The Role of PKM2 in Metabolic Reprogramming: Insights into the Regulatory Roles of Non-Coding RNAs


Abstract
:1. Introduction
2. PKM2: Uncovering the Origin
2.1. PKM2 Transcription and Dynamic Regulation
2.2. Impact of PKM2 Mutations on Gene Expression
2.3. Regulation of PKM2 Subcellular Distribution
3. “Metabolic” and “Non-Metabolic” Functions of PKM2
3.1. PKM2 Glycolytic Function
3.2. PKM2 Non-Glycolytic Functions
3.3. PKM2 and Apoptosis
3.4. PKM2 as an Inflammatory Regulator
3.5. PKM2 and Oncogenesis
4. Emerging Areas of Research Involving PKM2
4.1. Role of PKM2 beyond Cancer
4.2. PKM2 and Metabolic Homeostasis
4.3. Regulators of PKM2
4.3.1. PKM2 Activators
4.3.2. PKM2 Inhibitors
4.3.3. Natural Compounds
4.3.4. PKM2 microRNAs
microRNA Introduction
miRNAs in Cancer
miRNAs Regulation of PKM2 in Cancer
Regulation of PKM2 by miRNAs in Non-Cancerous Tissues
4.3.5. Long Non-Coding RNA Targeting of PKM2
Tumor-Suppressive PKM2/lncRNAs
Oncogenic PKM2/lncRNAs
PKM2/lncRNAs in Non-Cancer Diseases
4.3.6. CircularRNA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACL | ATP-citrate lyase |
| ACR | albumin to creatinine ratio |
| AIF | apoptosis-inducing factor |
| AIM2 | absent in melanoma 2 |
| AKI | acute kidney injury |
| AKR1A1 | aldo-keto reductase family 1, member A1 |
| ALDOA | aldolase A |
| AMPK | adenosine monophosphate-activated protein kinase |
| Apaf-1 | apoptotic protease activating factor-1 |
| APC/C-Cdh1 | anaphase-promoting complex/cyclosome-Cdh1 |
| APC | adenomatous polyposis coli |
| ATF6 | activating transcription factor 6 |
| ATG | autophagy related protein |
| ATP | adenosine triphosphate |
| BAD | blc-2 associated agonist of cell death |
| BAPN | β-aminopropionitrile fumarate |
| BAX | BCL2-associated X protein |
| BC | breast cancer |
| Bcl2 | b-cell lymphoma 2 |
| BCL-2 | b-cell lymphoma 2 |
| bFGF | basic fibroblast growth factor |
| BH3 | bcl-2 homology domain 3 |
| BIM | Bcl-2-like protein 11 |
| BMPR2 | bone morphogenetic protein receptor 2 |
| BUN | blood urea nitrogen |
| CAD | coronary artery disease |
| CARM | coactivator-associated arginine methyltransferase |
| Caspase | cysteine-aspartic acid protease |
| CCI | chronic constriction injury |
| ccRCC | clear-cell renal cell carcinoma |
| CD | Crohn’s disease |
| CD31 | cluster of differentiation 31 |
| CGNP | cerebellar granule neuron progenitor |
| CHOP | C/EBP homologous protein |
| CircRNA | circular RNA |
| COX | cyclooxygenase |
| CRC | colorectal cancer |
| DHAP | dihydroxyacetone phosphate |
| DN | diabetes nephropathy |
| DSS | dextrin sulfate sodium |
| EAE | autoimmune encephalomyelitis |
| eEF2K | eukaryotic elongation factor-2 |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor |
| EIF2AK2 | eukaryotic translation initiation factor 2 alpha kinase 2 |
| EIF2α | eukaryotic translation initiation factor 2 alpha |
| EMT | epithelial-mesenchymal transition |
| eNOS | endothelial NO synthase |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| ESRD | end-stage renal disease |
| FADD | fas-associated protein with death domain |
| EXP5 | exportin 5 |
| FBP | fructose 1,6-bisphosphate |
| FGF21 | fibroblast growth factor 21 |
| FIH-1 | asparaginyl hydroxylase factor inhibiting |
| GFR | glomerular filtration rate |
| GLUT1 | glucose transporter 1 |
| GM-CSF | granulocyte-macrophage colony-stimulating factor |
| GPR78 | G protein-coupled receptor |
| GSK3β | glycogen synthase kinase 3 beta |
| H2O2 | hydrogen peroxide |
| HDAC | histone deacetylase |
| HFD | high-fat diet |
| hGBM | human glioblastoma multiforme |
| HGC | human gastric cancer |
| HIF 1α | hypoxia-inducible factor 1-alpha |
| HK | hexokinase |
| HMGB | high mobility group box |
| hnRNPA | heterogenous nuclear ribonucleoprotein A |
| HSP | heat shock protein |
| IBD | inflammatory bowel disease |
| ICAM | intracellular adhesion molecule |
| IFNγ | interferon gamma |
| IGF | insulin growth factor |
| IL-1β | interleukin-1beta |
| IL-6 | interleukin-6 |
| IRE1α | inositol-requiring transmembrane kinase/endoribonuclease 1 alpha |
| JAK | Janus kinase |
| JNKs | c-Jun N-terminal kinases |
| LC3 | microtubule-associated protein 1A/1B-light chain 3 |
| LDHA | lactate dehydrogenase A |
| LMW-PTPs | low molecular weight protein tyrosine phosphatase |
| lncRNA | long non-coding RNA |
| LPS | lipopolysaccharides |
| MAPK | mitogen-activated protein kinase |
| MDM2 | mouse double minute 2 |
| MEF | mouse embryonic fibroblasts |
| miRNA | microRNA |
| MLC-2 | myosin regulatory light chain 2 |
| MS | multiple sclerosis |
| mTOR | mammalian target of rapamycin |
| NAC | n-acetyl-L-cysteine |
| NADH | nicotinamide adenine dinucleotide |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NF-κB | nuclear factor kappa light chain enhancer of activated B-cells |
| NK | natural killer cells |
| NLRC4 | NLR family CARD domain containing 4 |
| NLRP3 | NOD-, LRR- and pyrin domain-containing protein 3 |
| NLS | nuclear localization signal |
| NO | nitric oxide |
| NO | nitric oxide |
| OAA | oxaloacetate |
| NPC | nuclear pore complex |
| NSCLC | non-small-cell lung cancer |
| OGD | oxygen-glucose deprived |
| OXPHOS | oxidative phosphorylation |
| PARP | poly (ADP-ribose) polymerase |
| PDAC | pancreatic ductal adenocarcinoma |
| PDK | pyruvate dehydrogenase kinase |
| PD-L1 | programmed death-1 (PD-1) ligand 1 |
| PEP | phosphoenolpyruvate |
| PERK | protein kinase RNA-like endoplasmic reticulum kinase |
| PFK | phosphofructokinase |
| PHD | pyruvate dehydrogenase |
| PHD3 | prolyl hydroxylase 3 |
| PHGDH | phosphoglycerate dehydrogenase |
| PI3K | phosphoinositide 3-kinase |
| PIN1 | peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 |
| PiRNA | piwi-interacting RNA |
| PK | pyruvate kinase |
| PKC | protein kinase C |
| PKM | pyruvate kinase M |
| Pol II | polymerase II |
| PSAT1 | phosphoserine aminotransferase |
| PTB | polypyrimidine tract binding protein |
| PTEN | phosphatase and tensin homolog |
| PTP | protein-tyrosine phosphatase |
| PTP1B | protein tyrosine phosphatase 1B |
| RBM4 | RNA-binding motif |
| RISC | RNA-induced silencing complex |
| ROS | reactive oxygen species |
| rPKM2 | recombinant PKM2 |
| SCC | squamous cell carcinoma |
| SIR | systematic inflammatory responses |
| SIRT | sirtuins |
| SOCS3 | suppressor of cytokine signaling 3 |
| SNAP | synaptosome-associated protein |
| SNP | single nucleotide polymorphism |
| SOD2 | superoxide dismutase |
| SREBP | sterol regulatory element binding proteins |
| SRSF3 | serine/arginine-rich splicing factor |
| STAT | signal transducer and activator of transcription |
| T2D | type 2 diabetes |
| TGIF2 | TGFB-induced factor homeobox 2 |
| TAAD | thoracic aortic aneurysm and dissection |
| Th17 | T helper 17 |
| TLR | toll-like receptor |
| TNF-α | tumor necrosis factor alpha |
| TRIM35 | tripartite motif containing 35 |
| UC | ulcerative colitis |
| UCP1 | uncoupling protein 1 |
| VDAC | voltage-dependent anion channel |
| VEGF | vascular endothelial growth factor |
| Wnt | wingless-related integration site |
| WT1 | Wilms’ tumor 1 |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.A.; Owen, G.I. Glucose transporters: Expression, regulation and cancer. Biol. Res. 2002, 35, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, Y.D.; Babu, E.; Ramachandran, S.; Ganapathy, V. Amino Acid transporters in cancer and their relevance to “glutamine addiction”: Novel targets for the design of a new class of anticancer drugs. Cancer Res. 2015, 75, 1782–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Commisso, C.; Davidson, S.M.; Soydaner-Azeloglu, R.G.; Parker, S.J.; Kamphorst, J.J.; Hackett, S.; Grabocka, E.; Nofal, M.; Drebin, J.A.; Thompson, C.B.; et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 2013, 497, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cluntun, A.A.; Lukey, M.J.; Cerione, R.A.; Locasale, J.W. Glutamine Metabolism in Cancer: Understanding the Heterogeneity. Trends Cancer 2017, 3, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J. Energy metabolism of cancer: Glycolysis versus oxidative phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef] [Green Version]
- Marín-Hernández, A.; Gallardo-Pérez, J.C.; Ralph, S.J.; Rodríguez-Enríquez, S.; Moreno-Sánchez, R. HIF-1alpha modulates energy metabolism in cancer cells by inducing over-expression of specific glycolytic isoforms. Mini. Rev. Med. Chem. 2009, 9, 1084–1101. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.M.; Thomas, S.D.; Islam, A.; Muench, D.; Sedoris, K. c-Myc and cancer metabolism. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 5546–5553. [Google Scholar] [CrossRef] [Green Version]
- Magaway, C.; Kim, E.; Jacinto, E. Targeting mTOR and Metabolism in Cancer: Lessons and Innovations. Cells 2019, 8, 1584. [Google Scholar] [CrossRef] [Green Version]
- Ganapathy-Kanniappan, S.; Geschwind, J.F. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.S.; Manda, G. Metabolic Pathways of the Warburg Effect in Health and Disease: Perspectives of Choice, Chain or Chance. Int. J. Mol. Sci. 2017, 18, 2755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and glycine metabolism in cancer. Trends Biochem. Sci. 2014, 39, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Alquraishi, M.; Puckett, D.L.; Alani, D.S.; Humidat, A.S.; Frankel, V.D.; Donohoe, D.R.; Whelan, J.; Bettaieb, A. Pyruvate kinase M2: A simple molecule with complex functions. Free Radic. Biol. Med. 2019, 143, 176–192. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. THE METABOLISM OF TUMORS IN THE BODY. J. Gen. Physiol. 1927, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Qian, Y.; Wu, S. The Warburg Effect: Evolving Interpretations of an Established Concept. Free Radic. Biol. Med. 2015, 0, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem. Soc. Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, H.G. Observations on the carbohydrate metabolism of tumours. Biochem. J. 1929, 23, 536–545. [Google Scholar] [CrossRef]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, H.; Chun, Y.S.; Lewis, B.C.; Dang, C.V. A unique glucose-dependent apoptotic pathway induced by c-Myc. Proc. Natl. Acad. Sci. USA. 1998, 95, 1511–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafov, A.; Alessi, D.R. Uncoupling the Warburg effect from cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 19135–19136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, A.; Bolaños, J.P.; Moncada, S. E3 ubiquitin ligase APC/C-Cdh1 accounts for the Warburg effect by linking glycolysis to cell proliferation. Proc. Natl. Acad. Sci. 2010, 107, 738–741. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Keenan, H.A.; Li, Q.; Ishikado, A.; Kannt, A.; Sadowski, T.; Yorek, M.A.; Wu, I.H.; Lockhart, S.; Coppey, L.J.; et al. Pyruvate kinase M2 activation may protect against the progression of diabetic glomerular pathology and mitochondrial dysfunction. Nat. Med. 2017, 23, 753. [Google Scholar] [CrossRef] [PubMed]
- Bluemlein, K.; Grüning, N.-M.; Feichtinger, R.G.; Lehrach, H.; Kofler, B.; Ralser, M. No evidence for a shift in pyruvate kinase PKM1 to PKM2 expression during tumorigenesis. Oncotarget 2011, 2, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Semenza, G.L. Emerging roles of PKM2 in cell metabolism and cancer progression. Trends Endocrinol. Metab. 2012, 23, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, S.; Boschek, C.B.; Hugo, F.; Eigenbrodt, E. Pyruvate kinase type M2 and its role in tumor growth and spreading. Semin. Cancer Biol. 2005, 15, 300–308. [Google Scholar] [CrossRef]
- Wong, N.; De Melo, J.; Tang, D. PKM2, a Central Point of Regulation in Cancer Metabolism. Int. J. Cell Biol. 2013, 2013, 242513. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, S. Pyruvate kinase type M2: A key regulator of the metabolic budget system in tumor cells. Int. J. Biochem. Cell Biol. 2011, 43, 969–980. [Google Scholar] [CrossRef]
- Valentini, G.; Chiarelli, L.R.; Fortin, R.; Dolzan, M.; Galizzi, A.; Abraham, D.J.; Wang, C.; Bianchi, P.; Zanella, A.; Mattevi, A. Structure and function of human erythrocyte pyruvate kinase. Molecular basis of nonspherocytic hemolytic anemia. J. Biol. Chem. 2002, 277, 23807–23814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dombrauckas, J.D.; Santarsiero, B.D.; Mesecar, A.D. Structural basis for tumor pyruvate kinase M2 allosteric regulation and catalysis. Biochemistry 2005, 44, 9417–9429. [Google Scholar] [CrossRef]
- Israelsen, W.J.; Vander Heiden, M.G. Pyruvate kinase: Function, regulation and role in cancer. Semin. Cell Dev. Biol. 2015, 43, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Lang, N.; Wang, C.; Zhao, J.; Shi, F.; Wu, T.; Cao, H. Long non-coding RNA BCYRN1 promotes glycolysis and tumor progression by regulating the miR-149/PKM2 axis in non-small-cell lung cancer. Mol. Med. Rep. 2020, 21, 1509–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattevi, A.; Bolognesi, M.; Valentini, G. The allosteric regulation of pyruvate kinase. FEBS Lett. 1996, 389, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, J.M.; Dyson, R.D. Mammalian pyruvate kinase hybrid isozymes: Tissue distribution and physiological significance. J. Exp. Zool. 1978, 204, 361–367. [Google Scholar] [CrossRef]
- Zahra, K.; Dey, T.; Ashish; Mishra, S.P.; Pandey, U. Pyruvate Kinase M2 and Cancer: The Role of PKM2 in Promoting Tumorigenesis. Front. Oncol. 2020, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secrest, M.H.; Storm, M.; Carrington, C.; Casso, D.; Gilroy, K.; Pladson, L.; Boscoe, A.N. Prevalence of Pyruvate Kinase Deficiency: A Systematic Literature Review. Eur. J. Haematol. 2020, 105, 173–184. [Google Scholar] [CrossRef]
- Noguchi, T.; Inoue, H.; Tanaka, T. The M1- and M2-type isozymes of rat pyruvate kinase are produced from the same gene by alternative RNA splicing. J. Biol. Chem. 1986, 261, 13807–13812. [Google Scholar] [CrossRef]
- Izumi, S.; Manabe, A.; Tomoyasu, A.; Kihara-Negishi, F.; Ariga, H. Molecular cloning of the complementary DNA for the mouse pyruvate kinase M-2 gene whose expression is dependent upon cell differentiation. Biochim. Et. Biophys. Acta 1995, 1267, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Van Heyningen, V.; Bobrow, M.; Bodmer, W.F.; Gardiner, S.E.; Povey, S.; Hopkinson, D.A. Chromosome assignment of some human enzyme loci: Mitochondrial malate dehydrogenase to 7, mannosephosphate isomerase and pyruvate kinase to 15 and probably, esterase D to 13. Ann. Hum. Genet. 1975, 38, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Noguchi, T.; Sadahiro, S.; Hirai, H.; Yamada, K.; Matsuda, T.; Imai, E.; Tanaka, T. Isolation and characterization of the human pyruvate kinase M gene. Eur. J. Biochem. 1991, 198, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Clower, C.V.; Chatterjee, D.; Wang, Z.; Cantley, L.C.; Vander Heiden, M.G.; Krainer, A.R. The alternative splicing repressors hnRNP A1/A2 and PTB influence pyruvate kinase isoform expression and cell metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 1894–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, C.J.; Chen, M.; Assanah, M.; Canoll, P.; Manley, J.L. HnRNP proteins controlled by c-Myc deregulate pyruvate kinase mRNA splicing in cancer. Nature 2010, 463, 364–368. [Google Scholar] [CrossRef]
- Wang, Z.; Chatterjee, D.; Jeon, H.Y.; Akerman, M.; Vander Heiden, M.G.; Cantley, L.C.; Krainer, A.R. Exon-centric regulation of pyruvate kinase M alternative splicing via mutually exclusive exons. J. Mol. Cell Biol. 2012, 4, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Deng, X.; Liu, Y.; Liu, Y.; Sun, L.; Chen, F. PKM2, function and expression and regulation. Cell Biosci 2019, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Tani, K.; Fujii, H.; Tsutsumi, H.; Sukegawa, J.; Toyoshima, K.; Yoshida, M.C.; Noguchi, T.; Tanaka, T.; Miwa, S. Human liver type pyruvate kinase: cDNA cloning and chromosomal assignment. Biochem. Biophys. Res. Commun. 1987, 143, 431–438. [Google Scholar] [CrossRef]
- Tani, K.; Yoshida, M.C.; Satoh, H.; Mitamura, K.; Noguchi, T.; Tanaka, T.; Fujii, H.; Miwa, S. Human M2-type pyruvate kinase: cDNA cloning, chromosomal assignment and expression in hepatoma. Gene 1988, 73, 509–516. [Google Scholar]
