
Impact of an MT-RNR1 Gene Polymorphism on Hepatocellular Carcinoma Progression and Clinical Characteristics

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
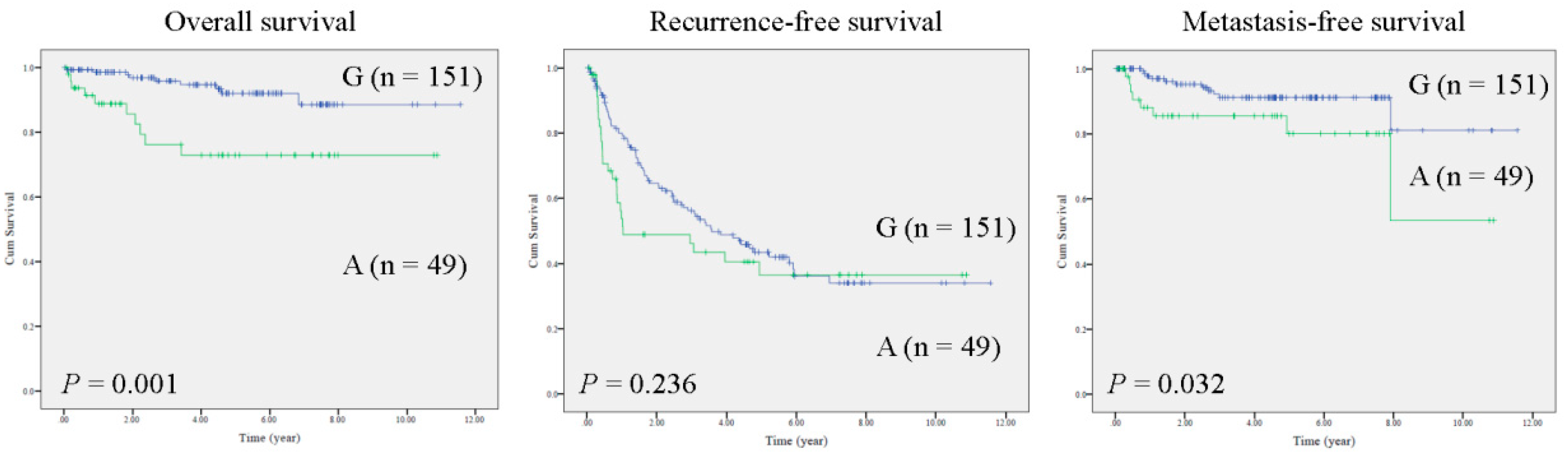
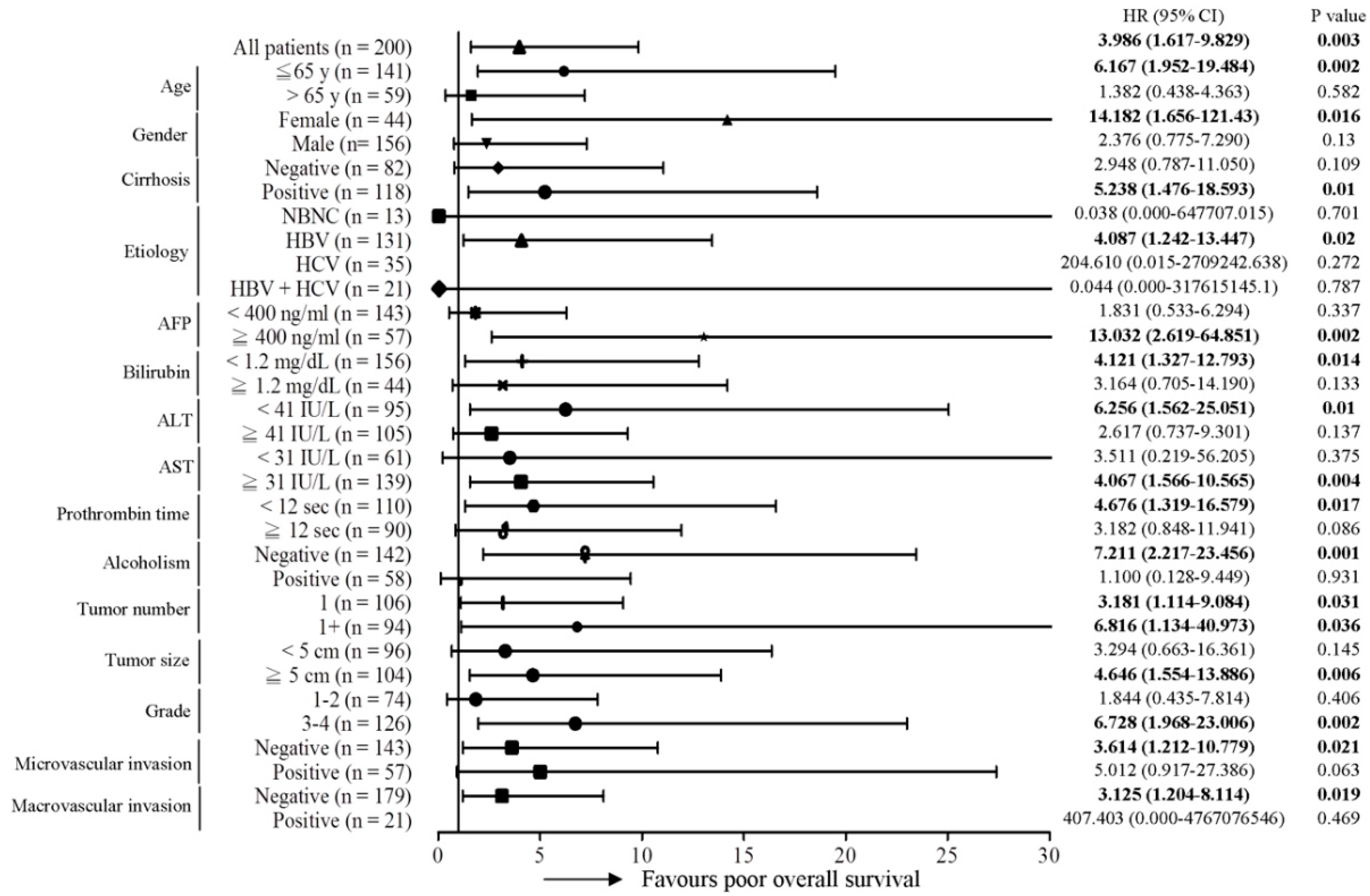
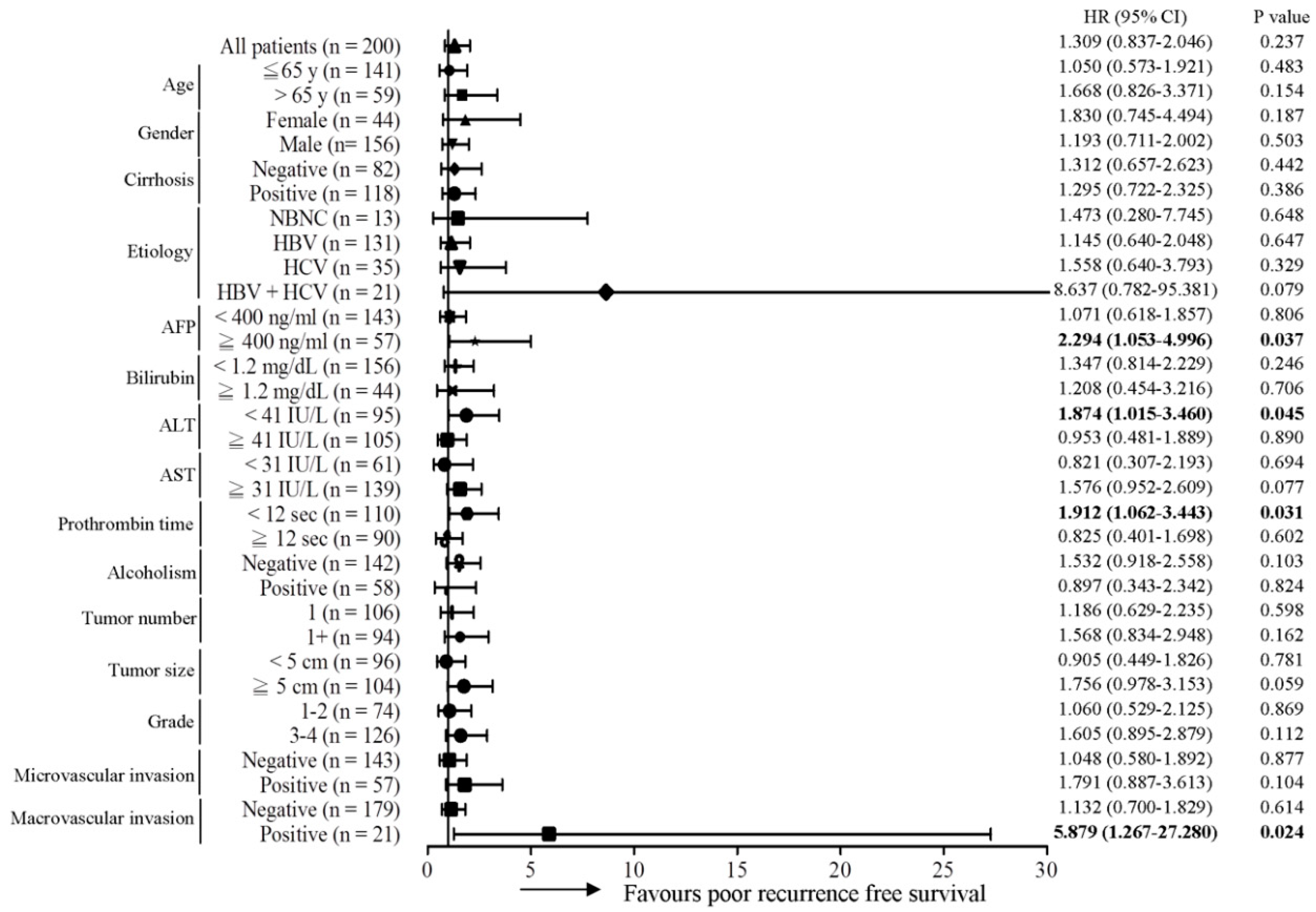
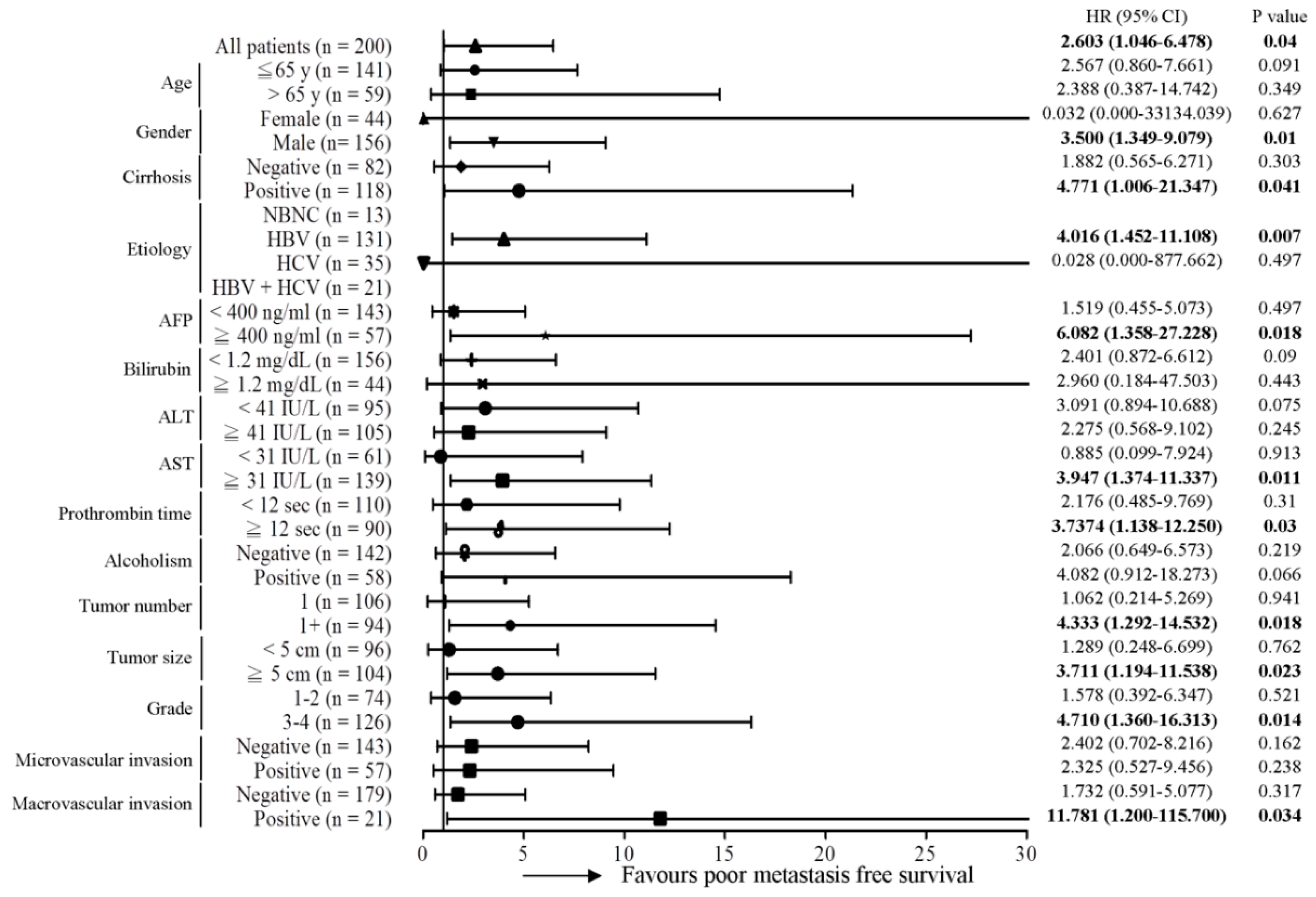
2.1. The MT-RNR1 (G709A) Polymorphism Is an Independent Prognostic Factor Associated with Survival in HCC
2.2. MT-RNR1 709A Is an Effective Prognostic Predictor in Subgroups of HCC
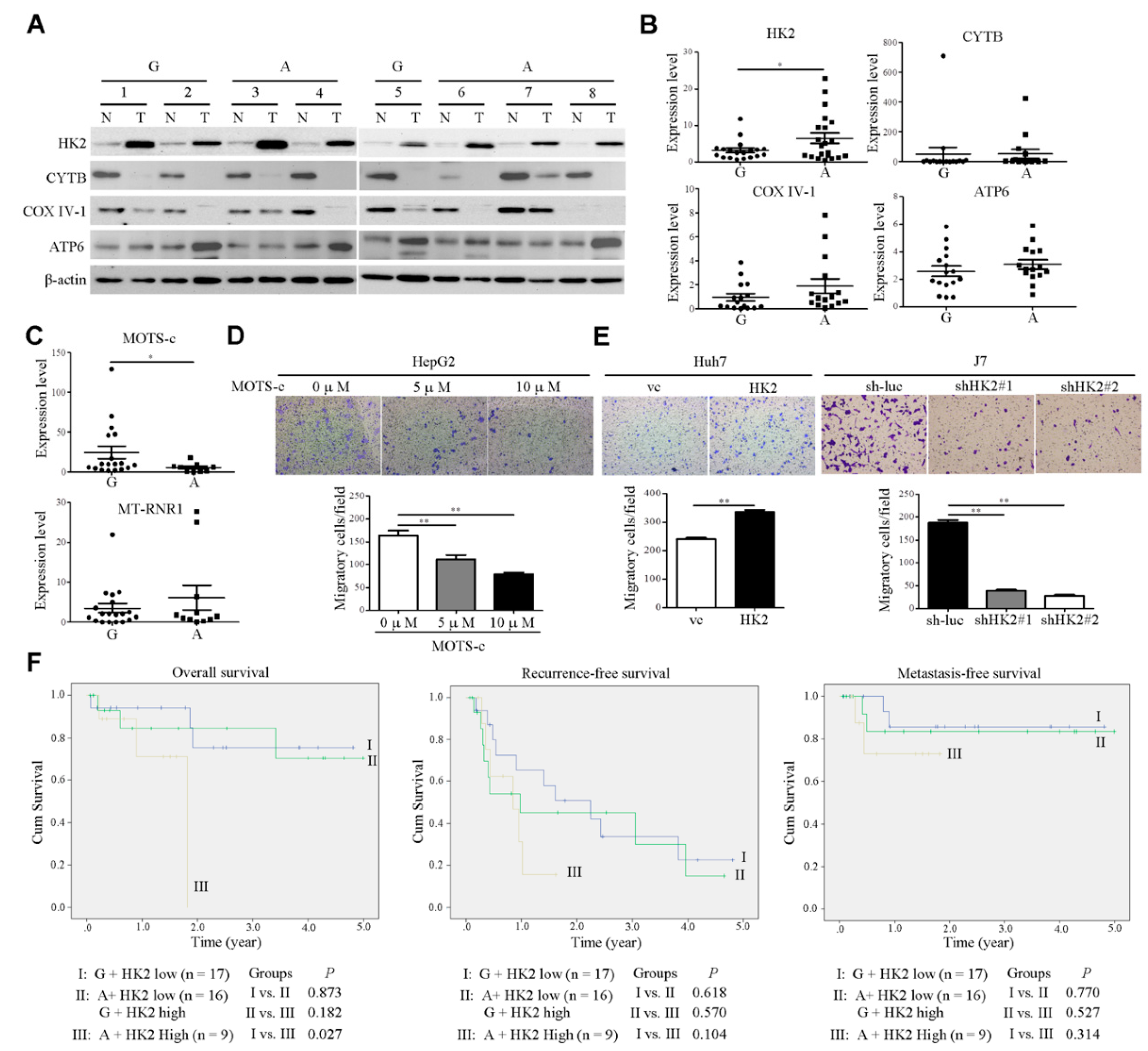
2.3. The Impact of MT-RNR1 G709A on Hexokinase 2 and Mitochondrial Open-Reading-Frame of the Twelve S rRNA-c Expression
3. Discussion
4. Materials and Methods
4.1. Human HCC Samples
4.2. Mutation Analysis of Entire Mitochondrial Genome
4.3. Immunoblot Analysis
4.4. Establishment of HK2 Overexpression or Knockdown Stable Cell Lines
4.5. In Vitro Migration Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| mtDNA | mitochondrial DNA |
| HCC | hepatocellular carcinoma |
| OXPHOS | oxidative phosphorylation |
| ATP6 | mitochondrially encoded ATP synthase membrane subunit 6 |
| ND | NADH dehydrogenase |
| ROS | reactive oxygen species |
| NSCLC | non-small cell lung cancer |
| cybrid | cytoplasmic hybrid |
| SNP | single nucleotide polymorphism |
| AFP | alpha-fetoprotein |
| Bil | bilirubin |
| ALT | alanine transaminase |
| AST | aspartate transaminase |
| PT | prothrombin time |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| HR | hazard ratio |
| HK2 | hexokinase 2 |
