
Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite

 ,
,
Abstract
:1. Introduction
2. Results
2.1. VASP Analysis
2.2. Immunoblot Analysis
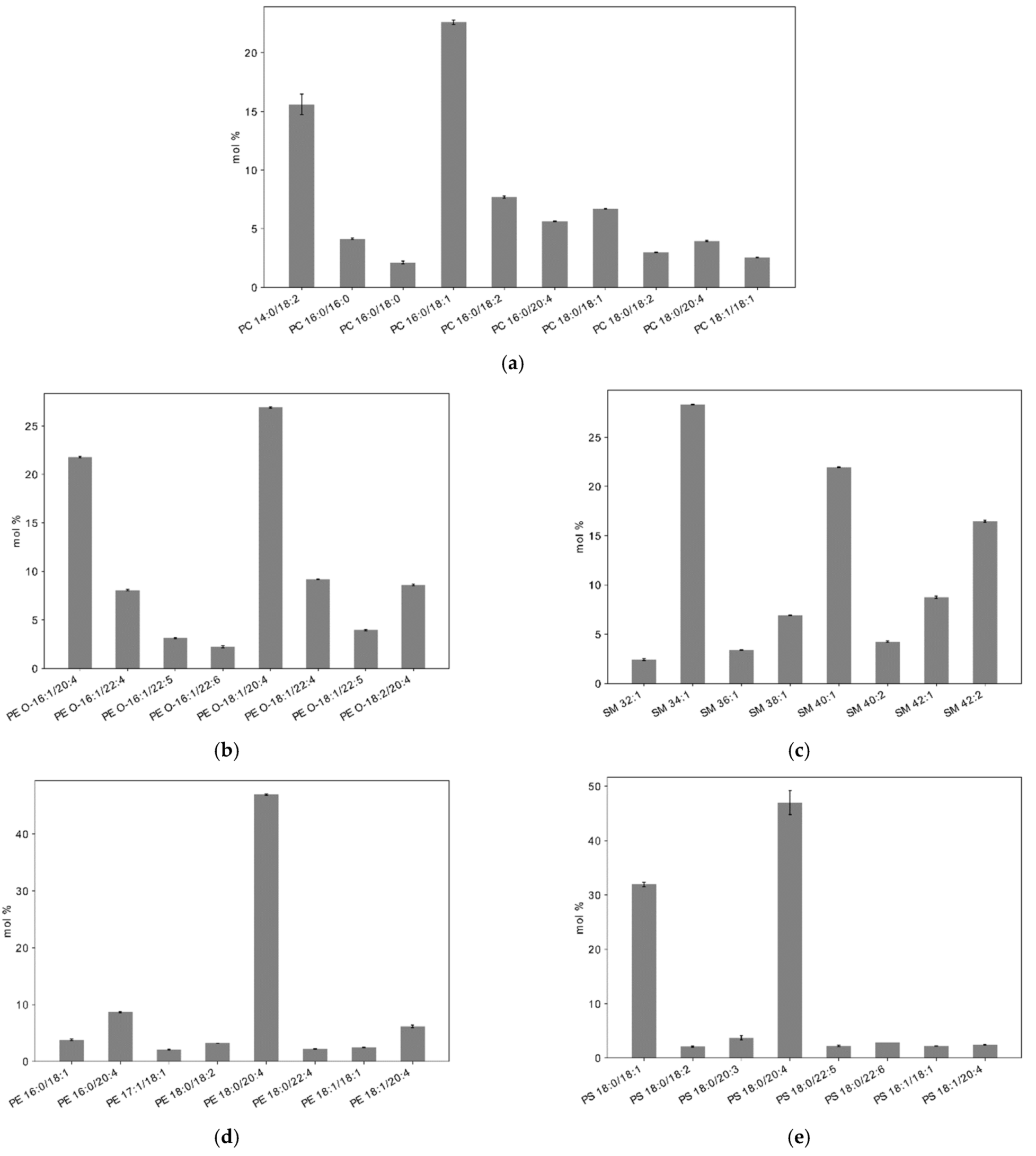
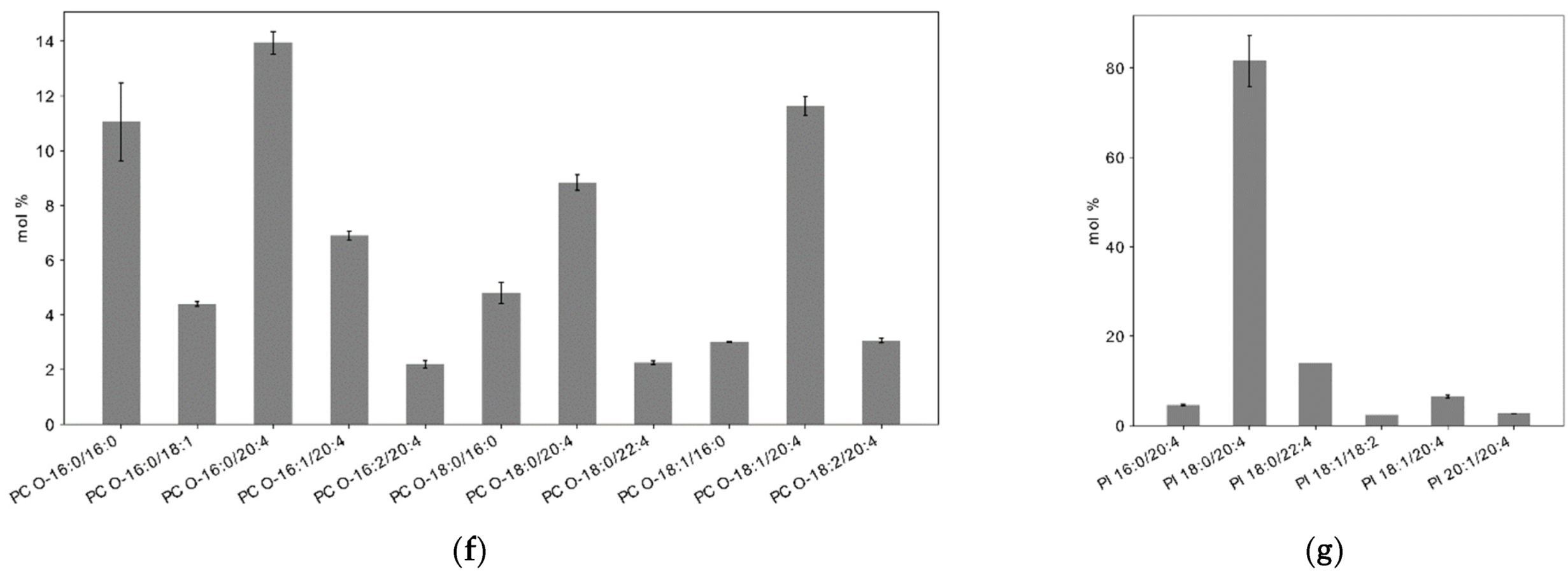
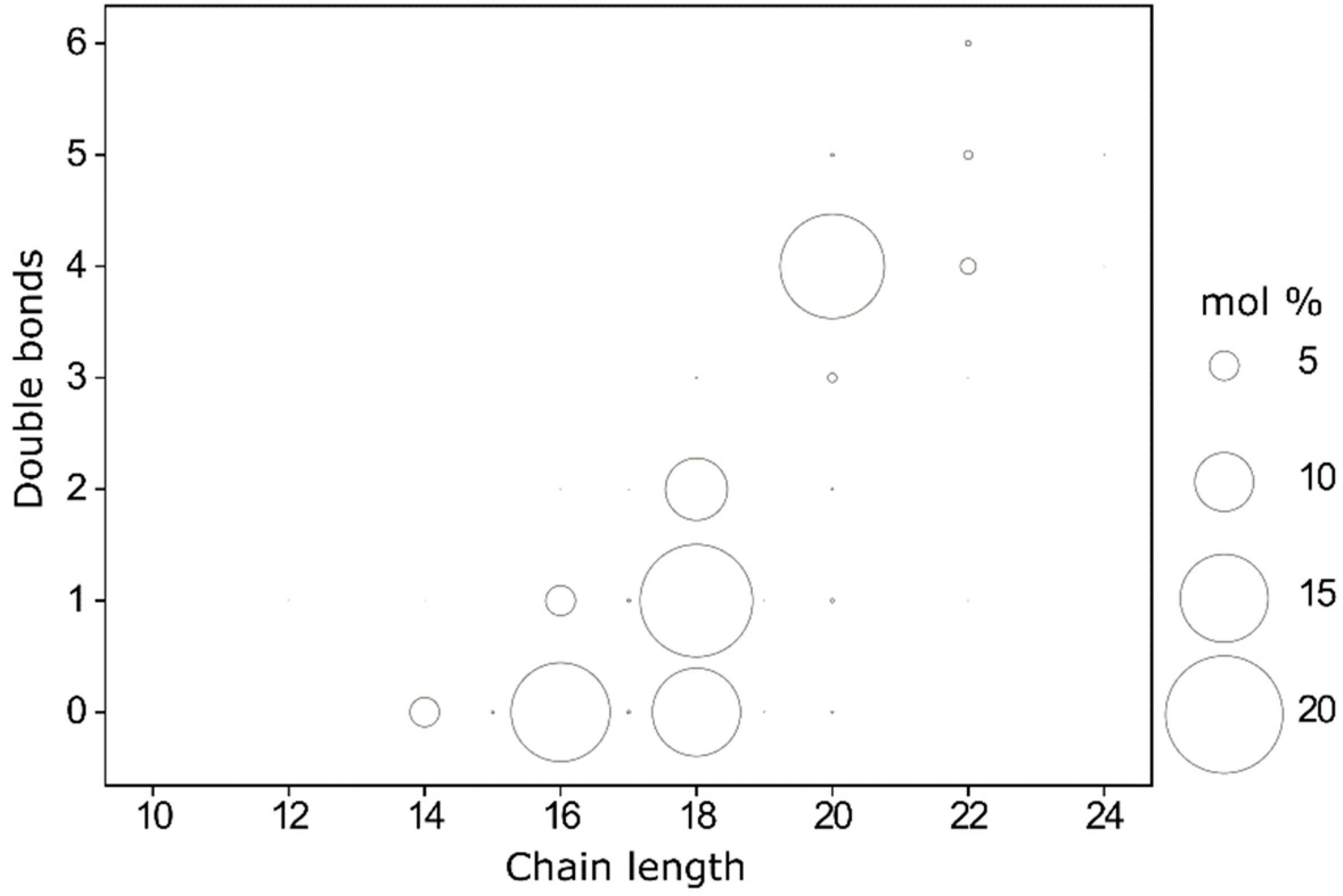
2.3. Lipid Composition of Plasma Membrane in Resting Platelets
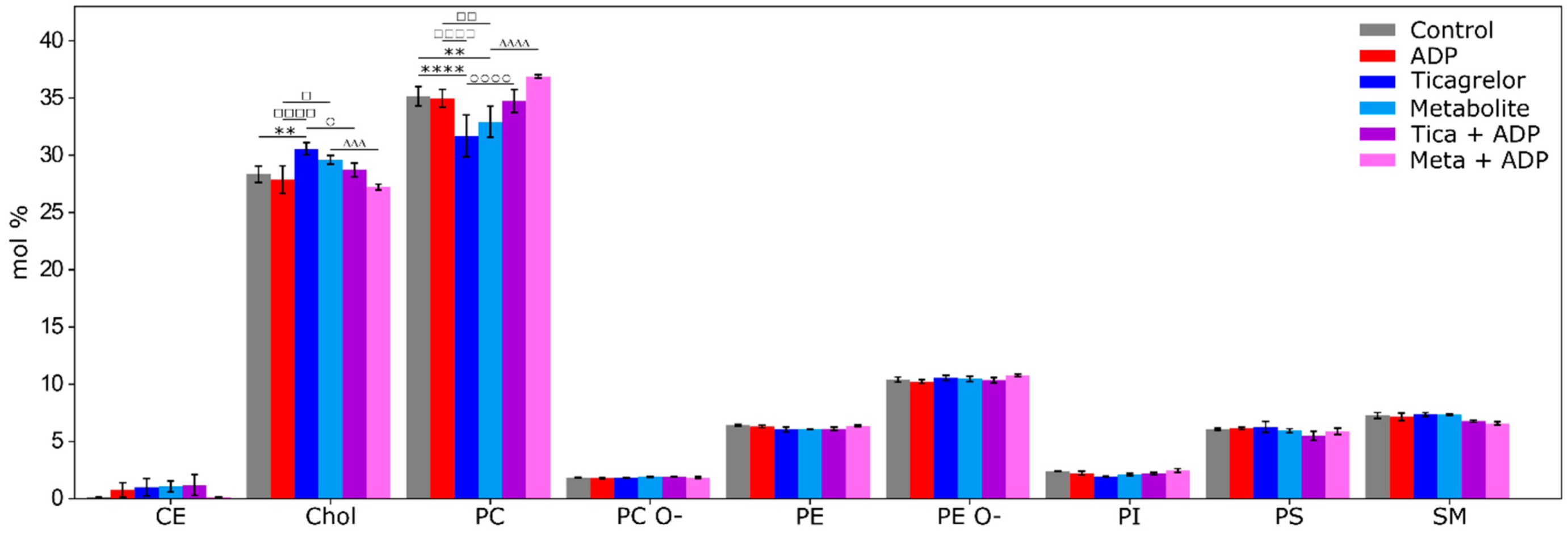
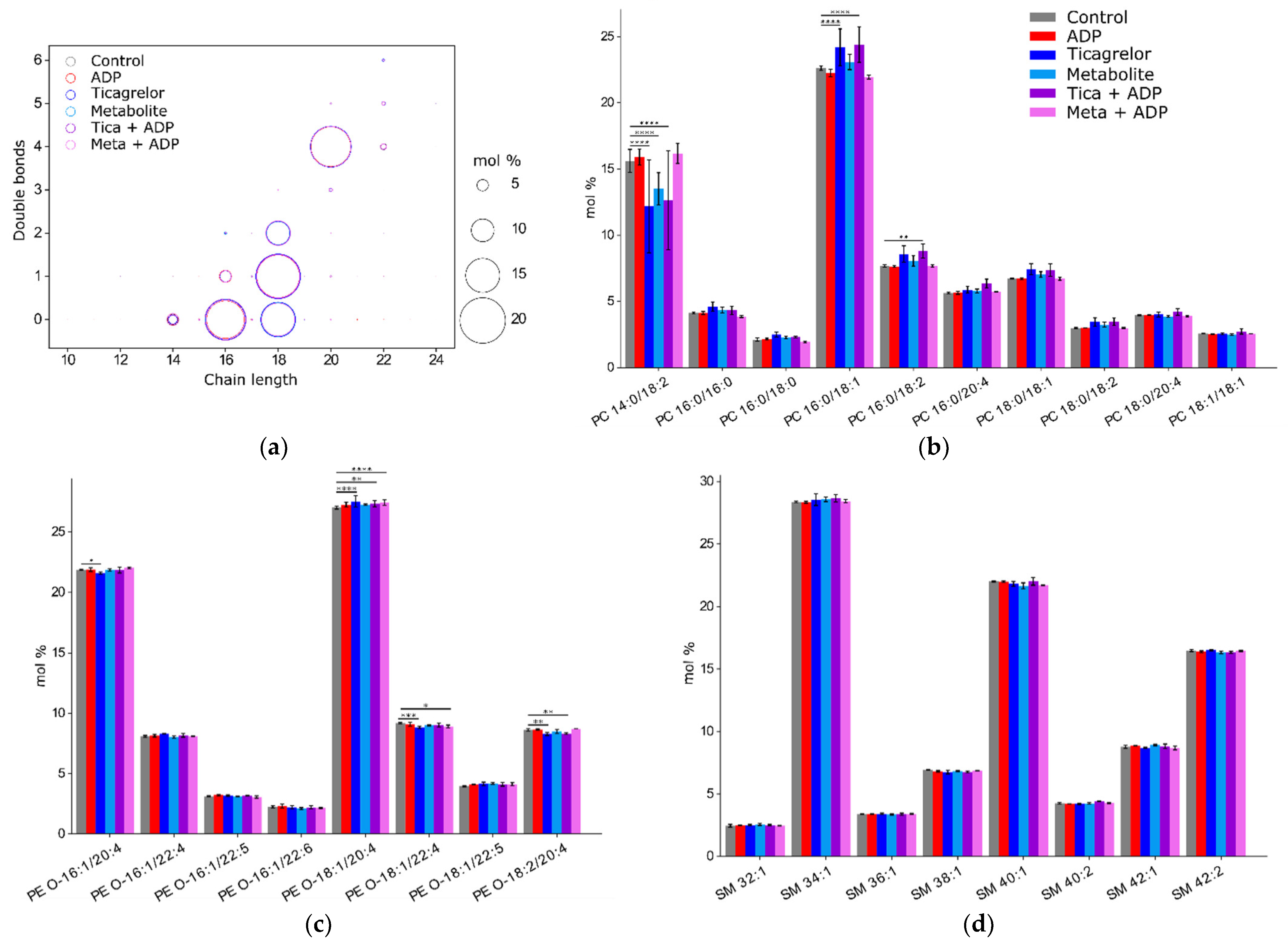
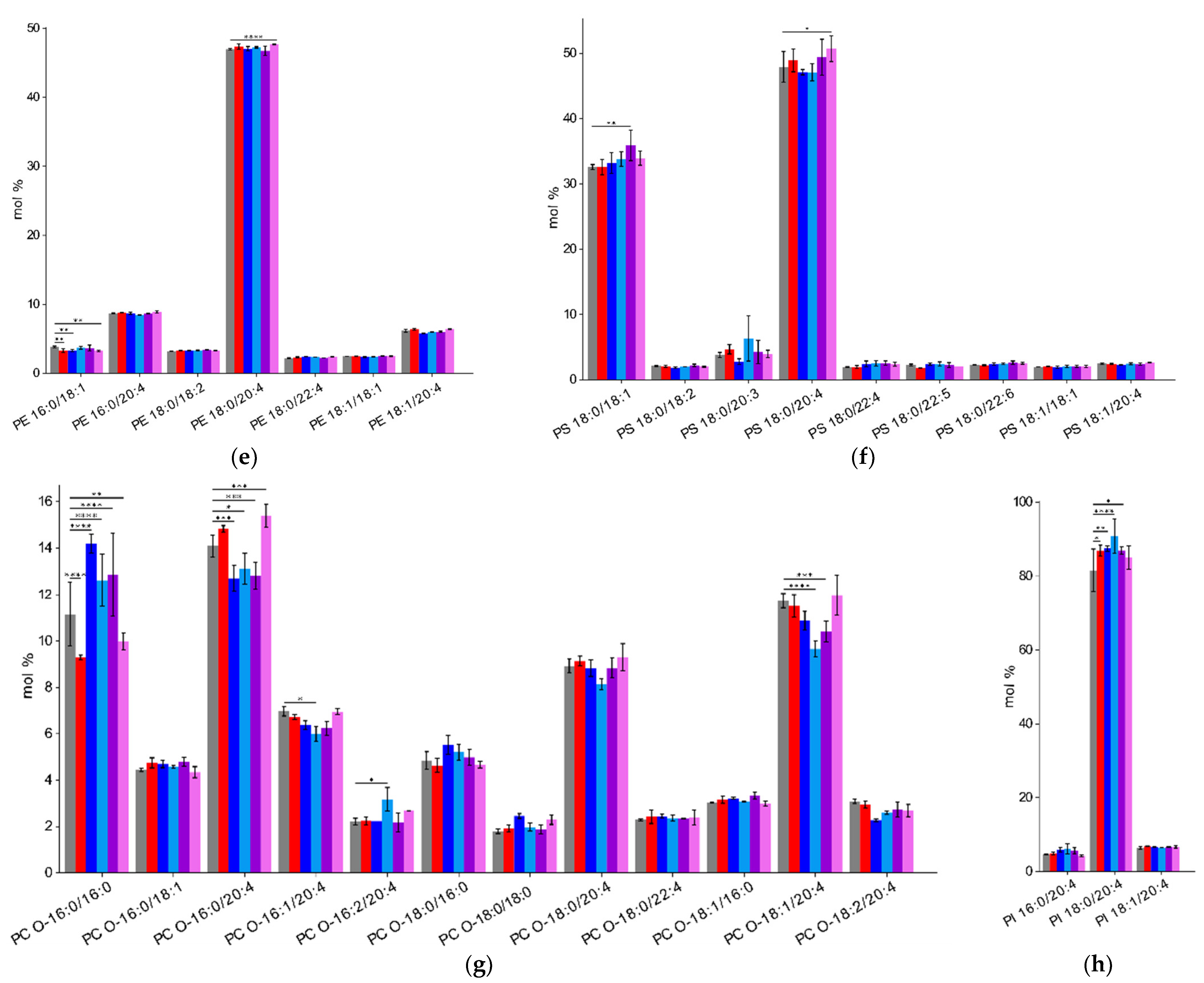
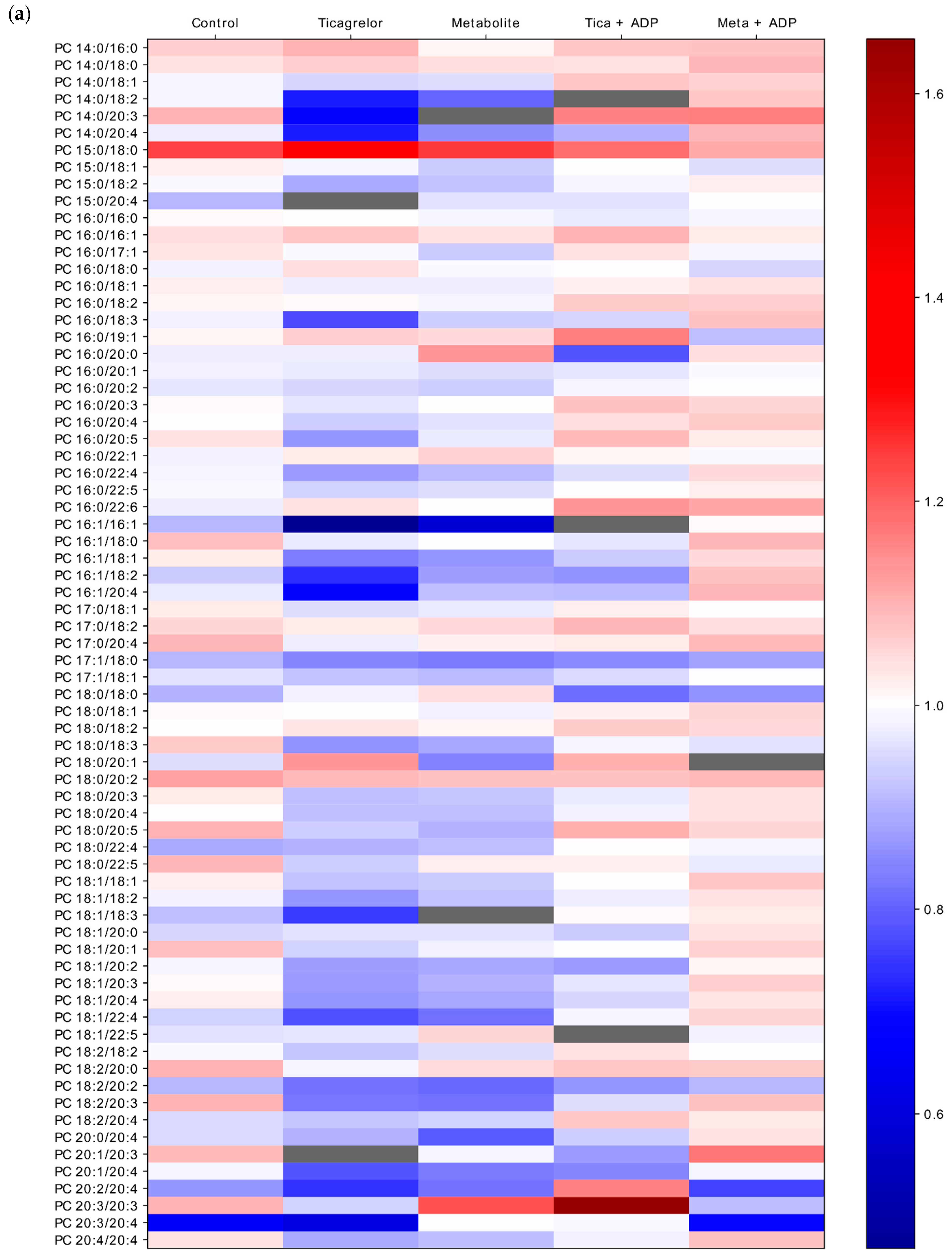
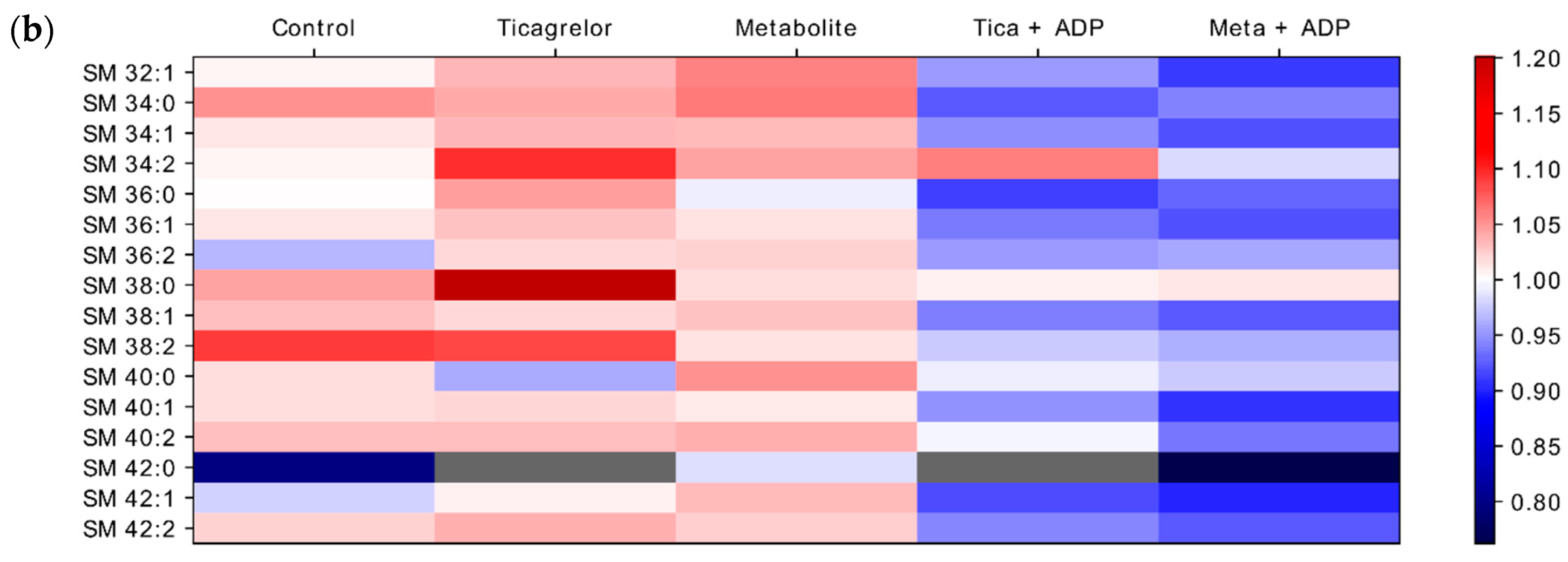
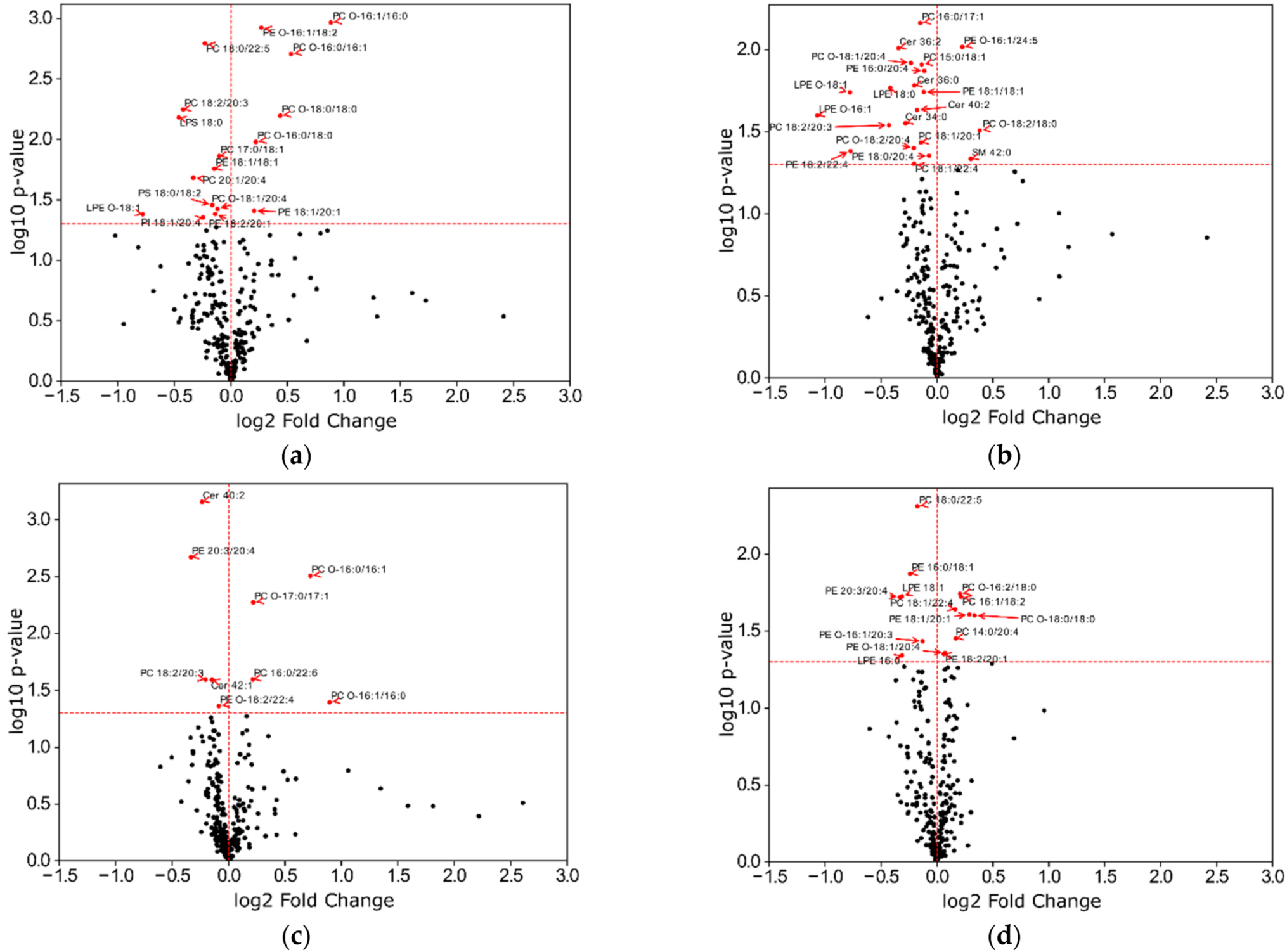
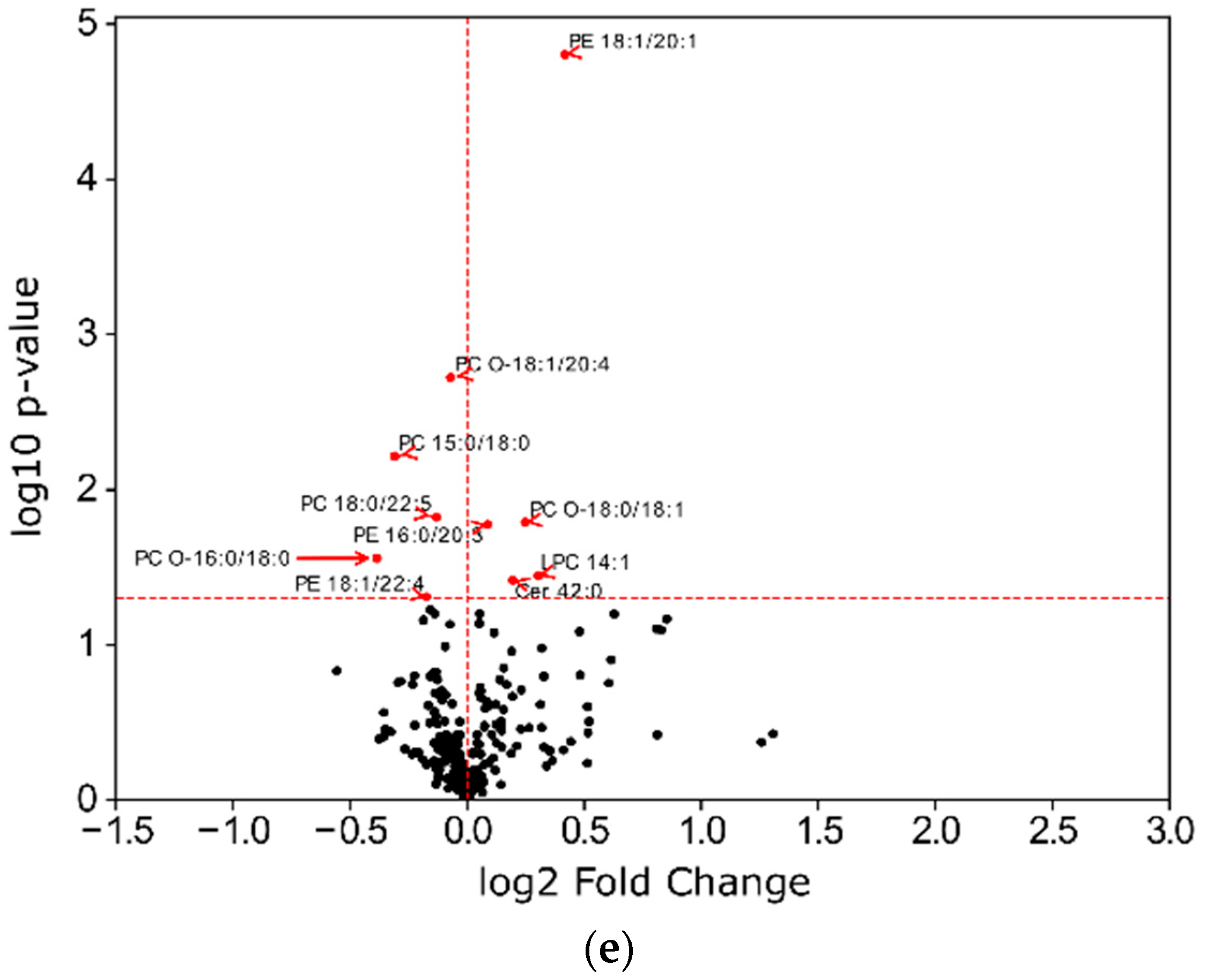
2.4. Impact of ADP, Ticagrelor and Its Active Metabolite on Lipid Classes of Plasma Membrane
3. Discussion
4. Material and Methods
4.1. Experimental Groups
4.2. Extraction of Platelet Plasma
4.3. VASP Phosphorylation Analysis
4.4. Immunoblot Assay
4.5. Lipid Extraction for Mass Spectrometry Lipidomics
4.6. MS Data Acquisition
4.7. Data Analysis and Post-Processing
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| BSA | Bovine Serum Albumin |
| CL | Cardiolipin |
| Cer | Ceramide |
| Chol | Cholesterol |
| CE | Cholesteryl ester |
| COX IV | Cytochrome c oxidase IV |
| DAG | Diacylglycerol |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| ENT1 | Equilibrative Nucleoside Transporter 1 |
| GP1b | Glycoprotein Ib |
| HexCer | Hexosylceramide |
| LPA | Lyso-phosphatidates |
| LPC | Lyso-phosphatidylcholines |
| LPE | Lyso-phosphatidylethanolamines |
| LPG | Lyso-phosphatidylglycerols |
| LPI | Lyso-phosphatidylinositols |
| LPS | Lyso-phosphatidylserines |
| MFI | Mean Fluorescence Intensity |
| NCF1 | Neutrophil cytosolic factor 1 |
| PA | Phosphatidate |
| PCA | Principal Component Analysis |
| PC | Phosphatidylcholines |
| PC O- | Phosphatidylcholines ether |
| PE | Phosphatidylethanolamines |
| PE O- | Phosphatidylethanolamines ether |
| PG | Phosphatidylglycerols |
| PI | Phosphatidylinositols |
| PS | Phosphatidylserines |
| PGE1 | Prostaglandin E1 |
| PRI | Platelet Reactivity Index |
| PUFA | Polyunsaturated Fatty Acids |
| SM | Sphingomyelins |
| TAG | Triacylglycerol |
| VASP | Vasodilator-Stimulated Phosphoprotein |
References
- Jurk, K.; Kehrel, B.E. Platelets: Physiology and Biochemistry. Semin. Thromb. Hemost. 2005, 31, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.G. 3-The Structure of Resting and Activated Platelets. In Platelets, 4th ed.; Michelson, A.D., Ed.; Academic Press: New York, NY, USA, 2019; pp. 47–77. ISBN 978-0-12-813456-6. [Google Scholar]
