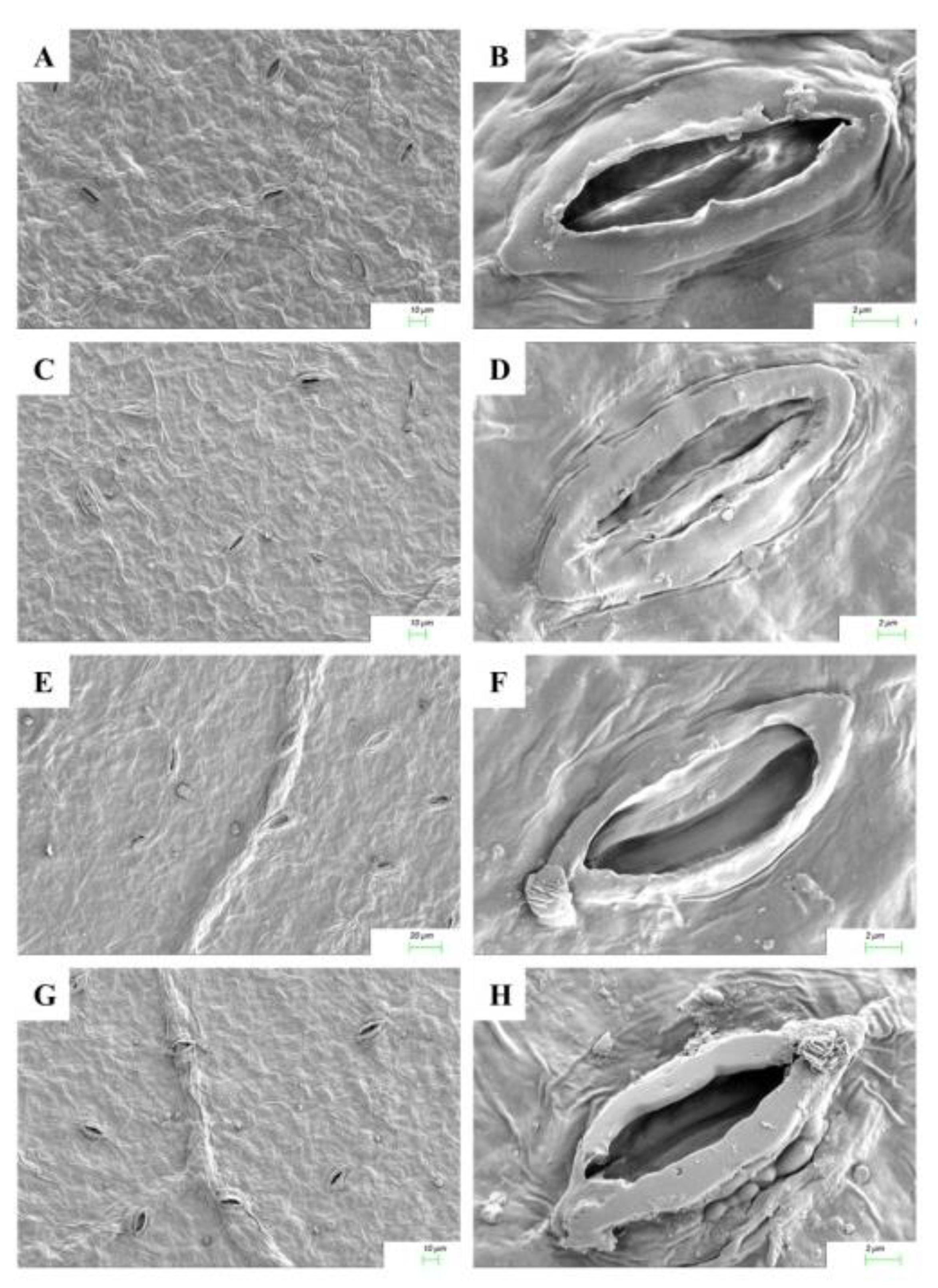
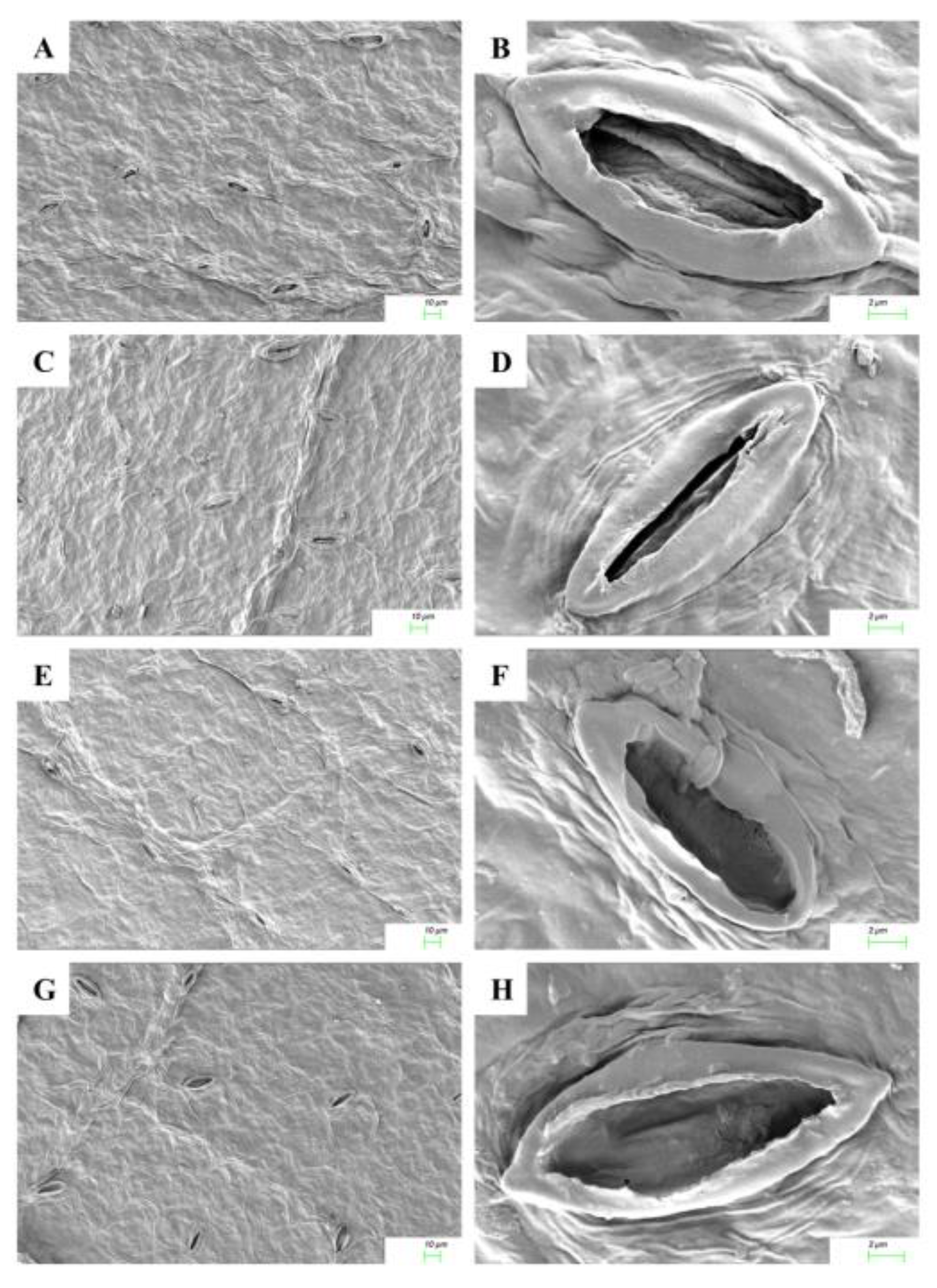
2.5. Cytokinin Mediated Stomatal Responses
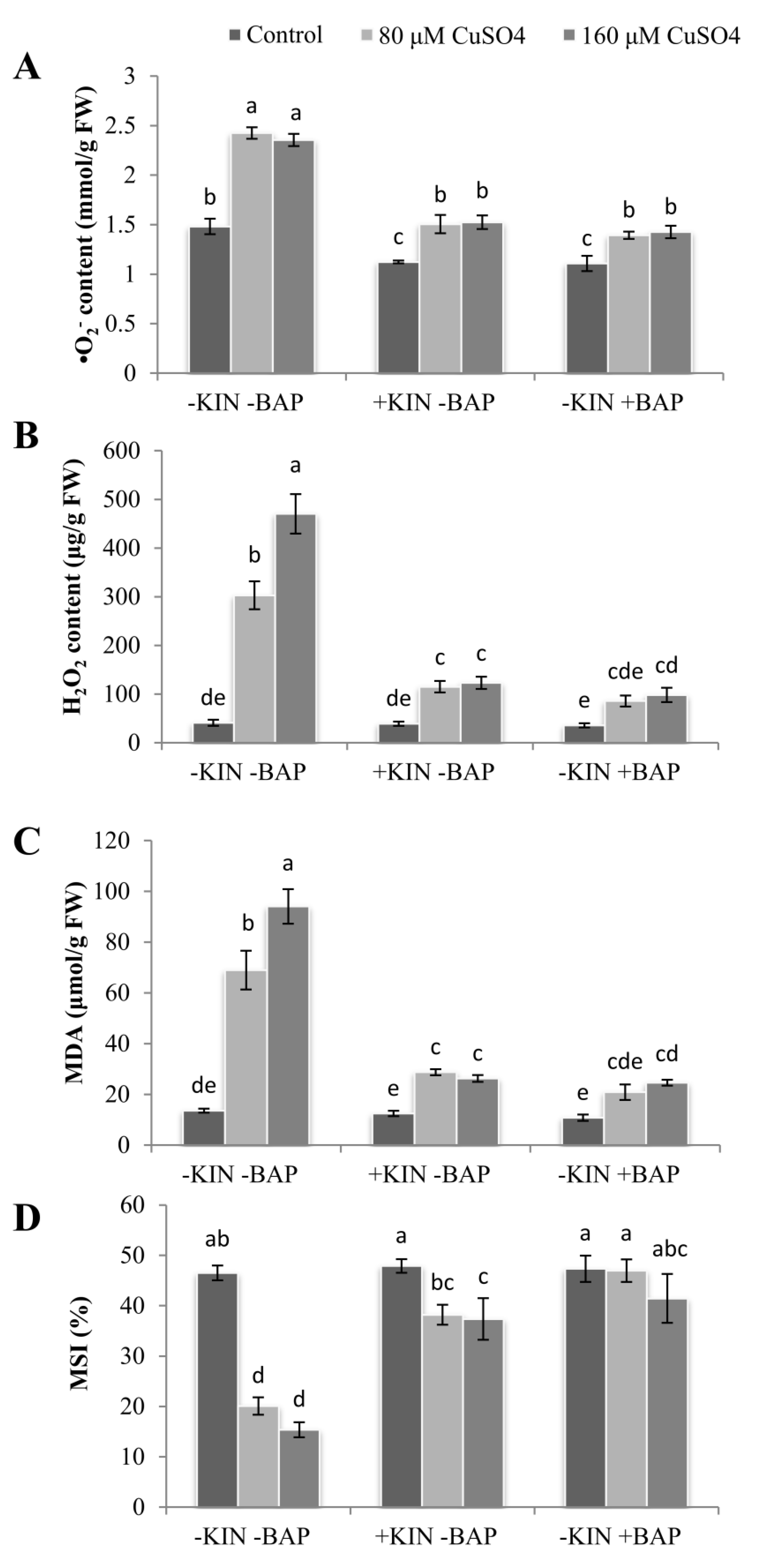
2.6. Cytokinin Induced Decline in ROS and MDA Accumulation
2.7. Maintenance of Membrane Stability Index (MSI) in Response to Cytokinins
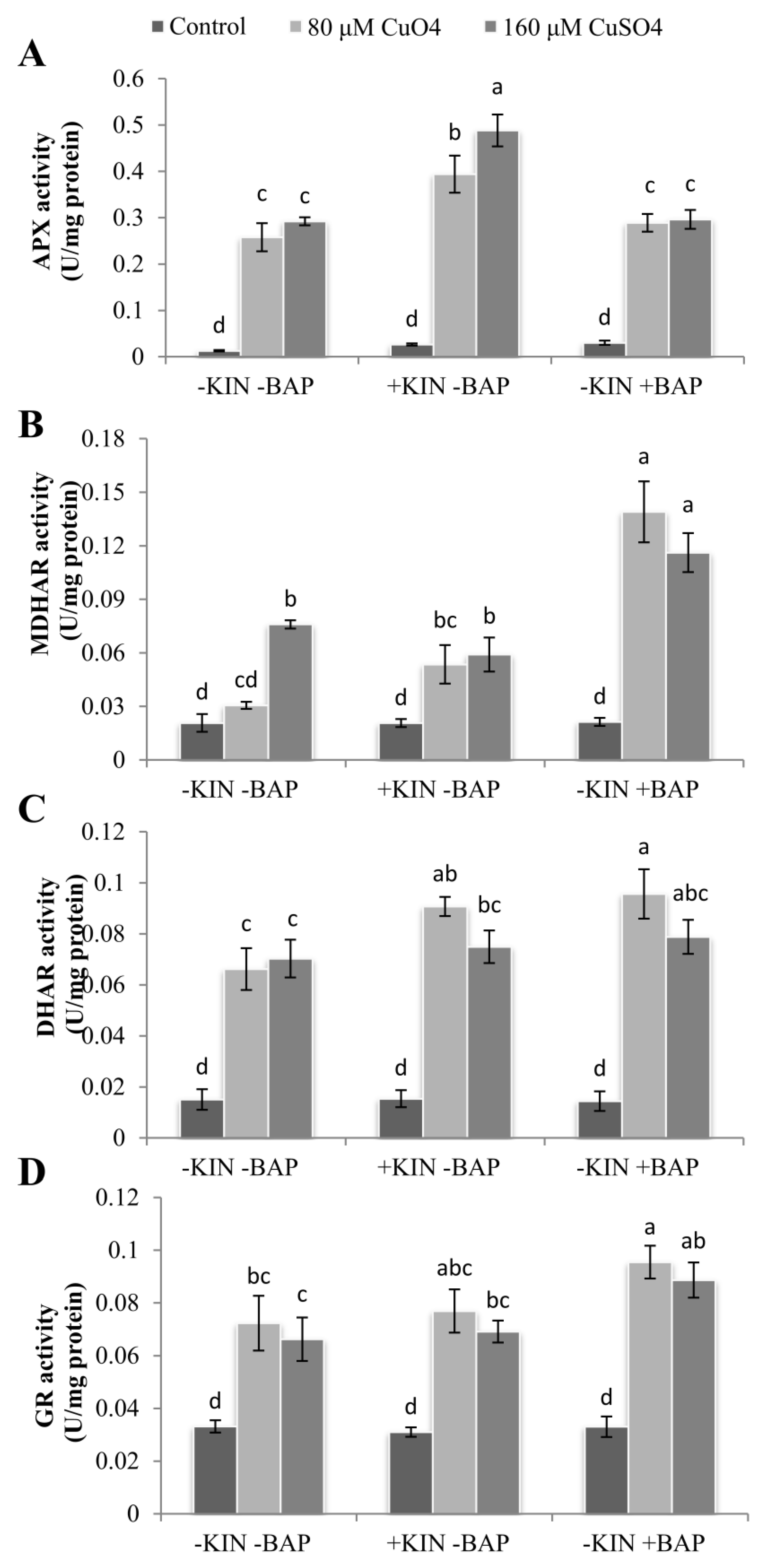
2.8. Cytokinin-Mediated Enhancementsin Antioxidant Enzyme Activities
2.9. Influence of Cytokinins on Accumulation of Non-Enzymatic Antioxidants during Cu Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Treatment with CuSO4 and Cytokinins
4.3. Measurement of Tissue Water Status
4.4. Estimation of Photosynthetic Pigment Composition
4.5. Analysis of Photosystem Activities
4.6. Analysis of Chlorophyll a Fluorescence Parameters
4.7. Analysis of Leaf Micromorphological Characters
4.8. Analysis of ROS Molecules
4.9. Estimation of Malondialdehyde Content
4.10. Estimation of the Membrane Stability Index
4.11. Assay of Enzymatic Antioxidant Activities
4.12. Estimation of Non-Enzymatic Antioxidants
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farooq, M.A.; Ali, S.; Hameed, A.; Bharwana, S.A.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium stress in cotton seedlings: Physiological, photosynthesis and oxidative damages alleviated by glycinebetaine. S. Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Siregar, A.S.; Sulistyo, I.; Prayogo, N.A. Heavy metal contamination in water, sediments and Planiliza subviridis tissue in the Donan River, Indonesia. J. Water Land Dev. 2020, 45, 157–164. [Google Scholar] [CrossRef]
- Kohli, S.k.; Handa, N.; Bali, S.; Arora, S.; Sharma, A.; Kaur, R.; Bhardwaj, R. Modulation of antioxidative defense expression and osmolyte content by co-application of 24-epibrassinolide and salicylic acid in Pb exposed Indian mustard plants. Ecotoxicol. Environ. Saf. 2018, 147, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.; Zaid, A.; Abo-Baker, A.B.A.E.; Salem, W.; Abu Alhmad, M.F. Mitigation of copper stress in maize by inoculation with Paenibacillus polymyxa and Bacillus circulans. Plants 2020, 9, 1513. [Google Scholar] [CrossRef] [PubMed]
