
Availability of mRNA Obtained from Peripheral Blood Mononuclear Cells for Testing Mutation Consequences in Dystrophic Epidermolysis Bullosa


Abstract
:1. Introduction
2. Results
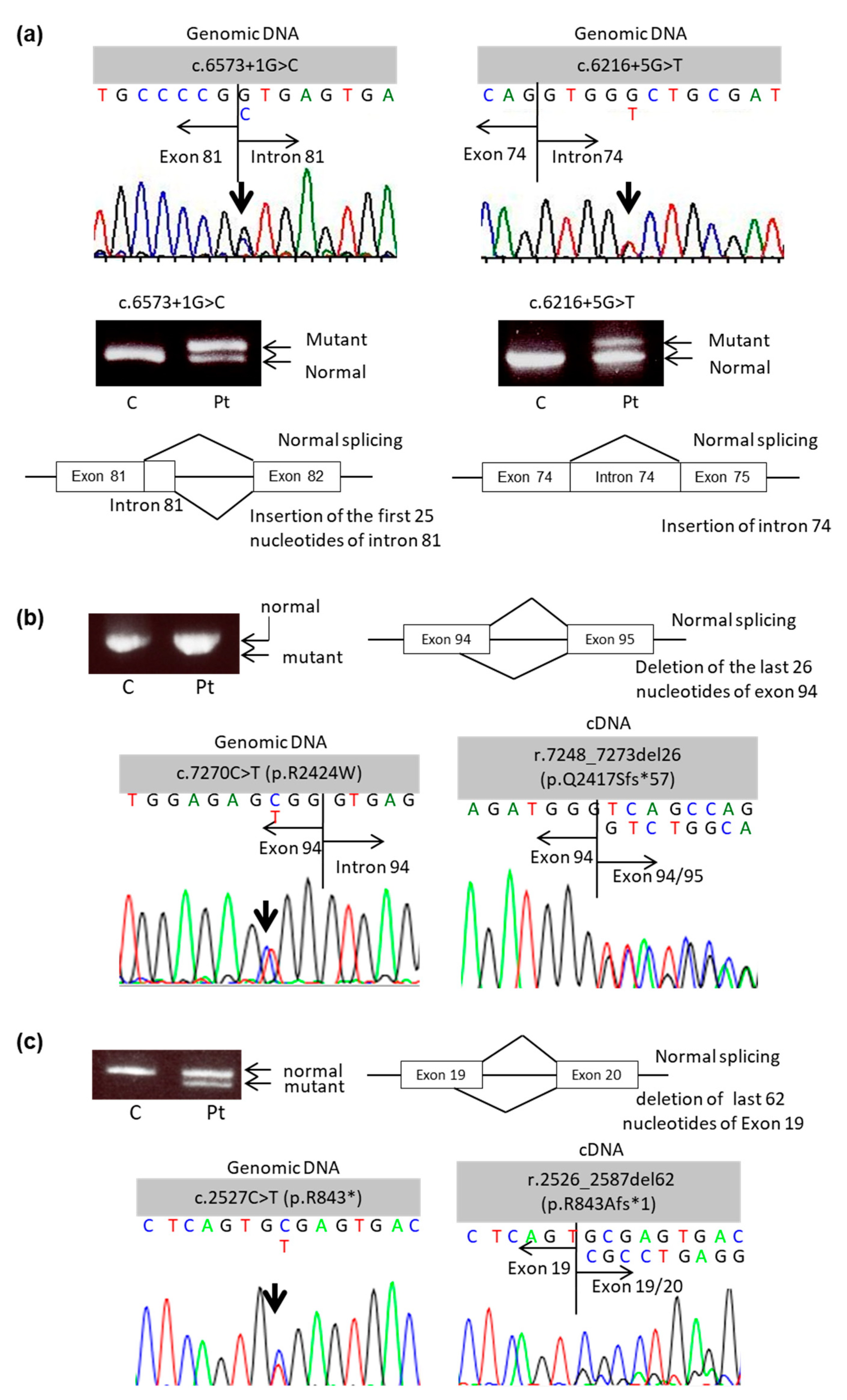
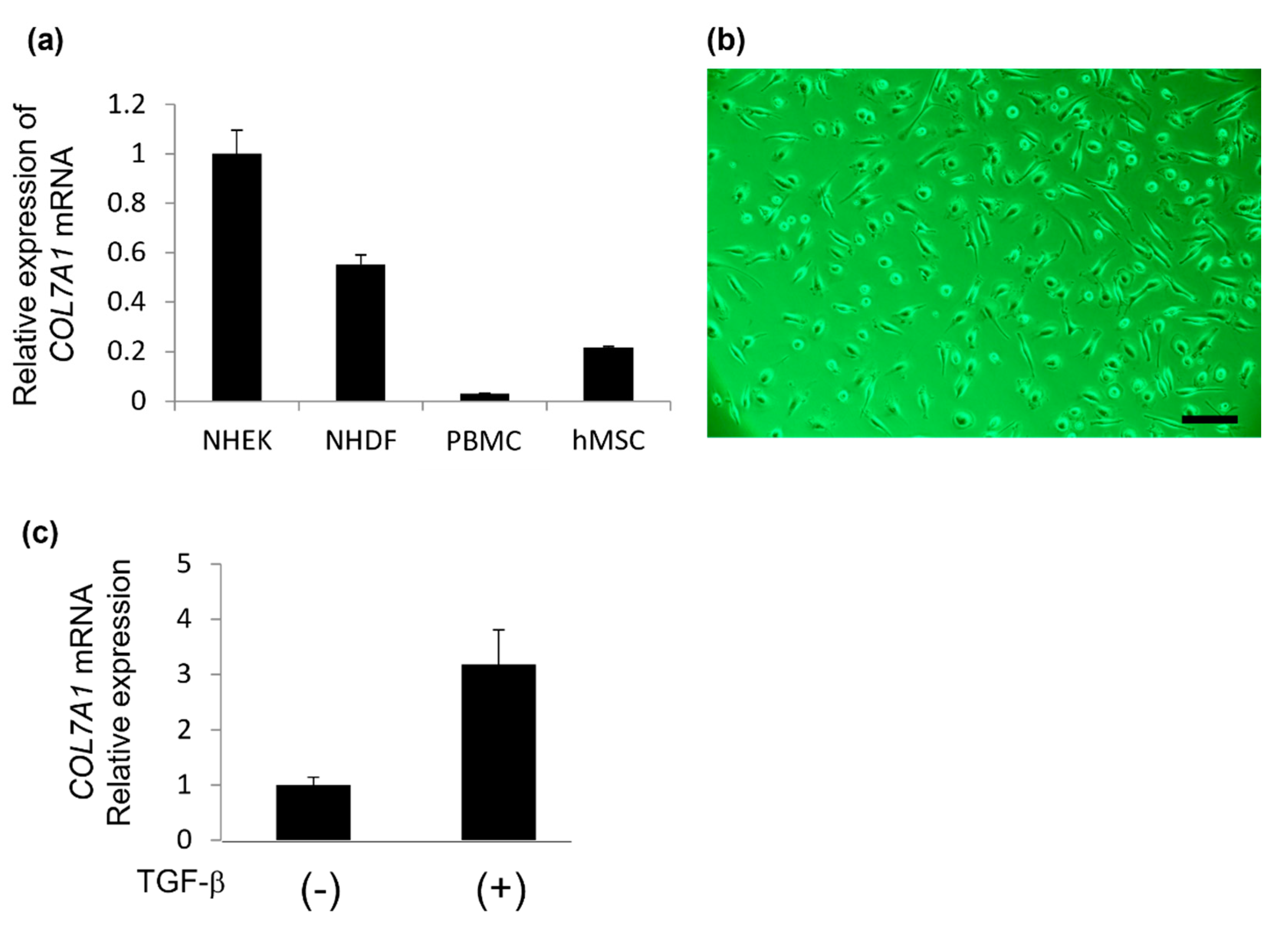
2.1. Mutational Analysis of COL7A1 Using mRNA Obtained from PBMCs
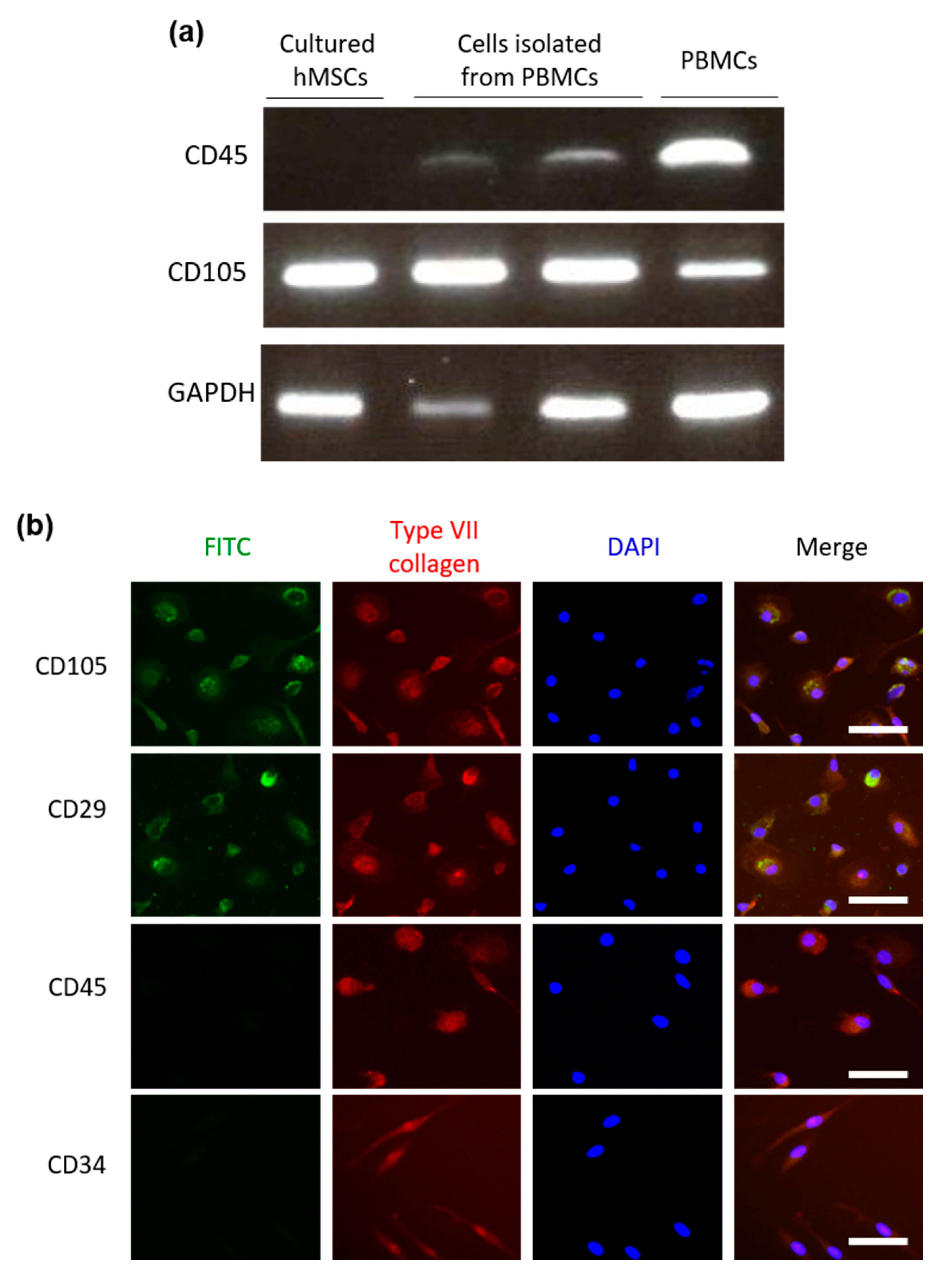
2.2. Type VII Collagen Expression in Isolated Cells Obtained from PBMCs Using the Human Mesenchymal Stem Cell Enrichment Cocktail
2.3. Cells Isolated from PBMCs Using the Human Mesenchymal Stem Cell Enrichment Cocktail Have a Similar Expression Pattern of Cell Surface Markers as Human MSCs
3. Discussion
4. Materials and Methods
4.1. Mutational Analysis
4.2. Cell Culture
4.3. Isolation of Mesenchymal Stromal Cells from PBMCs
4.4. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
4.5. Immunofluorescence of Peripheral Blood-Derived Mesenchymal Stromal Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Has, C.; Bauer, J.W.; Bodemer, C.; Bolling, M.; Bruckner-Tuderman, L.; Diem, A.; Fine, J.-D.; Heagerty, A.; Hovnanian, A.; Marinkovich, P.; et al. Consensus re-classification of inherited epidermolysis bullosa and other disorders with skin fragility. Br. J. Dermatol. 2020, 183, 614–627. [Google Scholar] [CrossRef] [Green Version]
- Varki, R.; Sadowski, S.; Uitto, J.; Pfendner, E. Epidermolysis bullosa. II. Type VII collagen mutations and phenotype–genotype correlations in the dystrophic subtypes. J. Med. Genet. 2007, 44, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Kern, J.S.; Grüninger, G.; Imsak, R.; Müller, M.L.; Schumann, H.; Kiritsi, D.; Emmert, S.; Borozdin, W.; Kohlhase, J.; Bruckner-Tuderman, L.; et al. Forty-two novel COL7A1 mutations and the role of a frequent single nucleotide polymorphism in the MMP1 promoter in modulation of disease severity in a large European dystrophic epidermolysis bullosa cohort. Br. J. Dermatol. 2009, 161, 1089–1097. [Google Scholar] [CrossRef]