- Imamura, K.; Tanaka, T. Multimolecular forms of pyruvate kinase from rat and other mammalian tissues. I. Electrophoretic studies. J. Biochem. 1972, 71, 1043–1051. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Mao, Q.; Xia, W.; Xu, Y.; Wang, J.; Xu, L.; Jiang, F. PKM2 and cancer: The function of PKM2 beyond glycolysis. Oncol. Lett. 2016, 11, 1980–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, H.P.; O’Reilly, F.J.; Wear, M.A.; O’Neill, J.R.; Fothergill-Gilmore, L.A.; Hupp, T.; Walkinshaw, M.D. M2 pyruvate kinase provides a mechanism for nutrient sensing and regulation of cell proliferation. Proc. Natl. Acad. Sci. USA 2013, 110, 5881–5886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Le, H. Dual roles of PKM2 in cancer metabolism. Acta Biochim. Biophys. Sin. 2013, 45, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macpherson, J.A.; Theisen, A.; Masino, L.; Fets, L.; Driscoll, P.C.; Encheva, V.; Snijders, A.P.; Martin, S.R.; Kleinjung, J.; Barran, P.E.; et al. Functional cross-talk between allosteric effects of activating and inhibiting ligands underlies PKM2 regulation. eLife 2019, 8, e45068. [Google Scholar] [CrossRef]
- Morita, M.; Sato, T.; Nomura, M.; Sakamoto, Y.; Inoue, Y.; Tanaka, R.; Ito, S.; Kurosawa, K.; Yamaguchi, K.; Sugiura, Y.; et al. PKM1 Confers Metabolic Advantages and Promotes Cell-Autonomous Tumor Cell Growth. Cancer Cell 2018, 33, 355–367.e7. [Google Scholar] [CrossRef] [Green Version]
- Ekman, P.; Dahlqvist, U.; Humble, E.; Engström, L. Comparative kinetic studies on the L-type pyruvate kinase from rat liver and the enzyme phosphorylated by cyclic 3’, 5’-AMP-stimulated protein kinase. Biochim. Biophys. Acta 1976, 429, 374–382. [Google Scholar] [CrossRef]
- Adachi, K.; Ghory, P.K.; Asakura, T.; Schwartz, E. A monomeric form of pyruvate kinase in human pyruvate kinase deficiency. Proc. Natl. Acad. Sci. USA 1977, 74, 501–504. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Taniguchi, N.; Noguchi, T. Dominant negative role of the glutamic acid residue conserved in the pyruvate kinase M(1) isozyme in the heterotropic allosteric effect involving fructose-1,6-bisphosphate. J. Biol. Chem. 2000, 275, 9150–9156. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Noguchi, T. Nutrient and hormonal regulation of pyruvate kinase gene expression. Biochem. J. 1999, 337 Pt 1, 1–11. [Google Scholar] [CrossRef]
- Brinck, U.; Fischer, G.; Eigenbrodt, E.; Oehmke, M.; Mazurek, S. L- and M2- pyruvate kinase expression in renal cell carcinomas and their metastases. Virchows Arch. 1994, 424, 177–185. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, X.; Ma, J.; Peng, H.; Wang, F.; Zha, X.; Wang, Y.; Jing, Y.; Yang, H.; Chen, R.; et al. Mammalian target of rapamycin up-regulation of pyruvate kinase isoenzyme type M2 is critical for aerobic glycolysis and tumor growth. Proc. Natl. Acad. Sci. USA 2011, 108, 4129–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Wang, H.; Yang, J.J.; Liu, X.; Liu, Z.-R. Pyruvate Kinase M2 Regulates Gene Transcription by Acting as a Protein Kinase. Mol. Cell 2012, 45, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.A.; Siddiqui, F.A.; Chaman, N.; Gupta, V.; Kumar, B.; Gopinath, P.; Bamezai, R.N. Missense mutations in pyruvate kinase M2 promote cancer metabolism, oxidative endurance, anchorage independence, and tumor growth in a dominant negative manner. J. Biol. Chem. 2014, 289, 8098–8105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anitha, M.; Kaur, G.; Baquer, N.Z.; Bamezai, R. Dominant negative effect of novel mutations in pyruvate kinase-M2. DNA Cell Biol. 2004, 23, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.J.; Wang, H.J.; Liu, J.S.; Cheng, H.H.; Hsu, S.C.; Wu, M.C.; Lu, C.H.; Wu, Y.F.; Wu, J.W.; Liu, Y.Y.; et al. Mutations in the PKM2 exon-10 region are associated with reduced allostery and increased nuclear translocation. Commun. Biol. 2019, 2, 105. [Google Scholar] [CrossRef]
- Fan, Z.; He, J.; Fu, T.; Zhang, W.; Yang, G.; Qu, X.; Liu, R.; Lv, L.; Wang, J. Arsenic trioxide inhibits EMT in hepatocellular carcinoma by promoting lncRNA MEG3 via PKM2. Biochem. Biophys. Res. Commun. 2019, 513, 834–840. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, Y.; Xia, Y.; Ji, H.; Chen, X.; Guo, F.; Lyssiotis, C.A.; Aldape, K.; Cantley, L.C.; Lu, Z. ERK1/2-dependent phosphorylation and nuclear translocation of PKM2 promotes the Warburg effect. Nat. Cell Biol. 2012, 14, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Lu, Z. Regulation and function of pyruvate kinase M2 in cancer. Cancer Lett. 2013, 339, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Hunter, T. Prolyl isomerase Pin1 in cancer. Cell Res. 2014, 24, 1033–1049. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.; Holyoak, T.; McDonald, G.; Gui, C.; Fenton, A.W. Differentiating a ligand’s chemical requirements for allosteric interactions from those for protein binding. Phenylalanine inhibition of pyruvate kinase. Biochemistry 2006, 45, 5421–5429. [Google Scholar] [CrossRef]
- Christofk, H.R.; Vander Heiden, M.G.; Harris, M.H.; Ramanathan, A.; Gerszten, R.E.; Wei, R.; Fleming, M.D.; Schreiber, S.L.; Cantley, L.C. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature 2008, 452, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; Locasale, J.W. Glucose Metabolism in Cancer: The Saga of Pyruvate Kinase Continues. Cancer Cell 2018, 33, 337–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaneton, B.; Hillmann, P.; Zheng, L.; Martin, A.C.L.; Maddocks, O.D.K.; Chokkathukalam, A.; Coyle, J.E.; Jankevics, A.; Holding, F.P.; Vousden, K.H.; et al. Serine is a natural ligand and allosteric activator of pyruvate kinase M2. Nature 2012, 491, 458–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, K.E.; Tan, I.S.; Lee, Y.S. SAICAR stimulates pyruvate kinase isoform M2 and promotes cancer cell survival in glucose-limited conditions. Science 2012, 338, 1069–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiou, D.; Poulogiannis, G.; Asara, J.M.; Boxer, M.B.; Jiang, J.K.; Shen, M.; Bellinger, G.; Sasaki, A.T.; Locasale, J.W.; Auld, D.S.; et al. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 2011, 334, 1278–1283. [Google Scholar] [CrossRef] [Green Version]
- Xiangyun, Y.; Xiaomin, N.; Linping, G.; Yunhua, X.; Ziming, L.; Yongfeng, Y.; Zhiwei, C.; Shun, L. Desuccinylation of pyruvate kinase M2 by SIRT5 contributes to antioxidant response and tumor growth. Oncotarget 2017, 8, 6984–6993. [Google Scholar] [CrossRef] [Green Version]
- Masaki, S.; Hashimoto, K.; Kihara, D.; Tsuzuki, C.; Kataoka, N.; Suzuki, K. The cysteine residue at 424th of pyruvate kinase M2 is crucial for tetramerization and responsiveness to oxidative stress. Biochem. Biophys. Res. Commun. 2020, 526, 973–977. [Google Scholar] [CrossRef]
- Sawicka, A.; Seiser, C. Histone H3 phosphorylation-a versatile chromatin modification for different occasions. Biochimie 2012, 94, 2193–2201. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Liu, L.Z.; Yin, Y.; He, J.; Li, Q.; Qian, X.; You, Y.; Lu, Z.; Peiper, S.C.; Shu, Y.; et al. Regulatory circuit of PKM2/NF-kappaB/miR-148a/152-modulated tumor angiogenesis and cancer progression. Oncogene 2015, 34, 5482–5493. [Google Scholar] [CrossRef]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2009, 29, 625. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Hu, H.; Chang, R.; Zhong, J.; Knabel, M.; O’Meally, R.; Cole, R.N.; Pandey, A.; Semenza, G.L. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell 2011, 145, 732–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoud, G.N.; Li, W. HIF-1alpha pathway: Role, regulation and intervention for cancer therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speer, R.E.; Karuppagounder, S.S.; Basso, M.; Sleiman, S.F.; Kumar, A.; Brand, D.; Smirnova, N.; Gazaryan, I.; Khim, S.J.; Ratan, R.R. Hypoxia-inducible factor prolyl hydroxylases as targets for neuroprotection by “antioxidant” metal chelators: From ferroptosis to stroke. Free Radic. Biol. Med. 2013, 62, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Ziello, J.E.; Jovin, I.S.; Huang, Y. Hypoxia-Inducible Factor (HIF)-1 regulatory pathway and its potential for therapeutic intervention in malignancy and ischemia. Yale J. Biol. Med. 2007, 80, 51–60. [Google Scholar] [PubMed]
- Lu, Z.; Hunter, T. Wnt-independent beta-catenin transactivation in tumor development. Cell Cycle (Georget. Tex.) 2004, 3, 571. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Li, Z.; Yang, P.; Zhang, L.; Fan, Y.; Li, Z. PKM2 depletion induces the compensation of glutaminolysis through [beta]-catenin/c-Myc pathway in tumor cells. Cell Signal. 2014, 26, 2397. [Google Scholar] [CrossRef]
- Yang, W.; Xia, Y.; Ji, H.; Zheng, Y.; Liang, J.; Huang, W.; Gao, X.; Aldape, K.; Lu, Z. Nuclear PKM2 regulates [beta]-catenin transactivation upon EGFR activation.(RESEARCH: LETTER)(epidermal growth factor receptor)(Report). Nature 2011, 480, 118. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Z.; Wang, H.; Chen, X.; Jin, C. Activation of AMPK by metformin promotes renal cancer cell proliferation under glucose deprivation through its interaction with PKM2. Int. J. Biol. Sci. 2019, 15, 617–627. [Google Scholar] [CrossRef]
- Zheng, Q.; Lin, Z.; Xu, J.; Lu, Y.; Meng, Q.; Wang, C.; Yang, Y.; Xin, X.; Li, X.; Pu, H.; et al. Long noncoding RNA MEG3 suppresses liver cancer cells growth through inhibiting beta-catenin by activating PKM2 and inactivating PTEN. Cell Death Dis. 2018, 9, 253. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, Q.; Feng, J.; Zheng, W.; Du, J.; Meng, X.; Wang, Y.; Wang, J. Activation of AMPK promotes thyroid cancer cell migration through its interaction with PKM2 and beta-catenin. Life Sci. 2019, 239, 116877. [Google Scholar] [CrossRef]
- Ji, L.; Ruixiu, C.; Xiongjun, W.; Yajuan, Z.; Pan, W.; Hong, G.; Chen, L.; Fan, Y.; Rong, Z.; Ping, W.; et al. Mitochondrial PKM2 regulates oxidative stress-induced apoptosis by stabilizing Bcl2. Cell Res. 2016, 27, 329–351. [Google Scholar]
- Qi, H.; Ning, X.; Yu, C.; Ji, X.; Jin, Y.; McNutt, M.A.; Yin, Y. Succinylation-dependent mitochondrial translocation of PKM2 promotes cell survival in response to nutritional stress. Cell Death Dis. 2019, 10, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Wang, D.; Jin, F.; Bian, Z.; Li, L.; Liang, H.; Li, M.; Shi, L.; Pan, C.; Zhu, D.; et al. Pyruvate kinase type M2 promotes tumour cell exosome release via phosphorylating synaptosome-associated protein 23. Nat. Commun. 2017, 8, 14041. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.-C.; Hung, W.-C. Pyruvate kinase M2 fuels multiple aspects of cancer cells: From cellular metabolism, transcriptional regulation to extracellular signaling. Mol. Cancer 2018, 17, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, V.R.; Dias, M.S.; Hainaut, P. Tumor-cell-derived microvesicles as carriers of molecular information in cancer. Curr. Opin. Oncol. 2013, 25, 66–75. [Google Scholar] [CrossRef]
- Tamada, M.; Suematsu, M.; Saya, H. Pyruvate Kinase M2: Multiple Faces for Conferring Benefits on Cancer Cells. Clin. Cancer Res. 2012, 18, 5554. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, S.; Boschek, C.; Eigenbrodt, E. The Role of Phosphometabolites in Cell Proliferation, Energy Metabolism, and Tumor Therapy. J. Bioenerg. Biomembr. 1997, 29, 315–330. [Google Scholar] [CrossRef]
- Prigione, A.; Rohwer, N.; Hoffmann, S.; Mlody, B.; Drews, K.; Bukowiecki, R.; Blumlein, K.; Wanker, E.E.; Ralser, M.; Cramer, T.; et al. HIF1[alpha] Modulates Cell Fate Reprogramming Through Early Glycolytic Shift and Upregulation of PDK1-3 and PKM2. Stem Cells 2014, 32, 364. [Google Scholar] [CrossRef] [Green Version]
- Salani, B.; Ravera, S.; Amaro, A.; Salis, A.; Passalacqua, M.; Millo, E.; Damonte, G.; Marini, C.; Pfeffer, U.; Sambuceti, G.; et al. IGF1 regulates PKM2 function through Akt phosphorylation. Cell Cycle (Georget. Tex) 2015, 14, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G. HIF-1 mediates the Warburg effect in clear cell renal carcinoma. J. Bioenerg. Biomembr. 2007, 39, 231–234. [Google Scholar] [CrossRef]
- Saleme, B.; Gurtu, V.; Zhang, Y.; Kinnaird, A.; Boukouris, A.E.; Gopal, K.; Ussher, J.R.; Sutendra, G. Tissue-specific regulation of p53 by PKM2 is redox dependent and provides a therapeutic target for anthracycline-induced cardiotoxicity. Sci. Transl. Med. 2019, 11, eaau8866. [Google Scholar] [CrossRef] [PubMed]
- Brimacombe, K.R.; Anastasiou, D.; Hong, B.S.; Tempel, W.; Dimov, S.; Veith, H.; Auld, D.S.; Vander Heiden, M.G.; Thomas, C.J.; Park, H.W.; et al. ML285 affects reactive oxygen species’ inhibition of pyruvate kinase M2. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2010. [Google Scholar]
- Zhu, X.; Huang, L.; Gong, J.; Shi, C.; Wang, Z.; Ye, B.; Xuan, A.; He, X.; Long, D.; Zhu, X.; et al. NF-kappaB pathway link with ER stress-induced autophagy and apoptosis in cervical tumor cells. Cell Death Discov. 2017, 3, 17059. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.J.; Neoh, K. The Stat family of transcription factors have diverse roles in mammary gland development. Semin. Cell Dev. Biol. 2008, 19, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.P.; Luo, L.J.; Chen, H.Z.; Chen, Q.T.; Bian, X.L.; Wu, S.F.; Zhou, J.X.; Zhao, W.X.; Liu, J.M.; Wang, X.M.; et al. Ectosomal PKM2 Promotes HCC by Inducing Macrophage Differentiation and Remodeling the Tumor Microenvironment. Mol. Cell 2020, 78, 1192–1206.e10. [Google Scholar] [CrossRef]
- Hosios, A.M.; Fiske, B.P.; Gui, D.Y.; Vander Heiden, M.G. Lack of Evidence for PKM2 Protein Kinase Activity. Mol. Cell 2015, 59, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yu, Z.; Li, J.; Zhang, A.; Zhang, X.; Kan, Q. Impact of PKM2 gene silencing on biological behavior of HepG2 cells. Int J. Clin. Exp. Med. 2016, 7, 13475–13483. [Google Scholar]
- Dong, D.; Dong, Y.; Fu, J.; Lu, S.; Yuan, C.; Xia, M.; Sun, L. Bcl2 inhibitor ABT737 reverses the Warburg effect via the Sirt3-HIF1α axis to promote oxidative stress-induced apoptosis in ovarian cancer cells. Life Sci. 2020, 255, 117846. [Google Scholar] [CrossRef]
- Kwon, O.-H.; Kang, T.-W.; Kim, J.-H.; Kim, M.; Noh, S.-M.; Song, K.-S.; Yoo, H.-S.; Kim, W.-H.; Xie, Z.; Pocalyko, D.; et al. Pyruvate kinase M2 promotes the growth of gastric cancer cells via regulation of Bcl-xL expression at transcriptional level. Biochem. Biophys. Res. Commun. 2012, 423, 38–44. [Google Scholar] [CrossRef]
- Hu, W.; Lu, S.X.; Li, M.; Zhang, C.; Liu, L.L.; Fu, J.; Jin, J.T.; Luo, R.Z.; Zhang, C.Z.; Yun, J.P. Pyruvate kinase M2 prevents apoptosis via modulating Bim stability and associates with poor outcome in hepatocellular carcinoma. Oncotarget 2015, 6, 6570–6583. [Google Scholar] [CrossRef] [PubMed]
- Chipuk, J.E.; Bouchier-Hayes, L.; Green, D.R. Mitochondrial outer membrane permeabilization during apoptosis: The innocent bystander scenario. Cell Death Differ. 2006, 13, 1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Saxena, S.; Singh, B.K.; Kakkar, P. BH3-only protein BIM: An emerging target in chemotherapy. Eur J. Cell Biol 2017, 96, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Tait, S.W. Mitochondrial apoptosis: Killing cancer using the enemy within. Br. J. Cancer 2015, 112, 957–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Yang, P.; Hu, W.; Wang, Y.; Lu, Y.; Zhang, L.; Fan, Y.; Xiao, H.; Li, Z. Overexpression of PKM2 promotes mitochondrial fusion through attenuated p53 stability. Oncotarget 2016, 7, 78069–78082. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Wang, X.R.; Wang, X.L.; Liu, S.H.; Ding, X.W.; Chen, G.Q.; Lu, Y. A Novel Role for Pyruvate Kinase M2 as a Corepressor for P53 during the DNA Damage Response in Human Tumor Cells. J. Biol. Chem. 2016, 291, 26138–26150. [Google Scholar] [CrossRef] [Green Version]
- Meng, M.-B.; Wang, H.-H.; Guo, W.-H.; Wu, Z.-Q.; Zeng, X.-L.; Zaorsky, N.G.; Shi, H.-S.; Qian, D.; Niu, Z.-M.; Jiang, B.; et al. Targeting pyruvate kinase M2 contributes to radiosensitivity of non-small cell lung cancer cells in vitro and in vivo. Cancer Lett. 2015, 356, 985–993. [Google Scholar] [CrossRef]