- von Kügelgen, I. Structure, Pharmacology and Roles in Physiology of the P2Y12 Receptor. Adv. Exp. Med. Biol. 2017. [Google Scholar] [CrossRef]
- Valgimigli, M.; Bueno, H.; Byrne, R.A.; Collet, J.-P.; Costa, F.; Jeppsson, A.; Jüni, P.; Kastrati, A.; Kolh, P.; Mauri, L.; et al. 2017 ESC Focused Update on Dual Antiplatelet Therapy in Coronary Artery Disease Developed in Collaboration with EACTS: The Task Force for Dual Antiplatelet Therapy in Coronary Artery Disease of the European Society of Cardiology (ESC) and of the European Association for Cardio-Thoracic Surgery (EACTS). Eur. Heart J. 2018, 39, 213–260. [Google Scholar] [CrossRef] [PubMed]
- Cammisotto, V.; Biondi-Zoccai, G.; Frati, G.; Giordano, A. Prasugrel and Ticagrelor: The Romulus and Remus of Antiplatelet Therapy? Am. J. Cardiovasc Drugs 2019, 19, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Nylander, S.; Schulz, R. Effects of P2Y12 Receptor Antagonists beyond Platelet Inhibition—Comparison of Ticagrelor with Thienopyridines. Br. J. Pharm. 2016, 173, 1163–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aungraheeta, R.; Conibear, A.; Butler, M.; Kelly, E.; Nylander, S.; Mumford, A.; Mundell, S.J. Inverse Agonism at the P2Y12 Receptor and ENT1 Transporter Blockade Contribute to Platelet Inhibition by Ticagrelor. Blood 2016, 128, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Rabani, V.; Montange, D.; Meneveau, N.; Davani, S. Impact of Ticagrelor on P2Y1 and P2Y12 Localization and on Cholesterol Levels in Platelet Plasma Membrane. Platelets 2017, 1–7. [Google Scholar] [CrossRef]
- Rawicz, W.; Olbrich, K.C.; McIntosh, T.; Needham, D.; Evans, E. Effect of Chain Length and Unsaturation on Elasticity of Lipid Bilayers. Biophys. J. 2000, 79, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.; Geue, S.; Coman, C.; Münzer, P.; Kopczynski, D.; Has, C.; Hoffmann, N.; Manke, M.-C.; Lang, F.; Sickmann, A.; et al. Identification of Key Lipids Critical for Platelet Activation by Comprehensive Analysis of the Platelet Lipidome. Blood 2018. [Google Scholar] [CrossRef]
- O’Donnell, V.B.; Murphy, R.C.; Watson, S.P. Platelet Lipidomics: Modern Day Perspective on Lipid Discovery and Characterization in Platelets. Circ. Res. 2014, 114, 1185–1203. [Google Scholar] [CrossRef]
- Schwarz, U.R.; Geiger, J.; Walter, U.; Eigenthaler, M. Flow Cytometry Analysis of Intracellular VASP Phosphorylation for the Assessment of Activating and Inhibitory Signal Transduction Pathways in Human Platelets—Definition and Detection of Ticlopidine/Clopidogrel Effects. Thromb. Haemost. 1999, 82, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perret, B.; Chap, H.J.; Douste-Blazy, L. Asymmetric Distribution of Arachidonic Acid in the Plasma Membrane of Human Platelets. A Determination Using Purified Phospholipases and a Rapid Method for Membrane Isolation. Biochim. Biophys. Acta 1979, 556, 434–446. [Google Scholar] [CrossRef]