| MOTS-c | mitochondrial open-reading-frame of the twelve S rRNA-c |
| TUG1 | taurine upregulated gene 1 |
References
- Patananan, A.N.; Wu, T.-H.; Chiou, P.-Y.; Teitell, M.A. Modifying the Mitochondrial Genome. Cell Metab. 2016, 23, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruda, A.P.; Pers, B.M.; Parlakgül, G.; Güney, E.; Inouye, K.; Hotamisligil, G.S. Chronic enrichment of hepatic endoplasmic reticulum–mitochondria contact leads to mitochondrial dysfunction in obesity. Nat. Med. 2014, 20, 1427–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swerdlow, R.H. Mitochondria and cell bioenergetics: Increasingly recognized components and a possible etiologic cause of Alzheimer’s disease. Antioxid. Redox. Signal. 2012, 16, 1434–1455. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.M.; Carl, S.M.; Swerdlow, R.H. Cytoplasmic hybrid (cybrid) cell lines as a practical model for mitochondriopathies. Redox Biol. 2014, 2, 619–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeley, K.P.; Bray, A.W.; Westbrook, D.G.; Johnson, L.W.; Kesterson, R.A.; Ballinger, S.W.; Welch, D. Mitochondrial Genetics Regulate Breast Cancer Tumorigenicity and Metastatic Potential. Cancer Res. 2015, 75, 4429–4436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.V.; Dong, L.; Neuzil, J. Mitochondrial DNA in Tumor Initiation, Progression, and Metastasis: Role of Horizontal mtDNA Transfer. Cancer Res. 2015, 75, 3203–3208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivekanandan, P.; Daniel, H.; Yeh, M.M.; Torbenson, M. Mitochondrial mutations in hepatocellular carcinomas and fibrolamellar carcinomas. Mod. Pathol. 