
Role of CgTpo4 in Polyamine and Antimicrobial Peptide Resistance: Determining Virulence in Candida glabrata

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
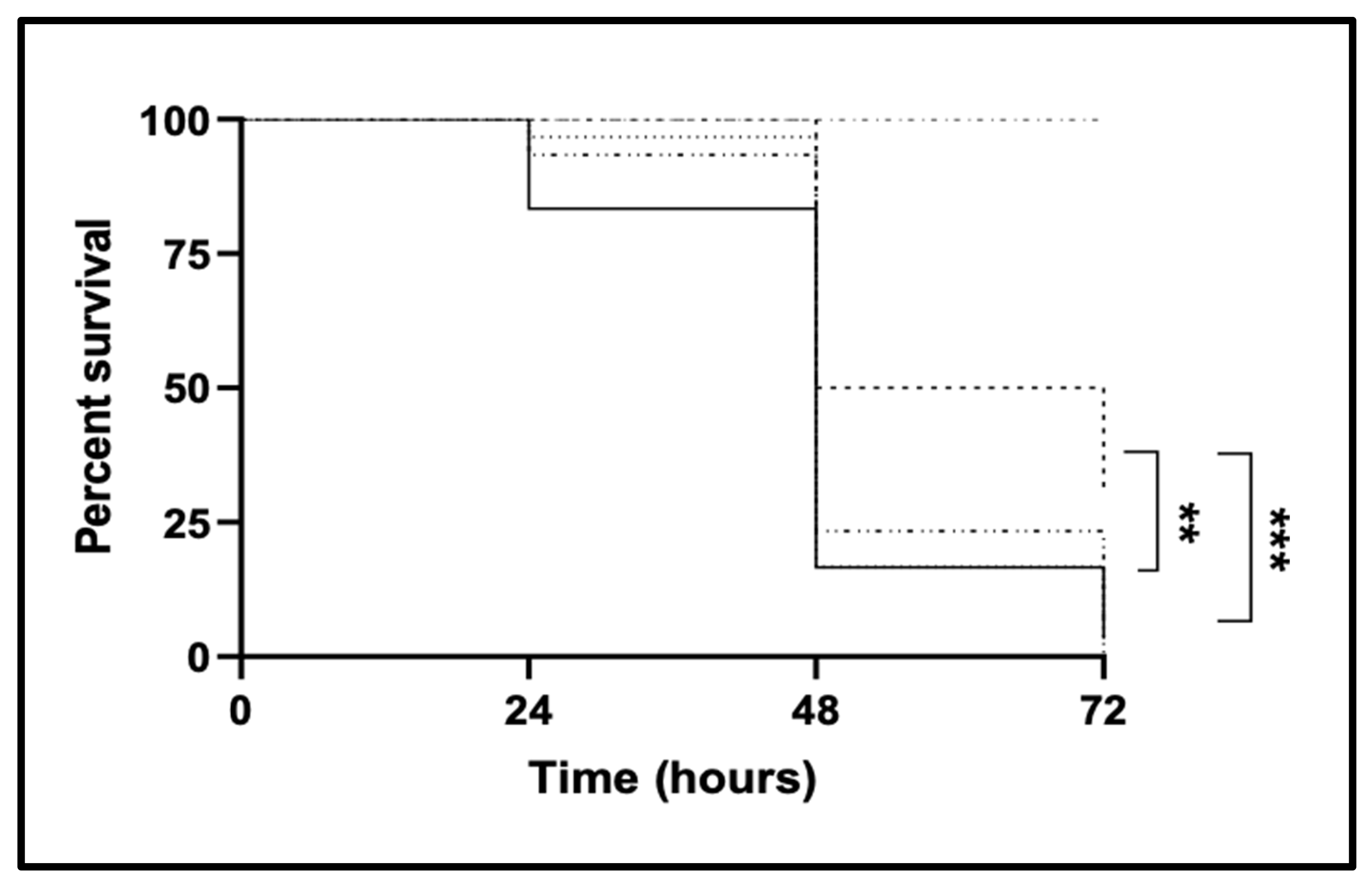
2.1. CgTPO4 Is a Determinant of C. glabrata Virulence against the G. mellonella Infection Model
2.2. CgTPO4 Confers Resistance to the Human Antimicrobial Peptide Histatin-5, But Not to Phagocytosis
2.3. Antimicrobial Peptide Gene Expression Is Highly Activated in G. mellonella Larvae in Response to Infection by C. glabrata
2.4. CgTPO4 Is a Determinant of Polyamine Resistance in C. glabrata
2.5. CgTpo4 Is a Plasma Membrane Polyamine Exporter
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids and Growth Medium
4.2. Cloning of the C. glabrata CgTPO4 Gene (ORF CAGL0L10912g)
4.3. Disruption of C. glabrata CgURA3 (ORF CAGL0I03080g)
4.4. Galleria Mellonella Survival Assays
4.5. Galleria Mellonella Hemocyte–Yeast Interaction Assays
4.6. Gene Expression Measurement
4.7. Candidacidal Assays of Histatin-5
4.8. Subcellular Localization of the CgTpo4 Transporter
4.9. Susceptibility Assays in C. glabrata
4.10. Spermidine Accumulation Assays
4.11. Statistical Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanguinetti, M.; Posteraro, B.; Lass-Flörl, C. Antifungal drug resistance among Candida species: Mechanisms and clinical impact. Mycoses 2015, 58, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Garcia-Effron, G.; Lass-Florl, C.; Lopez, A.G.; Rodriguez-Tudela, J.-L.; Cuenca-Estrella, M.; Perlin, D.S. Echinocandin Susceptibility Testing of Candida Species: Comparison of EUCAST EDef 7.1, CLSI M27-A3, Etest, Disk Diffusion, and Agar Dilution Methods with RPMI and IsoSensitest Media. Antimicrob. Agents Chemother. 2010, 54, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Tscherner, M.; Schwarzmüller, T.; Kuchler, K. Pathogenesis and Antifungal Drug Resistance of the Human Fungal Pathogen Candida glabrata. Pharmaceuticals 2011, 4, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Bromley, M.J. How to bolster the antifungal pipeline. Science 2015, 347, 1414–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.; Prasad, T.; Sharma, N.; Payasi, A.; Prasad, R.; Gupta, D.; Singh, R. Pathogenicity and drug resistance in Candida albicans and other yeast species. Acta Microbiol. Immunol. Hung. 2007, 54, 201–235. [Google Scholar] [CrossRef]
- Kasper, L.; Seider, K.; Hube, B. Intracellular survival of Candida glabrata in macrophages: Immune evasion and persistence. FEMS Yeast Res. 2015, 15, fov042. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Shirtliff, M.E.; Jabra-Rizk, M.A. Antimicrobial peptides: Primeval molecules or future drugs? PLoS Pathog. 2010, 6, e1001067. [Google Scholar] [CrossRef]