- Skeaff, C.M.; Holub, B.J. Altered Phospholipid Composition of Plasma Membranes from Thrombin-Stimulated Human Platelets. Biochim. Biophys. Acta 1985, 834, 164–171. [Google Scholar] [CrossRef]
- Tsvetkova, N.M.; Crowe, J.H.; Walker, N.J.; Crowe, L.M.; Oliver, A.E.; Wolkers, W.F.; Tablin, F. Physical Properties of Membrane Fractions Isolated from Human Platelets: Implications for Chilling Induced Platelet Activation. Mol. Membr. Biol. 1999, 16, 265–272. [Google Scholar] [CrossRef]
- Biró, E.; Akkerman, J.W.N.; Hoek, F.J.; Gorter, G.; Pronk, L.M.; Sturk, A.; Nieuwland, R. The Phospholipid Composition and Cholesterol Content of Platelet-Derived Microparticles: A Comparison with Platelet Membrane Fractions. J. Thromb. Haemost. 2005, 3, 2754–2763. [Google Scholar] [CrossRef]
- Cohen, P.; Derksen, A. Comparison of Phospholipid and Fatty Acid Composition of Human Erythrocytes and Platelets. Br. J. Haematol. 1969, 17, 359–371. [Google Scholar] [CrossRef]
- Broekman, M.J.; Ward, J.W.; Marcus, A.J. Phospholipid Metabolism in Stimulated Human Platelets. Changes in Phosphatidylinositol, Phosphatidic Acid, and Lysophospholipids. J. Clin. Investig. 1980, 66, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Hamid, M.A.; Kunicki, T.J.; Aster, R.H. Lipid Composition of Freshly Prepared and Stored Platelet Concentrates. Blood 1980, 55, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Owen, J.S.; Hutton, R.A.; Day, R.C.; Bruckdorfer, K.R.; McIntyre, N. Platelet Lipid Composition and Platelet Aggregation in Human Liver Disease. J. Lipid Res. 1981, 22, 423–430. [Google Scholar] [CrossRef]
- Dougherty, R.M.; Galli, C.; Ferro-Luzzi, A.; Iacono, J.M. Lipid and Phospholipid Fatty Acid Composition of Plasma, Red Blood Cells, and Platelets and How They Are Affected by Dietary Lipids: A Study of Normal Subjects from Italy, Finland, and the USA. Am. J. Clin. Nutr 1987, 45, 443–455. [Google Scholar] [CrossRef]
- Watanabe, M.; Shiraishi, K.; Itakura, M.; Matsuzaki, S. Relationship between Platelet Membrane Lipid Compositions and Platelet Aggregability in Alcoholic Liver Disease. Alcohol Clin. Exp. Res. 1998, 22, 97S–102S. [Google Scholar] [CrossRef] [PubMed]
