
Metabolomic Profiling of Blood-Derived Microvesicles in Breast Cancer Patients

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
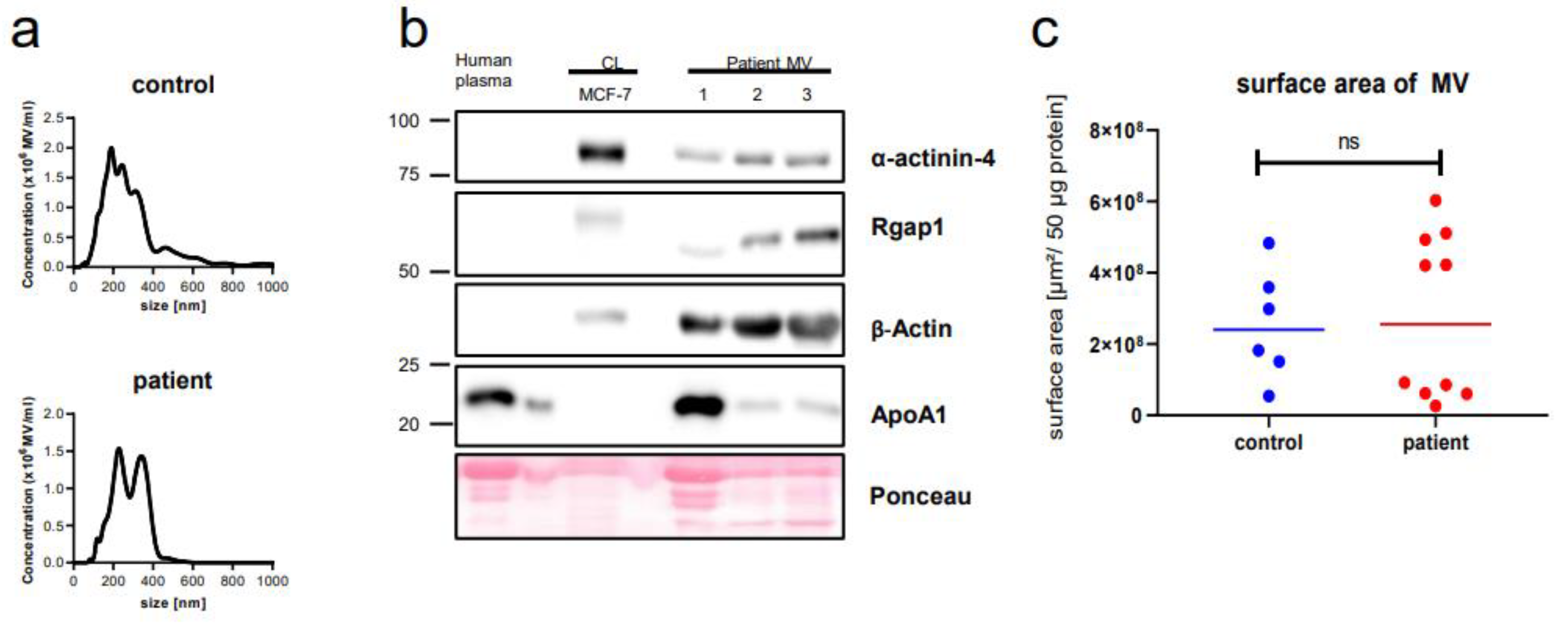
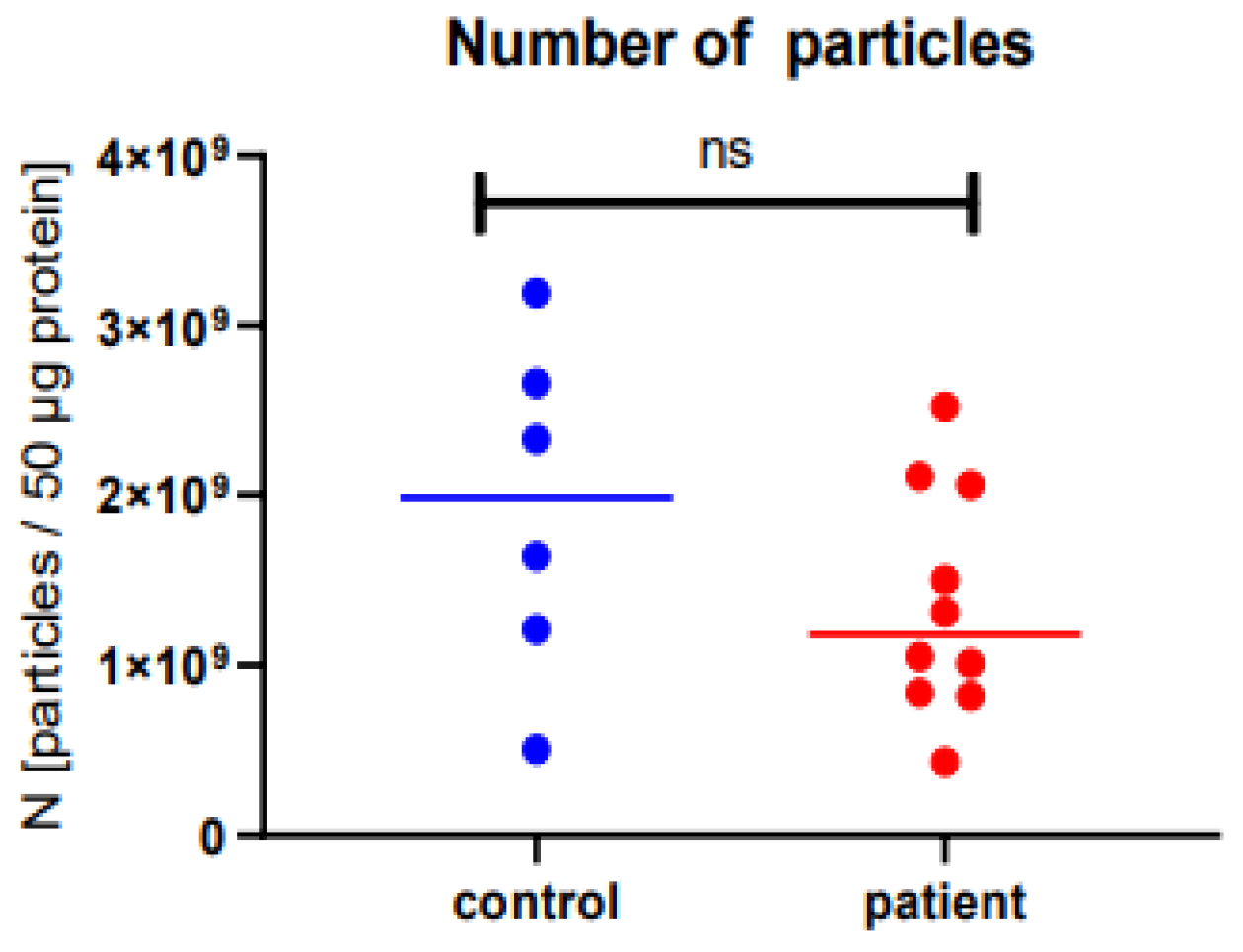
2.1. Microvesicle Harvest and Characterization
2.2. Metabolomics by Mass Spectrometry Is Feasible in Blood-Derived Microvesicles
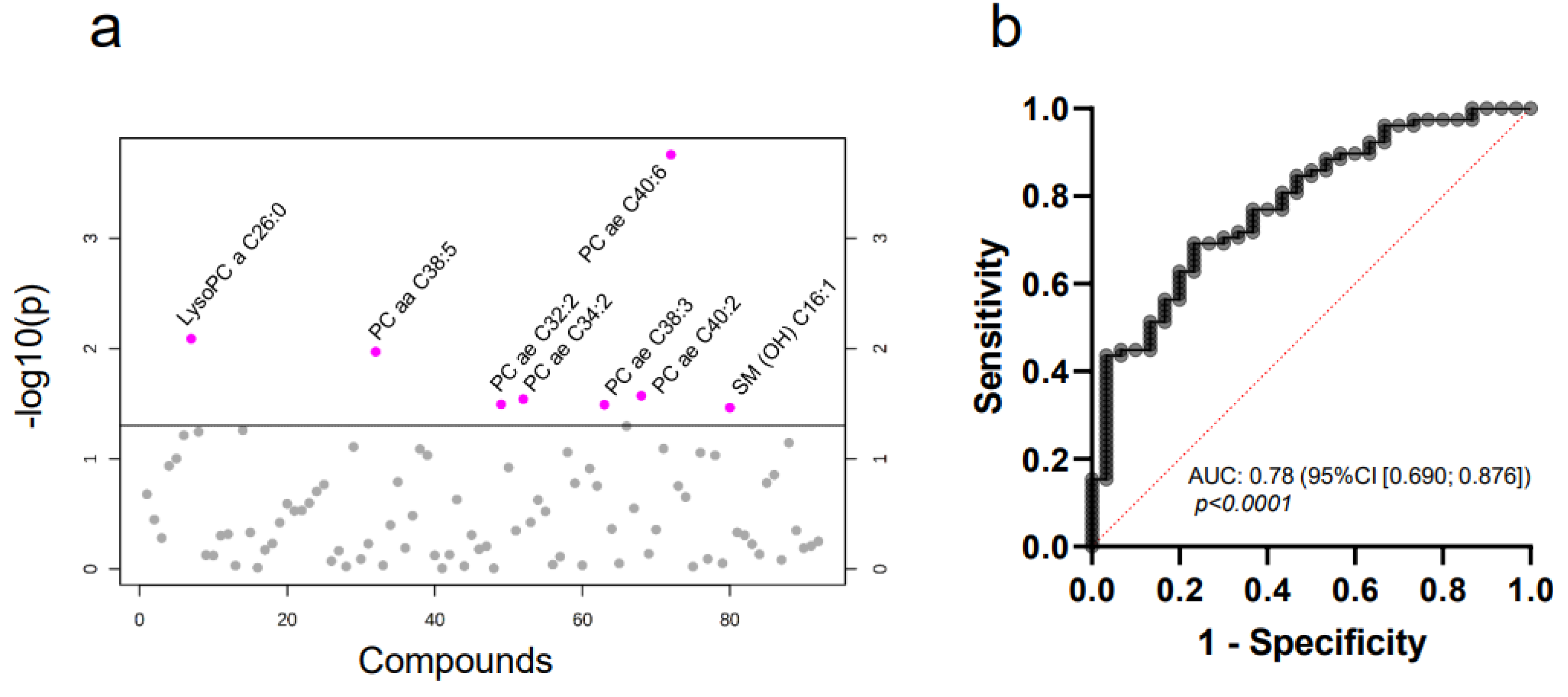
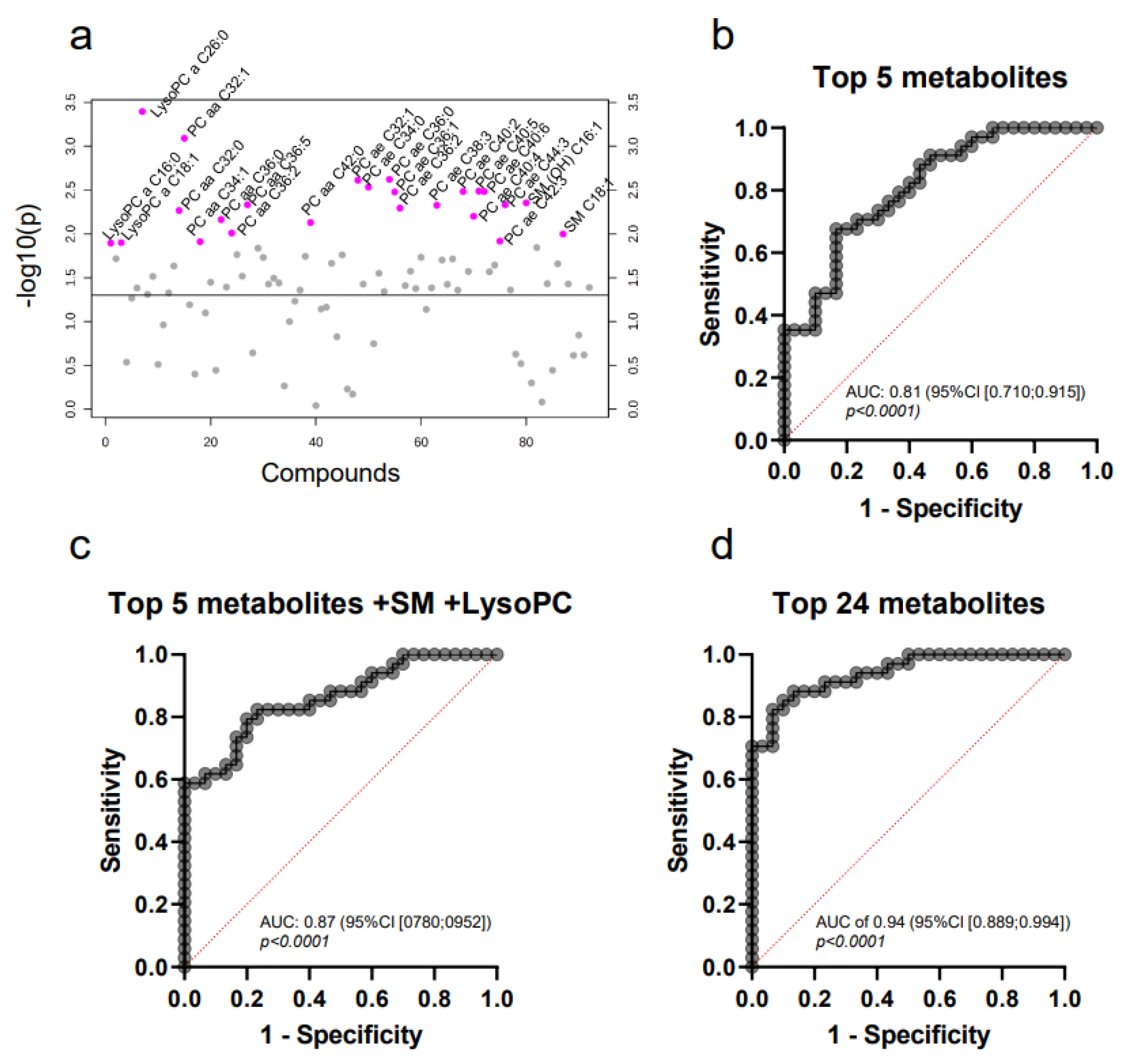
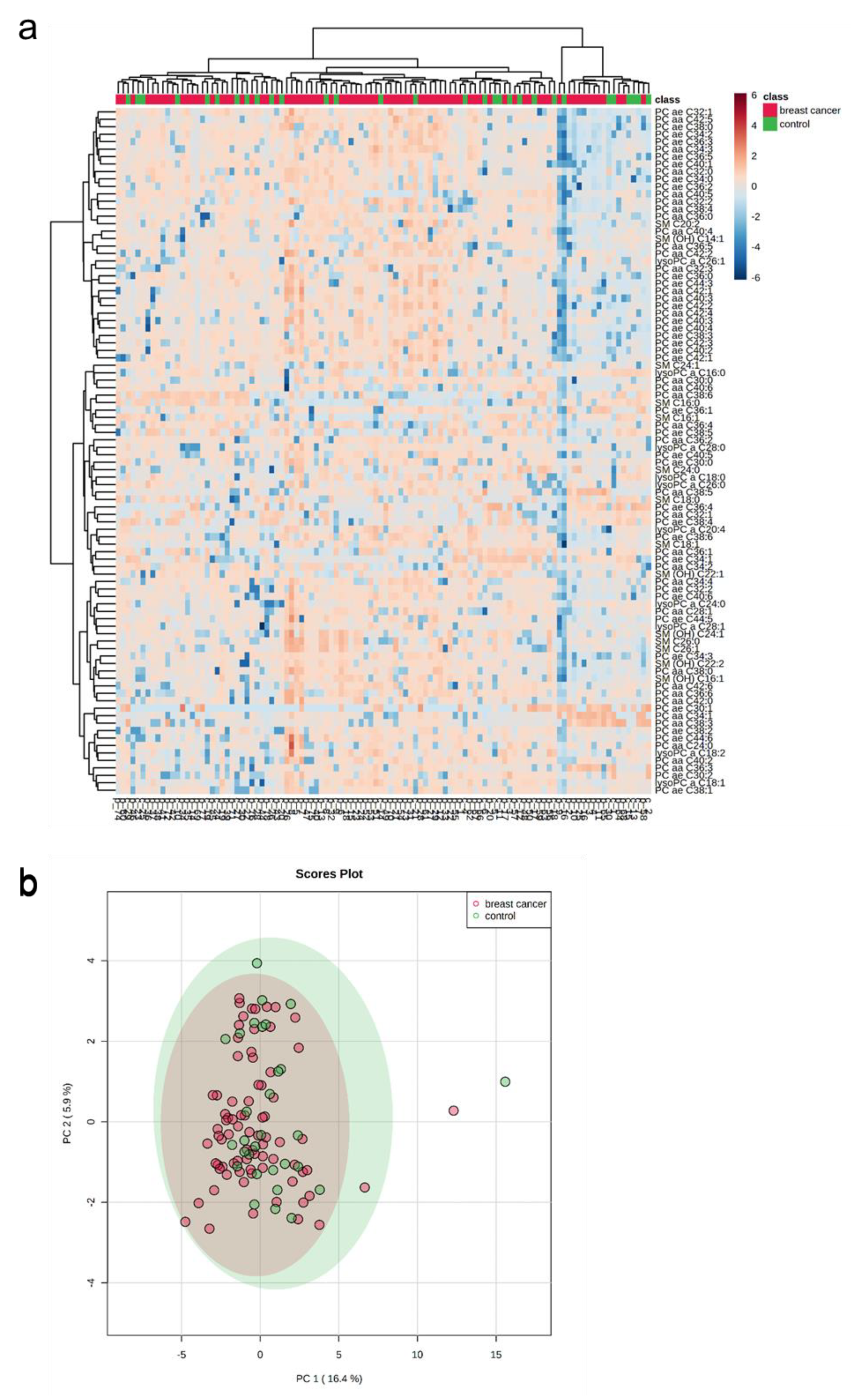
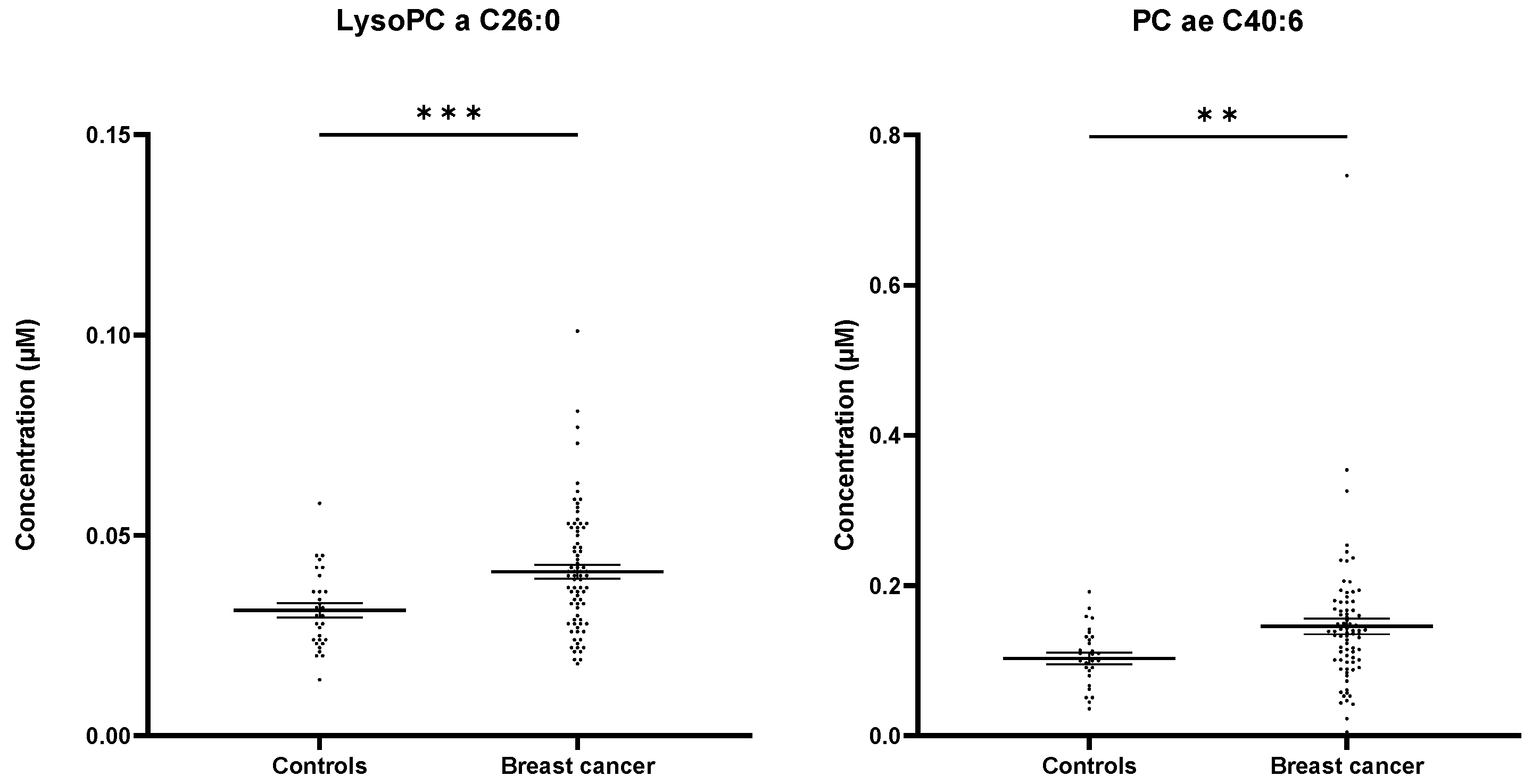
2.3. Targeted Mass Spectrometry Reveals Differences in the Whole MV Metabolome of Breast Cancer Patients and Controls
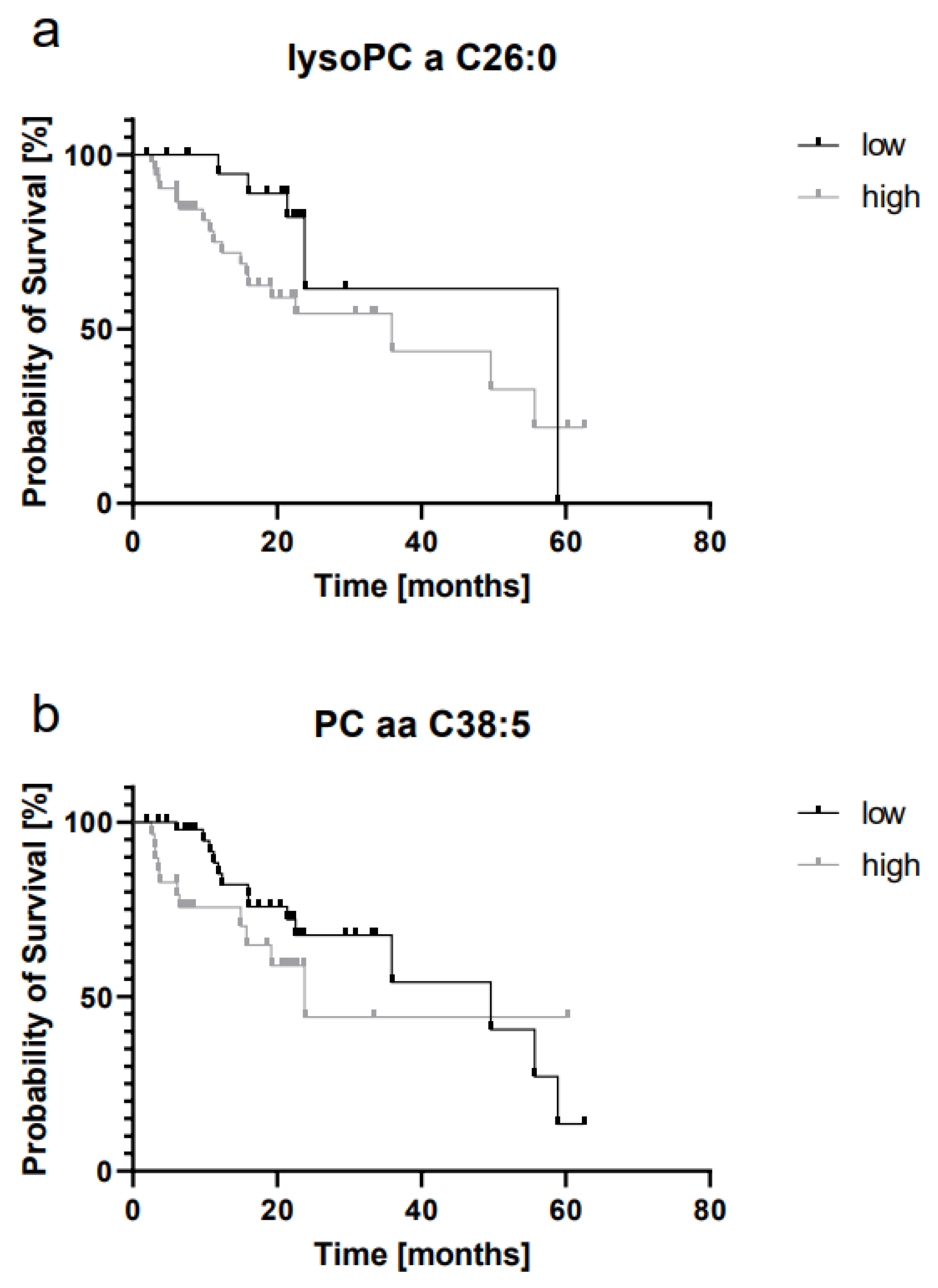
2.4. LysoPC a C26:0 and PC aa C38:5 Levels Are Prognostic for Overall Survival
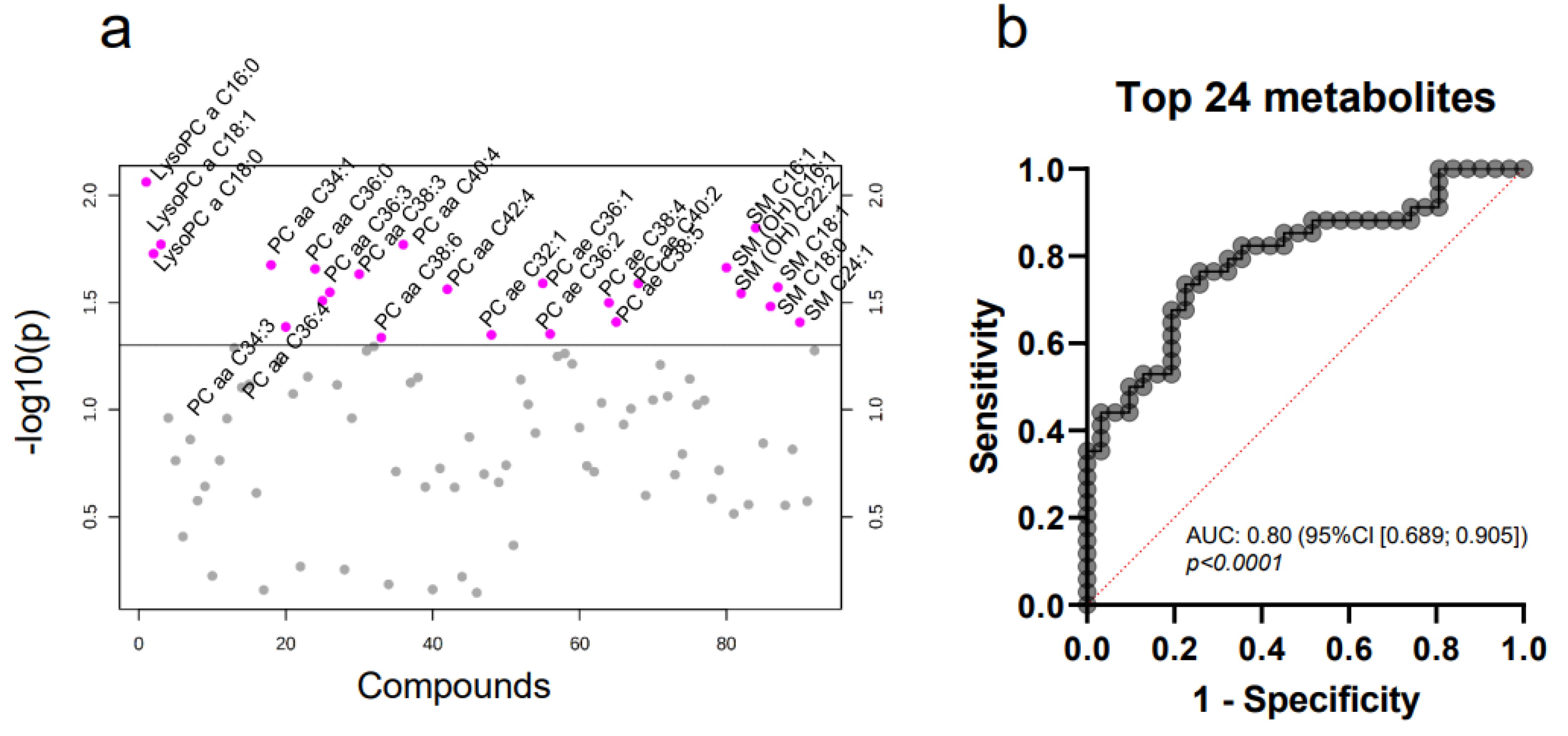
2.5. The Whole Blood MV Metabolome Differentiates between Molecular Breast Cancer Subtypes
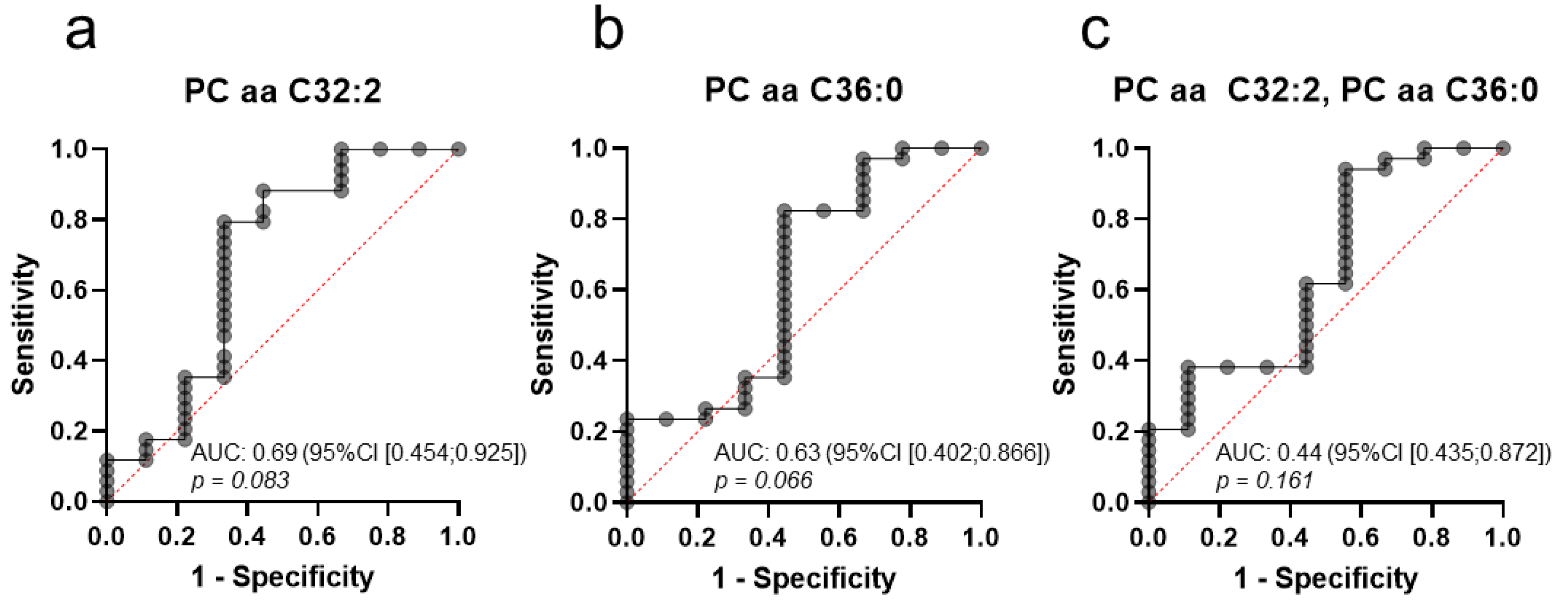
2.6. A Distinct Metabolic Profile Discriminates between Subtype Luminal B and Healthy Controls
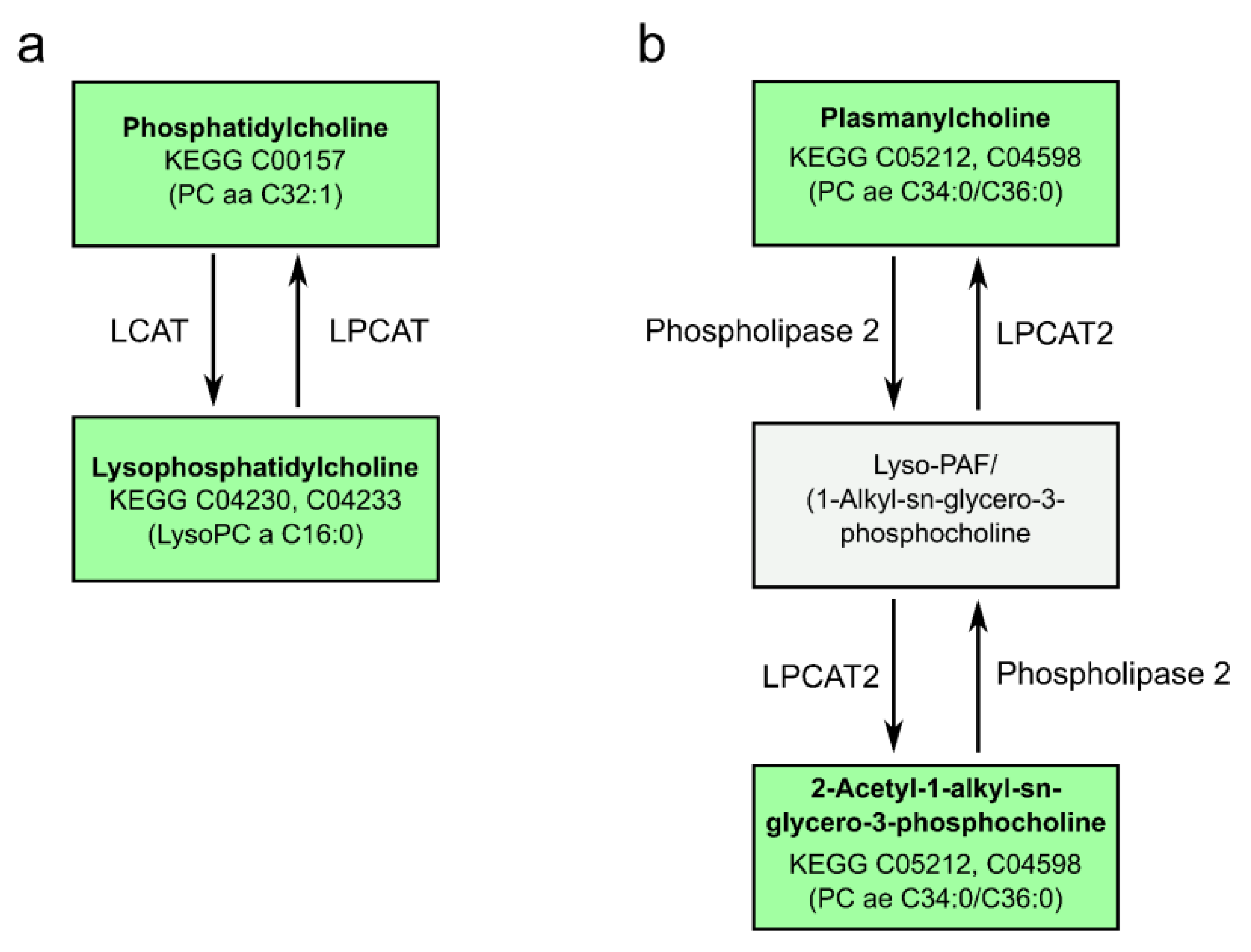
2.7. Pathway Analysis Reveals Alterations in Glycerophospholipid and Ether Lipid Metabolism as Well as Linoleic Acid Metabolism
3. Discussion
4. Materials and Methods
4.1. Patient Recruitment and Data Extraction
4.2. Sample Preparation and Mass Spectrometry
4.3. Nanoparticle Tracking Analysis (NTA)
4.4. Western Blot
4.5. Statistical Analysis and Bioinformatics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Class | Metabolites Detected |
|---|---|
| LysoPC | lysoPC a C16:0 lysoPC a C18:0 lysoPC a C18:1 lysoPC a C18:2 lysoPC a C20:4 lysoPC a C24:0 lysoPC a C26:0 lysoPC a C26:1 lysoPC a C28:0 lysoPC a C28:1 |
| PC aa | PC aa C24:0 PC aa C28:1 PC aa C30:0 PC aa C32:0 PC aa C32:1 PC aa C32:2 PC aa C32:3 PC aa C34:1 PC aa C34:2 PC aa C34:3 PC aa C34:4 PC aa C36:0 PC aa C36:1 PC aa C36:2 PC aa C36:3 PC aa C36:4 PC aa C36:5 PC aa C36:6 PC aa C38:0 PC aa C38:3 PC aa C38:4 PC aa C38:5 PC aa C38:6 PC aa C40:2 PC aa C40:3 PC aa C40:4 PC aa C40:5 PC aa C40:6 PC aa C42:0 PC aa C42:1 PC aa C42:2 PC aa C42:4 PC aa C42:5 PC aa C42:6 |
| PC ae | PC ae C30:0 PC ae C30:1 PC ae C30:2 PC ae C32:1 PC ae C32:2 PC ae C34:0 PC ae C34:1 PC ae C34:2 PC ae C34:3 PC ae C36:0 PC ae C36:1 PC ae C36:2 PC ae C36:3 PC ae C36:4 PC ae C36:5 PC ae C38:0 PC ae C38:1 PC ae C38:2 PC ae C38:3 PC ae C38:4 PC ae C38:5 PC ae C38:6 PC ae C40:1 PC ae C40:2 PC ae C40:3 PC ae C40:4 PC ae C40:5 PC ae C40:6 PC ae C42:1 PC ae C42:2 PC ae C42:3 PC ae C44:3 PC ae C44:5 PC ae C44:6 |
| SM | SM C16:0 SM C16:1 SM C18:0 SM C18:1 SM C20:2 SM C24:0 SM C24:1 SM C26:0 SM C26:1 |
| SM (OH) | SM (OH) C14:1 SM (OH) C16:1 SM (OH) C22:1 SM (OH) C22:2 SM (OH) C24:1 |
| Metabolite | t-Value | p-Value | FDR | |
|---|---|---|---|---|
| Her2-enriched Luminal A | PC aa C24:0 | 2.3912 | 0.02266 | 0.9786 |