- Tam, P.; Gee, K.; Piechocinski, M.; Macreadie, I. Candida glabrata, Friend and Foe. J. Fungi 2015, 1, 277–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory Circuitry Governing Fungal Development, Drug Resistance, and Disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, P.; Costa, C.; Pires, C.; Shimizu, K.; Chibana, H.; Teixeira, M.C. Membrane Proteome-Wide Response to the Antifungal Drug Clotrimazole in Candida glabrata: Role of the Transcription Factor CgPdr1 and the Drug:H+ Antiporters CgTpo1_1 and CgTpo1_2. Mol. Cell. Proteom. 2016, 15, 57–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, C.; Dias, P.J.; Sá-Correia, I.; Teixeira, M.C. MFS multidrug transporters in pathogenic fungi: Do they have real clinical impact? Front. Physiol. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomitori, H.; Kashiwagi, K.; Asakawa, T.; Kakinuma, Y.; Michael, A.J.; Igarashi, K. Multiple polyamine transport systems on the vacuolar membrane in yeast. Biochem. J. 2001, 353, 681–688. [Google Scholar] [PubMed]
- Chen, K.-H.H.; Miyazaki, T.; Tsai, H.-F.F.; Bennett, J.E. The bZip transcription factor Cgap1p is involved in multidrug resistance and required for activation of multidrug transporter gene CgFLR1 in Candida glabrata. Gene 2007, 386, 63–72. [Google Scholar] [CrossRef]
- Costa, C.; Nunes, J.; Henriques, A.; Mira, N.P.; Nakayama, H.; Chibana, H.; Teixeira, M.C. Candida glabrata drug:H+ antiporter CgTpo3 (ORF CAGL0I10384g): Role in azole drug resistance and polyamine homeostasis. J. Antimicrob. Chemother. 2014, 69, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Costa, C.; Henriques, A.; Pires, C.; Nunes, J.; Ohno, M.; Chibana, H.; Sá-Correia, I.; Teixeira, M.C. The dual role of Candida glabrata drug:H+ antiporter CgAqr1 (ORF CAGL0J09944g) in antifungal drug and acetic acid resistance. Front. Microbiol. 2013, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pais, P.; Pires, C.; Costa, C.; Okamoto, M.; Chibana, H.; Teixeira, M.C. Membrane proteomics analysis of the Candida glabrata response to 5-flucytosine: Unveiling the role and regulation of the drug efflux transporters CgFlr1 and CgFlr2. Front. Microbiol. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Romão, D.; Cavalheiro, M.; Mil-Homens, D.; Santos, R.; Pais, P.; Costa, C.; Takahashi-Nakaguchi, A.; Fialho, A.M.; Chibana, H.; Teixeira, M.C. A New Determinant of Candida glabrata Virulence: The Acetate Exporter CgDtr1. Front. Cell. Infect. Microbiol. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Becker, J.M.; Henry, L.K.; Jiang, W.; Koltin, Y. Reduced virulence of Candida albicans mutants affected in multidrug resistance. Infect. Immun. 