
Expression of the SARS-CoV-2 Receptor ACE2 and Proinflammatory Cytokines Induced by the Periodontopathic Bacterium Fusobacterium nucleatum in Human Respiratory Epithelial Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
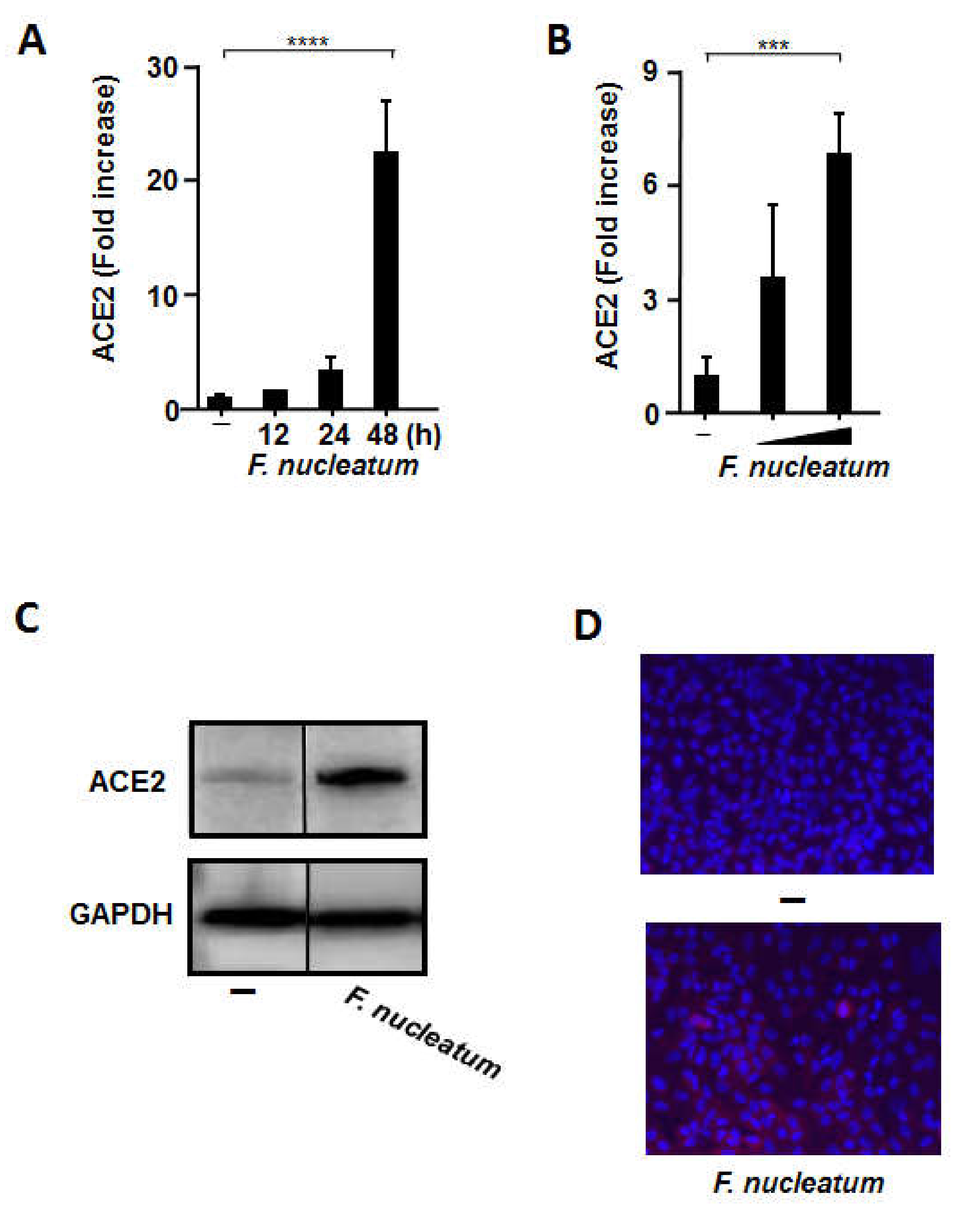
2.1. CSF Stimulated ACE2 Expression in Human Alveolar Epithelial Cells
2.2. CSF Induced IL-8 and IL-6 mRNA Expression and Protein Production by Human Alveolar Epithelial Cells
2.3. CSF Induced IL-8 and IL-6 Production by Human Primary Alveolar Epithelial Cells
2.4. CSF Induced IL-8 and IL-6 Production by Human Bronchial Epithelial Cells and Human Pharyngeal Epithelial Cells
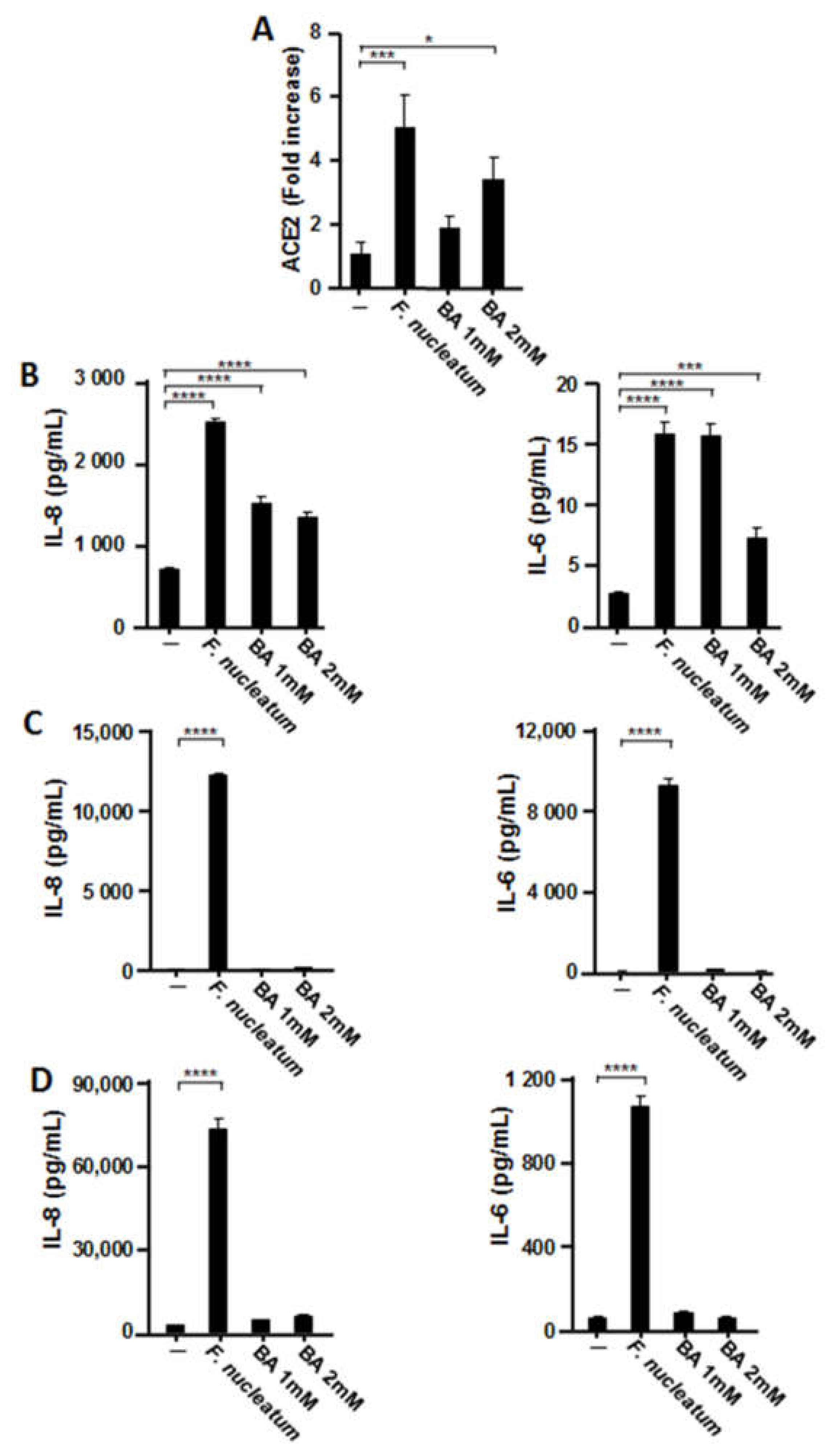
2.5. Effects of Butyric Acid (BA) on the Expression of ACE2 and Proinflammatory Cytokines in Human Respiratory Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Primary Human Alveolar Epithelial Cell Culture
4.3. Bacterial Strains and Culture
4.4. mRNA Preparation and RT-qPCR Assay
4.5. Western Blot Analysis
4.6. Immunofluorescence
4.7. IL-8 and IL-6 Measurements
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune Cross-Reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. HLH Across Speciality Collaboration, UK COVID-19: Consider Cytokine Storm Syndromes and Immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Predicts COVID-19 Severity and Survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Herold, T.; Jurinovic, V.; Arnreich, C.; Lipworth, B.J.; Hellmuth, J.C.; von Bergwelt-Baildon, M.; Klein, M.; Weinberger, T. Elevated Levels of IL-6 and CRP Predict the Need for Mechanical Ventilation in COVID-19. J. Allergy Clin. Immunol. 2020, 146, 128–136.e4. [Google Scholar] [CrossRef]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal Diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal Diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72 314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Wang, B.; Li, R.; Lu, Z.; Huang, Y. Does Comorbidity Increase the Risk of Patients with COVID-19: Evidence from Meta-Analysis. Aging 2020, 12, 6049–6057. [Google Scholar]