| Her2-enriched Luminal B | PC ae C40:4 | −2.0812 | 0.0446 | 0.9456 |
| Basal-like Luminal B | PC aa C32:2 | −2.6532 | 0.0113 | 0.3688 |
| PC aa C36:0 | −2.5427 | 0.0149 | 0.3688 | |
| Luminal A Luminal B | lysoPC a C16:0 | −2.7094 | 0.0087 | 0.1687 |
| SM C16:0 | −2.5227 | 0.0142 | 0.1687 | |
| lysoPC a C18:1 | −2.4528 | 0.0170 | 0.1687 | |
| PC aa C40:4 | −2.4522 | 0.0170 | 0.1687 | |
| lysoPC a C18:0 | −2.4133 | 0.0187 | 0.1687 | |
| PC aa C34:1 | −2.3647 | 0.0211 | 0.1687 | |
| SM OH C16:1 | −2.3531 | 0.0218 | 0.1687 | |
| PC aa C36:2 | −2.3475 | 0.0220 | 0.1687 | |
| PC aa C38:3 | −2.3247 | 0.0233 | 0.1687 | |
| PC ae C36:1 | −2.2838 | 0.0258 | 0.1687 | |
| PC ae C40:2 | −2.2831 | 0.0258 | 0.1687 | |
| SM C18:1 | −2.2667 | 0.0269 | 0.1687 | |
| PC aa C42:4 | −2.2577 | 0.0274 | 0.1687 | |
| PC aa C36:4 | −2.2442 | 0.0283 | 0.1687 | |
| SM OH C22:2 | −2.2387 | 0.0287 | 0.1687 | |
| PC aa C36:3 | −2.2061 | 0.0310 | 0.1687 | |
| PC ae C38:4 | −2.1969 | 0.0317 | 0.1687 | |
| SM C18:0 | −2.1799 | 0.0330 | 0.1687 | |
| PC ae C38:5 | −2.1078 | 0.0390 | 0.1723 | |
| SM C24:1 | −2.1067 | 0.0391 | 0.1723 | |
| PC aa C34:3 | −2.0851 | 0.0411 | 0.1723 | |
| PC ae C36:2 | −2.0515 | 0.0444 | 0.1723 | |
| PC ae C32:1 | −2.0474 | 0.0448 | 0.1723 | |
| PC aa C38:6 | −2.0347 | 0.0461 | 0.1723 |
| Metabolite | t-Value | p-Value | FDR | |
|---|---|---|---|---|
| Luminal A | lysoPC a C26:0 | −2.0632 | 0.0435 | 0.8630 |
| Luminal B | lysoPC a C26:0 | −3.7419 | 0.0004 | 0.0332 |
| PC aa C32:1 | −3.5215 | 0.0008 | 0.0332 | |
| PC ae C36:0 | −3.1676 | 0.0024 | 0.0332 | |
| PC ae C32:1 | −3.1676 | 0.0024 | 0.0332 | |
| PC ae C34:0 | −3.0978 | 0.0029 | 0.0332 | |
| PC ae C40:5 | −3.0619 | 0.0033 | 0.0332 | |
| PC ae C40:6 | −3.0584 | 0.0033 | 0.0332 | |
| PC ae C40:2 | −3.0581 | 0.0033 | 0.0332 | |
| PC ae C36:1 | −3.0542 | 0.0033 | 0.0332 | |
| SM OH C16:1 | −2.9544 | 0.0044 | 0.0332 | |
| PC aa C36:5 | −2.9370 | 0.0046 | 0.0332 | |
| PC ae C44:3 | −2.9368 | 0.0046 | 0.0332 | |
| PC ae C38:3 | −2.9318 | 0.0047 | 0.0332 | |
| PC ae C36:2 | −2.9057 | 0.0051 | 0.0332 | |
| PC aa C32:0 | −2.8831 | 0.0054 | 0.0332 | |
| PC ae C40:4 | −2.8284 | 0.0063 | 0.0362 | |
| PC aa C36:0 | −2.7971 | 0.0069 | 0.0371 | |
| PC aa C42:0 | −2.7682 | 0.0074 | 0.0379 | |
| PC aa C36:2 | −2.6660 | 0.0098 | 0.0461 | |
| SM C18:1 | −2.6564 | 0.0100 | 0.0461 | |
| PC ae C42:3 | −2.5854 | 0.0121 | 0.0487 | |
| PC aa C34:1 | −2.5802 | 0.0123 | 0.0487 | |
| lysoPC a C18:1 | −2.5700 | 0.0126 | 0.0487 | |
| lysoPC a C16:0 | −2.5666 | 0.0127 | 0.0487 | |
| Her2-enriched | lysoPC a C26:0 | −3.1351 | 0.0037 | 0.3374 |
| Author | Overlapping Metabolites (All Breast Cancer Patients) | Non-Overlapping Metabolites Published |
|---|---|---|
| Dìaz-Béltran et al. (blood plasma) | 0 | LyoPC a C14:0 LysoPC a C16:0 LysoPC a C23:0 Biliverdin |