- Leidl, K.; Liebisch, G.; Richter, D.; Schmitz, G. Mass Spectrometric Analysis of Lipid Species of Human Circulating Blood Cells. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2008, 1781, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Ruebsaamen, K.; Liebisch, G.; Boettcher, A.; Schmitz, G. Lipidomic Analysis of Platelet Senescence. Transfus. 2010, 50, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Yawata, Y. Cell Membrane: The Red Blood Cell as a Model.; Wiley VCH: Weinheim, Germany, 2003; ISBN 978-3-527-30463-9. [Google Scholar]
- Lorent, J.H.; Levental, K.R.; Ganesan, L.; Rivera-Longsworth, G.; Sezgin, E.; Doktorova, M.; Lyman, E.; Levental, I. Plasma Membranes Are Asymmetric in Lipid Unsaturation, Packing and Protein Shape. Nat. Chem. Biol. 2020. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes Review. Cell Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef]
- Baum, S.J.; Kris-Etherton, P.M.; Willett, W.C.; Lichtenstein, A.H.; Rudel, L.L.; Maki, K.C.; Whelan, J.; Ramsden, C.E.; Block, R.C. Fatty Acids in Cardiovascular Health and Disease: A Comprehensive Update. J. Clin. Lipidol 2012, 6, 216–234. [Google Scholar] [CrossRef]
- Harayama, T.; Riezman, H. Understanding the Diversity of Membrane Lipid Composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Pinot, M.; Vanni, S.; Pagnotta, S.; Lacas-Gervais, S.; Payet, L.-A.; Ferreira, T.; Gautier, R.; Goud, B.; Antonny, B.; Barelli, H. Lipid Cell Biology. Polyunsaturated Phospholipids Facilitate Membrane Deformation and Fission by Endocytic Proteins. Science 2014, 345, 693–697. [Google Scholar] [CrossRef]
- McMahon, H.T.; Boucrot, E. Membrane Curvature at a Glance. J. Cell Sci. 2015, 128, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The Mystery of Membrane Organization: Composition, Regulation and Roles of Lipid Rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Rabani, V.; Lagoutte-Renosi, J.; Series, J.; Valot, B.; Xuereb, J.-M.; Davani, S. Cholesterol-Rich Microdomains Contribute to PAR1 Signaling in Platelets Despite a Weak Localization of the Receptor in These Microdomains. Int. J. Mol. Sci. 2020, 21, 8065. [Google Scholar] [CrossRef] [PubMed]
- Rabani, V.; Montange, D.; Davani, S. Interactive Protein Network of FXIII-A1 in Lipid Rafts of Activated and Non-Activated Platelets. Platelets 2016, 27, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Corradi, V.; Sejdiu, B.I.; Mesa-Galloso, H.; Abdizadeh, H.; Noskov, S.Y.; Marrink, S.J.; Tieleman, D.P. Emerging Diversity in Lipid-Protein Interactions. Chem. Rev. 2019, 119, 5775–5848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, S.M.; Majerus, P.W. The Fatty Acid Composition of Phosphatidylinositol from Thrombin-Stimulated Human Platelets. J. Biol. Chem. 1981, 256, 579–582. [Google Scholar] [CrossRef]