1995, 63, 4515–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.H.; Singh, A.; Dhamgaye, S.; Chauhan, N.; Vandeputte, P.; Suneetha, K.J.; Kaur, R.; Mukherjee, P.K.; Chandra, J.; Ghannoum, M.; et al. Novel role of a family of major facilitator transporters in biofilm development and virulence of Candida albicans. Biochem. J. 2014, 460, 223–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada-Okabe, T.; Yamada-Okabe, H. Characterization of the CaNAG3, CaNAG4, and CaNAG6 genes of the pathogenic fungus Candida albicans: Possible involvement of these genes in the susceptibilities of cytotoxic agents. FEMS Microbiol. Lett. 2002, 212, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Costa, C.; Mil-Homens, D.; Romão, D.; de Carvalho, C.C.C.R.; Pais, P.; Mira, N.P.; Fialho, A.M.; Teixeira, M.C. The multidrug resistance transporters CgTpo1_1 and CgTpo1_2 play a role in virulence and biofilm formation in the human pathogen Candida glabrata. Cell. Microbiol. 2017, 19, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalheiro, M.; Pais, P.; Galocha, M.; Teixeira, M.C. Host-pathogen interactions mediated by MDR transporters in fungi: As pleiotropic as it gets! Genes 2018, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [Green Version]
- Edgerton, M.; Koshlukova, S.E.; Lo, T.E.; Chrzan, B.G.; Straubinger, R.M.; Raj, P.A. Candidacidal activity of salivary histatins. Identification of a histatin 5-binding protein on Candida albicans. J. Biol. Chem. 1998, 273, 20438–20447. [Google Scholar]
- Albertsen, M.; Bellahn, I.; Krämer, R.; Waffenschmidt, S. Localization and function of the yeast multidrug transporter Tpo1p. J. Biol. Chem. 2003, 278, 12820–12825. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Chadha, S.; Saraswat, D.; Bajwa, J.S.; Li, R.A.; Conti, H.R.; Edgerton, M. Histatin 5 Uptake by Candida albicans Utilizes Polyamine Transporters Dur3 and Dur31 Proteins. J. Biol. Chem. 2011, 286, 43748–43758. [Google Scholar] [CrossRef] [Green Version]
- Bergin, D.; Reeves, E.P.; Renwick, J.; Wientjes, F.B.; Kavanagh, K. Superoxide Production in Galleria mellonella Hemocytes: Identification of Proteins Homologous to the NADPH Oxidase Complex of Human Neutrophils. Infect. Immun. 2005, 73, 4161–4170. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, S.; Sanguinetti, M.; Torelli, R.; Posteraro, B.; Sanglard, D. Contribution of CgPDR1-regulated genes in enhanced virulence of azole-resistant Candida glabrata. PLoS ONE 2011, 6, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jemel, S.; Guillot, J.; Kallel, K.; Botterel, F.; Dannaoui, E. Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review. Microorganisms 2020, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiering, M.J. Primer on the Immune System. Alcohol Res. 2015, 37, 171–175. [Google Scholar] [PubMed]