| Ide et al., 2013 Uchiyama et al., 2014 Hosokawa et al., 2017 (tissue) | PC aa C38:5 | PC aa C36:1 PCaa C34:1 PC aa C38:6 PC aa C32:1 PC aa C34:0 PC aa C30:0 |
| Author | Overlapping metabolites (luminal B patients) | Non-overlapping metabolites published |
| Dìaz-Béltran et al. (blood plasma) | LysoPC a C16:0 | LyoPC a C14:0 LysoPC a C23:0 Biliverdin |
| Ide et al., 2013 Uchiyama et al., 2014 Hosokawa et al., 2017 (tissue) | PC aa C32:1 | PC aa C38:6 PC aa C34:0 PC aa C30:0 PC aa C38:5 PC aa C36:1 PC aa C34:1 |
References
- Menck, K.; Sivaloganathan, S.; Bleckmann, A.; Binder, C. Microvesicles in Cancer: Small Size, Large Potential. Int. J. Mol. Sci. 2020, 21, 5373. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezouar, S.; Mege, D.; Darbousset, R.; Farge, D.; Debourdeau, P.; Dignat-George, F.; Panicot-Dubois, L.; Dubois, C. Involvement of platelet-derived microparticles in tumor progression and thrombosis. Semin. Oncol. 2014, 41, 346–358. [Google Scholar] [CrossRef]
- Wang, H.-X.; Gires, O. Tumor-derived extracellular vesicles in breast cancer: From bench to bedside. Cancer Lett. 2019, 460, 54–64. [Google Scholar] [CrossRef]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menck, K.; Bleckmann, A.; Wachter, A.; Hennies, B.; Ries, L.; Schulz, M.; Balkenhol, M.; Pukrop, T.; Schatlo, B.; Rost, U.; et al. Characterisation of tumour-derived microvesicles in cancer patients’ blood and correlation with clinical outcome. J. Extracell. Vesicles 2017, 6, 1340745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, R.; O’Driscoll, L. Extracellular vesicles in blood: Are they viable as diagnostic and predictive tools in breast cancer? Drug Discov. Today 2020, 26, 778–785. [Google Scholar] [CrossRef]
- Tellez-Gabriel, M.; Knutsen, E.; Perander, M. Current Status of Circulating Tumor Cells, Circulating Tumor DNA, and Exosomes in Breast Cancer Liquid Biopsies. Int. J. Mol. Sci. 2020, 21, 9457. [Google Scholar] [CrossRef]
- Díaz-Beltrán, L.; González-Olmedo, C.; Luque-Caro, N.; Díaz, C.; Martín-Blázquez, A.; Fernández-Navarro, M.; Ortega-Granados, A.L.; Gálvez-Montosa, F.; Vicente, F.; Pérez Del Palacio, J.; et al. Human Plasma Metabolomics for Biomarker Discovery: Targeting the Molecular Subtypes in Breast Cancer. Cancers 2021, 13, 147. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhou, B.; Su, M.; Baxter, S.; Zheng, X.; Zhao, X.; Yen, Y.; Jia, W. Mass Spectrometry-Based Quantitative Metabolomics Revealed a Distinct Lipid Profile in Breast Cancer Patients. Int. J. Mol. Sci. 2013, 14, 8047–8061. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, Y.; Masaki, N.; Takei, S.; Horikawa, M.; Matsushita, S.; Sugiyama, E.; Ogura, H.; Shiiya, N.; Setou, M. Recurrent triple-negative breast cancer (TNBC) tissues contain a higher amount of phosphatidylcholine (32:1) than non-recurrent TNBC tissues. PLoS ONE 2017, 12, e0183724. [Google Scholar] [CrossRef] [Green Version]