- Rather, B.A.; Masood, A.; Sehar, Z.; Majid, A.; Anjum, N.A.; Khan, N.A. Mechanisms and role of nitric oxide in phytotoxicity-mitigation of copper. Front. Plant Sci. 2020, 11, 675. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Natasha Murtaza, G.; Dumat, C.; Shahid, M. Copper uptake, essentiality, toxicity, detoxification and risk assessment in soil-plant environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef] [PubMed]
- Radziemska, M.; Jeznach, J.; Mazur, Z.; Fronczyk, J.; Bilgin, A. Assessment of the effect of reactive materials on the content of selected elements in Indian mustard grown in Cu-contaminated soils. J. Water Land Dev. 2016, 28, 53–60. [Google Scholar] [CrossRef]
- Shu, X.; Yin, L.Y.; Zhang, Q.F.; Wang, W.B. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef]
- Küpper, H.; Šetlík, I.; Spiller, M.; Küpper, F.C.; Prášil, O. Heavy metal-induced inhibition of photosynthesis: Targets of in vivoheavy metal chlorophyll formation. J. Phycol. 2002, 38, 429–441. [Google Scholar] [CrossRef]
- Küpper, H.; Šetlík, I.; Setlikova, E.; Ferimazova, N.; Spiller, M.; Küpper, F.C. Copper-induced inhibition of photosynthesis: Limiting steps of in vivo copper chlorophyll formation in Scmedesmus quadricanda. Funct. Plant Biol. 2003, 30, 1187–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plant 2017, 23, 249–268. [Google Scholar] [CrossRef]
- Janeeshma, E.; Kalaji, H.M.; Puthur, J.T. Differential responses in the photosynthetic efficiency of Oryza sativa and Zea mays on exposure to Cd and Zn toxicity. Acta Physiol. Plant. 2021, 43, 12. [Google Scholar] [CrossRef]
- Bauddh, K.; Singh, K.; Singh, B.; Singh, R.P. Ricinus communis: A Robust plant for bio-energy and phytoremediation of toxic metals from contaminated soil. Ecol. Eng. 2015, 84, 640–652. [Google Scholar] [CrossRef]
- Sameena, P.P.; Puthur, J.T. Heavy metal phytoremediation by bioenergy plants and associated tolerance mechanisms. Soil Sediment Contam. Int. J. 2021, 30, 253–274. [Google Scholar] [CrossRef]
- Sameena, P.P.; Puthur, J.T. Cotyledonary leaves effectively shield the true leaves in Ricinus communis L. from copper toxicity. Int. J. Phytoremediat. 2021, 23, 492–504. [Google Scholar] [CrossRef]
- Thomas, J.C.; Perron, M.; LaRosa, P.C.; Smigocki, A.C. Cytokinin and the regulation of a tobacco metallothionein-like gene during copper stress. Physiol. Plant. 2005, 123, 262–271. [Google Scholar] [CrossRef]