- Vahidnezhad, H.; Youssefian, L.; Zeinali, S.; Saeidian, A.H.; Sotoudeh, S.; Mozafari, N.; Abiri, M.; Kajbafzadeh, A.-M.; Barzegar, M.; Ertel, A.; et al. Dystrophic Epidermolysis Bullosa: COL7A1 Mutation Landscape in a Multi-Ethnic Cohort of 152 Extended Families with High Degree of Customary Consanguineous Marriages. J. Investig. Dermatol. 2017, 137, 660–669. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, Z.; Mi, Z.; Sun, L.; Fu, X.; Yu, G.; Pang, Z.; Liu, H.; Zhang, F. Epidermolysis Bullosa in Chinese Patients: Genetic Analysis and Mutation Landscape in 57 Pedigrees and Sporadic Cases. Acta Dermatol. Venereol. 2021, 101, adv00503. [Google Scholar] [CrossRef]
- Nilay, M.; Saxena, D.; Mandal, K.; Moirangthem, A.; Phadke, S.R. Novel pathogenic variants in an Indian cohort with epidermolysis bullosa: Expanding the genotypic spectrum. Eur. J. Med. Genet. 2021, 64, 104345. [Google Scholar] [CrossRef]
- Jiang, W.; Bu, D.; Yang, Y.; Zhu, X. A novel splice site mutation in collagen type VII gene in a Chinese family with dominant dystrophic epidermolysis bullosa pruriginosa. Acta Dermatol. Venereol. 2002, 82, 187–191. [Google Scholar] [CrossRef]
- Jiang, W.; Sun, T.; Lei, P.; Zhu, X. Genotype-phenotype correlation in Chinese patients with dystrophic epidermolysis bullosa pruriginosa. Acta Dermatol. Venereol. 2012, 92, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Tamai, K.; Ishida-Yamamoto, A.; Matsuo, S.; Iizuka, H.; Hashimoto, I.; Christiano, A.M.; Uitto, J.; McGrath, J.A. Compound heterozygosity for a nonsense mutation and a splice site mutation in the type VII collagen gene (COL7A1) in recessive dystrophic epidermolysis bullosa. Lab. Investig. 1997, 76, 209–217. [Google Scholar]