- Ide, Y.; Waki, M.; Hayasaka, T.; Nishio, T.; Morita, Y.; Tanaka, H.; Sasaki, T.; Koizumi, K.; Matsunuma, R.; Hosokawa, Y. Human breast cancer tissues contain abundant phosphatidylcholine (36:1) with high stearoyl-CoA desaturase-1 expression. PLoS ONE 2013, 8, e61204. [Google Scholar] [CrossRef]
- Yamashita, Y.; Nishiumi, S.; Kono, S.; Takao, S.; Azuma, T.; Yoshida, M. Differences in elongation of very long chain fatty acids and fatty acid metabolism between triple-negative and hormone receptor-positive breast cancer. BMC Cancer 2017, 17, 589. [Google Scholar] [CrossRef]
- Xavier Assad, D.; Acevedo, A.C.; Cançado Porto Mascarenhas, E.; Costa Normando, A.G.; Pichon, V.; Chardin, H.; Neves Silva Guerra, E.; Combes, A. Using an Untargeted Metabolomics Approach to Identify Salivary Metabolites in Women with Breast Cancer. Metabolites 2020, 10, 506. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.L.; Olival, A.; Perestrelo, R.; Silva, P.; Tomás, H.; Câmara, J.S. Untargeted Urinary 1H NMR-Based Metabolomic Pattern as a Potential Platform in Breast Cancer Detection. Metabolites 2019, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Nishida-Aoki, N.; Izumi, Y.; Takeda, H.; Takahashi, M.; Ochiya, T.; Bamba, T. Lipidomic Analysis of Cells and Extracellular Vesicles from High- and Low-Metastatic Triple-Negative Breast Cancer. Metabolites 2020, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Roberg-Larsen, H.; Lund, K.; Seterdal, K.E.; Solheim, S.; Vehus, T.; Solberg, N.; Krauss, S.; Lundanes, E.; Wilson, S.R. Mass spectrometric detection of 27-hydroxycholesterol in breast cancer exosomes. J. Steroid Biochem. Mol. Biol. 2017, 169, 22–28. [Google Scholar] [CrossRef]
- Clos-Garcia, M.; Loizaga-Iriarte, A.; Zuñiga-Garcia, P.; Sánchez-Mosquera, P.; Rosa Cortazar, A.; González, E.; Torrano, V.; Alonso, C.; Pérez-Cormenzana, M.; Ugalde-Olano, A. Metabolic alterations in urine extracellular vesicles are associated to prostate cancer pathogenesis and progression. J. Extracell. Vesicles 2018, 7, 1470442. [Google Scholar] [CrossRef] [Green Version]
- Puhka, M.; Takatalo, M.; Nordberg, M.-E.; Valkonen, S.; Nandania, J.; Aatonen, M.; Yliperttula, M.; Laitinen, S.; Velagapudi, V.; Mirtti, T.; et al. Metabolomic Profiling of Extracellular Vesicles and Alternative Normalization Methods Reveal Enriched Metabolites and Strategies to Study Prostate Cancer-Related Changes. Theranostics 2017, 7, 3824–3841. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Lee, J.C.; Byeon, S.K.; Rha, K.H.; Moon, M.H. Size Dependent Lipidomic Analysis of Urinary Exosomes from Patients with Prostate Cancer by Flow Field-Flow Fractionation and Nanoflow Liquid Chromatography-Tandem Mass Spectrometry. Anal. Chem. 2017, 89, 2488–2496. [Google Scholar] [CrossRef]