2010, 23, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Chinnery, P.F.; Hudson, G. Mitochondrial genetics. Br. Med Bull. 2013, 106, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Usami, S.; Nishio, S. Nonsyndromic Hearing Loss and Deafness, Mitochondrial. In GeneReviews; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Mirzaa, G., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Guaran, V.; Astolfi, L.; Castiglione, A.; Simoni, E.; Olivetto, E.; Galasso, M.; Trevisi, P.; Busi, M.; Volinia, S.; Martini, A. Association between idiopathic hearing loss and mitochondrial DNA mutations: A study on 169 hearing-impaired subjects. Int. J. Mol. Med. 2013, 32, 785–794. [Google Scholar] [CrossRef]
- Abril, J.; De Heredia, M.L.; González, L.; Clèries, R.; Nadal, M.; Condom, E.; Aguiló, F.; Gómez-Zaera, M.; Nunes, V. Altered expression of12S/MT-RNR1,MT-CO2/COX2, andMT-ATP6mitochondrial genes in prostate cancer. Prostate 2008, 68, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Bragoszewski, P.; Kupryjanczyk, J.; Bartnik, E.; Rachinger, A.; Ostrowski, J. Limited clinical relevance of mitochondrial DNA mutation and gene expression analyses in ovarian cancer. BMC Cancer 2008, 8, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffarpour, M.; Mahdian, R.; Fereidooni, F.; Goh, Y.-M.; Moazami, N.; Houshmand, M. The mitochondrial ATPase6 gene is more susceptible to mutation than the ATPase8 gene in breast cancer patients. Cancer Cell Int. 2014, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshikawa, N.; Akimoto, M.; Hayashi, J.