- Xu, Q.; Tu, J.; Dou, C.; Zhang, J.; Yang, L.; Liu, X.; Lei, K.; Liu, Z.; Wang, Y.; Li, L.; et al. HSP90 promotes cell glycolysis, proliferation and inhibits apoptosis by regulating PKM2 abundance via Thr-328 phosphorylation in hepatocellular carcinoma (Report). Mol. Cancer 2017, 16, 178. [Google Scholar] [CrossRef]
- Wubben, T.J.; Pawar, M.; Weh, E.; Smith, A.; Sajjakulnukit, P.; Zhang, L.; Dai, L.; Hager, H.; Pai, M.P.; Lyssiotis, C.A.; et al. Small molecule activation of metabolic enzyme pyruvate kinase muscle isozyme 2, PKM2, circumvents photoreceptor apoptosis. Sci. Rep. 2020, 10, 2990. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Han, L.; Jian, Y.; Ma, Y.; Yan, W.; Chen, X.; Xu, H.; Li, L. Resveratrol induces apoptosis in human melanoma cell through negatively regulating Erk/PKM2/Bcl-2 axis. Oncotargets Ther. 2018, 11, 8995. [Google Scholar] [CrossRef] [Green Version]
- Paraiso, K.H.; Haarberg, H.E.; Wood, E.; Rebecca, V.W.; Chen, Y.A.; Xiang, Y.; Ribas, A.; Lo, R.S.; Weber, J.S.; Sondak, V.K.; et al. The HSP90 inhibitor XL888 overcomes BRAF inhibitor resistance mediated through diverse mechanisms. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 2502–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Xu, J.; Liu, X.; Xia, X.; Li, N.; Bi, X. Shikonin induces apoptosis in the human gastric cancer cells HGC-27 through mitochondria-mediated pathway. Pharmacogn. Mag. 2015, 11, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomonosova, E.; Chinnadurai, G. BH3-only proteins in apoptosis and beyond: An overview. Oncogene 2008, 27, S2–S19. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Nakagawara, A. Role of p53 in Cell Death and Human Cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves-Filho, J.C.; Palsson-McDermott, E.M. Pyruvate Kinase M2: A Potential Target for Regulating Inflammation. Front. Immunol. 2016, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palsson-McDermott, E.M.; Curtis, A.M.; Goel, G.; Lauterbach, M.A.; Sheedy, F.J.; Gleeson, L.E.; van den Bosch, M.W.; Quinn, S.R.; Domingo-Fernandez, R.; Johnston, D.G.; et al. Pyruvate kinase M2 regulates Hif-1alpha activity and IL-1beta induction and is a critical determinant of the warburg effect in LPS-activated macrophages. Cell Metab. 2015, 21, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Li, Z.; Li, H.; Lu, Y.; Wu, H.; Li, Z. Pyruvate kinase M2 accelerates pro-inflammatory cytokine secretion and cell proliferation induced by lipopolysaccharide in colorectal cancer. Cell Signal. 2015, 27, 1525–1532. [Google Scholar] [CrossRef]
- Shirai, T.; Nazarewicz, R.R.; Wallis, B.B.; Yanes, R.E.; Watanabe, R.; Hilhorst, M.; Tian, L.; Harrison, D.G.; Giacomini, J.C.; Assimes, T.L.; et al. The glycolytic enzyme PKM2 bridges metabolic and inflammatory dysfunction in coronary artery disease. J. Exp. Med. 2016, 213, 337–354. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Jiang, C.; Wang, X.; Geng, M.; Peng, Y.; Guo, Y.; Wang, S.; Li, X.; Tao, P.; Zhang, F.; et al. Upregulated PKM2 in Macrophages Exacerbates Experimental Arthritis via STAT1 Signaling. J. Immunol. 2020, 205, 181–192. [Google Scholar] [CrossRef]
- Andersson, U.; Wang, H.; Palmblad, K.; Aveberger, A.C.; Bloom, O.; Erlandsson-Harris, H.; Janson, A.; Kokkola, R.; Zhang, M.; Yang, H.; et al. High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J. Exp. Med. 2000, 192, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xie, M.; Yang, M.; Yu, Y.; Zhu, S.; Hou, W.; Kang, R.; Lotze, M.T.; Billiar, T.R.; Wang, H.; et al. PKM2 regulates the Warburg effect and promotes HMGB1 release in sepsis. Nat. Commun. 2014, 5, 4436. [Google Scholar] [CrossRef] [PubMed]
- Magna, M.; Pisetsky, D.S. The role of HMGB1 in the pathogenesis of inflammatory and autoimmune diseases. Mol. Med. 2014, 20, 138–146. [Google Scholar] [CrossRef]
- Xie, M.; Yu, Y.; Kang, R.; Zhu, S.; Yang, L.; Zeng, L.; Sun, X.; Yang, M.; Billiar, T.R.; Wang, H.; et al. PKM2-dependent glycolysis promotes NLRP3 and AIM2 inflammasome activation. Nat. Commun. 2016, 7, 13280. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Xie, J.; Wu, Y.; Wang, G.; Qi, X.; Liu, Z.; Wang, Y.; Wang, X.; Hoque, A.; Oakhill, J.; et al. The eEF2 kinase-induced STAT3 inactivation inhibits lung cancer cell proliferation by phosphorylation of PKM2. Cell Commun. Signal. 2020, 18, 25. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Proud, C.G. Eukaryotic elongation factor 2 kinase as a drug target in cancer, and in cardiovascular and neurodegenerative diseases. Acta Pharmacol. Sin. 2016, 37, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Kasembeli, M.M.; Bharadwaj, U.; Robinson, P.; Tweardy, D.J. Contribution of STAT3 to Inflammatory and Fibrotic Diseases and Prospects for its Targeting for Treatment. Int. J. Mol. Sci. 2018, 19, 2299. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Tang, L.; Yang, X.; Chen, R.; Zhang, Z.; Leng, Y.; Chen, A.F. Gene Regulatory Effect of Pyruvate Kinase M2 is Involved in Renal Inflammation in Type 2 Diabetic Nephropathy. Exp. Clin. Endocrinol. Diabetes 2020, 128, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, C.; Aboushousha, R.; Manuel, A.M.; Chia, S.B.; Erickson, C.; MacPherson, M.B.; van der Velden, J.L.; Anathy, V.; Dixon, A.E.; Irvin, C.G.; et al. Pyruvate Kinase M2 Promotes Expression of Proinflammatory Mediators in House Dust Mite-Induced Allergic Airways Disease. J. Immunol 2020, 204, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Z.; Wang, F.; Liu, S.; Li, H.; Wang, Y. Ablation of PKM2 ameliorated ER stress-induced apoptosis and associated inflammation response in IL-1β-treated chondrocytes via blocking Rspo2-mediated Wnt/β-catenin signaling. J. Cell. Biochem. 2020, 121, 4204–4213. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.L.; Hou, G.G.; Liu, J.; Ru, T.; Xu, Y.Z.; Zhao, S.Y.; Ye, H.; Zhang, L.Y.; Chen, K.X.; Guo, Y.W.; et al. Synthesis and Target Identification of Benzoxepane Derivatives as Potential Anti-Neuroinflammatory Agents for Ischemic Stroke. Angew. Chem. Int. Ed. 2020, 59, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Tae, I.H.; Son, J.Y.; Lee, S.H.; Ahn, M.Y.; Yoon, K.; Yoon, S.; Moon, H.R.; Kim, H.S. A new SIRT1 inhibitor, MHY2245, induces autophagy and inhibits energy metabolism via PKM2/mTOR pathway in human ovarian cancer cells. Int. J. Biol. Sci. 2020, 16, 1901–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.; Bhagat, S.D.; Gupta, A.; Samaiya, A.; Srivastava, A.; Shukla, S. Dietary-phytochemical mediated reversion of cancer-specific splicing inhibits Warburg effect in head and neck cancer. BMC Cancer 2019, 19, 1031. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.; Ding, M.; Wang, B.; Lu, Z.; Zhao, Q.; Shaw, K.; Yung, W.K.; Weinstein, J.N.; Tan, M.; Yao, J. Tissue-specific isoform switch and DNA hypomethylation of the pyruvate kinase PKM gene in human cancers. Oncotarget 2014, 5, 8202–8210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Zhang, J.; Manley, J.L. Turning on a fuel switch of cancer: hnRNP proteins regulate alternative splicing of pyruvate kinase mRNA. Cancer Res. 2010, 70, 8977–8980. [Google Scholar] [CrossRef] [Green Version]
- Zhan, C.; Yan, L.; Wang, L.; Ma, J.; Jiang, W.; Zhang, Y.; Shi, Y.; Wang, Q. Isoform switch of pyruvate kinase M1 indeed occurs but not to pyruvate kinase M2 in human tumorigenesis. PLoS ONE 2015, 10, e0118663. [Google Scholar] [CrossRef]
- Israelsen, W.J.; Dayton, T.L.; Davidson, S.M.; Fiske, B.P.; Hosios, A.M.; Bellinger, G.; Li, J.; Yu, Y.; Sasaki, M.; Horner, J.W.; et al. PKM2 Isoform-Specific Deletion Reveals a Differential Requirement for Pyruvate Kinase in Tumor Cells. Cell 2013, 155, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Jiang, Z.; Wang, B.; Wang, Y.; Hu, X. Vitamin K3 and K5 are inhibitors of tumor pyruvate kinase M2. Cancer Lett. 2012, 316, 204–210. [Google Scholar] [CrossRef]
- Li, N.; Feng, L.; Liu, H.; Wang, J.; Kasembeli, M.; Tran, M.K.; Tweardy, D.J.; Lin, S.H.; Chen, J. PARP Inhibition Suppresses Growth of EGFR-Mutant Cancers by Targeting Nuclear PKM2. Cell Rep. 2016, 15, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Asai, Y.; Yamada, K.; Watanabe, T.; Keng, V.W.; Noguchi, T. Insulin Stimulates Expression of the Pyruvate Kinase M Gene in 3T3-L1 Adipocytes. Biosci. Biotechnol. Biochem. 2003, 67, 1272–1277. [Google Scholar] [CrossRef]
- Tracey, B. What to do with PKM2. Sci. Bus. Exch. 2013, 6, 28. [Google Scholar]
- Basse, A.L.; Isidor, M.S.; Winther, S.; Skjoldborg, N.B.; Murholm, M.; Andersen, E.S.; Pedersen, S.B.; Wolfrum, C.; Quistorff, B.; Hansen, J.B. Regulation of glycolysis in brown adipocytes by HIF-1α. Sci. Rep. 2017, 7, 4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettaieb, A.; Bakke, J.; Nagata, N.; Matsuo, K.; Xi, Y.; Liu, S.; AbouBechara, D.; Melhem, R.; Stanhope, K.; Cummings, B.; et al. Protein Tyrosine Phosphatase 1B Regulates Pyruvate Kinase M2 Tyrosine Phosphorylation. J. Biol. Chem. 2013, 288, 17360–17371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.J.; Park, Y.S.; Kang, M.G.; You, Y.-M.; Jung, Y.; Koo, H.; Kim, J.-A.; Kim, M.-J.; Hong, S.-M.; Lee, K.B.; et al. Pyruvate kinase isoenzyme M2 is a therapeutic target of gemcitabine-resistant pancreatic cancer cells. Exp. Cell Res. 2015, 336, 119–129. [Google Scholar] [CrossRef]
- Nakatsu, D.; Horiuchi, Y.; Kano, F.; Noguchi, Y.; Sugawara, T.; Takamoto, I.; Kubota, N.; Kadowaki, T.; Murata, M. l-cysteine reversibly inhibits glucose-induced biphasic insulin secretion and ATP production by inactivating PKM2. Proc. Natl. Acad. Sci. USA 2015, 112, E1067–E1076. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yang, Z.; Gao, Y.; Li, Q.; Su, Y.; Wang, Y.; Zhang, Y.; Man, H.; Liu, H. Pyruvate kinase, muscle isoform 2 promotes proliferation and insulin secretion of pancreatic β-cells via activating Wnt/CTNNB1 signaling. Int. J. Clin. Exp. Pathol. 2015, 8, 14441–14448. [Google Scholar]
- Seino, S.; Shibasaki, T.; Minami, K. Dynamics of insulin secretion and the clinical implications for obesity and diabetes. J. Clin. Investig. 2011, 121, 2118. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, A.; Stefan, N.; Hardt, E.; Haring, H.; Stumvoll, M. Characterisation of beta-cell dysfunction of impaired glucose tolerance: Evidence for impairment of incretin-induced insulin secretion. Diabetologia 2000, 43, 852–858. [Google Scholar] [CrossRef] [Green Version]
- Brunzell, J.D.; Robertson, R.P.; Lerner, R.L.; Hazzard, W.R.; Ensinck, J.W.; Bierman, E.L.; Porte, D., Jr. Relationships between fasting plasma glucose levels and insulin secretion during intravenous glucose tolerance tests. J. Clin. Endocrinol. Metab. 1976, 42, 222–229. [Google Scholar] [CrossRef]
- Prentki, M.; Matschinsky, F.M. Ca2+, cAMP, and phospholipid-derived messengers in coupling mechanisms of insulin secretion. Physiol. Rev. 1987, 67, 1185–1248. [Google Scholar] [CrossRef]
- Wollheim, C.B.; Sharp, G.W. Regulation of insulin release by calcium. Physiol. Rev. 1981, 61, 914–973. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.M. ATP-sensitive potassium channelopathies: Focus on insulin secretion. J. Clin. Investig. 2005, 115, 2047–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henquin, J.C. Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 2000, 49, 1751–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, R.P. Beta-cell deterioration during diabetes: What’s in the gun? Trends Endocrinol. Metab. 2009, 20, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillis, A.L.; Lau, A.N.; Devoe, C.X.; Dayton, T.L.; Danai, L.V.; Di Vizio, D.; Vander Heiden, M.G. PKM2 is not required for pancreatic ductal adenocarcinoma. Cancer Metab. 2018, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, A.S. The epidemic of obesity and diabetes: Trends and treatments. Tex Heart Inst. J. 2011, 38, 142–144. [Google Scholar] [PubMed]
- Elshorbagy, A.K.; Smith, A.D.; Kozich, V.; Refsum, H. Cysteine and obesity. Obes. (Silver SpringMd) 2012, 20, 473–481. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Isidor, M.S.; Winther, S.; Markussen, L.K.; Basse, A.L.; Quistorff, B.; Nedergaard, J.; Emanuelli, B.; Hansen, J.B. Pyruvate kinase M2 represses thermogenic gene expression in brown adipocytes. FEBS Lett. 2019, 594, 1218–1225. [Google Scholar] [CrossRef]
- Zhou, H.L.; Zhang, R.; Anand, P.; Stomberski, C.T.; Qian, Z.; Hausladen, A.; Wang, L.; Rhee, E.P.; Parikh, S.M.; Karumanchi, S.A.; et al. Metabolic reprogramming by the S-nitroso-CoA reductase system protects against kidney injury. Nature 2019, 565, 96–100. [Google Scholar] [CrossRef]
- Chen, J.; Xie, J.; Jiang, Z.; Wang, B.; Wang, Y.; Hu, X. Shikonin and its analogs inhibit cancer cell glycolysis by targeting tumor pyruvate kinase-M2. Oncogene 2011, 30, 4297–4306. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.A.; Bamezai, R.N.K. Resveratrol Inhibits Cancer Cell Metabolism by Down Regulating Pyruvate Kinase M2 via Inhibition of Mammalian Target of Rapamycin (Resveratrol PKM2 and Cancer Metabolism). PLoS ONE 2012, 7, e36764. [Google Scholar] [CrossRef]
- Anastasiou, D.; Yu, Y.; Israelsen, W.J.; Jiang, J.K.; Boxer, M.B.; Hong, B.S.; Tempel, W.; Dimov, S.; Shen, M.; Jha, A.; et al. Pyruvate kinase M2 activators promote tetramer formation and suppress tumorigenesis. Nat. Chem. Biol. 2012, 8, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Walsh, M.J.; Brimacombe, K.R.; Anastasiou, D.; Yu, Y.; Israelsen, W.J.; Hong, B.S.; Tempel, W.; Dimov, S.; Veith, H.; et al. ML265: A potent PKM2 activator induces tetramerization and reduces tumor formation and size in a mouse xenograft model. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2010. [Google Scholar]
- Ren, R.; Guo, J.; Shi, J.; Tian, Y.; Li, M.; Kang, H. PKM2 regulates angiogenesis of VR-EPCs through modulating glycolysis, mitochondrial fission, and fusion. J. Cell. Physiol. 2020, 235, 6204–6217. [Google Scholar] [CrossRef]
- Li, R.Z.; Fan, X.X.; Shi, D.F.; Zhu, G.Y.; Wang, Y.W.; Luo, L.X.; Pan, H.D.; Yao, X.J.; Leung, E.L.H.; Liu, L. Identification of a new pyruvate kinase M2 isoform (PKM2) activator for the treatment of non-small-cell lung cancer (NSCLC). Chem. Biol. Drug Des. 2018, 92, 1851–1858. [Google Scholar] [CrossRef]
- Guo, J.; Ren, R.; Yao, X.; Ye, Y.; Sun, K.; Lin, J.; Wang, G.; Guo, F.; Xiao, J.; Xu, T. PKM2 suppresses osteogenesis and facilitates adipogenesis by regulating beta-catenin signaling and mitochondrial fusion and fission. Aging (Albany NY) 2020, 12, 3976–3992. [Google Scholar] [CrossRef]
- Palsson-McDermott, E.M.; Dyck, L.; Zaslona, Z.; Menon, D.; McGettrick, A.F.; Mills, K.H.G.; O’Neill, L.A. Pyruvate Kinase M2 Is Required for the Expression of the Immune Checkpoint PD-L1 in Immune Cells and Tumors. Front. Immunol 2017, 8, 1300. [Google Scholar] [CrossRef] [Green Version]
- Kythreotou, A.; Siddique, A.; Mauri, F.A.; Bower, M.; Pinato, D.J. Pd-L1. J. Clin. Pathol. 2018, 71, 189–194. [Google Scholar] [CrossRef]
- Angiari, S.; Runtsch, M.C.; Sutton, C.E.; Palsson-McDermott, E.M.; Kelly, B.; Rana, N.; Kane, H.; Papadopoulou, G.; Pearce, E.L.; Mills, K.H.G.; et al. Pharmacological Activation of Pyruvate Kinase M2 Inhibits CD4(+) T Cell Pathogenicity and Suppresses Autoimmunity. Cell Metab. 2020, 31, 391–405.e8. [Google Scholar] [CrossRef] [Green Version]
- Seki, S.M.; Posyniak, K.; McCloud, R.; Rosen, D.A.; Fernandez-Castaneda, A.; Beiter, R.M.; Serbulea, V.; Nanziri, S.C.; Hayes, N.; Spivey, C.; et al. Modulation of PKM activity affects the differentiation of TH17 cells. Sci. Signal. 2020, 13, eaay9217. [Google Scholar] [CrossRef]
- Walls, J.F.; Subleski, J.J.; Palmieri, E.M.; Gonzalez-Cotto, M.; Gardiner, C.M.; McVicar, D.W.; Finlay, D.K. Metabolic but not transcriptional regulation by PKM2 is important for natural killer cell responses. eLife 2020, 9, e59166. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.H.; Yang, Y.X.; Meng, X.; Luo, X.Y.; Li, X.M.; Shuai, Z.W.; Ye, D.Q.; Pan, H.F. NLRP3: A promising therapeutic target for autoimmune diseases. Autoimmun. Rev. 2018, 17, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Leng, K.; Liu, Y.; Sun, H.; Gao, J.; Ren, Q.; Zhou, T.; Dong, J.; Xia, J. The impact of hyperglycaemia on PKM2-mediated NLRP3 inflammasome/stress granule signalling in macrophages and its correlation with plaque vulnerability: An in vivo and in vitro study. Metab. Clin. Exp. 2020, 107, 154231. [Google Scholar] [CrossRef] [PubMed]