- Fan, T.W.M.; Zhang, X.; Wang, C.; Yang, Y.; Kang, W.-Y.; Arnold, S.; Higashi, R.M.; Liu, J.; Lane, A.N. Exosomal lipids for classifying early and late stage non-small cell lung cancer. Anal. Chim. Acta 2018, 1037, 256–264. [Google Scholar] [CrossRef]
- Tao, L.; Zhou, J.; Yuan, C.; Zhang, L.; Li, D.; Si, D.; Xiu, D.; Zhong, L. Metabolomics identifies serum and exosomes metabolite markers of pancreatic cancer. Metab. Off. J. Metab. Soc. 2019, 15, 86. [Google Scholar] [CrossRef]
- Menck, K.; Bleckmann, A.; Schulz, M.; Ries, L.; Binder, C. Isolation and Characterization of Microvesicles from Peripheral Blood. J. Vis. Exp. 2017, 119, e55057. [Google Scholar] [CrossRef] [Green Version]
- Sódar, B.W.; Kittel, Á.; Pálóczi, K.; Vukman, K.V.; Osteikoetxea, X.; Szabó-Taylor, K.; Németh, A.; Sperlágh, B.; Baranyai, T.; Giricz, Z.; et al. Low-density lipoprotein mimics blood plasma-derived exosomes and microvesicles during isolation and detection. Sci. Rep. 2016, 6, 24316. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, Y.; Hayasaka, T.; Masaki, N.; Watanabe, Y.; Masumoto, K.; Nagata, T.; Katou, F.; Setou, M. Imaging mass spectrometry distinguished the cancer and stromal regions of oral squamous cell carcinoma by visualizing phosphatidylcholine (16:0/16:1) and phosphatidylcholine (18:1/20:4). Anal. Bioanal. Chem. 2014, 406, 1307–1316. [Google Scholar] [CrossRef]
- Camp, R.L.; Dolled-Filhart, M.; Rimm, D.L. X-tile: A new bio-informatics tool for biomarker assessment and outcome-based cut-point optimization. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 7252–7259. [Google Scholar] [CrossRef] [Green Version]
- Prat, A.; Pineda, E.; Adamo, B.; Galván, P.; Fernández, A.; Gaba, L.; Díez, M.; Viladot, M.; Arance, A.; Muñoz, M. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast Edinb. Scotl. 2015, 24 (Suppl. 2), S26–S35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.J.; Swain, S.M. Luminal A Breast Cancer and Molecular Assays: A Review. Oncologist 2018, 23, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Kurabe, N.; Hayasaka, T.; Ogawa, M.; Masaki, N.; Ide, Y.; Waki, M.; Nakamura, T.; Kurachi, K.; Kahyo, T.; Shinmura, K.; et al. Accumulated phosphatidylcholine (16:0/16:1) in human colorectal cancer; possible involvement of LPCAT4. Cancer Sci. 2013, 104, 1295–1302. [Google Scholar] [CrossRef]
- Gerashchenko, T.S.; Zolotaryova, S.Y.; Kiselev, A.M.; Tashireva, L.A.; Novikov, N.M.; Krakhmal, N.V.; Cherdyntseva, N.V.; Zavyalova, M.V.; Perelmuter, V.M.; Denisov, E.V. The Activity of KIF14, Mieap, and EZR in a New Type of the Invasive Component, Torpedo-Like Structures, Predetermines the Metastatic Potential of Breast Cancer. Cancers 2020, 12, 1909. [Google Scholar] [CrossRef] [PubMed]