- Kon, A.; Pulkkinen, L.; Ishida-Yamamoto, A.; Hashimoto, I.; Uitto, J. Novel COL7A1 mutations in dystrophic forms of epidermolysis bullosa. J. Investig. Dermatol. 1998, 111, 534–537. [Google Scholar] [CrossRef]
- Hamada, T.; Fukuda, S.; Ishii, N.; Sakaguchi, S.; Ishikawa, T.; Abe, T.; Yasumoto, S.; Hashimoto, T.; Nakano, H.; Sawamura, D. Genotype-phenotype correlation in non-Hallopeau-Siemens recessive dystrophic epidermolysis bullosa: The splice site mutation c.6216+5G > T in the COL7A1 gene results in aberrant and normal splicings. J. Dermatol. Sci. 2008, 50, 147–150. [Google Scholar] [CrossRef]
- Masunaga, T.; Saito, M.; Sasaki, T.; Kubo, A.; Amagai, M.; Ishiko, A. Japanese recurrent mutation c.6216+5G>T in COL7A1 leads to a mild phenotype of dystrophic epidermolysis bullosa. J. Dermatol. Sci. 2015, 80, 220–223. [Google Scholar] [CrossRef]
- Kern, J.S.; Kohlhase, J.; Bruckner-Tuderman, L.; Has, C. Expanding the COL7A1 mutation database: Novel and recurrent mutations and unusual genotype-phenotype constellations in 41 patients with dystrophic epidermolysis bullosa. J. Investig. Dermatol. 2006, 126, 1006–1012. [Google Scholar] [CrossRef] [Green Version]
- Chino, T.; Tamai, K.; Yamazaki, T.; Otsuru, S.; Kikuchi, Y.; Nimura, K.; Endo, M.; Nagai, M.; Uitto, J.; Kitajima, Y.; et al. Bone marrow cell transfer into fetal circulation can ameliorate genetic skin diseases by providing fibroblasts to the skin and inducing immune tolerance. Am. J. Pathol. 2008, 173, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Iinuma, S.; Aikawa, E.; Tamai, K.; Fujita, R.; Kikuchi, Y.; Chino, T.; Kikuta, J.; McGrath, J.; Uitto, J.; Ishii, M.; et al. Transplanted bone marrow-derived circulating PDGFRα+ cells restore type VII collagen in recessive dystrophic epidermolysis bullosa mouse skin graft. J. Immunol. 2015, 194, 1996–2003. [Google Scholar] [CrossRef] [Green Version]