-I.; Nagase, H.; Takenaga, K. Association of predicted pathogenic mutations in mitochondrial ND genes with distant metastasis in NSCLC and colon cancer. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Takenaga, K.; Akimoto, M.; Koshikawa, N.; Yamaguchi, A.; Imanishi, H.; Nakada, K.; Honma, Y.; Hayashi, J.-I. ROS-Generating Mitochondrial DNA Mutations Can Regulate Tumor Cell Metastasis. Science 2008, 320, 661–664. [Google Scholar] [CrossRef] [Green Version]
- Goswami, P.C. Mutant mitochondria and cancer cell metastasis: Quest for a mechanism. Cancer Biol. Ther. 2009, 8, 1386–1388. [Google Scholar] [CrossRef] [Green Version]
- Kulawiec, M.; Owens, K.M.; Singh, K.K. Cancer cell mitochondria confer apoptosis resistance and promote metastasis. Cancer Biol. Ther. 2009, 8, 1378–1385. [Google Scholar] [CrossRef] [Green Version]
- Mehlen, P.; Puisieux, A. Metastasis: A question of life or death. Nat. Rev. Cancer 2006, 6, 449–458. [Google Scholar] [CrossRef]
- Tang, Z.-Y. Hepatocellular Carcinoma-Cause, Treatment and Metastasis. World J. Gastroenterol. 2001, 7, 445–454. [Google Scholar] [CrossRef]
- Yamada, S.; Nomoto, S.; Fujii, T.; Kaneko, T.; Takeda, S.; Inoue, S.; Kanazumi, N.; Nakao, A. Correlation between copy number of mitochondrial DNA and clinico-pathologic parameters of hepatocellular carcinoma. Eur. J. Surg. Oncol. (EJSO) 2006, 32, 303–307. [Google Scholar] [CrossRef]