- Hahnefeld, L.; Gruber, L.; Schömel, N.; Fischer, C.; Mattjus, P.; Gurke, R.; Beretta, M.; Ferreirós, N.; Geisslinger, G.; Wegner, M.-S. Ether lipid and sphingolipid expression patterns are estrogen receptor-dependently altered in breast cancer cells. Int. J. Biochem. Cell. Biol. 2020, 127, 105834. [Google Scholar] [CrossRef]
- Yi, S.-W.; Ohrr, H.; Shin, S.-A.; Yi, J.-J. Sex-age-specific association of body mass index with all-cause mortality among 12.8 million Korean adults: A prospective cohort study. Int. J. Epidemiol. 2015, 44, 1696–1705. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbassi-Ghadi, N.; Kumar, S.; Huang, J.; Goldin, R.; Takats, Z.; Hanna, G.B. Metabolomic profiling of oesophago-gastric cancer: A systematic review. Eur. J. Cancer. 2013, 49, 3625–3637. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Rutigliano, M.; Galleggiante, V.; Giglio, A.; Palazzo, S.; Ferro, M.; Simone, C.; Bettocchi, C.; Battaglia, M.; Ditonno, P. Metabolomic profiling for the identification of novel diagnostic markers in prostate cancer. Expert Rev. Mol. Diagn. 2015, 15, 1211–1224. [Google Scholar] [CrossRef]
- Pandey, R.; Caflisch, L.; Lodi, A.; Brenner, A.J.; Tiziani, S. Metabolomic signature of brain cancer. Mol. Carcinog. 2017, 56, 2355–2371. [Google Scholar] [CrossRef] [PubMed]
- Kozar, N.; Kruusmaa, K.; Bitenc, M.; Argamasilla, R.; Adsuar, A.; Takač, I.; Arko, D. Identification of Novel Diagnostic Biomarkers in Breast Cancer Using Targeted Metabolomic Profiling. Clin. Breast Cancer 2020, 21, e204–e211. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Misra, B.B. Challenges and Opportunities in Cancer Metabolomics. Proteomics 2019, 19, 1900042. [Google Scholar] [CrossRef] [PubMed]
- Kotland, T.S.; Ekroos, K.; Kauhanen, D.; Simolin, H.; Seierstad, T.; Berge, V.; Sandvig, K.; Llorente, A. Molecular lipid species in urinary exosomes as potential prostate cancer biomarkers. Eur. J. Cancer 2017, 70, 122–132. [Google Scholar] [CrossRef]
- Menck, K.; Sönmezer, C.; Worst, T.S.; Schulz, M.; Dihazi, G.H.; Streit, F.; Erdmann, G.; Kling, S.; Boutros, M.; Binder, C. Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J. Extracell. Vesicles 2017, 6, 1378056. [Google Scholar] [CrossRef]
- Yan, F.; Zhao, H.; Zeng, Y. Lipidomics: A promising cancer biomarker. Clin. Transl. Med. 2018, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wang, X.; Qiu, L.; Qin, X.; Liu, H.; Wang, Y.; Li, F.; Wang, X.; Chen, G.; Song, G. Probing gender-specific lipid metabolites and diagnostic biomarkers for lung cancer using Fourier transform ion cyclotron resonance mass spectrometry. Clin. Chim. Acta Int. J. Clin. Chem. 2012, 414, 135–141. [Google Scholar] [CrossRef]