- El-Darouti, M.; Fawzy, M.; Amin, I.; Abdel Hay, R.; Hegazy, R.; Gabr, H.; El Maadawi, Z. Treatment of dystrophic epidermolysis bullosa with bone marrow non-hematopoeitic stem cells: A randomized controlled trial. Dermatol. Ther. 2016, 29, 96–100. [Google Scholar] [CrossRef]
- Wu, G.; Pan, M.; Wang, X.; Wen, J.; Cao, S.; Li, Z.; Li, Y.; Qian, C.; Liu, Z.; Wu, W.; et al. Osteogenesis of peripheral blood mesenchymal stem cells in self assembling peptide nanofiber for healing critical size calvarial bony defect. Sci. Rep. 2015, 5, 16681. [Google Scholar] [CrossRef] [Green Version]
- Petrof, G.; Lwin, S.M.; Martinez-Queipo, M.; Abdul-Wahab, A.; Tso, S.; Mellerio, J.E.; Slaper-Cortenbach, I.; Boelens, J.J.; Tolar, J.; Veys, P.; et al. Potential of Systemic Allogeneic Mesenchymal Stromal Cell Therapy for Children with Recessive Dystrophic Epidermolysis Bullosa. J. Investig. Dermatol. 2015, 135, 2319–2321. [Google Scholar] [CrossRef] [Green Version]
- Ganier, C.; Titeux, M.; Gaucher, S.; Peltzer, J.; Le Lorc’h, M.; Lataillade, J.-J.; Ishida-Yamamoto, A.; Hovnanian, A. Intradermal Injection of Bone Marrow Mesenchymal Stromal Cells Corrects Recessive Dystrophic Epidermolysis Bullosa in a Xenograft Model. J. Investig. Dermatol. 2018, 138, 2483–2486. [Google Scholar] [CrossRef] [Green Version]
- Rashidghamat, E.; Kadiyirire, T.; Ayis, S.; Petrof, G.; Liu, L.; Pullabhatla, V.; Ainali, C.; Guy, A.; Aristodemou, S.; McMillan, J.R.; et al. Phase I/II open-label trial of intravenous allogeneic mesenchymal stromal cell therapy in adults with recessive dystrophic epidermolysis bullosa. J. Am. Acad. Dermatol. 2020, 83, 447–454. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, S.-J.; Kim, S.-E.; Kim, K.; Cho, B.; Roh, K.; Kim, S.-C. Intravenous allogeneic umbilical cord blood-derived mesenchymal stem cell therapy in recessive dystrophic epidermolysis bullosa patients. JCI Insight 2021, 6, 143606. [Google Scholar] [CrossRef]