- Yin, P.-H.; Wu, C.-C.; Lin, J.-C.; Chi, C.-W.; Wei, Y.-H.; Lee, H.-C. Somatic mutations of mitochondrial genome in hepatocellular carcinoma. Mitochondrion 2010, 10, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Haran, M.; Gross, A. Balancing glycolysis and mitochondrial OXPHOS: Lessons from the hematopoietic system and exercising muscles. Mitochondrion 2014, 19, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J. Energy metabolism of cancer: Glycolysis versus oxidative phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, K.C.; Wang, Q.; Bhaskar, P.T.; Miller, L.; Wang, Z.; Wheaton, W.; Chandel, N.; Laakso, M.; Muller, W.J.; Allen, E.L.; et al. Hexokinase 2 is required for tumor initiation and maintenance and its systemic deletion is therapeutic in mouse models of cancer. Cancer Cell. 2013, 24, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.-S.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.-J.; Mehta, H.; Hevener, A.L.; De Cabo, R.; et al. The Mitochondrial-Derived Peptide MOTS-c Promotes Metabolic Homeostasis and Reduces Obesity and Insulin Resistance. Cell Metab. 2015, 21, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Kim, K.H.; Cohen, P. MOTS-c: A novel mitochondrial-derived peptide regulating muscle and fat metabolism. Free. Radic. Biol. Med. 2016, 100, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Gan, L.; Liu, Z.; Liu, L.; Chang, J.-R.; Yin, D.-C.; Cao, H.-L.; Su, X.; Smith, W.W. Mitochondrial-Derived Peptide MOTS-c Attenuates Vascular Calcification and Secondary Myocardial Remodeling via Adenosine Monophosphate-Activated Protein Kinase Signaling Pathway. Cardiorenal Med. 2019, 10, 42–50. [Google Scholar] [CrossRef]