- Klupczynska, A.; Plewa, S.; Kasprzyk, M.; Dyszkiewicz, W.; Kokot, Z.J.; Matysiak, J. Serum lipidome screening in patients with stage I non-small cell lung cancer. Clin. Exp. Med. 2019, 19, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Ros-Mazurczyk, M.; Jelonek, K.; Marczyk, M.; Binczyk, F.; Pietrowska, M.; Polanska, J.; Dziadziuszko, R.; Jassem, J.; Rzyman, W.; Widlak, P. Serum lipid profile discriminates patients with early lung cancer from healthy controls. Lung Cancer 2017, 112, 69–74. [Google Scholar] [CrossRef]
- Schmidt, J.A.; Fensom, G.K.; Rinaldi, S.; Scalbert, A.; Appleby, P.N.; Achaintre, D.; Gicquiau, A.; Gunter, M.J.; Ferrari, P.; Kaaks, R.; et al. Patterns in metabolite profile are associated with risk of more aggressive prostate cancer: A prospective study of 3057 matched case-control sets from EPIC. Int. J. Cancer 2020, 146, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhou, X.; Xia, T.-S.; Chen, Z.; Li, J.; Liu, Q.; Alolga, R.N.; Chen, Y.; Lai, M.-D.; Li, P.; et al. Human plasma metabolomics for identifying differential metabolites and predicting molecular subtypes of breast cancer. Oncotarget 2016, 7, 9925–9938. [Google Scholar] [CrossRef] [Green Version]
- Menck, K.; Klemm, F.; Gross, J.C.; Pukrop, T.; Wenzel, D.; Binder, C. Induction and transport of Wnt 5a during macrophage-induced malignant invasion is mediated by two types of extracellular vesicles. Oncotarget 2013, 4, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- Munir, M.T.; Kay, M.K.; Kang, M.H.; Rahman, M.M.; Al-Harrasi, A.; Choudhury, M.; Moustaid-Moussa, N.; Hussain, F.; Rahman, S.M. Tumor-Associated Macrophages as Multifaceted Regulators of Breast Tumor Growth. Int. J. Mol. Sci. 2021, 22, 6526. [Google Scholar] [CrossRef]
- Piano, V.; Benjamin, D.I.; Valente, S.; Nenci, S.; Marrocco, B.; Mai, A.; Aliverti, A.; Nomura, D.K.; Mattevi, A. Discovery of Inhibitors for the Ether Lipid-Generating Enzyme AGPS as Anti-Cancer Agents. ACS Chem. Biol. 2015, 10, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmholtz Institute Index of /metap2/metabolites/Details. Available online: http://metap.helmholtz-muenchen.de/metap2/metabolites/Details/ (accessed on 24 April 2021).
- KEGG PATHWAY Database KEGG PATHWAY Database. Available online: https://www.genome.jp/kegg/pathway.html (accessed on 5 October 2021).






| Clinical Characteristics | |
|---|---|
| Age median (patients, min-max range) Age median (healthy controls, min-max range) | 63 (27–92) 39 (21–65) |
| Histological subtype | |
| Invasive ductal | 49 |
| Invasive lobular | 14 |
| Invasive ductal/lobular | 1 |
| Other | 3 |
| Not defined | 11 |
| Molecular subtype | |
| Luminal A | 31 |
| Luminal B | 34 |
| Basal-like | 9 |
| Her2-enriched | 4 |
| Metastasis | |
| Oligometastasis (≤3 metastasis) | 18 |
| Polymetastasis (>3 metastasis) | 50 |
| Location of metastasis | |
| Bone | 48 |
| Brain | 15 |
| Lung | 25 |
| Other | 33 |