- Aldesuquy, H.; Baka, Z.; Mickky, B. Kinetin and spermine mediated induction of salt tolerance in wheat plants: Leaf area, photosynthesis and chloroplast ultrastructure of flag leaf at ear emergence. Egypt. J. Basic Appl. Sci. 2014, 1, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, H.; Yasmeen, A.; Anjum, M.A.; Hussain, N. Exogenous application of growth enhancers mitigate water stress in wheat by antioxidant elevation. Front. Plant Sci. 2016, 7, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Kumar, S.; Prakash, P. Exogenous application of cytokinin (6-BAP) ameliorates the adverse effect of combined drought and high temperature stress in wheat seedling. J. Pharmacogn. Phytochem. 2018, 7, 1176–1180. [Google Scholar]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Zhang, W.; Peng, K.; Cui, F.; Wang, D.; Zhao, J.; Zhang, Y.; Yu, N.; Wang, Y.; Zeng, D.; Wang, Y.; et al. Cytokinin oxidase/dehydrogenase OsCKX11 coordinates source and sink relationship in rice by simultaneous regulation of leaf senescence and grain number. Plant Biotechnol. J. 2020, 19, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, A.; Srivastava, P.K.; Prasad, S.M. Cadmium toxicity and its amelioration by kinetin in tomato seedlings vis-à-vis ascorbate-glutathione cycle. J. Photochem. Photobiol. B Biol. 2018, 178, 76–84. [Google Scholar] [CrossRef]
- Kamran, M.; Danish, M.; Saleem, M.H.; Malik, Z.; Parveen, A.; Abbasi, G.H.; Jamil, M.; Ali, S.; Afzal, S.; Riaz, M.; et al. Application of abscisic acid and 6-benzylaminopurine modulated morpho-physiological and antioxidative defense responses of tomato (Solanum lycopersicum L.) by minimizing cobalt uptake. Chemosphere 2021, 263, 128169. [Google Scholar] [CrossRef]
- Sameena, P.P.; Puthur, J.T. Exogenous application of cytokinins confers copper stress tolerance in Ricinus communisL. seedlings. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of lead and copper on photosynthetic apparatus in citrus (Citrus aurantium L.) plants. The role of antioxidants in oxidative damage as a response to heavy metal stress. Plants 2021, 10, 155. [Google Scholar] [CrossRef]