- Prisco, D.; Tufano, A.; Cenci, C.; Pignatelli, P.; Santilli, F.; Di Minno, G.; Perticone, F. Position Paper of the Italian Society of Internal Medicine (SIMI) on Prophylaxis and Treatment of Venous Thromboembolism in Patients with Cancer. Intern. Emerg Med. 2019, 14, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Okpala, I.; Ghebremeskel, K.; Ugochukwu, C.C.; Ibegbulam, O.; Crawford, M. Blood Mononuclear Cells and Platelets Have Abnormal Fatty Acid Composition in Homozygous Sickle Cell Disease. Ann. Hematol 2005, 84, 578–583. [Google Scholar] [CrossRef]
- García-Rubio, D.; Rodríguez-Varela, M.; Martínez-Vieyra, I.; de la Mora, M.B.; Méndez-Méndez, J.V.; Durán-Álvarez, J.C.; Cerecedo, D. Alterations to the Contents of Plasma Membrane Structural Lipids Are Associated with Structural Changes and Compartmentalization in Platelets in Hypertension. Exp. Cell Res. 2019, 385, 111692. [Google Scholar] [CrossRef]
- Sengupta, D.; Chattopadhyay, A. Molecular Dynamics Simulations of GPCR-Cholesterol Interaction: An Emerging Paradigm. Biochim Biophys Acta 2015, 1848, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, I.; Liu, X.; Neya, S.; Hoshino, T. Influence of Lipid Composition on the Structural Stability of G-Protein Coupled Receptor. Chem. Pharm Bull. (Tokyo) 2013, 61, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Watała, C.; Gwoździński, K. Effect of Aspirin on Conformation and Dynamics of Membrane Proteins in Platelets and Erythrocytes. Biochem. Pharm. 1993, 45, 1343–1349. [Google Scholar] [CrossRef]
- Alves, A.C.; Ribeiro, D.; Horta, M.; Lima, J.L.F.C.; Nunes, C.; Reis, S. A Biophysical Approach to Daunorubicin Interaction with Model Membranes: Relevance for the Drug’s Biological Activity. J. R Soc. Interface 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.T.; Nguyen, T.Q.; Dao, U.P.N.; Nguyen, T.T. On the Interaction between Fluoxetine and Lipid Membranes: Effect of the Lipid Composition. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 191, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Winocour, P.D.; Watala, C.; Perry, D.W.; Kinlough-Rathbone, R.L. Decreased Platelet Membrane Fluidity Due to Glycation or Acetylation of Membrane Proteins. Thromb. Haemost. 1992, 68, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Doktorova, M.; Molugu, T.R.; Heberle, F.A.; Scott, H.L.; Dzikovski, B.; Nagao, M.; Stingaciu, L.-R.; Standaert, R.F.; Barrera, F.N.; et al. How Cholesterol Stiffens Unsaturated Lipid Membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 21896–21905. [Google Scholar] [CrossRef] [PubMed]