- Benayoun, B.A.; Lee, C. MOTS-c: A Mitochondrial-Encoded Regulator of the Nucleus. Bioessays 2019, 41, e1900046. [Google Scholar] [CrossRef]
- Lu, J.; Sharma, L.K.; Bai, Y. Implications of mitochondrial DNA mutations and mitochondrial dysfunction in tumorigenesis. Cell Res. 2009, 19, 802–815. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Lee, H.-C.; Wei, Y.-H. Mitochondrial DNA alterations and mitochondrial dysfunction in the progression of hepatocellular carcinoma. World J. Gastroenterol. 2013, 19, 8880–8886. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.M.; Costa, S.M.D.S.; do Amor Divino Miranda, P.M.; Ramos, P.Z.; Marconi, T.G.; Oliveira, G.S.; Castilho, A.M.; Sartorato, E.L. Analysis of mitochondrial alterations in Brazilian patients with sensorineural hearing loss using MALDI-TOF mass spectrometry. BMC Med Genet. 2016, 17, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wan, P.; Peng, T.; Xiao, K.; Su, M.; Shang, L.; Xu, B.; Su, Z.; Ye, X.; Peng, N.; et al. Associations between sequence variations in the mitochondrial DNA D-loop region and outcome of hepatocellular carcinoma. Oncol. Lett. 2016, 11, 3723–3728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Baez, S.; Pruyas, M.; Diaz, A.; Calvo, A.; Riquelme, E.; Wistuba, I. Mitochondrial DNA mutation at the D310 (displacement loop) mononucleotide sequence in the pathogenesis of gallbladder carcinoma. Clin. Cancer Res. 2004, 10, 1041–1046. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Lu, Y.; Zhang, Y.; Chen, Z.; Xing, G.; Cao, X. [Mutation analysis of mitochondrial 12S rRNA gene G709A in a maternally inherited pedigree with non-syndromic deafness]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2009, 26, 610–614. [Google Scholar]