- Sharma, A.; Wang, J.; Xu, D.; Tao, S.; Chong, S.; Yan, D.; Li, Z.; Yuan, H.; Zheng, B. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Caryacathayensis plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, Z.; Li, X.; Zha, D. Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters and antioxidative system in seedlings of eggplant (Solanum melongena L.) under salinity stress. Acta Physiol. Plant. 2012, 34, 2105–2114. [Google Scholar] [CrossRef]
- Wu, W.; Du, K.; Kang, X.; Wei, H. The diverse roles of cytokinins in regulating leaf development. Hortic. Res. 2021, 8, 118. [Google Scholar] [CrossRef]
- Sameena, P.P.; Puthur, J.T. Differential modulation of photosynthesis and defense strategies towards copper toxicity in primary and cotyledonary leaves of Ricinus communisL. J. Photochem. Photobiol. 2021, 8, 100059. [Google Scholar] [CrossRef]
- Talla, S.K.; Panigrahy, M.; Kappara, S.; Nirosha, P.; Neelamraju, S.; Ramanan, R. Cytokinin delays dark-induced senescence in rice by maintaining the chlorophyll cycle and photosynthetic complexes. J. Exp. Bot. 2016, 67, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, X.; Li, K.; Wu, M.; Zhang, R.; Zhang, L.; Chen, G. Photosynthetic responses ofOryza sativaL. seedlings to cadmium stress: Physiological, biochemical and ultrastructural analyses. Biometals 2014, 27, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Shackira, A.M.; Puthur, J.T. Cd2+ influences metabolism and elemental distribution in roots of Acanthus ilicifoliusL. Int. J. Phytoremediat. 2019, 21, 866–877. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Ilias, I.; Kabataidis, M.; Chatzimichail, A. Comparative study of nutrient deficiencies on growth and photochemistry of tobacco. J. Plant Nutr. 2003, 26, 1605–1616. [Google Scholar] [CrossRef]
- Gururani, M.A.; Mohanta, T.K.; Bae, H. Current understanding of the interplay between phytohormones and photosynthesis under environmental stress. Int. J. Mol. Sci. 2015, 16, 19055–19085. [Google Scholar] [CrossRef] [Green Version]
- Mathur, S.; Kalaji, H.M.; Jajoo, A. Investigation of deleterious effects of chromium phytotoxicity and photosynthesis in wheat plant. Photosynthetica 2016, 54, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Bashri, G.; Singh, S.; Prasad, S.M.; Ansari, M.J.; Usmani, S.; Alfarraj, S.; Alharbi, S.A.; Brestic, M. Kinetin mitigates Cd-induced damages to growth, photosynthesis and PS II photochemistry of Trigonellaseedlings by up-regulating ascorbate-glutathione cycle. PLoS ONE 2021, 16, e0249230. [Google Scholar] [CrossRef] [PubMed]