- Le, S.; Zhang, H.; Huang, X.; Chen, S.; Wu, J.; Chen, S.; Ding, X.; Chen, S.; Zhao, J.; Xu, H.; et al. PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1beta Secretion. J. Cardiovasc. Pharm. 2020, 25, 364–376. [Google Scholar] [CrossRef]
- Wang, F.; Wang, K.; Xu, W.; Zhao, S.; Ye, D.; Wang, Y.; Xu, Y.; Zhou, L.; Chu, Y.; Zhang, C.; et al. SIRT5 Desuccinylates and Activates Pyruvate Kinase M2 to Block Macrophage IL-1beta Production and to Prevent DSS-Induced Colitis in Mice. Cell Rep. 2017, 19, 2331–2344. [Google Scholar] [CrossRef] [Green Version]
- Ning, X.; Qi, H.; Li, R.; Li, Y.; Jin, Y.; McNutt, M.A.; Liu, J.; Yin, Y. Discovery of novel naphthoquinone derivatives as inhibitors of the tumor cell specific M2 isoform of pyruvate kinase. Eur. J. Med. Chem. 2017, 138, 343–352. [Google Scholar] [CrossRef]
- Ning, X.; Qi, H.; Li, R.; Jin, Y.; McNutt, M.A.; Yin, Y. Synthesis and antitumor activity of novel 2, 3-didithiocarbamate substituted naphthoquinones as inhibitors of pyruvate kinase M2 isoform. J. Enzym. Inhib. Med. Chem. 2018, 33, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Ha, J.H.; Jang, J.; Chung, S.I.; Yoon, Y. AMPK and SREBP-1c mediate the anti-adipogenic effect of beta-hydroxyisovalerylshikonin. Int. J. Mol. Med. 2016, 37, 816–824. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.J.; Jung, C.H.; Ahn, J.; Gwon, S.Y.; Ha, T.Y. Shikonin inhibits adipogenic differentiation via regulation of mir-34a-FKBP1B. Biochem. Biophys. Res. Commun. 2015, 467, 941–947. [Google Scholar] [CrossRef]
- Gwon, S.Y.; Ahn, J.Y.; Chung, C.H.; Moon, B.; Ha, T.Y. Lithospermum erythrorhizon suppresses high-fat diet-induced obesity, and acetylshikonin, a main compound of Lithospermum erythrorhizon, inhibits adipocyte differentiation. J. Agric. Food Chem. 2012, 60, 9089–9096. [Google Scholar] [CrossRef]
- Lee, H.; Bae, S.; Kim, K.; Kim, W.; Chung, S.I.; Yang, Y.; Yoon, Y. Shikonin inhibits adipogenesis by modulation of the WNT/beta-catenin pathway. Life Sci. 2011, 88, 294–301. [Google Scholar] [CrossRef]
- Lee, H.; Kang, R.; Yoon, Y. Shikonin inhibits fat accumulation in 3T3-L1 adipocytes. Phytother. Res. Ptr 2010, 24, 344–351. [Google Scholar] [CrossRef]
- Tang, W.; Liu, Z.L.; Mai, X.Y.; Qi, X.; Li, D.H.; Gu, Q.Q.; Li, J. Identification of Gliotoxin isolated from marine fungus as a new pyruvate kinase M2 inhibitor. Biochem. Biophys. Res. Commun. 2020, 528, 594–600. [Google Scholar] [CrossRef]
- Whyte, L.; Huang, Y.Y.; Torres, K.; Mehta, R.G. Molecular mechanisms of resveratrol action in lung cancer cells using dual protein and microarray analyses. Cancer Res. 2007, 67, 12007–12017. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [Green Version]
- Sgambato, A.; Ardito, R.; Faraglia, B.; Boninsegna, A.; Wolf, F.I.; Cittadini, A. Resveratrol, a natural phenolic compound, inhibits cell proliferation and prevents oxidative DNA damage. Mutat. Res. 2001, 496, 171–180. [Google Scholar] [CrossRef]
- Kim, Y.A.; Lee, W.H.; Choi, T.H.; Rhee, S.H.; Park, K.Y.; Choi, Y.H. Involvement of p21WAF1/CIP1, pRB, Bax and NF-kappaB in induction of growth arrest and apoptosis by resveratrol in human lung carcinoma A549 cells. Int. J. Oncol. 2003, 23, 1143–1149. [Google Scholar]
- Lin, J.N.; Lin, V.C.; Rau, K.M.; Shieh, P.C.; Kuo, D.H.; Shieh, J.C.; Chen, W.J.; Tsai, S.C.; Way, T.D. Resveratrol modulates tumor cell proliferation and protein translation via SIRT1-dependent AMPK activation. J. Agric. Food Chem. 2010, 58, 1584–1592. [Google Scholar] [CrossRef]
- Liu, M.; Wilk, S.A.; Wang, A.; Zhou, L.; Wang, R.H.; Ogawa, W.; Deng, C.; Dong, L.Q.; Liu, F. Resveratrol inhibits mTOR signaling by promoting the interaction between mTOR and DEPTOR. J. Biol. Chem. 2010, 285, 36387–36394. [Google Scholar] [CrossRef] [Green Version]
- Hey, E. Vitamin K -- what, why, and when. (Review). Arch. Dis. Child. Fetal Neonatal Ed. 2003, 88, F80–F83. [Google Scholar] [CrossRef] [Green Version]
- Aris, R.M.; Merkel, P.A.; Bachrach, L.K.; Borowitz, D.S.; Boyle, M.P.; Elkin, S.L.; Guise, T.A.; Hardin, D.S.; Haworth, C.S.; Holick, M.F.; et al. Guide to bone health and disease in cystic fibrosis. J. Clin. Endocrinol. Metab. 2005, 90, 1888. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, M.; Nakai, S.; Deguchi, A.; Nonomura, T.; Masaki, T.; Uchida, N.; Yoshiji, H.; Kuriyama, S. Vitamins K2, K3 and K5 exert antitumor effects on established colorectal cancer in mice by inducing apoptotic death of tumor cells. Int. J. Oncol. 2007, 31, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Lamson, D.W.; Plaza, S.M. The anticancer effects of vitamin K. Altern. Med. Rev. 2003, 8, 303–318. [Google Scholar]
- Wu, F.Y.H.; Liao, W.C.; Chang, H.M. Comparison of antitumor activity of vitamins K1, K2 and K3 on human tumor cells by two (MTT and SRB) cell viability assays. Life Sci. 1993, 52, 1797–1804. [Google Scholar] [CrossRef]
- Hitomi, M.; Yokoyama, F.; Kita, Y.; Nonomura, T.; Masaki, T.; Yoshiji, H.; Inoue, H.; Kinekawa, F.; Kurokohchi, K.; Uchida, N.; et al. Antitumor effects of vitamins K1, K2 and K3 on hepatocellular carcinoma in vitro and in vivo. Int. J. Oncol. 2005, 26, 713–720. [Google Scholar] [CrossRef]
- Gilloteaux, J.; Jamison, J.M.; Lorimer, H.E.; Jarjoura, D.; Taper, H.S.; Calderon, P.B.; Neal, D.R.; Summers, J.L. Autoschizis: A new form of cell death for human ovarian carcinoma cells following ascorbate: Menadione treatment nuclear and DNA degradation. Tissue Cell 2004, 36, 197–210. [Google Scholar] [CrossRef]
- Verrax, J.; Cadrobbi, J.; Delvaux, M.; Jamison, J.M.; Gilloteaux, J.; Summers, J.L.; Taper, H.S.; Buc Calderon, P. The association of vitamins C and K3 kills cancer cells mainly by autoschizis, a novel form of cell death. Basis for their potential use as coadjuvants in anticancer therapy. Eur. J. Med. Chem. 2003, 38, 451–457. [Google Scholar] [CrossRef]
- Lakhter, A.J.; Hamilton, J.; Dagher, P.C.; Mukkamala, S.; Hato, T.; Dong, X.C.; Mayo, L.D.; Harris, R.A.; Shekhar, A.; Ivan, M.; et al. Ferroxitosis: A cell death from modulation of oxidative phosphorylation and PKM2-dependent glycolysis in melanoma. Oncotarget 2014, 5, 12694–12703. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Lu, M.; Mei, M.; Wang, H.; Han, Z.; Chen, M.; Yao, H.; Song, N.; Ding, X.; Ding, J.; et al. Pyridoxine induces glutathione synthesis via PKM2-mediated Nrf2 transactivation and confers neuroprotection. Nat. Commun. 2020, 11, 941. [Google Scholar] [CrossRef] [Green Version]
- Ahn, B.Z.; Baik, K.U.; Kweon, G.R.; Lim, K.; Hwang, B.D. Acylshikonin analogues: Synthesis and inhibition of DNA topoisomerase-I. J. Med. Chem. 1995, 38, 1044–1047. [Google Scholar] [CrossRef]
- Kim, S.H.; Kang, I.C.; Yoon, T.J.; Park, Y.M.; Kang, K.S.; Song, G.Y.; Ahn, B.Z. Antitumor activities of a newly synthesized shikonin derivative, 2-hyim-DMNQ-S-33. Cancer Lett. 2001, 172, 171–175. [Google Scholar] [CrossRef]
- Hu, X.; Xuan, Y. Bypassing cancer drug resistance by activating multiple death pathways--a proposal from the study of circumventing cancer drug resistance by induction of necroptosis. Cancer Lett. 2008, 259, 127–137. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, P.; Huang, H.; Chen, D.; Ma, N.; Cui, Q.C.; Shen, S.; Dong, W.; Zhang, X.; Lian, W.; et al. Shikonin exerts antitumor activity via proteasome inhibition and cell death induction in vitro and in vivo. Int. J. Cancer 2009, 124, 2450–2459. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, F.; Wu, X.-R. Inhibition of Pyruvate Kinase M2 Markedly Reduces Chemoresistance of Advanced Bladder Cancer to Cisplatin. Sci. Rep. 2017, 7, 45983. [Google Scholar] [CrossRef] [Green Version]
- Nigorikawa, K.; Yoshikawa, K.; Sasaki, T.; Iida, E.; Tsukamoto, M.; Murakami, H.; Maehama, T.; Hazeki, K.; Hazeki, O. A Naphthoquinone Derivative, Shikonin, Has Insulin-Like Actions by Inhibiting Both Phosphatase and Tensin Homolog Deleted on Chromosome 10 and Tyrosine Phosphatases. Mol. Pharmacol. 2006, 70, 1143. [Google Scholar] [CrossRef]
- Lu, Y.-M.; Cheng, F.; Teng, L.-S. The association between phosphatase and tensin homolog hypermethylation and patients with breast cancer, a meta-analysis and literature review. Sci. Rep. 2016, 6, 32723. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, F.; Sellers, W.R. The PTEN tumor suppressor protein: An antagonist of phosphoinositide 3-kinase signaling. Biochim. Biophys. Acta 2000, 1470, M21–M35. [Google Scholar] [CrossRef]
- Dubé, N.; Tremblay, M.L. Involvement of the small protein tyrosine phosphatases TC-PTP and PTP1B in signal transduction and diseases: From diabetes, obesity to cell cycle, and cancer. Biochim. Biophys. Acta 2005, 1754, 108–117. [Google Scholar] [CrossRef]
- Liu, T.; Li, S.; Wu, L.; Yu, Q.; Li, J.; Feng, J.; Zhang, J.; Chen, J.; Zhou, Y.; Ji, J.; et al. Experimental Study of Hepatocellular Carcinoma Treatment by Shikonin Through Regulating PKM2. J. Hepatocell. Carcinoma 2020, 7, 19–31. [Google Scholar] [CrossRef] [Green Version]
- James, A.D.; Richardson, D.A.; Oh, I.-W.; Sritangos, P.; Attard, T.; Barrett, L.; Bruce, J.I.E. Cutting off the fuel supply to calcium pumps in pancreatic cancer cells: Role of pyruvate kinase-M2 (PKM2). Br. J. Cancer 2020, 122, 266–278. [Google Scholar] [CrossRef]
- Guo, C.; He, J.; Song, X.; Tan, L.; Wang, M.; Jiang, P.; Li, Y.; Cao, Z.; Peng, C. Pharmacological properties and derivatives of shikonin—A review in recent years. Pharmacol. Res. 2019, 149, 104463. [Google Scholar] [CrossRef]
- Kumar, A.; Gupta, P.; Rana, M.; Chandra, T.; Dikshit, M.; Barthwal, M.K. Role of pyruvate kinase M2 in oxidized LDL-induced macrophage foam cell formation and inflammation. J. Lipid Res. 2020, 61, 351–364. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, D.; Li, M.; Plecita-Hlavata, L.; D’Alessandro, A.; Tauber, J.; Riddle, S.; Kumar, S.; Flockton, A.; McKeon, B.A.; et al. Metabolic and Proliferative State of Vascular Adventitial Fibroblasts in Pulmonary Hypertension Is Regulated Through a MicroRNA-124/PTBP1 (Polypyrimidine Tract Binding Protein 1)/Pyruvate Kinase Muscle Axis. Circulation 2017, 136, 2468–2485. [Google Scholar] [CrossRef]
- Gomez-Escudero, J.; Clemente, C.; Garcia-Weber, D.; Acin-Perez, R.; Millan, J.; Enriquez, J.A.; Bentley, K.; Carmeliet, P.; Arroyo, A.G. PKM2 regulates endothelial cell junction dynamics and angiogenesis via ATP production. Sci. Rep. 2019, 9, 15022. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Jiang, Y.; Qu, C.; Yuan, H.; Hu, K.; He, L.; Chen, P.; Li, J.; Tu, M.; Lin, L.; et al. Pyruvate Kinase M2 Tetramerization Protects against Hepatic Stellate Cell Activation and Liver Fibrosis. Am. J. Pathol. 2020, 190, 2267–2281. [Google Scholar] [CrossRef]
- Vinaik, R.; Barayan, D.; Auger, C.; Abdullahi, A.; Jeschke, M.G. Regulation of glycolysis and the Warburg effect in wound healing. Jci Insight 2020, 5, e138949. [Google Scholar] [CrossRef]
- Siragusa, M.; Thole, J.; Bibli, S.I.; Luck, B.; Loot, A.E.; de Silva, K.; Wittig, I.; Heidler, J.; Stingl, H.; Randriamboavonjy, V.; et al. Nitric oxide maintains endothelial redox homeostasis through PKM2 inhibition. EMBO J. 2019, 38, e100938. [Google Scholar] [CrossRef]
- Ding, H.; Jiang, L.; Xu, J.; Bai, F.; Zhou, Y.; Yuan, Q.; Luo, J.; Zen, K.; Yang, J. Inhibiting aerobic glycolysis suppresses renal interstitial fibroblast activation and renal fibrosis. Am. J. Physiol. Ren. Physiol. 2017, 313, F561–F575. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Liu, B.; Shao, Y.; Hua, L.; Fu, R.; Wang, H.; Wang, T.; Qi, W.; Shao, Z.; Liu, C. Effects of Shikonin on the Functions of Myeloid Dendritic Cells in a Mouse Model of Severe Aplastic Anemia. Mediat. Inflamm 2020, 2020, 9025705. [Google Scholar] [CrossRef]
- Damasceno, L.E.A.; Prado, D.S.; Veras, F.P.; Fonseca, M.M.; Toller-Kawahisa, J.E.; Rosa, M.H.; Publio, G.A.; Martins, T.V.; Ramalho, F.S.; Waisman, A.; et al. PKM2 promotes Th17 cell differentiation and autoimmune inflammation by fine-tuning STAT3 activation. J. Exp. Med. 2020, 217, e20190613. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Maeda, K.; Stocton-Gavanescu, I.; Pan, W.; Umeda, M.; Katsuyama, E.; Burbano, C.; Orite, S.Y.K.; Vukelic, M.; Tsokos, M.G.; et al. Pyruvate kinase M2 is requisite for Th1 and Th17 differentiation. JCI Insight 2019, 4, e127395. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.L.; Dang, G.H.; Deng, J.C.; Liu, H.Y.; Liu, B.; Yang, J.; Ma, X.L.; Miao, Y.T.; Jiang, C.T.; Xu, Q.B.; et al. Shikonin attenuates hyperhomocysteinemia-induced CD4(+) T cell inflammatory activation and atherosclerosis in ApoE(-/-) mice by metabolic suppression. Acta Pharm. Sin. 2020, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Lu, S.; Liu, H.; Liu, B.; Jiang, C.; Xu, Q.; Feng, J.; Wang, X. Homocysteine Activates B Cells via Regulating PKM2-Dependent Metabolic Reprogramming. J. Immunol. 2017, 198, 170–183. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, N.; Damianakos, H.; Chinou, I.; Andujar, I.; Rios, J.; Kunert, O.; Boechzelt, H.; Bauer, R. Comparison of the cytotoxicity and antimicrobial activity of several isohexenylnaphthazarins. Planta Med. 2011, 77, PM199. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, D.Y.; Kim, Y.B.; Lee, S.W.; Cha, S.W.; Park, H.W.; Kim, G.S.; Kwon, D.Y.; Lee, M.H.; Han, S.H. The Mechanism Underlying the Antibacterial Activity of Shikonin against Methicillin-Resistant Staphylococcus aureus. Evid. Based Complementary Altern. Med. Ecam 2015, 2015, 520578. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.; Kato, H.; Taguchi, Y.; Umeda, M. Biological Effects of Shikonin in Human Gingival Fibroblasts via ERK 1/2 Signaling Pathway. Molecules 2019, 24, 3542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Han, H.; Sun, L.; Qiu, H.; Lin, H.; Yu, L.; Zhu, W.; Qi, J.; Yang, R.; Pang, Y.; et al. Antiviral activity of shikonin ester derivative PMM-034 against enterovirus 71 in vitro. Braz. J. Med. Biol. Res. 2017, 50, e6586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Han, H.; Qiu, H.; Lin, H.; Yu, L.; Zhu, W.; Qi, J.; Yang, R.; Pang, Y.; Wang, X.; et al. Antiviral activity of a synthesized shikonin ester against influenza A (H1N1) virus and insights into its mechanism. Biomed. Pharm. 2017, 93, 636–645. [Google Scholar] [CrossRef]
- Chen, X.; Yang, L.; Zhang, N.; Turpin, J.A.; Buckheit, R.W.; Osterling, C.; Oppenheim, J.J.; Howard, O.M. Shikonin, a component of chinese herbal medicine, inhibits chemokine receptor function and suppresses human immunodeficiency virus type 1. Antimicrob. Agents Chemother 2003, 47, 2810–2816. [Google Scholar] [CrossRef] [Green Version]
- Gwon, S.Y.; Choi, W.H.; Lee, D.H.; Ahn, J.Y.; Jung, C.H.; Moon, B.; Ha, T.Y. Shikonin protects against obesity through the modulation of adipogenesis, lipogenesis, and β-oxidation in vivo. J. Funct. Foods 2015, 16, 484–493. [Google Scholar] [CrossRef]
- Bettaieb, A.; Hosein, E.; Chahed, S.; Abdulaziz, A.; Kucera, H.R.; Gaikwad, N.W.; Haj, F.G. Decreased adiposity and enhanced glucose tolerance in shikonin treated mice. Obesity (Silver Spring) 2015, 23, 2269–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.; Huang, W.; Zhu, B. Acetylshikonin from Zicao Prevents Obesity in Rats on a High-Fat Diet by Inhibiting Lipid Accumulation and Inducing Lipolysis. PLoS ONE 2016, 11, e0146884. [Google Scholar] [CrossRef] [Green Version]
- Gwon, S.Y.; Ahn, J.; Jung, C.H.; Moon, B.; Ha, T.Y. Shikonin Attenuates Hepatic Steatosis by Enhancing Beta Oxidation and Energy Expenditure via AMPK Activation. Nutrients 2020, 12, 1133. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yang, M.; Yao, H.; Ma, Y.; Ren, X. Shikonin from Zicao prevents against non-alcoholic fatty liver disease induced by high-fat diet in rats. 2017. Available online: https://www.academiapublishing.org/print/Yang (accessed on 24 January 2021).