- Zhu, Y.; Gu, X.; Xu, C. A Mitochondrial DNA A8701G Mutation Associated with Maternally Inherited Hypertension and Dilated Cardiomyopathy in a Chinese Pedigree of a Consanguineous Marriage. Chin. Med. J. 2016, 129, 259–266. [Google Scholar] [CrossRef]
- Lu, H.; Tang, S.; Xue, C.; Liu, Y.; Wang, J.; Zhang, W.; Luo, W.-J.; Chen, J.-Y. Mitochondrial-Derived Peptide MOTS-c Increases Adipose Thermogenic Activation to Promote Cold Adaptation. Int. J. Mol. Sci. 2019, 20, 2456. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Suo, C.; Li, S.-T.; Zhang, H.; Gao, P. Metabolic reprogramming for cancer cells and their microenvironment: Beyond the Warburg Effect. Biochim. Biophys. Acta Bioenerg. 2018, 1870, 51–66. [Google Scholar] [CrossRef]
- Hay, N. Reprogramming glucose metabolism in cancer: Can it be exploited for cancer therapy? Nat. Rev. Cancer 2016, 16, 635–649. [Google Scholar] [CrossRef] [Green Version]
- DeWaal, D.; Nogueira, V.; Terry, A.R.; Patra, K.C.; Jeon, S.M.; Guzman, G.; Au, J.; Long, C.P.; Antoniewicz, M.R.; Hay, N. Hexokinase-2 depletion inhibits glycolysis and induces oxidative phosphorylation in hepatocellular carcinoma and sensitizes to metformin. Nat Commun. 2018, 9, 446. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Wu, M.-H.; Huang, Y.-H.; Yeh, C.-T.; Cheng, M.-L.; Chi, H.-C.; Tsai, C.