- Cortleven, A.; Nitschke, S.; Klaumunzer, M.; AbdElgawad, H.; Asard, H.; Grimm, B.; Riefler, M.; Schmulling, T. A novel protective function for cytokinin in the light stress response is mediated by the ARABIDOPSIS HISTIDINE KINASE2 and ARABIDOPSIS HISTIDINE KINASE3 receptors. Plant Physiol. 2014, 164, 1470–1483. [Google Scholar] [CrossRef] [Green Version]
- Tongra, T.; Mehta, P.; Mathur, S.; Agrawal, D.; Bharti, S.; Los, D.A.; Allakhverdiev, S.I.; Jajoo, A. Computational analysis of fluorescence induction curves in intact spinach leaves treated at different pH. Biosystems 2011, 103, 158–163. [Google Scholar] [CrossRef]
- Tomar, R.S.; Sharma, A.; Jajoo, A. Assessment of phytotoxicity ofanthracene in soybean (Glycine max) with a quick method ofchlorophyll fluorescence. Plant Biol. 2015, 17, 870–876. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [Green Version]
- Sagardoy, R.; Vázquez, S.; Florez-Sarasa, I.D.; Albacete, A.; Ribas-Carbó, M.; Flexas, J.; Abadía, J.; Morales, F. Stomatal and mesophyll conductances to CO2 are the main limitations to photosynthesis in sugar beet (Betavulgaris) plants grown with excess zinc. New Phytol. 2010, 187, 145–158. [Google Scholar] [CrossRef]
- Rucińska-Sobkowiak, R. Water relations in plants subjected to heavy metal stresses. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.Q.; Sesin, V.; Kisiala, A.; Emery, R.N. Phytohormonal roles in plant responses to heavy metal stress: Implications for using macrophytes in phytoremediation of aquatic ecosystems. Environ. Toxicol. Chem. 2021, 40, 7–22. [Google Scholar] [CrossRef]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Exogenous kinetin promotes the nonenzymatic antioxidant system and photosynthetic activity of coffee (Coffea arabica L.) plants under cold stress conditions. Plants 2020, 9, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.G.; Liu, W.; Yu, W.; Zhang, Y.; Ding, S.; Li, H.; Mrak, T.; Kraigher, H.; Bin, L.Z. Abscisic acid enhances lead translocation from the roots to the leaves and alleviates its toxicity in Populuscanescens. J. Hazard. Mater. 2019, 362, 275–285. [Google Scholar] [CrossRef]