- Osamah, H.; Mira, R.; Sorina, S.; Shlomo, K.; Michael, A. Reduced Platelet Aggregation after Fluvastatin Therapy Is Associated with Altered Platelet Lipid Composition and Drug Binding to the Platelets. Br. J. Clin. Pharm. 1997, 44, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Storey, R.F.; Husted, S.; Harrington, R.A.; Heptinstall, S.; Wilcox, R.G.; Peters, G.; Wickens, M.; Emanuelsson, H.; Gurbel, P.; Grande, P.; et al. Inhibition of Platelet Aggregation by AZD6140, a Reversible Oral P2Y12 Receptor Antagonist, Compared with Clopidogrel in Patients with Acute Coronary Syndromes. J. Am. Coll Cardiol. 2007, 50, 1852–1856. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Rojo, N.; Riezman, H. On the Road to Unraveling the Molecular Functions of Ether Lipids. FEBS Lett. 2019, 593, 2378–2389. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.M.; Lodhi, I.J. Structural and Functional Roles of Ether Lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Koivuniemi, A. The Biophysical Properties of Plasmalogens Originating from Their Unique Molecular Architecture. FEBS Lett. 2017, 591, 2700–2713. [Google Scholar] [CrossRef]
- Dorninger, F.; Forss-Petter, S.; Wimmer, I.; Berger, J. Plasmalogens, Platelet-Activating Factor and beyond—Ether Lipids in Signaling and Neurodegeneration. Neurobiol Dis 2020, 145, 105061. [Google Scholar] [CrossRef]
- Wood, P.L.; Unfried, G.; Whitehead, W.; Phillipps, A.; Wood, J.A. Dysfunctional Plasmalogen Dynamics in the Plasma and Platelets of Patients with Schizophrenia. Schizophr. Res. 2015, 161, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of Plasmalogen Lipids in Health and Disease. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletis, C.D. Alkyl-Acylglycerols and the Important Clinical Ramifications of Raising Plasmalogens in Dementia and Alzheimer’s Disease. Integr. Med. (Encinitas) 2020, 19, 12–16. [Google Scholar] [PubMed]
- Astudillo, A.M.; Balboa, M.A.; Balsinde, J. Selectivity of Phospholipid Hydrolysis by Phospholipase A2 Enzymes in Activated Cells Leading to Polyunsaturated Fatty Acid Mobilization. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R. Arachidonic Acid as a Bioactive Molecule. J. Clin. Investig. 2001, 107, 1339–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnweber, T.; Pizzini, A.; Nairz, M.; Weiss, G.; Tancevski, I. Arachidonic Acid Metabolites in Cardiovascular and Metabolic Diseases. Int. J. Mol. Sci. 2018, 19, 3285. [Google Scholar] [CrossRef] [Green Version]