- Yamasaki, K.; Kawanami, T.; Yatera, K.; Fukuda, K.; Noguchi, S.; Nagata, S.; Nishida, C.; Kido, T.; Ishimoto, H.; Taniguchi, H.; et al. Significance of Anaerobes and Oral Bacteria in Community-Acquired Pneumonia. PLoS ONE 2013, 8, e63103. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, K.; Yanagihara, K.; Harada, Y.; Yamada, K.; Migiyama, Y.; Morinaga, Y.; Izumikawa, K.; Kohno, S. Quantitative Detection of Periodontopathic Bacteria in Lower Respiratory Tract Specimens by Real-Time PCR. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2017, 23, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.G.; Finegold, S.M. Anaerobic Infections of the Lung and Pleural Space. Am. Rev. Respir. Dis. 1974, 110, 56–77. [Google Scholar] [CrossRef] [PubMed]
- Awano, S.; Ansai, T.; Takata, Y.; Soh, I.; Akifusa, S.; Hamasaki, T.; Yoshida, A.; Sonoki, K.; Fujisawa, K.; Takehara, T. Oral Health and Mortality Risk from Pneumonia in the Elderly. J. Dent. Res. 2008, 87, 334–339. [Google Scholar] [CrossRef]
- De Melo Neto, J.P.; Melo, M.S.; dos Santos-Pereira, S.A.; Martinez, E.F.; Okajima, L.S.; Saba-Chujfi, E. Periodontal Infections and Community-Acquired Pneumonia: A Case-Control Study. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2013, 32, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Matsumoto, K.; Furuta, M.; Fukuyama, S.; Takeshita, T.; Ogata, H.; Suma, S.; Shibata, Y.; Shimazaki, Y.; Hata, J.; et al. Periodontitis Is Associated with Chronic Obstructive Pulmonary Disease. J. Dent. Res. 2019, 98, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Ochiai, K.; Okamoto, T. Reactivation of Latent HIV-1 Infection by the Periodontopathic Bacterium Porphyromonas gingivalis Involves Histone Modification. J. Immunol. 2009, 182, 3688–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.; Yamada, K.; Tamura, M.; Ochiai, K.; Okamoto, T. Reactivation of Latent HIV-1 by a Wide Variety of Butyric Acid-Producing Bacteria. Cell Mol. Life Sci. 2012, 69, 2583–2592. [Google Scholar] [CrossRef]
- Imai, K.; Inoue, H.; Tamura, M.; Cueno, M.E.; Inoue, H.; Takeichi, O.; Kusama, K.; Saito, I.; Ochiai, K. The Periodontal Pathogen Porphyromonas gingivalis Induces the Epstein–Barr Virus Lytic Switch Transactivator ZEBRA by Histone Modification. Biochimie 2012, 94, 839–846. [Google Scholar]
- Yoneyama, T.; Yoshida, M.; Matsui, T.; Sasaki, H. Oral Care and Pneumonia. Oral Care Working Group. Lancet 1999, 354, 515. [Google Scholar]