- Tamai, K.; Yamazaki, T.; Chino, T.; Ishii, M.; Otsuru, S.; Kikuchi, Y.; Iinuma, S.; Saga, K.; Nimura, K.; Shimbo, T.; et al. PDGFRalpha-positive cells in bone marrow are mobilized by high mobility group box 1 (HMGB1) to regenerate injured epithelia. Proc. Natl. Acad. Sci. USA 2011, 108, 6609–6614. [Google Scholar] [CrossRef] [Green Version]
- Abe, S.; Sato, S.; Aono, Y.; Azuma, M.; Kishi, M.; Koyama, K.; Takahashi, N.; Kagawa, K.; Kawano, H.; Nishioka, Y. Functional analysis of human fibrocytes derived from monocytes reveals their profibrotic phenotype through paracrine effects. J. Med. Investig. 2020, 67, 102–112. [Google Scholar] [CrossRef]
- Christiano, A.M.; Hoffman, G.G.; Zhang, X.; Xu, Y.; Tamai, Y.; Greenspan, D.S.; Uitto, J. Strategy for identification of sequence variants in COL7A1 and a novel 2-bp deletion mutation in recessive dystrophic epidermolysis bullosa. Hum. Mutat. 1997, 10, 408–414. [Google Scholar] [CrossRef]
- Valenti, M.T.; Dalle Carbonare, L.; Donatelli, L.; Bertoldo, F.; Zanatta, M.; Lo Cascio, V. Gene expression analysis in osteoblastic differentiation from peripheral blood mesenchymal stem cells. Bone 2008, 43, 1084–1092. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient No. | Age (years) | Sex | Clinical Subtype of DEB | Genotype (Genomic DNA) | Expected Consequence | Genotype (cDNA) | Expected Consequence |
|---|---|---|---|---|---|---|---|
| 1 | 27 | F | Intermediate RDEB | c.6573+1G > C | Splice site mutation | Insertion of 25 nucle- otides of intron 81 | |
| c.6216+5G > T | Splice site mutation | Insertion of intron 74 | |||||
| 2 | 40 | M | Intermediate RDEB | c.409C > T (p.R137*) | Nonsense mutation | c.409C > T (p.R137*) | Nonsense mutation |
| c.7270C > T (p.R2424W) | Splice site mutation | r.7248_7273del26 (p.Q2417Sfs*57) | Splice site mutation leading to PTC | ||||
| 3 | 19 | M | Severe RDEB | c.8053C > T (p.R2685*) | Nonsense mutation | c.8053C > T (p.R2685*) | Nonsense mutation |