- Tallima, H.; El Ridi, R. Arachidonic Acid: Physiological Roles and Potential Health Benefits—A Review. J. Adv. Res. 2017, 11, 33–41. [Google Scholar] [CrossRef]
- Hamberg, M.; Svensson, J.; Samuelsson, B. Thromboxanes: A New Group of Biologically Active Compounds Derived from Prostaglandin Endoperoxides. Proc. Natl. Acad. Sci. USA 1975, 72, 2994–2998. [Google Scholar] [CrossRef] [Green Version]
- Lebrero, P.; Astudillo, A.M.; Rubio, J.M.; Fernández-Caballero, L.; Kokotos, G.; Balboa, M.A.; Balsinde, J. Cellular Plasmalogen Content Does Not Influence Arachidonic Acid Levels or Distribution in Macrophages: A Role for Cytosolic Phospholipase A2γ in Phospholipid Remodeling. Cells 2019, 8, 799. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Hayashi, Y.; Nemoto-Sasaki, Y.; Ito, M.; Oka, S.; Tanikawa, T.; Waku, K.; Sugiura, T. Acyltransferases and Transacylases That Determine the Fatty Acid Composition of Glycerolipids and the Metabolism of Bioactive Lipid Mediators in Mammalian Cells and Model Organisms. Prog. Lipid Res. 2014, 53, 18–81. [Google Scholar] [CrossRef]
- Nieto, M.L.; Venable, M.E.; Bauldry, S.A.; Greene, D.G.; Kennedy, M.; Bass, D.A.; Wykle, R.L. Evidence That Hydrolysis of Ethanolamine Plasmalogens Triggers Synthesis of Platelet-Activating Factor via a Transacylation Reaction. J. Biol. Chem. 1991, 266, 18699–18706. [Google Scholar] [CrossRef]
- Beckett, C.S.; Kell, P.J.; Creer, M.H.; McHowat, J. Phospholipase A2-Catalyzed Hydrolysis of Plasmalogen Phospholipids in Thrombin-Stimulated Human Platelets. Thromb. Res. 2007, 120, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freynhofer, M.K.; Brozovic, I.; Bruno, V.; Farhan, S.; Vogel, B.; Jakl, G.; Willheim, M.; Hübl, W.; Wojta, J.; Huber, K. Multiple Electrode Aggregometry and Vasodilator Stimulated Phosphoprotein-Phosphorylation Assay in Clinical Routine for Prediction of Postprocedural Major Adverse Cardiovascular Events. Thromb. Haemost. 2011, 106, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.L.; Gerl, M.J.; Klose, C.; Ejsing, C.S.; Beug, H.; Simons, K.; Shevchenko, A. Membrane Lipidome of an Epithelial Cell Line. Proc. Natl. Acad. Sci. USA 2011, 108, 1903–1907. [Google Scholar] [CrossRef] [Green Version]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global Analysis of the Yeast Lipidome by Quantitative Shotgun Mass Spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Surma, M.A.; Herzog, R.; Vasilj, A.; Klose, C.; Christinat, N.; Morin-Rivron, D.; Simons, K.; Masoodi, M.; Sampaio, J.L. An Automated Shotgun Lipidomics Platform for High Throughput, Comprehensive, and Quantitative Analysis of Blood Plasma Intact Lipids. Eur. J. Lipid Sci. Technol. 2015, 117, 1540–1549. [Google Scholar] [CrossRef] [Green Version]