- Abe, S.; Ishihara, K.; Adachi, M.; Sasaki, H.; Tanaka, K.; Okuda, K. Professional Oral Care Reduces Influenza Infection in Elderly. Arch. Gerontol. Geriatr. 2006, 43, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Han, J.; Liu, Z.; Song, Y.; Wang, Z.; Sun, Z. Effects of Periodontal Treatment on Lung Function and Exacerbation Frequency in Patients with Chronic Obstructive Pulmonary Disease and Chronic Periodontitis: A 2-Year Pilot Randomized Controlled Trial. J. Clin. Periodontol. 2014, 41, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, S.; Nitta, H.; Nagasawa, T.; Uchimura, I.; Izumiyama, H.; Inagaki, K.; Kikuchi, T.; Noguchi, T.; Kanazawa, M.; Matsuo, A.; et al. Multi-Center Intervention Study on Glycohemoglobin (HbA1C) and Serum, High-Sensitivity CRP (hs-CRP) After Local Anti-Infectious Periodontal Treatment in type 2 Diabetic Patients with Periodontal Disease. Diabetes Res. Clin. Pr. 2009, 83, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Simpson, T.C.; Weldon, J.C.; Worthington, H.V.; Needleman, I.; Wild, S.H.; Moles, D.R.; Stevenson, B.; Furness, S.; Iheozor-Ejiofor, Z. Treatment of Periodontal Disease for Glycaemic Control in People with Diabetes Mellitus. Cochrane Database Syst. Rev. 2015, 2015, CD004714. [Google Scholar] [CrossRef]
- Shen, Z.; Xiao, Y.; Kang, L.; Ma, W.; Shi, L.; Zhang, L.; Zhou, Z.; Yang, J.; Zhong, J.; Yang, D.; et al. Genomic Diversity of Severe Acute Respiratory Syndrome-Coronavirus 2 in Patients with Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 713–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riggs, M.G.; Whittaker, R.G.; Neumann, J.R.; Ingram, V.M. n-Butyrate Causes Histone Modification in HeLa and Friend Erythroleukaemia Cells. Nature 1977, 268, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Towatari, M.; Kosugi, H.; Saito, H. Up-Regulation of costimulatory/Adhesion Molecules by Histone Deacetylase Inhibitors in Acute Myeloid Leukemia Cells. Blood 2000, 96, 3847–3856. [Google Scholar] [PubMed]
- Long, C.W.; Suk, W.A.; Snead, R.M.; Christensen, W.L. Cell Cycle-Specific Enhancement of type C Virus Activation by Sodium n-Butyrate. Cancer Res. 1980, 40, 3886–3890. [Google Scholar]