-Y.; Chung-Ying, T.; Chen, C.-Y.; Lin, K.-H. Taurine up-regulated gene 1 functions as a master regulator to coordinate glycolysis and metastasis in hepatocellular carcinoma. Hepatology 2018, 67, 188–203. [Google Scholar] [CrossRef] [PubMed]
- Shadel, G.S. Expression and maintenance of mitochondrial DNA: New insights into human disease pathology. Am. J. Pathol. 2008, 172, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-H.; Chu, Y.-D.; Lim, S.-N.; Chen, C.-W.; Yeh, C.-T.; Lin, W.-R. Impact of an MT-RNR1 Gene Polymorphism on Hepatocellular Carcinoma Progression and Clinical Characteristics. Int. J. Mol. Sci. 2021, 22, 1119. https://doi.org/10.3390/ijms22031119
Lin Y-H, Chu Y-D, Lim S-N, Chen C-W, Yeh C-T, Lin W-R. Impact of an MT-RNR1 Gene Polymorphism on Hepatocellular Carcinoma Progression and Clinical Characteristics. International Journal of Molecular Sciences. 2021; 22(3):1119. https://doi.org/10.3390/ijms22031119
Chicago/Turabian StyleLin, Yang-Hsiang, Yu-De Chu, Siew-Na Lim, Chun-Wei Chen, Chau-Ting Yeh, and Wey-Ran Lin. 2021. "Impact of an MT-RNR1 Gene Polymorphism on Hepatocellular Carcinoma Progression and Clinical Characteristics" International Journal of Molecular Sciences 22, no. 3: 1119. https://doi.org/10.3390/ijms22031119
APA StyleLin, Y. -H., Chu, Y. -D., Lim, S. -N., Chen, C. -W., Yeh, C. -T., & Lin, W. -R. (2021). Impact of an MT-RNR1 Gene Polymorphism on Hepatocellular Carcinoma Progression and Clinical Characteristics. International Journal of Molecular Sciences, 22(3), 1119. https://doi.org/10.3390/ijms22031119