| c.2527C > T (p.R843*) | Nonsense mutation | r.2526_2587del62 (p.R843Afs*1) | Splice site mutation leading to PTC |
| Antibody | Host Animal | Dilution | Manufacturer (Location) |
|---|---|---|---|
| Anti-collagen type VII (LH7.2) | Mouse | 1:50 | Sigma-Aldrich (St. Louis, MO, USA) |
| Anti-CD105 | Rabbit | 1:200 | Abcam (Cambridge, UK) |
| Anti-CD29 | Rabbit | 1:50 | Abcam (Cambridge, UK) |
| Anti-CD45 | Rabbit | 1:200 | Abcam (Cambridge, UK) |
| Anti-CD34 | Rabbit | 1:50 | Abcam (Cambridge, UK) |
| Anti-mouse IgG - Alexa Fluor 647 | Donkey | 1:200 | Abcam (Cambridge, UK) |
| Anti-rabbit IgG - FITC | Goat | 1:200 | Sigma-Aldrich (St. Louis, MO, USA) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akasaka, E.; Nakano, H.; Sawamura, D. Availability of mRNA Obtained from Peripheral Blood Mononuclear Cells for Testing Mutation Consequences in Dystrophic Epidermolysis Bullosa. Int. J. Mol. Sci. 2021, 22, 13369. https://doi.org/10.3390/ijms222413369
Akasaka E, Nakano H, Sawamura D. Availability of mRNA Obtained from Peripheral Blood Mononuclear Cells for Testing Mutation Consequences in Dystrophic Epidermolysis Bullosa. International Journal of Molecular Sciences. 2021; 22(24):13369. https://doi.org/10.3390/ijms222413369
Chicago/Turabian StyleAkasaka, Eijiro, Hajime Nakano, and Daisuke Sawamura. 2021. "Availability of mRNA Obtained from Peripheral Blood Mononuclear Cells for Testing Mutation Consequences in Dystrophic Epidermolysis Bullosa" International Journal of Molecular Sciences 22, no. 24: 13369. https://doi.org/10.3390/ijms222413369
APA StyleAkasaka, E., Nakano, H., & Sawamura, D. (2021). Availability of mRNA Obtained from Peripheral Blood Mononuclear Cells for Testing Mutation Consequences in Dystrophic Epidermolysis Bullosa. International Journal of Molecular Sciences, 22(24), 13369. https://doi.org/10.3390/ijms222413369