- Gernapudi, R.; Yao, Y.; Zhang, Y.; Wolfson, B.; Roy, S.; Duru, N.; Eades, G.; Yang, P.; Zhou, Q. Targeting exosomes from preadipocytes inhibits preadipocyte to cancer stem cell signaling in early-stage breast cancer. Breast Cancer Res. Treat. 2015, 150, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Gwon, S.Y.; Ahn, J.Y.; Jung, C.H.; Moon, B.K.; Ha, T.Y. Shikonin suppresses ERK 1/2 phosphorylation during the early stages of adipocyte differentiation in 3T3-L1 cells. BMC Complement. Altern. Med. 2013, 13, 207. [Google Scholar] [CrossRef] [Green Version]
- Madoka Kawara, R.M. Yuko Yamamoto, Go Yoneda, Rika Fujino, Kazuhiko Nishi, Hirofumi Jono and Hideyuki Saito, Nephropreventive Effect of Shikonin on Murine LPS-induced Septic Acute Kidney Injury via Nrf2 Activation with Antioxidative Responses. J. Clin. Exp. Nephrol. 2016, 1, 19. [Google Scholar]
- Tong, Y.; Chuan, J.; Bai, L.; Shi, J.; Zhong, L.; Duan, X.; Zhu, Y. The protective effect of shikonin on renal tubular epithelial cell injury induced by high glucose. Biomed. Pharm. 2018, 98, 701–708. [Google Scholar] [CrossRef]
- Wei, Q.; Su, J.; Dong, G.; Zhang, M.; Huo, Y.; Dong, Z. Glycolysis inhibitors suppress renal interstitial fibrosis via divergent effects on fibroblasts and tubular cells. Am. J. Physiol. Ren. Physiol. 2019, 316, F1162–F1172. [Google Scholar] [CrossRef]
- Guo, T.; Jiang, Z.B.; Tong, Z.Y.; Zhou, Y.; Chai, X.P.; Xiao, X.Z. Shikonin Ameliorates LPS-Induced Cardiac Dysfunction by SIRT1-Dependent Inhibition of NLRP3 Inflammasome. Front. Physiol 2020, 11, 570441. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, T.; Pan, Z.; Ge, X.; Sun, C.; Lu, C.; Chen, H.; Xiao, Z.; Zhang, B.; Dai, Y.; et al. Shikonin inhibits myeloid differentiation protein 2 to prevent LPS-induced acute lung injury. Br. J. Pharmacol. 2018, 175, 840–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, H.; Wang, M.; Gao, S.; Hong, L.; Hou, T.; Zhang, Y.; Zhu, Y.; Qian, F. Shikonin ameliorates lipoteichoic acidinduced acute lung injury via promotion of neutrophil apoptosis. Mol. Med. Rep. 2021, 23, 133. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Lin, J.; Lin, L.; Xu, Y.; Feng, J. Shikoninmediated inhibition of nestin affects hypoxiainduced proliferation of pulmonary artery smooth muscle cells. Mol. Med. Rep. 2018, 18, 3476–3482. [Google Scholar] [PubMed] [Green Version]
- Guo, H.; Sun, J.; Li, D.; Hu, Y.; Yu, X.; Hua, H.; Jing, X.; Chen, F.; Jia, Z.; Xu, J. Shikonin attenuates acetaminophen-induced acute liver injury via inhibition of oxidative stress and inflammation. Biomed. Pharm. 2019, 112, 108704. [Google Scholar] [CrossRef]
- Tian, Y.; Li, Z.; Shen, B.; Wu, L.; Han, L.; Zhang, Q.; Feng, H. The protective effects of Shikonin on lipopolysaccharide/d-galactosamine-induced acute liver injury via inhibiting MAPK and NF-κB and activating Nrf2/HO-1 signaling pathways. RSC Adv. 2017, 7, 34846–34856. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, Q.; Mo, W.; Yu, Q.; Xu, S.; Li, J.; Li, S.; Feng, J.; Wu, L.; Lu, X.; et al. The protective effects of shikonin on hepatic ischemia/reperfusion injury are mediated by the activation of the PI3K/Akt pathway. Sci. Rep. 2017, 7, 44785. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Xu, L.; Wang, C.; Chen, K.; Xia, Y.; Li, J.; Li, S.; Wu, L.; Feng, J.; Xu, S.; et al. Alleviation of hepatic fibrosis and autophagy via inhibition of transforming growth factor-beta1/Smads pathway through shikonin. J. Gastroenterol. Hepatol. 2019, 34, 263–276. [Google Scholar] [CrossRef]
- Liu, T.; Xia, Y.; Li, J.; Li, S.; Feng, J.; Wu, L.; Zhang, R.; Xu, S.; Cheng, K.; Zhou, Y.; et al. Shikonin Attenuates Concanavalin A-Induced Acute Liver Injury in Mice via Inhibition of the JNK Pathway. Mediat. Inflamm. 2016, 2016, 2748367. [Google Scholar] [CrossRef]
- Song, M.; Zhang, H.; Chen, Z.; Yang, J.; Li, J.; Shao, S.; Liu, J. Shikonin reduces hepatic fibrosis by inducing apoptosis and inhibiting autophagy via the platelet-activating factor-mitogen-activated protein kinase axis. Exp. Med. 2021, 21, 28. [Google Scholar]
- Andujar, I.; Recio, M.C.; Bacelli, T.; Giner, R.M.; Rios, J.L. Shikonin reduces oedema induced by phorbol ester by interfering with IkappaBalpha degradation thus inhibiting translocation of NF-kappaB to the nucleus. Br. J. Pharmacol. 2010, 160, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, T.; Gan, L.; Wang, T.; Yuan, X.; Zhang, B.; Chen, H.; Zheng, Q. Shikonin protects mouse brain against cerebral ischemia/reperfusion injury through its antioxidant activity. Eur. J. Pharmacol. 2010, 643, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Nasrollahzadeh Sabet, M.; Biglari, S.; Khorram Khorshid, H.R.; Esmaeilzadeh, E. Shikonin ameliorates experimental autoimmune encephalomyelitis (EAE) via immunomodulatory, anti-apoptotic and antioxidative activity. J. Pharm. Pharm. 2020, 72, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.N.; Son, M.S.; Park, J.H.; Lee, E.H. Shikonins attenuate microglial inflammatory responses by inhibition of ERK, Akt, and NF-kappaB: Neuroprotective implications. Neuropharmacology 2008, 55, 819–825. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509. [Google Scholar] [CrossRef]
- Hibner, G.; Kimsa-Furdzik, M.; Francuz, T. Relevance of MicroRNAs as Potential Diagnostic and Prognostic Markers in Colorectal Cancer. Int. J. Mol. Sci. 2018, 19, 2944. [Google Scholar] [CrossRef] [Green Version]
- Nama, S.; Muhuri, M.; Di Pascale, F.; Quah, S.; Aswad, L.; Fullwood, M.; Sampath, P. MicroRNA-138 is a Prognostic Biomarker for Triple-Negative Breast Cancer and Promotes Tumorigenesis via TUSC2 repression. Sci. Rep. 2019, 9, 12718. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I. MiR-15a and miR-16-1 cluster functions in human leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal. Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhang, C.; Zhao, Y.; Feng, Z. MicroRNA Control of p53. J. Cell. Biochem. 2017, 118, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Takwi, A.A.; Li, Y.; Becker Buscaglia, L.E.; Zhang, J.; Choudhury, S.; Park, A.K.; Liu, M.; Young, K.H.; Park, W.Y.; Martin, R.C.; et al. A statin-regulated microRNA represses human c-Myc expression and function. Embo Mol. Med. 2012, 4, 896–909. [Google Scholar] [CrossRef] [PubMed]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. MicroRNA let-7a down-regulates MYC and reverts MYC-induced growth in Burkitt lymphoma cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, Y.; Shimizu, Y.; Hasegawa, H.; Ouchi, Y.; Qiao, S.; Nagahara, M.; Ichihara, M.; Lee, J.D.; Adachi, K.; Hamaguchi, M.; et al. Forced expression of miR-143 represses ERK5/c-Myc and p68/p72 signaling in concert with miR-145 in gut tumors of Apc(Min) mice. PLoS ONE 2012, 7, e42137. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Xie, G.; Tong, J.; Peng, Y.; Huang, H.; Li, J.; Wang, N.; Liang, H. Overexpression of microRNA-122 re-sensitizes 5-FU-resistant colon cancer cells to 5-FU through the inhibition of PKM2 in vitro and in vivo. Cell Biochem. Biophys. 2014, 70, 1343–1350. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Y.; Ruan, Y.; Bai, C.; Qiu, L.; Cui, Y.; Ying, G.; Li, B. PKM2 promotes reductive glutamine metabolism. Cancer Biol Med. 2018, 15, 389–399. [Google Scholar]
- Li, Q.; Pan, X.; Zhu, D.; Deng, Z.; Jiang, R.; Wang, X. Circular RNA MAT2B Promotes Glycolysis and Malignancy of Hepatocellular Carcinoma Through the miR-338-3p/PKM2 Axis Under Hypoxic Stress. Hepatology 2019, 70, 1298–1316. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Y.; Chen, X.; He, Y.; Hu, Q.; Li, H.; Han, Q.; Ren, F.; Li, J.; Li, C.; et al. MiR-139-5p is associated with poor prognosis and regulates glycolysis by repressing PKM2 in gallbladder carcinoma. Cell Prolif. 2018, 51, e12510. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Kong, W.; Zhang, J.; Chen, Y.; Xue, W.; Liu, D.; Huang, Y. c-Myc modulates glucose metabolism via regulation of miR-184/PKM2 pathway in clear-cell renal cell carcinoma. Int. J. Oncol. 2016, 49, 1569–1575. [Google Scholar] [CrossRef]
- Wen, Y.Y.; Liu, W.T.; Sun, H.R.; Ge, X.; Shi, Z.M.; Wang, M.; Li, W.; Zhang, J.Y.; Liu, L.Z.; Jiang, B.H. IGF-1-mediated PKM2/beta-catenin/miR-152 regulatory circuit in breast cancer. Sci. Rep. 2017, 7, 15897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, B.; Chen, J.; Hu, L.; Zhao, C. MiR-338-3p targets pyruvate kinase M2 and affects cell proliferation and metabolism of ovarian cancer. Am. J. Transl. Res. 2016, 8, 3266–3273. [Google Scholar] [PubMed]
- Liu, A.M.; Xu, Z.; Shek, F.H.; Wong, K.F.; Lee, N.P.; Poon, R.T.; Chen, J.; Luk, J.M. miR-122 targets pyruvate kinase M2 and affects metabolism of hepatocellular carcinoma. PLoS ONE 2014, 9, e86872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.; Wang, X.; Shi, K.; Zheng, Y.; Li, J.; Chen, Y.; Jin, L.; Pan, Z. MiR-122 Reverses the Doxorubicin-Resistance in Hepatocellular Carcinoma Cells through Regulating the Tumor Metabolism. PLoS ONE 2016, 11, e0152090. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhao, X.; Zhou, Y.; Hu, Y. miR-124, miR-137 and miR-340 regulate colorectal cancer growth via inhibition of the Warburg effect. Oncol. Rep. 2012, 28, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Sun, W.; Shen, Y.; Hu, Y.; Liu, H.; Li, W.; Wang, Y. PKM2 functions as a potential oncogene and is a crucial target of miR-148a and miR-326 in thyroid tumorigenesis. Am. J. Transl. Res. 2018, 10, 1793–1801. [Google Scholar]
- Guo, M.; Zhao, X.; Yuan, X.; Jiang, J.; Li, P. MiR-let-7a inhibits cell proliferation, migration, and invasion by down-regulating PKM2 in cervical cancer. Oncotarget 2017, 8, 28226–28236. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.S.; Liu, X.B.; Chung-Wai Ho, A.; Po-Wing Yuen, A.; Wai-Man Ng, R.; Ignace Wei, W. Identification of pyruvate kinase type M2 as potential oncoprotein in squamous cell carcinoma of tongue through microRNA profiling. Int. J. Cancer 2008, 123, 251–257. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, C.; Zhang, M.; Zeng, A.; Si, L.; Yu, N.; Bai, M. miR-625-5p/PKM2 negatively regulates melanoma glycolysis state. J. Cell. Biochem. 2019, 120, 2964–2972. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, M.; Jiang, H.; Liu, F.; Liu, H.; Li, Y. Down-regulation of miR-214 inhibits proliferation and glycolysis in non-small-cell lung cancer cells via down-regulating the expression of hexokinase 2 and pyruvate kinase isozyme M2. Biomed. Pharm. 2018, 105, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, M.; Tu, J.; Pang, L.; Cai, W.; Liu, X. MicroRNA-122 affects cell aggressiveness and apoptosis by targeting PKM2 in human hepatocellular carcinoma. Oncol. Rep. 2015, 34, 2054–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhang, H.; Hong, H.; Zhang, Z. MiR-374b re-sensitizes hepatocellular carcinoma cells to sorafenib therapy by antagonizing PKM2-mediated glycolysis pathway. Am. J. Cancer Res. 2019, 9, 765–778. [Google Scholar] [PubMed]
- Xu, Q.; Dou, C.; Liu, X.; Yang, L.; Ni, C.; Wang, J.; Guo, Y.; Yang, W.; Tong, X.; Huang, D. Oviductus ranae protein hydrolysate (ORPH) inhibits the growth, metastasis and glycolysis of HCC by targeting miR-491-5p/PKM2 axis. Biomed. Pharm. 2018, 107, 1692–1704. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, E.; Jung, J.; Lee, J.W.; Kim, H.J.; Kim, J.; Yoo, H.J.; Lee, H.J.; Chae, S.Y.; Jeon, S.M.; et al. microRNA-155 positively regulates glucose metabolism via PIK3R1-FOXO3a-cMYC axis in breast cancer. Oncogene 2018, 37, 2982–2991. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zheng, X.; Lu, J.; Chen, W.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 Inhibits the Warburg Effect Via Modulating DNMT3A/MiR-532-3p/HK2 Pathway in Ovarian Cancer Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 45, 2548–2559. [Google Scholar] [CrossRef]
- Teng, Y.; Zhang, Y.; Qu, K.; Yang, X.; Fu, J.; Chen, W.; Li, X. MicroRNA-29B (mir-29b) regulates the Warburg effect in ovarian cancer by targeting AKT2 and AKT3. Oncotarget 2015, 6, 40799–40814. [Google Scholar] [CrossRef] [Green Version]
- Kefas, B.; Comeau, L.; Erdle, N.; Montgomery, E.; Amos, S.; Purow, B. Pyruvate kinase M2 is a target of the tumor-suppressive microRNA-326 and regulates the survival of glioma cells. Neuro-oncology 2010, 12, 1102–1112. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Li, Y.I.; Gao, X. Overexpression of microRNA-133b sensitizes non-small cell lung cancer cells to irradiation through the inhibition of glycolysis. Oncol Lett 2016, 11, 2903–2908. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhao, Z.; Zhou, Z.; Liu, R. Linc-ROR confers gemcitabine resistance to pancreatic cancer cells via inducing autophagy and modulating the miR-124/PTBP1/PKM2 axis. Cancer Chemother. Pharmacol. 2016, 78, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, Y.; Wu, C.; Yang, P.; Li, H.; Li, Z. Resveratrol Induces Cancer Cell Apoptosis through MiR-326/PKM2-Mediated ER Stress and Mitochondrial Fission. J. Agric. Food Chem. 2016, 64, 9356–9367. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, Z.; Guo, W.; Zhang, Z.; Zhao, F.; Zhao, Y.; Jia, D.; Ding, J.; Wang, H.; Yao, M.; et al. TRIM35 Interacts with pyruvate kinase isoform M2 to suppress the Warburg effect and tumorigenicity in hepatocellular carcinoma. Oncogene 2015, 34, 3946–3956. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Zhang, W.; Chang, Z.; Pan, Y.; Zong, H.; Fan, Q.; Wang, L. miR-4417 Targets Tripartite Motif-Containing 35 (TRIM35) and Regulates Pyruvate Kinase Muscle 2 (PKM2) Phosphorylation to Promote Proliferation and Suppress Apoptosis in Hepatocellular Carcinoma Cells. Med. Sci. Monit.: Int. Med. J. Exp. Clin. Res. 2017, 23, 1741–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, R.; Karantza-Wadsworth, V.; White, E. Role of autophagy in cancer. Nat. Rev. Cancer 2007, 7, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, K.; Sakai, M.; Sugito, N.; Kumazaki, M.; Shinohara, H.; Yamada, N.; Nakayama, T.; Ueda, H.; Nakagawa, Y.; Ito, Y.; et al. PTBP1-associated microRNA-1 and -133b suppress the Warburg effect in colorectal tumors. Oncotarget 2016, 7, 18940–18952. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Zhang, Y.; Zhou, L.; Wang, X.; Mu, J.; Jiang, L.; Hu, Y.; Dong, P.; Liu, Y. miR-122 inhibits cancer cell malignancy by targeting PKM2 in gallbladder carcinoma. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015. [Google Scholar] [CrossRef]
- Tang, R.; Yang, C.; Ma, X.; Wang, Y.; Luo, D.; Huang, C.; Xu, Z.; Liu, P.; Yang, L. MiR-let-7a inhibits cell proliferation, migration, and invasion by down-regulating PKM2 in gastric cancer. Oncotarget 2016, 7, 5972–5984. [Google Scholar] [CrossRef]