| Stable vs. progressive disease | |
| Progressive disease | 29 |
| Stable disease | 49 |
| Metabolic comorbidities of patients | |
| Obesity | 14/76 * |
| Diabetes mellitus | 8/77 ** |
| Lipometabolic disorders | 6/77 ** |
| Lysophosphatidylcholines |
|---|
| LysoPC a C16:0, LysoPC a C18:0, Lyso PC a C18:1, LysoPC a C18:2, LysoPC a C20:4, LysoPC a C24:0 |
| Sphingomyelins |
| SM (OH) C14:1, SM (OH) C22:1, SM(OH) C22:2, SM (OH) C24:1, SM C18:0, SM C18:1, SM C20:2, SM C24:0, SM C26:1 |
| Metabolite | t-Value | p-Value | FDR |
|---|---|---|---|
| PC ae C40:6 | −3.8915 | 0.0002 | 0.016 |
| lysoPC a C26:0 | −2.6967 | 0.0081 | 0.3277 |
| PC aa C38:5 | −2.5989 | 0.0107 | 0.3277 |
| PC ae C40:2 | −2.2459 | 0.0268 | 0.3946 |
| PC ae C34:2 | −2.2170 | 0.0288 | 0.3946 |
| PC ae C32:2 | −2.1730 | 0.0320 | 0.3946 |
| PC ae C38:3 | −2.1701 | 0.0322 | 0.3946 |
| SM (OH) C16:1 | −2.1441 | 0.0343 | 0.3946 |
| Metabolite | Group | N (Patients) | N (Events) | Cut-Off (µmol/L) |
|---|---|---|---|---|
| lysoPC a C26:0 | low | 26 | 5 | 0.03 |
| high | 52 | 30 | ||
| PC aa C38:5 | low | 49 | 13 | 2.56 |
| high | 29 | 9 | ||
| PC ae C32:2 | low | 33 | 8 | 0.03 |
| high | 45 | 17 | ||
| PC ae C34:2 | low | 24 | 6 | 0.39 |
| high | 54 | 19 | ||
| PC ae C38:3 | low | 19 | 6 | 0:27 |
| high | 59 | 19 | ||
| PC ae C40:2 | low | 56 | 18 | 0.18 |
| high | 22 | 7 | ||
| PC ae C40:6 | low | 39 | 10 | 0.14 |
| high | 39 | 15 | ||
| SM (OH) C16:1 | low | 49 | 16 | 0.27 |
| high | 29 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buentzel, J.; Klemp, H.G.; Kraetzner, R.; Schulz, M.; Dihazi, G.H.; Streit, F.; Bleckmann, A.; Menck, K.; Wlochowitz, D.; Binder, C. Metabolomic Profiling of Blood-Derived Microvesicles in Breast Cancer Patients. Int. J. Mol. Sci. 2021, 22, 13540. https://doi.org/10.3390/ijms222413540
Buentzel J, Klemp HG, Kraetzner R, Schulz M, Dihazi GH, Streit F, Bleckmann A, Menck K, Wlochowitz D, Binder C. Metabolomic Profiling of Blood-Derived Microvesicles in Breast Cancer Patients. International Journal of Molecular Sciences. 2021; 22(24):13540. https://doi.org/10.3390/ijms222413540
Chicago/Turabian StyleBuentzel, Judith, Henry Gerd Klemp, Ralph Kraetzner, Matthias Schulz, Gry Helene Dihazi, Frank Streit, Annalen Bleckmann, Kerstin Menck, Darius Wlochowitz, and Claudia Binder. 2021. "Metabolomic Profiling of Blood-Derived Microvesicles in Breast Cancer Patients" International Journal of Molecular Sciences 22, no. 24: 13540. https://doi.org/10.3390/ijms222413540
APA StyleBuentzel, J., Klemp, H. G., Kraetzner, R., Schulz, M., Dihazi, G. H., Streit, F., Bleckmann, A., Menck, K., Wlochowitz, D., & Binder, C. (2021). Metabolomic Profiling of Blood-Derived Microvesicles in Breast Cancer Patients. International Journal of Molecular Sciences, 22(24), 13540. https://doi.org/10.3390/ijms222413540