- Viegas, S.C.; Mil-Homens, D.; Fialho, A.M.; Arraiano, C.M. The virulence of Salmonella enterica Serovar Typhimurium in the insect model Galleria mellonella is impaired by mutations in RNase E and RNase III. Appl. Environ. Microbiol. 2013, 79, 6124–6133. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.R.; Schroeder, G.N.; Reynolds, S.; Kosta, A.; Collins, J.W.; Mousnier, A.; Frankel, G. Legionella pneumophila Pathogenesis in the Galleria mellonella Infection Model. Infect. Immun. 2012, 80, 2780–2790. [Google Scholar] [CrossRef] [Green Version]
- Mak, P.; Zdybicka-Barabas, A.; Cytryńska, M. A different repertoire of Galleria mellonella antimicrobial peptides in larvae challenged with bacteria and fungi. Dev. Comp. Immunol. 2010, 34, 1129–1136. [Google Scholar] [CrossRef]
- Sowa-Jasiłek, A.; Zdybicka-Barabas, A.; Stączek, S.; Wydrych, J.; Skrzypiec, K.; Mak, P.; Deryło, K.; Tchórzewski, M.; Cytryńska, M. Galleria mellonella lysozyme induces apoptotic changes in Candida albicans cells. Microbiol. Res. 2016, 193, 121–131. [Google Scholar] [CrossRef]
- Schuhmann, B.; Seitz, V.; Vilcinskas, A.; Podsiadlowski, L. Cloning and expression of gallerimycin, an antifungal peptide expressed in immune response of greater wax moth larvae, Galleria mellonella. Arch. Insect Biochem. Physiol. 2003, 53, 125–133. [Google Scholar] [CrossRef]
- Langen, G.; Imani, J.; Altincicek, B.; Kieseritzky, G.; Kogel, K.-H.; Vilcinskas, A. Transgenic expression of gallerimycin, a novel antifungal insect defensin from the greater wax moth Galleria mellonella, confers resistance to pathogenic fungi in tobacco. Biol. Chem. 2006, 387, 549–557. [Google Scholar] [CrossRef]
- Helmerhorst, E.J.; Alagl, A.S.; Siqueira, W.L.; Oppenheim, F.G. Oral fluid proteolytic effects on histatin 5 structure and function. Arch. Oral Biol. 2006, 51, 1061–1070. [Google Scholar] [CrossRef]
- Li, R.; Kumar, R.; Tati, S.; Puri, S.; Edgerton, M. Candida albicans Flu1-Mediated Efflux of Salivary Histatin 5 Reduces Its Cytosolic Concentration and Fungicidal Activity. Antimicrob. Agents Chemother. 2013, 57, 1832–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.; Cavalheiro, M.; Costa, C.; Takahashi-Nakaguchi, A.; Okamoto, M.; Chibana, H.; Teixeira, M.C. Screening the Drug:H+ Antiporter Family for a Role in Biofilm Formation in Candida glabrata. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Vargas, R.C.; García-Salcedo, R.; Tenreiro, S.; Teixeira, M.C.; Fernandes, A.R.; Ramos, J.; Sá-Correia, I. Saccharomyces cerevisiae multidrug resistance transporter Qdr2 is implicated in potassium uptake, providing a physiological advantage to quinidine-stressed cells. Eukaryot. Cell 2007, 6, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, M.C.; Cabrito, R.; Hanif, Z.M.; Vargas, R.C.; Tenreiro, S.; Sá-Correia, I. Yeast response and tolerance to polyamine toxicity