- Taniguchi, K.; Sugito, N.; Kumazaki, M.; Shinohara, H.; Yamada, N.; Nakagawa, Y.; Ito, Y.; Otsuki, Y.; Uno, B.; Uchiyama, K.; et al. MicroRNA-124 inhibits cancer cell growth through PTB1/PKM1/PKM2 feedback cascade in colorectal cancer. Cancer Lett. 2015, 363, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, Y.; Cao, W.; Liu, Y. miR-491-5p inhibits osteosarcoma cell proliferation by targeting PKM2. Oncol Lett 2018, 16, 6472–6478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Lu, Y.; Meng, Q.; Wang, C.; Li, X.; Yang, Y.; Xin, X.; Zheng, Q.; Xu, J.; Gui, X.; et al. miR372 Promotes Progression of Liver Cancer Cells by Upregulating erbB-2 through Enhancement of YB-1. Mol. Nucleic Acids 2018, 11, 494–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Qiu, Y.; Hao, J.; Zhao, C.; Deng, X.; Shu, G. Dauricine upregulates the chemosensitivity of hepatocellular carcinoma cells: Role of repressing glycolysis via miR-199a:HK2/PKM2 modulation. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 121, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, L.; Zuo, D.; He, W.; Wang, Y.; Zhang, Y.; Liu, W.; Yuan, Y. Mutual Regulation of MiR-199a-5p and HIF-1alpha Modulates the Warburg Effect in Hepatocellular Carcinoma. J. Cancer 2017, 8, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Yuan, J.D.; Cao, Z.L.; Huang, T.; Zhang, C.H.; Wang, L.; Zeng, F.Q. [MiR-124 suppresses the proliferation of human prostate cancer PC3 cells by targeting PKM2]. Zhonghua Nan Ke Xue 2014, 20, 495–499. [Google Scholar] [PubMed]
- Li, J.; Zhang, S.; Zou, Y.; Wu, L.; Pei, M.; Jiang, Y. miR-145 promotes miR-133b expression through c-myc and DNMT3A-mediated methylation in ovarian cancer cells. J. Cell. Physiol. 2020, 235, 4291–4301. [Google Scholar] [CrossRef]
- Yin, J.; Shi, Z.; Wei, W.; Lu, C.; Wei, Y.; Yan, W.; Li, R.; Zhang, J.; You, Y.; Wang, X. MiR-181b suppress glioblastoma multiforme growth through inhibition of SP1-mediated glucose metabolism. Cancer Cell Int. 2020, 20, 69. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.Y.; Liu, X.J.; Zhan, Y.; Liu, M.H.; Zhang, X.Y.; Li, Z.Y.; Lu, Y.Q.; Luo, J.M.; Yang, L. miR-140-5p induces cell apoptosis and decreases Warburg effect in chronic myeloid leukemia by targeting SIX1. Biosci. Rep. 2019, 39, BSR20190150. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Yu, H.; Chen, J.; Song, X.; Sun, L. Antitumor Effect of miR-1294/Pyruvate Kinase M2 Signaling Cascade in Osteosarcoma Cells. Onco Target 2020, 13, 1637–1647. [Google Scholar] [CrossRef] [Green Version]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [PubMed]
- Su, Q.; Tao, T.; Tang, L.; Deng, J.; Darko, K.O.; Zhou, S.; Peng, M.; He, S.; Zeng, Q.; Chen, A.F.; et al. Down-regulation of PKM2 enhances anticancer efficiency of THP on bladder cancer. J. Cell. Mol. Med. 2018, 22, 2774–2790. [Google Scholar] [CrossRef] [PubMed]
- Meoli, L.; Gupta, N.K.; Saeidi, N.; Panciotti, C.A.; Biddinger, S.B.; Corey, K.E.; Stylopoulos, N. Nonalcoholic fatty liver disease and gastric bypass surgery regulate serum and hepatic levels of pyruvate kinase isoenzyme M2. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E613–E621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.C.W.; Carella, M.A.; Papa, S.; Bubici, C. High Expression of Glycolytic Genes in Cirrhosis Correlates with the Risk of Developing Liver Cancer. Front. Cell Dev. Biol. 2018, 6, 138. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Han, S.N.; Arumugam, S.; Yousaf, M.N.; Qin, Y.; Jiang, J.X.; Torok, N.J.; Chen, Y.; Mankash, M.S.; Liu, J.; et al. Digoxin improves steatohepatitis with differential involvement of liver cell subsets in mice through inhibition of PKM2 transactivation. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G387–G397. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.; Zhao, X.; Wang, L.; Wang, X.; Wang, J.; Dong, B.; Gong, D. MicroRNA-122 targets genes related to goose fatty liver. Poult. Sci. 2018, 97, 643–649. [Google Scholar] [CrossRef]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, M.B. miR-122--a key factor and therapeutic target in liver disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Shao, F.; Yu, J.; Jiang, H.; Gong, D.; Gu, Z. MicroRNA-122 targets genes related to liver metabolism in chickens. Comp. Biochem. Physiol. Part. BBiochem. Mol. Biol. 2015, 184, 29–35. [Google Scholar] [CrossRef]
- Auguet, T.; Aragones, G.; Berlanga, A.; Guiu-Jurado, E.; Marti, A.; Martinez, S.; Sabench, F.; Hernandez, M.; Aguilar, C.; Sirvent, J.J.; et al. miR33a/miR33b* and miR122 as Possible Contributors to Hepatic Lipid Metabolism in Obese Women with Nonalcoholic Fatty Liver Disease. Int J. Mol. Sci 2016, 17, 1620. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Liu, S.; Fan, B.; Xu, X.; Chen, Y.; Lu, R.; Xu, Z.; Liu, X. PKM2 is involved in neuropathic pain by regulating ERK and STAT3 activation in rat spinal cord. J. Headache Pain 2018, 19, 7. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wei, L.; Liu, Z.R.; Yang, J.J.; Gu, X.; Wei, Z.Z.; Liu, L.P.; Yu, S.P. Pyruvate Kinase M2 Increases Angiogenesis, Neurogenesis, and Functional Recovery Mediated by Upregulation of STAT3 and Focal Adhesion Kinase Activities After Ischemic Stroke in Adult Mice. Neurother. J. Am. Soc. Exp. Neurother. 2018, 15, 770–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.L.; Wang, J.L.; Liu, X.; Zhang, J.; Liu, C.; Guo, L. Inhibition of miR-19a protects neurons against ischemic stroke through modulating glucose metabolism and neuronal apoptosis. Cell. Mol. Biol. Lett. 2019, 24, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Liu, N.; Zhang, J.; Wang, L.; Zhang, Z.; Zhu, J.; Li, Q.; Wang, Y. Inhibition of miR-143 during ischemia cerebral injury protects neurones through recovery of the hexokinase 2-mediated glucose uptake. Biosci. Rep. 2017, 37, BSR20170216. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.H.; Liu, B.; Wu, J.D.; Yan, Y.Y.; Wang, J.N. miR-143 is involved in endothelial cell dysfunction through suppression of glycolysis and correlated with atherosclerotic plaques formation. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4063–4071. [Google Scholar] [PubMed]
- Caruso, P.; Dunmore, B.J.; Schlosser, K.; Schoors, S.; Dos Santos, C.; Perez-Iratxeta, C.; Lavoie, J.R.; Zhang, H.; Long, L.; Flockton, A.R.; et al. Identification of MicroRNA-124 as a Major Regulator of Enhanced Endothelial Cell Glycolysis in Pulmonary Arterial Hypertension via PTBP1 (Polypyrimidine Tract Binding Protein) and Pyruvate Kinase M2. Circulation 2017, 136, 2451–2467. [Google Scholar] [CrossRef] [PubMed]
- Dewachter, L.; Adnot, S.; Guignabert, C.; Tu, L.; Marcos, E.; Fadel, E.; Humbert, M.; Dartevelle, P.; Simonneau, G.; Naeije, R.; et al. Bone morphogenetic protein signalling in heritable versus idiopathic pulmonary hypertension. Eur. Respir. J. 2009, 34, 1100–1110. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, J.; Chen, Q.D.; Qian, X.; Li, Q.; Yin, Y.; Shi, Z.M.; Wang, L.; Lin, J.; Liu, L.Z.; et al. Insulin promotes glucose consumption via regulation of miR-99a/mTOR/PKM2 pathway. PLoS ONE 2013, 8, e64924. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Guo, M.; Fei, D.; Zhang, L.; Xing, M. Targeting the miR-122/PKM2 autophagy axis relieves arsenic stress. J. Hazard. Mater. 2020, 383, 121217. [Google Scholar] [CrossRef]
- Wade, S.M.; Ohnesorge, N.; McLoughlin, H.; Biniecka, M.; Carter, S.P.; Trenkman, M.; Cunningham, C.C.; McGarry, T.; Canavan, M.; Kennedy, B.N.; et al. Dysregulated miR-125a promotes angiogenesis through enhanced glycolysis. EBioMedicine 2019, 47, 402–413. [Google Scholar] [CrossRef] [Green Version]
- Ohno, S. So much “junk” DNA in our genome. Brookhaven Symp. Biol. 1972, 23, 366–370. [Google Scholar]
- Dhanoa, J.K.; Sethi, R.S.; Verma, R.; Arora, J.S.; Mukhopadhyay, C.S. Long non-coding RNA: Its evolutionary relics and biological implications in mammals: A review. J. Anim. Sci. Technol. 2018, 60, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA-LncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [PubMed]
- Cui, S.; Yang, X.; Zhang, L.; Zhao, Y.; Yan, W. LncRNA MAFG-AS1 promotes the progression of colorectal cancer by sponging miR-147b and activation of NDUFA4. Biochem. Biophys. Res. Commun. 2018, 506, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.L.; Li, L.; Jia, Y.X.; Zhang, B.Z.; Li, J.C.; Zhu, Y.H.; Li, M.Q.; He, J.Z.; Zeng, T.T.; Ban, X.J.; et al. LINC01554-Mediated Glucose Metabolism Reprogramming Suppresses Tumorigenicity in Hepatocellular Carcinoma via Downregulating PKM2 Expression and Inhibiting Akt/mTOR Signaling Pathway. Theranostics 2019, 9, 796–810. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Wang, X.; Jiang, C.; Han, S.; Dong, K.; Shen, M.; Xu, D. LincRNA-p21 suppresses development of human prostate cancer through inhibition of PKM2. Cell Prolif. 2017, 50, e12395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Z.; Zhang, J.; Li, M.; Feng, Y.; Wang, X.; Zhang, J.; Yao, S.; Jin, G.; Du, J.; Han, W.; et al. LncRNA-FEZF1-AS1 Promotes Tumor Proliferation and Metastasis in Colorectal Cancer by Regulating PKM2 Signaling. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 4808–4819. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhu, Q.; Guo, Y.; Xiao, Z.; Hu, L.; Xu, Q. LncRNA LINC00689 promotes the growth, metastasis and glycolysis of glioma cells by targeting miR-338-3p/PKM2 axis. Biomed. Pharm. 2019, 117, 109069. [Google Scholar] [CrossRef]
- Zheng, X.; Zhou, Y.; Chen, W.; Chen, L.; Lu, J.; He, F.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 Prevents PKM2-Targeting miR-324-5p from H19 Sponging to Antagonize the Warburg Effect in Ovarian Cancer Cells. Cell. Physiol. Biochem.: Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 1340–1353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, L. Inhibition of breast cancer cell proliferation and tumorigenesis by long non-coding RNA RPPH1 down-regulation of miR-122 expression. Cancer Cell Int 2017, 17, 109. [Google Scholar] [CrossRef] [Green Version]
- Luan, X.; Wang, Y. LncRNA XLOC_006390 facilitates cervical cancer tumorigenesis and metastasis as a ceRNA against miR-331-3p and miR-338-3p. J. Gynecol. Oncol. 2018, 29, e95. [Google Scholar] [CrossRef]
- Xin, X.; Wu, M.; Meng, Q.; Wang, C.; Lu, Y.; Yang, Y.; Li, X.; Zheng, Q.; Pu, H.; Gui, X.; et al. Long noncoding RNA HULC accelerates liver cancer by inhibiting PTEN via autophagy cooperation to miR15a. Mol. Cancer 2018, 17, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Q.; Mi, B.; Xu, F.; Wen, J.; Zhao, L.; Liu, J.; Huang, G. Hypoxia-induced lncRNA-AC020978 promotes proliferation and glycolytic metabolism of non-small cell lung cancer by regulating PKM2/HIF-1α axis. Theranostics 2020, 10, 4762–4778. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, D.; Zheng, T.; Yang, G.; Wang, J.; Meng, F.; Liu, Y.; Zhang, G.; Zhang, L.; Han, J.; et al. lncRNA-SOX2OT promotes hepatocellular carcinoma invasion and metastasis through miR-122-5p-mediated activation of PKM2. Oncogenesis 2020, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Liu, R.; Wang, L.; Tang, M.; Li, S.R.; Hu, X. LncRNA RPPH1 attenuates Aβ(25-35)-induced endoplasmic reticulum stress and apoptosis in SH-SY5Y cells via miR-326/PKM2. Int. J. Neurosci. 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, H.; Du, Y.; Liu, P.; Zhang, J.; Li, Y.; Shen, H.; Xing, L.; Xue, X.; Chen, J.; et al. Long noncoding RNA TP53TG1 promotes pancreatic ductal adenocarcinoma development by acting as a molecular sponge of microRNA-96. Cancer Sci. 2019, 110, 2760–2772. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Gao, Y.; Li, D.; Cao, Y.; Hao, B. LncRNA-TP53TG1 Participated in the Stress Response Under Glucose Deprivation in Glioma. J. Cell. Biochem. 2017, 118, 4897–4904. [Google Scholar] [CrossRef]
- Huang, J.Z.; Chen, M.; Chen, D.; Gao, X.C.; Zhu, S.; Huang, H.; Hu, M.; Zhu, H.; Yan, G.R. A Peptide Encoded by a Putative lncRNA HOXB-AS3 Suppresses Colon Cancer Growth. Mol. Cell 2017, 68, 171–184.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Z.; Yao, X.; Sun, K.; Li, A.; Liu, S.; Wang, X. The Interaction Between lncRNA SNHG6 and hnRNPA1 Contributes to the Growth of Colorectal Cancer by Enhancing Aerobic Glycolysis Through the Regulation of Alternative Splicing of PKM. Front. Oncol. 2020, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, W.R.; Zhang, B.; Zhang, L.M.; Li, C.G.; Liu, C.; Zhang, H.; Huo, Y.S.; Ma, Y.C.; Tian, P.F.; et al. LncRNA H19 downregulation confers erlotinib resistance through upregulation of PKM2 and phosphorylation of AKT in EGFR-mutant lung cancers. Cancer Lett. 2020, 486, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, X.; Chen, Y.; Cao, D. Long non-coding RNA LINC00504 regulates the Warburg effect in ovarian cancer through inhibition of miR-1244. Mol. Cell. Biochem. 2020, 464, 39–50. [Google Scholar] [CrossRef]
- Ghemrawi, R.; Khair, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 6127. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Gong, Y.; Wu, L.; Li, J.; He, W.; Zhu, X.; Xiao, H. LncRNA-Malat1 is Involved in Lipotoxicity-Induced ss-cell Dysfunction and the Therapeutic Effect of Exendin-4 via Ptbp1. Endocrinology 2020, 161, bqaa065. [Google Scholar] [CrossRef]
- Yu, C.Y.; Kuo, H.C. The emerging roles and functions of circular RNAs and their generation. J. Biomed. Sci. 2019, 26, 29. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, A.; Banerjee, S.; Sharma, V.; Khandelia, P. Circular RNAs: Emerging Role in Cancer Diagnostics and Therapeutics. Front. Mol. Biosci. 2020, 7, 577938. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Xiao, Y.; Ma, J.; Tang, Y.; Tian, B.; Zhang, Y.; Li, X.; Wu, Z.; Yang, D.; Zhou, Y.; et al. Circular RNAs in Cancer: Emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol. Cancer 2019, 18, 90. [Google Scholar] [CrossRef]
- Shang, W.; Adzika, G.K.; Li, Y.; Huang, Q.; Ding, N.; Chinembiri, B.; Rashid, M.S.; Machuki, J.O. Molecular mechanisms of circular RNAs, transforming growth factor-beta, and long noncoding RNAs in hepatocellular carcinoma. Cancer Med. 2019, 8, 6684–6699. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Shen, M. Circular RNA hsa_circ_103809 suppresses hepatocellular carcinoma proliferation and invasion by sponging miR-620. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 555–566. [Google Scholar]
- Song, C.; Li, D.; Liu, H.; Sun, H.; Liu, Z.; Zhang, L.; Hu, Y. The competing endogenous circular RNA ADAMTS14 suppressed hepatocellular carcinoma progression through regulating microRNA-572/regulator of calcineurin 1. J. Cell. Physiol. 2019, 234, 2460–2470. [Google Scholar] [CrossRef]
- Wang, J.; Huang, K.; Shi, L.; Zhang, Q.; Zhang, S. CircPVT1 Promoted the Progression of Breast Cancer by Regulating MiR-29a-3p-Mediated AGR2-HIF-1alpha Pathway. Cancer Manag. Res. 2020, 12, 11477–11490. [Google Scholar] [CrossRef]
- Shangguan, H.; Feng, H.; Lv, D.; Wang, J.; Tian, T.; Wang, X. Circular RNA circSLC25A16 contributes to the glycolysis of non-small-cell lung cancer through epigenetic modification. Cell Death Dis. 2020, 11, 437. [Google Scholar] [CrossRef]