- Van der Maarel-Wierink, C.D.; Vanobbergen, J.N.; Bronkhorst, E.M.; Schols, J.M.; de Baat, C. Oral Health Care and Aspiration Pneumonia in Frail Older People: A Systematic Literature Review. Gerodontology 2013, 30, 3–9. [Google Scholar] [CrossRef]
- Terpenning, M.S.; Taylor, G.W.; Lopatin, D.E.; Kerr, C.K.; Dominguez, B.L.; Loesche, W.J. Aspiration Pneumonia: Dental and Oral Risk Factors in an Older Veteran Population. J. Am. Geriatr. Soc. 2001, 49, 557–563. [Google Scholar] [CrossRef]
- Marik, P.E.; Kaplan, D. Aspiration Pneumonia and Dysphagia in the Elderly. Chest 2003, 124, 328–336. [Google Scholar] [CrossRef]
- Ebihara, S.; Ebihara, T.; Kohzuki, M. Effect of Aging on Cough and Swallowing Reflexes: Implications for Preventing Aspiration Pneumonia. Lung 2012, 190, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Clayton, N.A.; Carnaby, G.D.; Peters, M.J.; Ing, A.J. Impaired Laryngopharyngeal Sensitivity in Patients with COPD: The Association with Swallow Function. Int. J. Speech Lang. Pathol. 2014, 16, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.C.; Sausville, E.L.; Girish, V.; Yuan, M.L.; Vasudevan, A.; John, K.M.; Sheltzer, J.M. Cigarette Smoke Exposure and Inflammatory Signaling Increase the Expression of the SARS-CoV-2 Receptor ACE2 in the Respiratory Tract. Dev. Cell 2020, 53, 514–529.e3. [Google Scholar] [PubMed]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets Across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Okuda, K.; Kimizuka, R.; Abe, S.; Kato, T.; Ishihara, K. Involvement of Periodontopathic Anaerobes in Aspiration Pneumonia. J. Periodontol. 2005, 76, 2154–2160. [Google Scholar] [PubMed]
- Nagaoka, K.; Yanagihara, K.; Morinaga, Y.; Nakamura, S.; Harada, T.; Hasegawa, H.; Izumikawa, K.; Ishimatsu, Y.; Kakeya, H.; Nishimura, M.; et al. Prevotella intermedia Induces Severe Bacteremic Pneumococcal Pneumonia in Mice with Upregulated Platelet-Activating Factor Receptor Expression. Infect. Immun. 2014, 82, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Scannapieco, F.A. Role of Oral Bacteria in Respiratory Infection. J. Periodontol. 1999, 70, 793–802. [Google Scholar] [CrossRef]
- Baggiolini, M.; Walz, A.; Kunkel, S.L. Neutrophil-Activating peptide-1/interleukin 8, a Novel Cytokine That Activates Neutrophils. J. Clin. Investig. 1989, 84, 1045–1049. [Google Scholar] [CrossRef]