involving the drug: H+ antiporter Qdr3 and the transcription factors Yap1 and Gcn4. Microbiology 2011, 157, 945–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, Y.; Itoi, T.; Sugimoto, M.; Sofuni, A.; Tsuchiya, T.; Tanaka, R.; Tonozuka, R.; Honjo, M.; Mukai, S.; Fujita, M.; et al. Elevated polyamines in saliva of pancreatic cancer. Cancers 2018, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, K.; Uno, J.; Nakayama, H.; Sasamoto, K.; Mikami, Y.; Chibana, H. Development of a Highly Efficient Gene Targeting System Induced by Transient Repression of YKU80 Expression in Candida glabrata. Eukaryot. Cell 2007, 6, 1239–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, P.; Vagueiro, S.; Mil-Homens, D.; Pimenta, A.I.; Viana, R.; Okamoto, M.; Chibana, H.; Fialho, A.M.; Teixeira, M.C. A new regulator in the crossroads of oxidative stress resistance and virulence in Candida glabrata: The transcription factor CgTog1. Virulence 2020, 11, 1522–1538. [Google Scholar] [CrossRef]
- Jansen, G.; Wu, C.; Schade, B.; Thomas, D.Y.; Whiteway, M. Drag&Drop cloning in yeast. Gene 2005, 344, 43–51. [Google Scholar] [CrossRef]
- Costa, C.; Pires, C.; Cabrito, T.R.; Renaudin, A.; Ohno, M.; Chibana, H.; Sá-Correia, I.; Teixeira, M.C. Candida glabrata drug:H+ antiporter CgQdr2 confers imidazole drug resistance, being activated by transcription factor CgPdr1. Antimicrob. Agents Chemother. 2013, 57, 3159–3167. [Google Scholar] [CrossRef] [Green Version]
- Vyas, V.K.; Bushkin, G.G.; Bernstein, D.A.; Getz, M.A.; Sewastianik, M.; Barrasa, M.I.; Bartel, D.P.; Fink, G.R. New CRISPR Mutagenesis Strategies Reveal Variation in Repair Mechanisms among Fungi. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Mil-Homens, D.; Fialho, A.M. A BCAM0223 Mutant of Burkholderia cenocepacia Is Deficient in Hemagglutination, Serum Resistance, Adhesion to Epithelial Cells and Virulence. PLoS ONE 2012, 7, e41747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmelo, V.; Santos, H.; Sá-Correia, I. Effect of extracellular acidification on the activity of plasma membrane ATPase and on the cytosolic and vacuolar pH of Saccharomyces cerevisiae. Biochim. Biophys. Acta 1997, 1325, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Altincicek, B.; Vilcinskas, A. Metamorphosis and collagen-IV-fragments stimulate innate immune response in the greater wax moth, Galleria mellonella. Dev. Comp. Immunol. 2006, 30, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I.; Kowalski, P.; Jakubowicz, T. Humoral immune response of Galleria mellonella larvae after infection by Beauveria bassiana under optimal and heat-shock conditions. J. Insect Physiol. 2009, 55, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Xu, L.; Yi, Y. Galleria mellonella larvae are capable of sensing the extent of priming agent and mounting proportionatal cellular and humoral immune responses. Immunol. Lett. 2016, 174, 45–52. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| CgTPO4 gene cloning | |