- Tian, S.; Han, G.; Lu, L.; Meng, X. Circ-FOXM1 contributes to cell proliferation, invasion, and glycolysis and represses apoptosis in melanoma by regulating miR-143-3p/FLOT2 axis. World J. Surg Oncol 2020, 18, 56. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, Y.; Xu, L.; Qu, C.; Zhang, L.; Xiao, X.; Chen, W.; Li, K.; Liang, Q.; Wu, H. circ-NRIP1 Promotes Glycolysis and Tumor Progression by Regulating miR-186-5p/MYH9 Axis in Gastric Cancer. Cancer Manag. Res. 2020, 12, 5945–5956. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Cai, Q.; Ma, M.; Jin, L.Y.; Weng, M.; Zhou, D.; Tang, Z.; Wang, J.D.; Quan, Z. Circular RNA FOXP1 promotes tumor progression and Warburg effect in gallbladder cancer by regulating PKLR expression. Mol. Cancer 2019, 18, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.; Li, H.; Goel, A. Piwi-interacting RNAs (piRNAs) and cancer: Emerging biological concepts and potential clinical implications. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.P.; Young, P.E.; McCorkindale, A.L.; Dang, T.H.; Clancy, J.L.; Humphreys, D.T.; Preiss, T.; Hutvagner, G.; Martin, D.I.; Cropley, J.E.; et al. The human Piwi protein Hiwi2 associates with tRNA-derived piRNAs in somatic cells. Nucleic Acids Res. 2014, 42, 8984–8995. [Google Scholar] [CrossRef] [PubMed]
- Dayton, T.L.; Gocheva, V.; Miller, K.M.; Israelsen, W.J.; Bhutkar, A.; Clish, C.B.; Davidson, S.M.; Luengo, A.; Bronson, R.T.; Jacks, T.; et al. Germline loss of PKM2 promotes metabolic distress and hepatocellular carcinoma. Genes Dev. 2016, 30, 1020–1033. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Kellner, K.; Solanki, A.; Amann, T.; Lao, N.; Barron, N. Targeting miRNAs with CRISPR/Cas9 to Improve Recombinant Protein Production of CHO Cells. Methods Mol. Biol. 2018, 1850, 221–235. [Google Scholar]
- Abdollah, N.A.; Kumitaa, T.D.; Narazah, M.Y.; Abdul Razak, S.R. Sequence-specific inhibition of microRNA-130a gene by CRISPR/Cas9 system in breast cancer cell line. J. Phys. Conf. Ser. 2017, 851, 012037. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, R.; Appel, B.; Stein, A.; Metz, Y.; Neheman, A.; Barak, M. The pyruvate kinase isoenzyme M2 (Tu M2-PK) as a tumour marker for renal cell carcinoma. Eur. J. Cancer Care 2007, 16, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.S.; Dew, T.; Lawton, F.G.; Papadopoulos, A.J.; Devaja, O.; Raju, K.S.; Sherwood, R.A. M2-PK as a novel marker in ovarian cancer. A prospective cohort study. Eur. J. Gynaecol. Oncol. 2007, 28, 83–88. [Google Scholar] [PubMed]
- Schneider, J. Tumor markers in detection of lung cancer. Adv. Clin. Chem. 2006, 42, 1–41. [Google Scholar] [PubMed]
- Wang, C.; Zhang, S.; Liu, J.; Tian, Y.; Ma, B.; Xu, S.; Fu, Y.; Luo, Y. Secreted Pyruvate Kinase M2 Promotes Lung Cancer Metastasis through Activating the Integrin Beta1/FAK Signaling Pathway. Cell Rep. 2020, 30, 1780–1797.e6. [Google Scholar] [CrossRef] [Green Version]
- Almousa, A.A.; Morris, M.; Fowler, S.; Jones, J.; Alcorn, J. Elevation of serum pyruvate kinase M2 (PKM2) in IBD and its relationship to IBD indices. Clin. Biochem. 2018, 53, 19–24. [Google Scholar] [CrossRef] [PubMed]






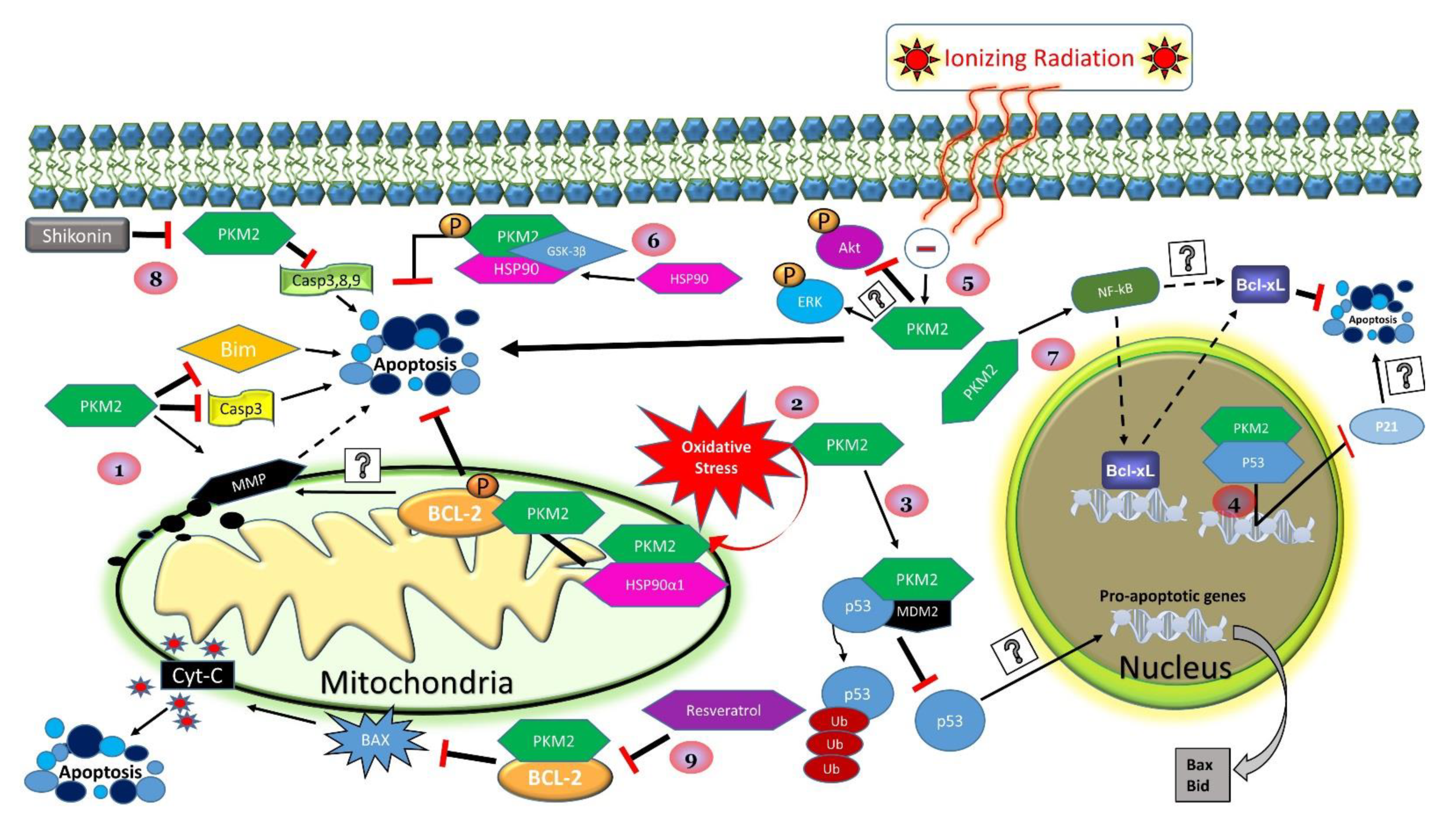{kind=link}
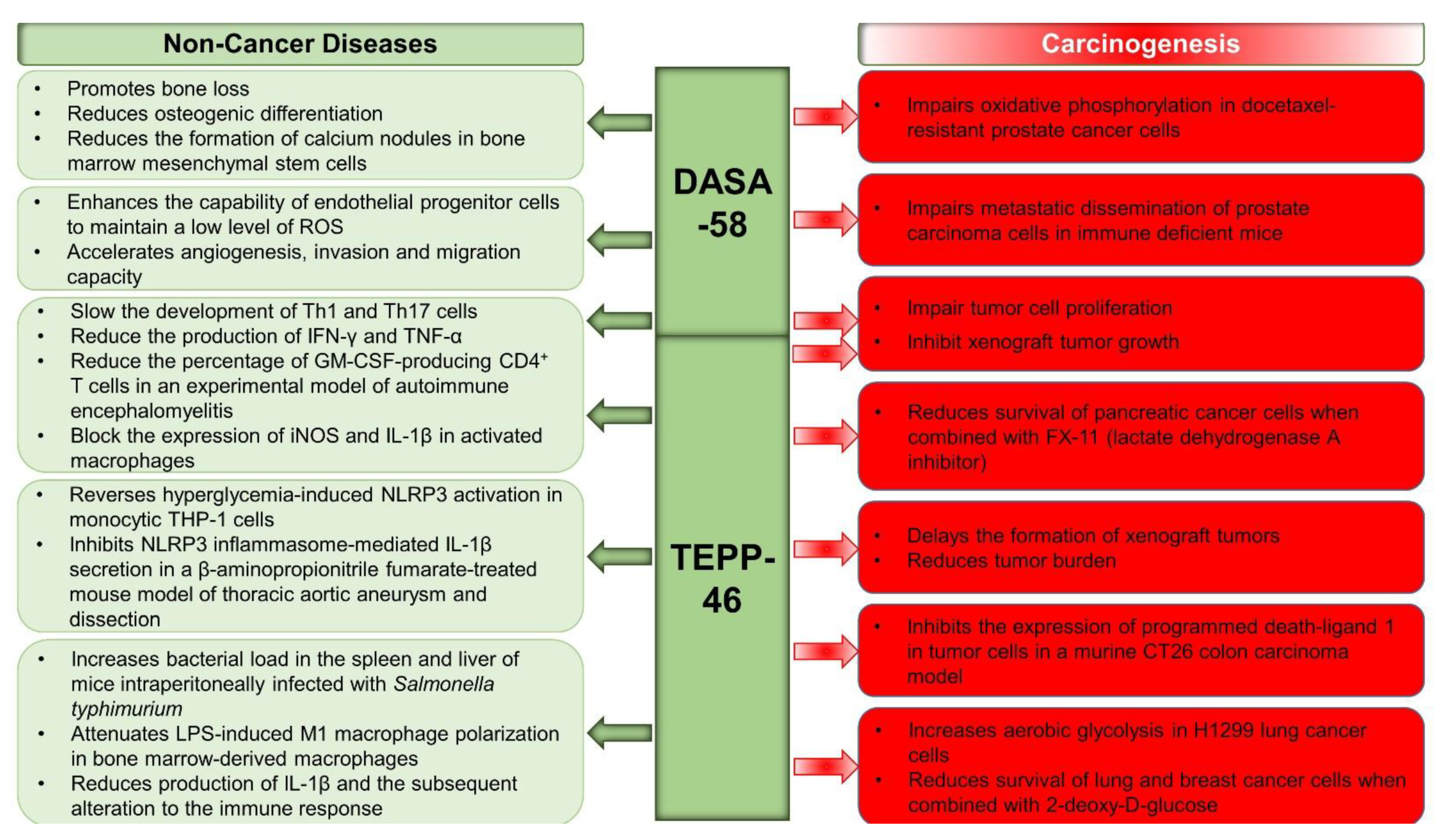{kind=link}
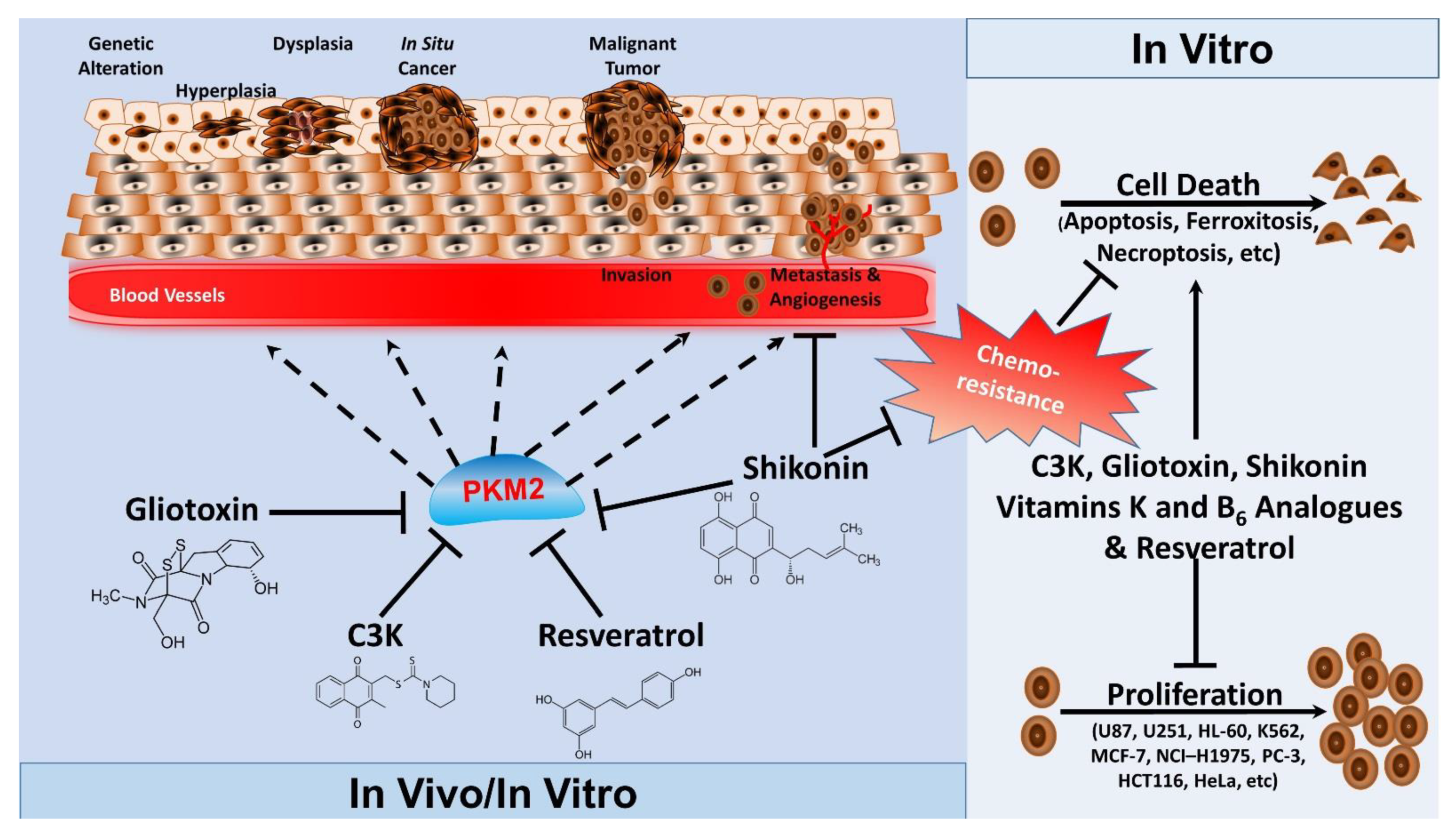{kind=link}
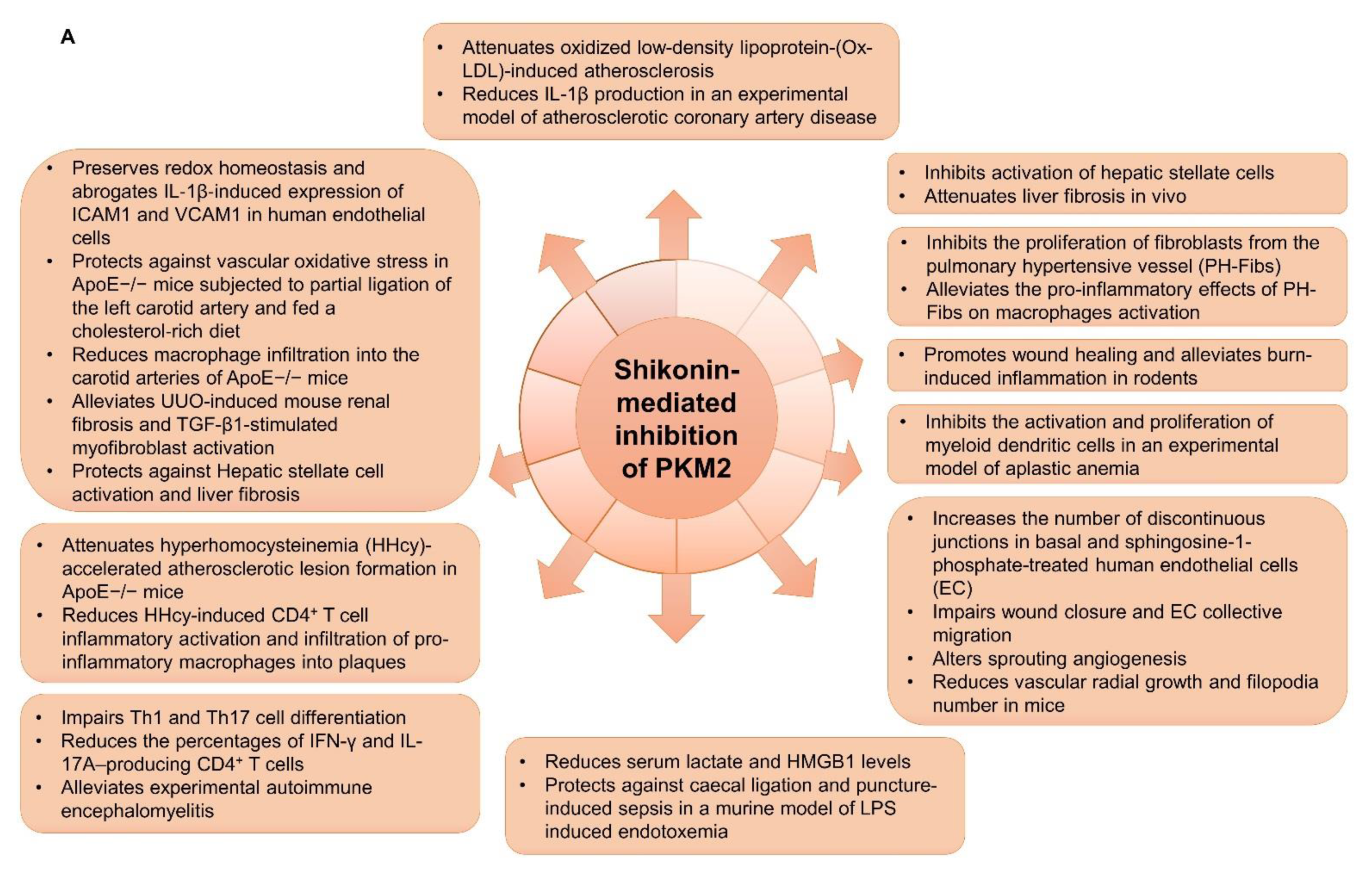{kind=link}
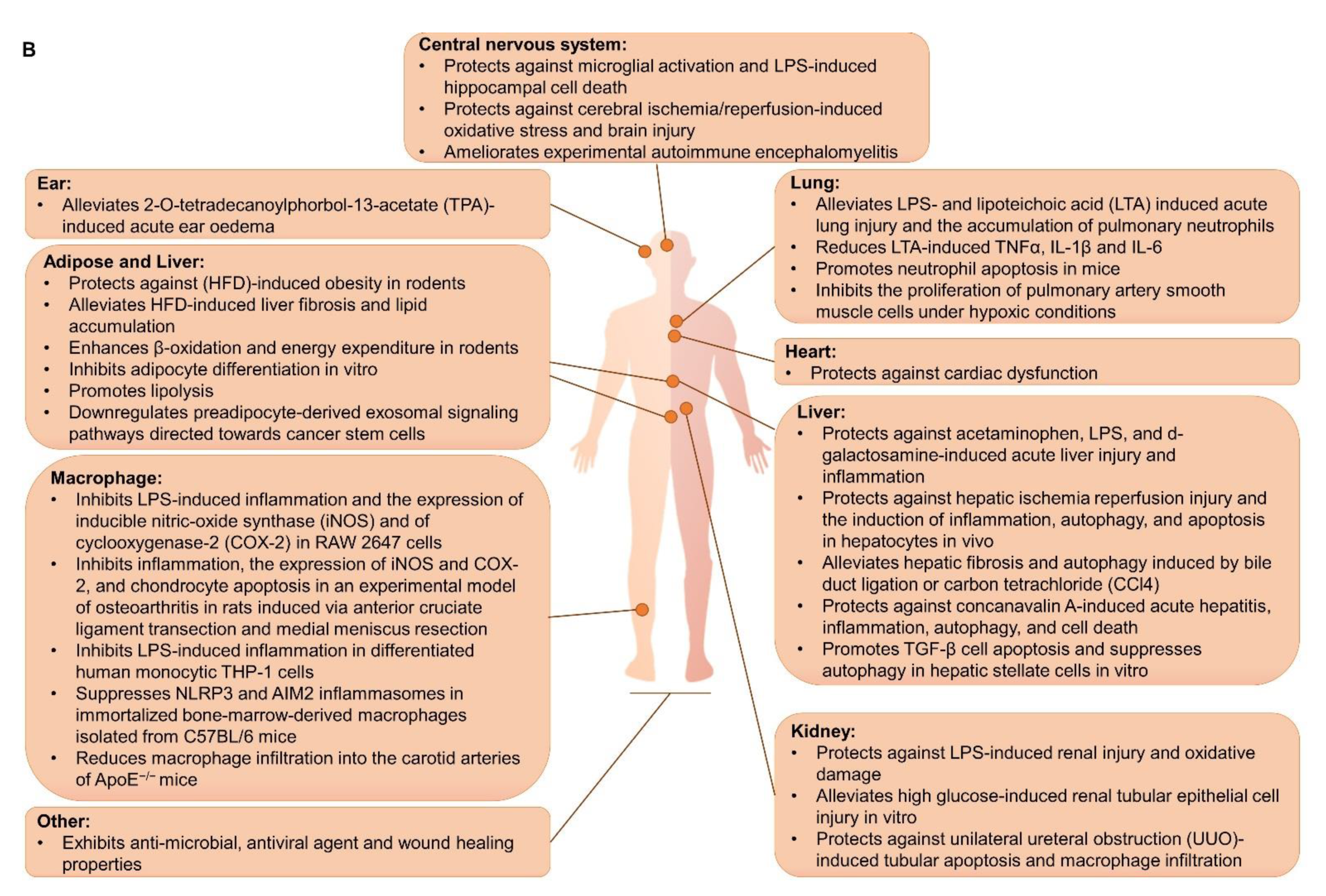{kind=link}
| miRNA | Research Model | Targeted Pathways | Effects on PKM2 Expression | Biological/Physiological Effects | Significance |
|---|---|---|---|---|---|
| miR-122 | Hepatocellular Carcinoma (HCC) | Glycolysis Proliferation Apoptosis | Decreased PKM2 mRNA and protein levels (direct targeting) | Reduction of lactate production Increased oxygen consumption [287] Decreased glucose uptake and ATP production [288] Induction of apoptosis [288,296] Reduced tumor cell proliferation [296] | Targeting PKM2 through its negative regulator miR-122 may serve as a therapeutic approach to improve chemosensitivity [288] and survival among patients with HCC [296] |
| Colon Cancer | Glycolysis | Decreased PKM2 protein levels through direct binding to PKM2 mRNA | Reduced glucose uptake and lactate production Reduced tumor cell viability and tumor volume Sensitization to 5-Fluorouracil therapy [280] | Targeting PKM2 through stabilization of miR-122 in colon cancer may enhance the effectiveness of chemotherapy in colon cancer | |
| Gallbladder Cancer | Proliferation Metastasis | Decreased PKM2 expression | Inhibits TGF-β-induced epithelium mesenchymal transformation Decreases proliferation and metastatic potential [312] | Targeting PKM2 through its negative regulator miR-122 may potentiate the effect on reducing cell invasion in gallbladder cancer | |
| miR-372 | HCC | β-catenin/Transcription Factor 4 Glycolysis | Increases the activity and expression of PKM2 | Promoted liver cancer cell cycle progression via activation of the cell cycle complex CDK2-cyclin E-P21/Cip1/WAF1. Effects are mediated through miR372-YB-1-β-catenin-LEF/TCF4-PKM2-erbB-2 axis [316] | Targeting miR-372 may be of therapeutic value for the treatment of HCC |
| miR-374b | Glycolysis | Decreased PKM2 mRNA level (indirect regulation mediated through hnRNPA1 inhibition) | Sensitization to sorafenib therapy Reduces the number of developed colonies [297] | Enhancing miR-374b expression may promote the effectiveness of chemotherapy, halt tumor growth, and increase patient survival likelihood. These effects are mediated through the suppression of hnRNPA1 and its downstream effector PKM2 | |
| miR-199a | Glycolysis | Decreased PKM2 mRNA and protein levels (direct regulation) | Decreased glucose consumption and lactate production [317] Decreased cancer cell proliferation and survival Reduced tumor growth [112] | Using natural compounds that enhance miR-199a expression to suppress PKM2 may potentiate the effectiveness of HCC treatment [317] | |
| miR-4417 | Proliferation Apoptosis | No effect on PKM2 mRNA or protein levels | Decreased expression of TRIM35 Increased PKM2 Tyr-105 phosphorylation Increased proliferation Reduced apoptosis [308] | Targeting miR-4417 may serve as an adjuvant therapy to induce apoptosis and halt HCC growth mediated by inhibiting PKM2 Tyr-105 tyrosine phosphorylation | |
| miR-199a-5p | HIF-1α | Decreased PKM2 mRNA level (indirectly by suppressing HIF-1α) | Decreased cell growth Reduced glucose uptake and lactate production [318] | Promoting miR-199a-5p expression may increase patient survival rate through suppressing the expression of HIF-1α and PKM2 | |
| miR-338-3p | Glycolysis Cell Migration | Decreased PKM2 mRNA level (direct binding leading to PKM2 mRNA degradation) | Sponging miR-338-3p mediated by circMAT2B resulted in increased glucose utilization, tumor expansion, and metastatic potential [282] | Enhancing miR-338-3p expression may reduce the progression of HCC and its metastatic potential | |
| MiR-139-5p | Gallbladder Carcinoma | Glycolysis | Decreased PKM2 mRNA and protein levels (direct regulation) | Decreased glucose consumption, lactate production, and ATP availability Reduced cell proliferation and migration [283] | Overexpressing miR-139-5p may serve as a therapeutic approach to halt GBC progression and improve GBC patient outcomes |
| MiR-let-7a | Gastric Cancer (GC) | c-Myc hnRNPA1 | Decreased PKM2 mRNA and protein levels (indirect regulation mediated through the reduction in c-Myc and hnRNPA1 expression) | Decreased cell proliferation and migration Reduced tumor size [313] | MiR-let-7a was reduced in GC tissues and enhancing its expression in GC might be capable of decreasing proliferation and metastatic potential |
| miR-124 | Colorectal Cancer (CRC) Colorectal Adenoma (CRA) | PTBP1 DDX6 | Increased PKM1 to PKM2 mRNAs ratio (indirect regulation mediated through the suppression of PTBP1) [314] | Increased oxidative stress, apoptosis, and autophagy [314] Modulation of the Warburg effect [314] | Expression of miR-124 is reduced in CRA and CRC. Its stabilization could suppress tumorigenesis through modulation of PKM1 to PKM2 ratio [314] |
| Prostate Cancer | Proliferation | Decreased PKM2 protein levels | Reduced cell proliferation [319] | May suppress tumor growth through reducing the proliferation of prostate cancer cells | |
| Pancreatic Cancer (PDAC) | Autophagy Apoptosis | Decreased PKM2 protein levels (possibly mediated through the suppression of PTBP1) | Reduced autophagy and enhanced gemcitabine-induced apoptosis [304] | miR-124/PTBP1/PKM2 axis could play a pivotal role in gemcitabine resistance and miR-124 overexpression may have therapeutic potential | |
| miR-124 miR-137 miR-340 | CRC | Glycolysis Oxidative Phosphorylation | Increased PKM1 to PKM2 mRNAs ratio. Reduced PKM2 expression through indirect regulation mediated by PTBP1, hnRNAPA1, and hnRNAPA2 | Decreased glycolysis rate and lactate production Increased oxygen consumption Inhibition of cell growth [289] | Enhancing the expression of any of the three miRNAs may increase survival rates and could exhibit antitumorigenic effects |
| miR-184 | Clear-Cell Renal Cell Carcinoma (ccRCC) | Glycolysis | Decreased PKM2 mRNA and protein levels (direct binding to PKM2 mRNA) | Reduced glucose consumption and lactate production Reduced cell proliferation [284] | The negative correlation between miR-184 and PKM2 expression in human ccRCC samples suggests that targeting miR-184 may be of therapeutic value |
| miR-152 | Breast Cancer (BC) | β-catenin Proliferation | Decreased PKM2 protein level (direct binding to PKM2 mRNA) | Reduced β-catenin Reduced cell proliferation and colony development [285] | Enhancing miR-152 may inhibit cancer cell proliferation and sensitize cancer cells to paclitaxel therapy through modulating the β-catenin pathway |
| miR-155 | PIK3R1 FOXO3a c-Myc Glycolysis | miR-155 knockout caused a reduction in PKM2 mRNA and protein levels (indirect regulation mediated through the FOXO3a/c-Myc axis) | Increased glucose utilization [299] | miR-155 may promote tumorigenesis through upregulation of PKM2 and glucose metabolism. Reducing miR-155 levels could serve as a new therapeutic approach for breast cancer | |
| miR-532-3p | Ovarian Cancer | Glycolysis DNMT3A | Decreased PKM2 protein level (an indirect regulation/unknown mechanism) | Decreased glucose consumption and lactate production [300] | Targeting miR-532-3p/PKM2 axis using glycosylated triterpenes such as 20(S)-Rg3 may prove a safe and effective therapeutic option and warrants further exploration |
| miR-29b | Glycolysis AKT2/3 | Inhibition of miR-29b increased PKM2 mRNA and protein levels (indirect regulation mediated through the activation of AKT2/3) | Reduced glucose uptake and lactate production Reduced xenograft tumor formation [301] | miR-29b may disrupt glucose metabolism and suppress epithelial ovarian cancer growth by reducing AKT2-AKT3 and PKM2 levels | |
| miR-338-3p | Glycolysis Proliferation | Decreased PKM2 mRNA and protein levels (direct binding to PKM2 mRNA) | Decreased ATP and lactate production Reduced cell proliferation [286] | Promoting miR-338-3p expression might be of therapeutic value due to its negative regulation of PKM2 in ovarian cancer cells | |
| miR-145 | Glycolysis Lactate Production Cell Growth | Decreased PKM2 protein levels (indirect mechanism mediated through upregulation of the c-Myc/miR133-b pathway) | Decreased glucose consumption and lactate production Reduced cell proliferation [320] | Promoting miR-145 expression might be a therapeutic strategy for the treatment of ovarian cancer through modulation of the Warburg effect | |
| miR-148a miR-326 | Thyroid Cancer | Cell Growth Apoptosis | Decreased PKM2 mRNA and protein levels (direct binding to PKM2 mRNA) | Reduced colony development and overall metastatic potential [290] miR-326 induced apoptosis in HeLa cells [305] | miR-148a and miR-326 may exhibit tumor-suppressive effects and reduce the metastatic potential of thyroid cancer cells [290] Identification of natural compounds that could regulate miR-326 such as resveratrol may serve as adjuvant therapy for thyroid cancer [305] |
| miR-326 | Glioma | Metabolic Activity | Decreased PKM2 protein levels through direct binding to PKM2 mRNA | Reduced ATP levels [302] | miR-326 may exhibit tumor-suppressive effects through decreased PKM2 expression |
| miR-181b | Glucose Metabolism Colony Formation | Decreased PKM2 protein levels mediated by downregulation of Specificity protein 1 (SP1) | Reduced glycolysis, proliferation, and colony number [321] | Promoting miR-181b expression might be of therapeutic potential for glioblastoma multiforme | |
| miR-214 | Non-Small-Cell Lung Cancer (NSCLC) | Glycolysis PTEN Akt mTOR | Increased PKM2 mRNA and protein levels (indirect regulation mediated through PTEN and the AKT/mTOR pathway) | Increased glucose consumption and lactate production Increased cell proliferation [295] | miR-214 may act as an oncogene. miR-214 suppression could reduce PKM2 expression and cancer growth |
| miR-625-5p | Melanoma | Glycolysis Proliferation | Decreased PKM2 mRNA and protein levels (direct regulation) | Decreased glucose consumption, ATP and lactate production Reduced cell proliferation [293] | miR-625-5p may serve as a potential target to reduce cancer growth and glycolysis in human melanoma |
| miR-133a miR-133b | Tongue Squamous Cell Carcinoma (SCC) | Proliferation Apoptosis | Decreased PKM2 protein levels (direct regulation) | Decreased cell proliferation and induction of apoptosis [292] | miR133a and miR-133b may exhibit antitumorigenic potential and could serve as a potential therapeutic strategy for SCC |
| miR-133b miR-1 | CRC | Proliferation Autophagy | Decreased PKM2 to PKM1 mRNA ratio and PKM2 protein expression levels (indirect regulation mediated through decreased PTBP1 levels) | Increased ROS Decreased lactate production Increased ATP level Induction of autophagy Reduced xenograft tumor volume [311] | Both miRNAs may have tumor-suppressive potential through their effects on PKM2 and glucose metabolism |
| miR-133b | NSCLC | Glycolysis | Decreased PKM2 protein levels (direct regulation) | Reduced glucose uptake and lactate production Sensitization of NSCLC to radiation [303] | May overcome radio-resistance and could decrease glycolysis |
| mIR-140-5p | Chronic Myeloid Leukemia | Glycolysis Proliferation Apoptosis | Decreased PKM2 protein levels (indirect regulation through modulation of sine oculis homeobox 1 (SIX1) gene) | Overexpression miR-140-5p in chronic myeloid leukemia cells inhibited cell proliferation and promoted cell apoptosis [322] | Targeting the miR-140-5p/SIX1 axis may be a potential therapeutic target for the treatment of chronic myeloid leukemia |
| miR-let-7a | Cervical Cancer | Proliferation Cell Migration Apoptosis | Decreased PKM2 protein level through direct binding to PKM2 mRNA | Decreased proliferation, metastasis, and tumor growth [291] | miR-let-7a may have tumor-suppressive potential against cervical cancer |