- Tóth, S.Z.; Nagy, V.; Puthur, J.T.; Kovács, L.; Garab, G. The physiological role of ascorbate as photosystem II electron donor: Protection against photoinactivation in heat-stressed leaves. Plant Physiol. 2011, 156, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Kiran, B.R.; Prasad, M.N.V. Responses of Ricinus communis L. (castor bean, phytoremediation crop) seedlings to lead (Pb) toxicity in hydroponics. Selcuk J. Agri. Food Sci. 2017, 31, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Weatherley, P. Studies in the water relations of the cotton plant. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls aandbof leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Mirshad, P.P.; Puthur, J.T. Arbuscular mycorrhizal association enhances drought tolerance potential of promising bioenergy grass (Saccharum arundinaceum Retz.). Environ. Monit. Assess. 2016, 188, 425. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Doke, N. Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol. 1983, 23, 345–357. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Analyt. Chem. 2014, 5, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Yin, D.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. Morphological and physiological responses of two Chrysanthemum cultivars differing in their tolerance to water logging. Environ. Exp. Bot. 2009, 67, 87–93. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbatereductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar] [CrossRef]
- Dalton, D.A.; Hanus, F.J.; Russell, S.A.; Evans, H.J. Purification, properties and distribution of ascorbate peroxidase in legume root nodules. Plant Physiol. 1987, 83, 789–794. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [CrossRef]
- Chen, J.X.; Wang, X.F. Guide to Plant Physiological Experiments; South China University of Technology Press: Guangzhou, China, 2002; pp. 123–127. [Google Scholar]
- Mancinelli, A.L.; Yang, C.P.H.; Lindquist, P.; Anderson, O.R.; Rabino, I. Photocontrol of anthocyanin synthesis: III. The action of streptomycin on the synthesis of chlorophyll and anthocyanin. Plant Physiol. 1975, 55, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirecki, R.M.; Teramura, A.H. Effects of ultraviolet-B irradiance on soybean: V. The dependence of plant sensitivity on the photosynthetic photon flux density during and after leaf expansion. Plant Physiol. 1984, 74, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | DW% | MC% | RWC |