- Strieter, R.M.; Belperio, J.A.; Keane, M.P. Host Innate Defenses in the Lung: The Role of Cytokines. Curr. Opin. Infect. Dis. 2003, 16, 193–198. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective Treatment of Severe COVID-19 Patients with Tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar]
- Hayata, M.; Watanabe, N.; Tamura, M.; Kamio, N.; Tanaka, H.; Nodomi, K.; Miya, C.; Nakayama, E.; Ueda, K.; Ogata, Y.; et al. The Periodontopathic Bacterium Fusobacterium nucleatum Induced Proinflammatory Cytokine Production by Human Respiratory Epithelial Cell Lines and in the Lower Respiratory Organs in Mice. Cell Physiol. Biochem. 2019, 53, 49–61. [Google Scholar]
- Watanabe, N.; Yokoe, S.; Ogata, Y.; Sato, S.; Imai, K. Exposure to Porphyromonas gingivalis Induces Production of Proinflammatory Cytokine via TLR2 from Human Respiratory Epithelial Cells. J. Clin. Med. 2020, 9, 3433. [Google Scholar] [CrossRef]
- Koike, R.; Cueno, M.E.; Nodomi, K.; Tamura, M.; Kamio, N.; Tanaka, H.; Kotani, A.; Imai, K. Heat-Killed Fusobacterium nucleatum Triggers Varying Heme-Related Inflammatory and Stress Responses Depending on Primary Human Respiratory Epithelial Cell Type. Molecules 2020, 25, 3839. [Google Scholar] [CrossRef]
- Hsueh, P.R.; Chen, P.J.; Hsiao, C.H.; Yeh, S.H.; Cheng, W.C.; Wang, J.L.; Chiang, B.L.; Chang, S.C.; Chang, F.Y.; Wong, W.W.; et al. SARS Research Group of National Taiwan University College of Medicine and National Taiwan University Hospital Patient Data, Early SARS Epidemic, Taiwan. Emerg. Infect. Dis. 2004, 10, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical Course and Risk Factors for Mortality of Adult Inpatients with COVID-19 in Wuhan, China: A Retrospective Cohort Study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical Course and Outcomes of Critically Ill Patients with SARS-CoV-2 Pneumonia in Wuhan, China: A Single-Centered, Retrospective, Observational Study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Margolis, H.C.; Duckworth, J.H.; Moreno, E.C. Composition and Buffer Capacity of Pooled Starved Plaque Fluid from Caries-Free and Caries-Susceptible Individuals. J. Dent. Res. 1988, 67, 1476–1482. [Google Scholar]