| miR-491-5p | Osteosarcoma | Proliferation | Decreased PKM2 mRNA and protein levels through direct binding to PKM2 mRNA | Decreased cell proliferation [315] | Targeting PKM2 through its negative regulator miR-491-5p by natural compounds such as Oviductus Ranae (OR) may reduce tumorigenesis [298] |
| HCC | Glycolysis Proliferation Cell Migration | Decreased PKM2 mRNA level through direct binding to PKM2 mRNA | Decreased glucose consumption and lactate production Reduced cell proliferation and metastasis [298] | ||
| miR-1294 | Osteosarcoma | Proliferation Cell Migration Invasion Apoptosis | Decreased PKM2 mRNA and protein levels through direct binding to PKM2 mRNA | Reduced cell proliferation, invasion, and tumor growth, while inducing apoptosis [323] | Enhancing miR-1294 expression modulates PKM2 expression to suppress tumorigenesis, which could serve as a therapeutic strategy for osteosarcoma |
| miRNA | Tissue Expression/Distribution | Research Model | Target Genes/Pathways | Effects on PKM2 Expression | Biological/Physiological Effects | Significance |
|---|---|---|---|---|---|---|
| miR-99a | Liver | Human liver cells (cancerous and noncancerous) | Glycolysis mTOR HIF-1α | Inhibited insulin-induced PKM2 expression | Regulates insulin-induced mTOR and HIF-1α expression Regulates insulin-induced glucose uptake and lactate production [340] | Insulin may inhibit miR-99a to regulate PKM2 expression, which provides a novel biological mechanism of regulating glucose metabolism in the liver that warrants further investigation |
| miR-122 | Chicken livers | Liver metabolism | Inhibition of miR-122 increased PKM2 mRNA level (direct regulation) | Inhibition of miR-122 decreased FABP5 mRNA level [331] | miR-122 may regulate aspects of liver function and homeostasis including lipid and glucose metabolism | |
| Chicken livers and hepatocytes | Autophagy Oxidative stress | Decreased PKM2 protein levels | Overexpression of miR-122 promoted autophagy and ameliorated arsenic-induced liver damage via decreasing PKM2 levels [341] | Reducing PKM2 expression through promoting miR-122 may lead to novel treatment strategies against arsenic toxicity | ||
| miR-19a-3p | Brain | Astrocytes and neurons Cerebral ischemic injury | Glycolysis Apoptosis (Bax, Caspase 3) Adipor2) | Decreased PKM2 protein levels | Decreased glycolytic enzymes, glucose consumption, and lactate production Increased markers of apoptosis [335] | miR-19a-3p may play a role in the regulation of neural cell function and could serve as a potential target against cerebral ischemic injury |
| miR-143 | Ischemic stroke Rat cortex neurons and astrocytes | Glycolysis HK2 | Decreased PKM2 mRNA and protein levels | Decreased glucose uptake and lactate production [336] | miR-143 inhibition may have neuroprotective potential during ischemic brain injury (IBI) | |
| Heart | Endothelial cells | Glycolysis HK2 LDHA | Decreased PKM2 protein levels | Decreased ATP/ADP ratio, glucose consumption, and lactate production [337] | Overexpression of miR-143 may contribute to EC dysfunction through the suppression of glycolytic activity | |
| miR-124 | Pulmonary arterial hypertension (PAH) | Proliferation Glycolysis Mitochondrial reprogramming | Decreased PKM2 mRNA levels (indirect regulation mediated through PTBP1) | Decreased glycolysis and lactate production Decreased the proliferation of blood outgrowth endothelial cells (BOECs) from patients with heritable PAH Restored mitochondrial function [338] | Targeting the miR-124-PTBP1-PKM2 axis may be of therapeutic potential for the treatment of PAH | |
| miR125a | Synovium | Psoriatic arthritis (PsA) | Glycolysis Migration Invasion | Suppression of PKM2 expression | miR-125a inhibition promoted EC tube formation, glycolysis, branching, migration, and invasion [342] | Potential therapeutic approach for the treatment of PsA |
| LncRNA | Research Model | Targeted Pathways | Mediator | Effects on PKM2 Expression | Biological/Physiological Effects | Significance |
|---|---|---|---|---|---|---|
| LncRNA MAFG-AS1 | CRC | Proliferation Glycolysis Cell cycle Apoptosis | MiR-147b | A general trend of increased PKM2 mRNA levels | Increased proliferation, cell cycle progression, and cell invasion Decreased apoptosis [346] | MAFG-AS1 may sponge miR-147b to promote tumorigenesis and may play a role in CRC progression |
| LncRNA-FEZF1-AS1 | Glycolysis | Direct binding to PKM2 | Increased PKM2 stability and protein levels through direct binding | Increased PK activity and lactate production [349] | It is upregulated in CRC and correlated with PKM2 expression. The suppression of LncRNA-FEZF1-AS1 could exhibit therapeutic potential | |
| LINC00689 | Glioma | Glycolysis Proliferation Migration Invasion | miR-338-3p | Increased PKM2 protein levels | Silencing of LINC00689 suppressed glycolysis, cell proliferation, migration, and invasion [350] | The LINC00689/miR-338-3p/PKM2 axis may have oncogenic potential and modulating this axis may prove effective in glioma therapy |
| BCYRN1 | NSCLC | Glycolysis Proliferation Invasion | miR-149 | Increased PKM2 mRNA and protein levels | Overexpression of BCYRN1 induced glycolysis, cell proliferation, and invasion [34] | Inhibiting BCYRN1 or/and AC020978 might be a therapeutic target for NSCLC |
| AC020978 | Glycolysis Proliferation | Increased PKM2 protein levels and stability through direct binding | AC020978 induced glycolysis and proliferation partially through enhancing PKM2 levels and its transactivation capability on HIF-1α [355] | |||
| LINC01554 | Liver Cancer | Glycolysis Cell growth Akt/mTOR | Decreased PKM2 protein levels (promotes ubiquitin-mediated degradation of PKM2) | Inhibited Akt/mTOR signaling to reduce glycolysis Reduced cell growth and colony formation [347] | LINC01554 may exhibit tumor suppressor activity and could serve as a prognostic biomarker | |
| lncRNA MEG3 | Epithelial to mesenchymal transition (EMT) Wound healing | Negatively correlates with PKM2 mRNA and protein levels | Arsenic trioxide inhibited HCC migration and EMT through promoting MEG3 and reducing PKM2 [66] | MEG3 may disrupt metastatic potential after arsenic trioxide treatment and may exhibit beneficial effects through suppressing PKM2 expression | ||
| Cell growth | miR-122 | Decreased PKM2 protein levels | Promoted the expression of miR-122 to downregulate PKM2 expression and its nuclear translocation Reduced β-catenin, cell proliferation, and tumor growth [89] | MEG3 may act as a tumor suppressor with potential in prognostic and therapeutic clinical application | ||
| lncRNA HULC | Autophagy PTEN AKT-PI3K-mTOR pathway | Increased PKM2 protein levels | Increased LC3II and Sirt1, resulting in elevated levels of autophagy Increased cell growth [354] | HULC may play a critical role in the progression of hepatocarcinogenesis | ||
| lncRNA-SOX2OT | Glycolysis Metastasis | miR-122-5p | Increased PKM2 protein levels | Increased glucose metabolism and PKM2 expression and exacerbates the metastatic potential of HCC [356] | Suppressing SOX2OT might be of therapeutic value to halt HCC metastasis | |
| Linc-ROR | PDAC | Autophagy Apoptosis PTBP1 | miR-124 | Silencing ROR decreased PKM2 protein levels | Overexpression of ROR increased autophagy Silencing ROR enhanced gemcitabine-induced apoptosis [304] | Linc-ROR may sponge miR-124 (a negative regulator of PTBP1) to increase PKM2 levels and confer chemoresistance |
| LncRNA XLOC_006390 | Cervical Cancer | Apoptosis Cell migration | miR-338-3p | Silencing XLOC_006390 decreased PKM2 mRNA and protein levels | XLOC_006390 may act as a negative regulator of miR-331-3p and miR-338-3p Promoted tumorigenesis and metastasis [353] | XLOC_006390 may hold value as a marker for cervical cancer with potential therapeutic targeting application |
| LncRNA RPPH1 | Breast Cancer | Cell cycle Proliferation Colony formation | miR-122 | Increased PKM2 mRNA levels | Promoted proliferation, colonization, and cell cycle progression [352] | RPPH1 may act in an oncogenic manner and its suppression could possess therapeutic potential |
| Alzheimer’s Disease | Apoptosis Endoplasmic reticulum (ER) stress | miR-326 | Increased PKM2 protein level | Ameliorated amyloid β (Aβ) induced ER stress and apoptosis in neuroblastoma cells [357] | RPPH1 might be of therapeutic potential for future Alzheimer’s treatment options | |
| TP53TG1 | PDAC Gliomas/Brain Tumor Cells | KRAS Proliferation Colony formation Glycolysis Proliferation Migration | miR-96 ND | ND Decreased PKM2 mRNA levels under high and low glucose levels | Promoted cell proliferation and invasion and inhibited apoptotic cell death [358] Promoted cell proliferation and migration and inhibited apoptotic cell death [359] | TP53TG1 may inhibit the Warburg effect and halt cancer cell growth and metastasis |
| HOXB-AS3 | CRC | Glycolysis PKM splicing miR-18a processing | hnRNP A1 | Inhibition of hnRNP A1-dependent PKM splicing and PKM2 expression. Effects are mediated through the HOXB-AS3 peptide and not the LncRNA. | HOXB-AS3 peptide inhibits tumorigenesis and metabolic reprogramming in CRC cells [360] | HOXB-AS3 and SNHG6 might be viable targets for disrupting CRC metabolism and tumor growth |
| SNHG6 | Glycolysis PKM splicing | Knockdown of SNHG6 decreased PKM2/M1 ratio | SNHG6 promotes hnRNPA1 to favor PKM2 expression and the subsequent increase in glycolysis [361] | |||
| LncRNA H19 | Ovarian Cancer | Glycolysis | miR-324-5p | Increased PKM2 protein level | Increased glucose consumption and lactate production [351] | 20(S)-Rg3 may inhibit the Warburg effect and halt cancer cell growth through the H19/miR-324-5p/PKM2 axis |
| NSCLC | EGFR AKT | Decreased PKM2 protein levels by promoting PKM2 ubiquitin mediated degradation | Suppression of H19 exacerbated erlotinib resistance (an EGFR inhibitor) by increasing PKM2, and the subsequent activation of AKT [362] | Targeting H19/PKM2 axis might be a viable option to overcome erlotinib resistance | ||
| LINC00504 | Glycolysis Proliferation Apoptosis | miR-1244 | Increased PKM2 mRNA and protein levels | LINC00504 knockdown suppressed proliferation and glycolysis, but induced apoptosis [363] | Inhibiting LINC00504 may exhibit therapeutic potential for ovarian cancer treatment | |
| LincRNA-p21 | Prostate Cancer | Glycolysis Proliferation Tumorigenesis | Decreased PKM2 mRNA and protein levels through indirect regulation mediated by PTEN/AKT/mTOR pathway | Reduced proliferation and tumorigenic potential Its downregulation increased glucose consumption, lactate production, and pyruvate levels [348] | LincRNA-p21 may be a viable target for disrupting PC metabolism and tumor growth |
| LncRNA | Research Model | Targeted pathways | Mediator | Effects on PKM2 Expression | Biological/physiological Effects | Significance |
|---|---|---|---|---|---|---|
| LncRNA RPPH1 | Alzheimer’s disease (AD) | Endoplasmic reticulum stress (ER stress) Apoptosis | miR-326 | Not determined (a possible increase through blocking the role of MiR-326 in downregulating PKM2 expression) | RPPH1 overexpression decreased β25-35-induced apoptosis in SH-SY5Y cells through downregulating ER stress and modulating PKM2 activity [357] | RPPH1 acts as a molecular sponge for miR-326. LncRNA RPPH1 could attenuate ERS and apoptosis in neurodegenerative disorders |
| LncRNA-Malat1 | Type 2 diabetes (T2D) | Glucose-stimulated insulin secretion Cell death | Ptbp1 (Increased) | Silencing malat1 leads to β-cell dysfunction and alterations in glucose-stimulated insulin secretion | LncRNA-Malat1 enhances Ptbp1 stability and PKM2 expression [365] | LncRNA-Malat1 could possibly serve as a potential therapeutic target for T2D through modulation of PKM2 activity |
| CircRNA | Research Model | Targeted Pathways | Mediator | Effects on PKM2 Expression | Biological/Physiological Effects | Significance |
|---|---|---|---|---|---|---|
| Circ-MAT2B | Hepatocellular Carcinoma (HCC) | Glycolysis Cell proliferation Invasion | miR-338-3p | Overexpression of Circ-MAT2B increased protein and mRNA PKM2 levels | Circ-MAT2B overexpression increases HCC glucose utilization, tumor growth, and metastasis in vivo Circ-MAT2B overexpression promoted glycolysis, cell proliferation, migration, and invasion in vitro under hypoxia [282] | Circ-MAT2B is associated with and predicts poor prognosis of HCC Targeting circ-MAT2B may alleviate HCC burden |
| Circ-FOXM1 | Melanoma | Proliferation Glycolysis Invasion Apoptosis | miR-143-3p | Silencing CircFOXM1 decreased PKM2 protein levels | Increased proliferation, glycolysis, motility, and decreased apoptosis [374] | Circ-FOXM1 may promote melanoma progression through the miR-143-3p/FLOT2 axis |
| Circ-NRIP1 | Gastric Cancer | Proliferation Migration Glycolysis Apoptosis | miR-186-5p | Silencing of circ-NRIP1 decreased PKM2 protein levels | Circ-NRIP1 KD decreased proliferation, migration, and glycolysis but induced apoptosis [375] | Circ-NRIP1 promoted carcinogenesis and may have potential in prognostic and clinical application |
| Circ-FOXP1 | Gallbladder Cancer | Proliferation Migration Invasion Apoptosis Glycolysis | miR-370 | Silencing of circFOXP1 resulted in a partial reduction in PKM2 protein level | Promotes proliferation, invasion, migration and decreases apoptosis [376] | Circ-FOXP1 may sponge miR-370 and promote PKLR expression, enhancing tumor progression |
| Circ-RNA hsa_circ_0005963 (ciRS-122) | CRC | Glycolysis Apoptosis | miR-122 | Exosome delivery of CiRS-122 upregulated PKM2 levels | CiRS-122 delivery may promote drug resistance and glycolysis, and exosome-delivered siRNA appeared to reverse the resistance to treatment [377] | CiRS-122 silencing may promote enhanced therapeutic effectiveness against oxaliplatin-resistant CRC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puckett, D.L.; Alquraishi, M.; Chowanadisai, W.; Bettaieb, A. The Role of PKM2 in Metabolic Reprogramming: Insights into the Regulatory Roles of Non-Coding RNAs. Int. J. Mol. Sci. 2021, 22, 1171. https://doi.org/10.3390/ijms22031171
Puckett DL, Alquraishi M, Chowanadisai W, Bettaieb A. The Role of PKM2 in Metabolic Reprogramming: Insights into the Regulatory Roles of Non-Coding RNAs. International Journal of Molecular Sciences. 2021; 22(3):1171. https://doi.org/10.3390/ijms22031171
Chicago/Turabian StylePuckett, Dexter L., Mohammed Alquraishi, Winyoo Chowanadisai, and Ahmed Bettaieb. 2021. "The Role of PKM2 in Metabolic Reprogramming: Insights into the Regulatory Roles of Non-Coding RNAs" International Journal of Molecular Sciences 22, no. 3: 1171. https://doi.org/10.3390/ijms22031171
APA StylePuckett, D. L., Alquraishi, M., Chowanadisai, W., & Bettaieb, A. (2021). The Role of PKM2 in Metabolic Reprogramming: Insights into the Regulatory Roles of Non-Coding RNAs. International Journal of Molecular Sciences, 22(3), 1171. https://doi.org/10.3390/ijms22031171