|---|---|---|---|
| Control | 10.169 ± 0.508 d | 89.831 ± 4.492 a | 86.179 ± 4.309 b |
| 80 μM CuSO4 | 25.806 ± 1.290 b | 74.194 ± 3.709 b | 37.097 ± 1.855 e |
| 160 μM CuSO4 | 34.783 ± 1.739 a | 65.217 ± 3.261 c | 27.273 ± 1.364 f |
| KIN | 8.511± 0.426 e | 91.489 ± 4.574 a | 91.489 ± 4.574 ab |
| 80 μM CuSO4 + KIN | 8.642 ± 0.432 de | 91.358 ± 4.568 a | 77.083 ± 3.854 c |
| 160 μM CuSO4 + KIN | 16.667 ± 0.833 c | 83.333 ± 4.167 a | 49.180 ± 2.459 d |
| BAP | 8.421 ± 0.421 e | 91.579 ± 4.579 a | 93.548 ± 4.677 a |
| 80 μM CuSO4 + BAP | 9.001 ± 0.450 de | 91.002 ± 4.550 a | 88.349 ± 4.417 ab |
| 160 μM CuSO4 + BAP | 9.211 ± 0.461 de | 90.789 ± 4.539 a | 76.667 ± 3.833 c |
| Treatments | Total Chlorophyll Content (mg/g FW) | Carotenoids Content (mg/g FW) |
|---|---|---|
| Control | 1.115 ± 0.011 cd | 0.508 ± 0.006 c |
| 80 µM CuSO4 | 0.804 ± 0.015 e | 0.248 ± 0.007 f |
| 160 µM CuSO4 | 0.684 ± 0.022 f | 0.213 ± 0.003 g |
| KIN | 1.301 ± 0.056 b | 0.559 ± 0.009 b |
| 80 µM CuSO4 + KIN | 1.064 ± 0.036 d | 0.284 ± 0.003 e |
| 160 µM CuSO4 + KIN | 1.075 ± 0.004 d | 0.385 ± 0.029 d |
| BAP | 1.477 ± 0.064 a | 0.503 ± 0.009 c |
| 80 µM CuSO4 + BAP | 1.433 ± 0.021 a | 0.622 ± 0.011 a |
| 160 µM CuSO4 + BAP | 1.209 ± 0.013 bc | 0.417 ± 0.002 d |
| Treatments | ABS/CSm | TRo/CSm | ETo/CSm | DIo/CSm | ABS/RC | TRo/RC | ETo/RC | DIo/RC |
|---|---|---|---|---|---|---|---|---|
| Control | 2525.333 ± 36.448 ab | 2102.333 ± 29.237 a | 1020.667 ± 23.355 a | 423.0 ± 14.572 d | 2.338 ± 0.082 b | 1.946 ± 0.063 d | 0.944 ± 0.014 a | 0.392 ± 0.021 b |
| 80 μM CuSO4 | 1869.667 ± 233.326 c | 1251.333 ± 273.866 c | 334.333 ± 121.436 c | 618.333 ± 41.882 ab | 4.286 ± 0.569 b | 2.707 ± 0.069 a | 0.658 ± 0.140 b | 1.579 ± 0.552 b |
| 160 μM CuSO4 | 748.0 ± 160.706 d | 158.667 ± 71.966 d | 5.0 ± 4.0 d | 589.333 ± 91.758 abc | 19.089 ± 6.566 a | 2.872 ± 0.097 a | 0.070 ± 0.029 c | 16.217 ± 6.511 a |
| KIN | 2633.667 ± 52.616 a | 2147.667 ± 44.946 a | 973.333 ± 26.187 a | 486.0 ± 10.599 bcd | 2.586 ± 0.068 b | 2.109 ± 0.051 cd | 0.955 ± 0.017 a | 0.477 ± 0.018 b |