- Moreno, E.C.; Margolis, H.C. Composition of Human Plaque Fluid. J. Dent. Res. 1988, 67, 1181–1189. [Google Scholar]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.Y.; Robinson, P.; Naleway, C. Short-Chain Carboxylic Acid Concentration in Human Gingival Crevicular Fluid. J. Dent. Res. 1997, 76, 575–579. [Google Scholar] [CrossRef]
- Koike, R.; Nodomi, K.; Watanabe, N.; Ogata, Y.; Takeichi, O.; Takei, M.; Kaneko, T.; Tonogi, M.; Kotani, A.I.; Imai, K. Butyric Acid in Saliva of Chronic Periodontitis Patients Induces Transcription of the EBV Lytic Switch Activator BZLF1: A Pilot Study. In Vivo 2020, 34, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Watanabe, N.; Kamio, N.; Kobayashi, R.; Iinuma, T.; Imai, K. Aspiration of Periodontopathic Bacteria Due to Poor Oral Hygiene Potentially Contributes to the Aggravation of COVID-19. J. Oral Sci. 2020, 63, 1–3. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, Y.; Watanabe, N.; Kamio, N.; Yokoe, S.; Suzuki, R.; Sato, S.; Iinuma, T.; Imai, K. Expression of the SARS-CoV-2 Receptor ACE2 and Proinflammatory Cytokines Induced by the Periodontopathic Bacterium Fusobacterium nucleatum in Human Respiratory Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 1352. https://doi.org/10.3390/ijms22031352
Takahashi Y, Watanabe N, Kamio N, Yokoe S, Suzuki R, Sato S, Iinuma T, Imai K. Expression of the SARS-CoV-2 Receptor ACE2 and Proinflammatory Cytokines Induced by the Periodontopathic Bacterium Fusobacterium nucleatum in Human Respiratory Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(3):1352. https://doi.org/10.3390/ijms22031352
Chicago/Turabian StyleTakahashi, Yuwa, Norihisa Watanabe, Noriaki Kamio, Sho Yokoe, Ryuta Suzuki, Shuichi Sato, Toshimitsu Iinuma, and Kenichi Imai. 2021. "Expression of the SARS-CoV-2 Receptor ACE2 and Proinflammatory Cytokines Induced by the Periodontopathic Bacterium Fusobacterium nucleatum in Human Respiratory Epithelial Cells" International Journal of Molecular Sciences 22, no. 3: 1352. https://doi.org/10.3390/ijms22031352
APA StyleTakahashi, Y., Watanabe, N., Kamio, N., Yokoe, S., Suzuki, R., Sato, S., Iinuma, T., & Imai, K. (2021). Expression of the SARS-CoV-2 Receptor ACE2 and Proinflammatory Cytokines Induced by the Periodontopathic Bacterium Fusobacterium nucleatum in Human Respiratory Epithelial Cells. International Journal of Molecular Sciences, 22(3), 1352. https://doi.org/10.3390/ijms22031352