| pGREG_CgTPO4 _Fw | GAATTCGATATCAAGCTTATCGATACCGTCGACAATGGCCGGTACAAATCAAG |
| pGREG_CgTPO4_Rev | GCGTGACATAACTAATTACATGACTCGAGGTCGACCTATACCATTCTAGAGGAG |
| pGREG576 GAL-to-MTI promotor replacement | |
| pGREG_MTI_ Fw | TTAACCCTCACTAAAGGGAACAAAAGCTGGAGCTCTGTACGACACGCATCATGTGGCAATC |
| pGREG_MTI_Rev | GAAAAGTTCTTCTCCTTTACTCATACTAGTGCGGCTGTGTTTGTTTTTGTATGTGTTTGTTG |
| CgURA3 gene disruption | |
| CgURA3 gRNA in pV1382 _Fw | GATCGACCGGCCAAGGTATCGTCACG |
| CgURA3 gRNA in pV1382 _Rev | AAAACGTGACGATACCTTGGCCGGTC |
| RT-PCR experiments | |
| P1RT Galle | CGCAATATCATTGGCCTTCT [53] |
| P2RT Galle | CCTGCAGTTAGCAATGCAC [53] |
| P1RT Galli | TCGTATCGTCACCGCAAAATG [54] |
| P2RT Galli | GCCGCAATGACCACCTTTATA [54] |
| P1RT IMPI | AGATGGCTATGCAAGGGATG [53] |
| P2RT IMPI | AGGACCTGTGCAGCATTTCT [53] |
| P1RT Lys | TCCCAACTCTTGACCGACGA [53] |
| P2RT Lys | AGTGGTTGCGCCATCCATAC [53] |
| P1RT Cecr | ATTTGCCTGCATCGTAGCG [55] |
| P2RT Cecr | CTTGTACTGCTGGACCAGCTTTT [55] |
| P1RT Act | ATCCTCACCCTGAAGTACCC [53] |
| P2RT Act | CCACACGCAGCTCATTGTA [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalheiro, M.; Romão, D.; Santos, R.; Mil-Homens, D.; Pais, P.; Costa, C.; Galocha, M.; Pereira, D.; Takahashi-Nakaguchi, A.; Chibana, H.; et al. Role of CgTpo4 in Polyamine and Antimicrobial Peptide Resistance: Determining Virulence in Candida glabrata. Int. J. Mol. Sci. 2021, 22, 1376. https://doi.org/10.3390/ijms22031376
Cavalheiro M, Romão D, Santos R, Mil-Homens D, Pais P, Costa C, Galocha M, Pereira D, Takahashi-Nakaguchi A, Chibana H, et al. Role of CgTpo4 in Polyamine and Antimicrobial Peptide Resistance: Determining Virulence in Candida glabrata. International Journal of Molecular Sciences. 2021; 22(3):1376. https://doi.org/10.3390/ijms22031376
Chicago/Turabian StyleCavalheiro, Mafalda, Daniela Romão, Rui Santos, Dalila Mil-Homens, Pedro Pais, Catarina Costa, Mónica Galocha, Diana Pereira, Azusa Takahashi-Nakaguchi, Hiroji Chibana, and et al. 2021. "Role of CgTpo4 in Polyamine and Antimicrobial Peptide Resistance: Determining Virulence in Candida glabrata" International Journal of Molecular Sciences 22, no. 3: 1376. https://doi.org/10.3390/ijms22031376
APA StyleCavalheiro, M., Romão, D., Santos, R., Mil-Homens, D., Pais, P., Costa, C., Galocha, M., Pereira, D., Takahashi-Nakaguchi, A., Chibana, H., Fialho, A. M., & Teixeira, M. C. (2021). Role of CgTpo4 in Polyamine and Antimicrobial Peptide Resistance: Determining Virulence in Candida glabrata. International Journal of Molecular Sciences, 22(3), 1376. https://doi.org/10.3390/ijms22031376