| 80 μM CuSO4 + KIN | 2194.667 ± 76.667 bc | 1691.333 ± 70.333 b | 600.0 ± 18.0 b | 503.333 ± 6.333 bcd | 2.880 ± 0.033 b | 2.219 ± 0.011 c | 0.788 ± 0.013 ab | 0.662 ± 0.022 b |
| 160 μM CuSO4 + KIN | 1027.0 ± 129.418 d | 373.0 ± 115.754 d | 29.0 ± 14.422 d | 654.0 ± 67.023 a | 8.438 ± 1.556 b | 2.702 ± 0.084 a | 0.186 ± 0.041 c | 5.736 ± 1.579 b |
| BAP | 2673.667 ± 13.220 a | 2184.667 ± 15.857 a | 995.667 ± 61.974 a | 489.0 ± 7.638 bcd | 2.482 ± 0.036 b | 2.028 ± 0.026 cd | 0.922 ± 0.041 a | 0.454 ± 0.013 b |
| 80 μM CuSO4 + BAP | 2679.333 ± 46.998 a | 2211.333 ± 28.386 a | 1027.333 ± 12.333 a | 468.0 ± 18.717 cd | 2.515 ± 0.054 b | 2.076 ± 0.037 cd | 0.964 ± 0.014 a | 0.439 ± 0.019 b |
| 160 μM CuSO4 + BAP | 2458.0 ± 75.020 ab | 1931.667 ± 86.822 ab | 649.333 ± 63.679 b | 526.333 ± 13.569 abcd | 3.065 ± 0.122 b | 2.404 ± 0.062 b | 0.803 ± 0.031 ab | 0.661 ± 0.059 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sameena, P.P.; Kalaji, H.M.; Żuk-Gołaszewska, K.; Horaczek, T.; Sierka, E.; Puthur, J.T. 6-Benzylaminopurine Alleviates the Impact of Cu2+ Toxicity on Photosynthetic Performance of Ricinus communis L. Seedlings. Int. J. Mol. Sci. 2021, 22, 13349. https://doi.org/10.3390/ijms222413349
Sameena PP, Kalaji HM, Żuk-Gołaszewska K, Horaczek T, Sierka E, Puthur JT. 6-Benzylaminopurine Alleviates the Impact of Cu2+ Toxicity on Photosynthetic Performance of Ricinus communis L. Seedlings. International Journal of Molecular Sciences. 2021; 22(24):13349. https://doi.org/10.3390/ijms222413349
Chicago/Turabian StyleSameena, Puthukkolli P., Hazem M. Kalaji, Krystyna Żuk-Gołaszewska, Tomasz Horaczek, Edyta Sierka, and Jos T. Puthur. 2021. "6-Benzylaminopurine Alleviates the Impact of Cu2+ Toxicity on Photosynthetic Performance of Ricinus communis L. Seedlings" International Journal of Molecular Sciences 22, no. 24: 13349. https://doi.org/10.3390/ijms222413349
APA StyleSameena, P. P., Kalaji, H. M., Żuk-Gołaszewska, K., Horaczek, T., Sierka, E., & Puthur, J. T. (2021). 6-Benzylaminopurine Alleviates the Impact of Cu2+ Toxicity on Photosynthetic Performance of Ricinus communis L. Seedlings. International Journal of Molecular Sciences, 22(24), 13349. https://doi.org/10.3390/ijms222413349