- Liebisch, G.; Binder, M.; Schifferer, R.; Langmann, T.; Schulz, B.; Schmitz, G. High Throughput Quantification of Cholesterol and Cholesteryl Ester by Electrospray Ionization Tandem Mass Spectrometry (ESI-MS/MS). Biochim. Biophys. Acta 2006, 1761, 121–128. [Google Scholar] [CrossRef]
- Herzog, R.; Schwudke, D.; Schuhmann, K.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. A Novel Informatics Concept for High-Throughput Shotgun Lipidomics Based on the Molecular Fragmentation Query Language. Genome Biol. 2011, 12, R8. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Schuhmann, K.; Schwudke, D.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. LipidXplorer: A Software for Consensual Cross-Platform Lipidomics. PLoS ONE 2012, 7, e29851. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cholesterol (%) | PC and Related Species (%) | PE and Related Species (%) | PI (%) | PS (%) | SM (%) | Other Lipids Species | Method | Ref/Years | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PC | PC O- | PE | PE O- | ||||||||
| Plasma membrane of resting platelet | Unknown | 35 | - | 26.1 | - | 5.6 | 9.9 | 23.4 | Two-dimensional thin layer chromatography and gas–liquid chromatography | Perret et al. [13]/1979 | |
| Unknown | 39.2 | - | 27.8 | - | 2.5 | 8.6 | 22.1 | Thin layer chromatography and gas–liquid chromatography after transmethylation | Skeaff et al. [14]/1985 | ||
| 7.5 | 34.8 | - | 25.3 | - | 5.8 | 6.4 | 14.6 | 5.2 | Rod like thin-layer chromatography | Tsvetkova et al. [15]/1999 | |
| Unknown | 27 | - | 29 | - | 2 | 19 | 22 | - | High performance thin layer chromatography | Biró et al. [16]/2005 | |
| 28.35 | 37.01 | 16.81 | 2.41 | 6.06 | 7.28 | 2.08 | Mass spectrometry | Result of our study | |||
| 35.17 | 1.84 | 6.41 | 10.40 | ||||||||
| Without cholesterol | 51.92 | 23.59 | 3.38 | 8.50 | 10.19 | 2.42 | Mass spectrometry | Result of our study | |||
| 49.34 | 2.58 | 9.00 | 14.59 | ||||||||
| Lipid composition of whole resting platelet | Unknown | 39.8 | 1.7 | 13.6 | 14.0 | 3.7 | 8.8 | 18.4 | Thin layer and gas–liquid chromatography | Cohen et al. [17]/1969 | |
| Unknown | 38.56 | - | 29.12 | - | 4.17 | 10.98 | 14.96 | 2.21 | Two-dimensional thin layer chromatography and gas chromatography | Broekman et al. [18]/1980 | |
| Unknown | 38.7 | - | 26.9 | - | 4.4 | 9.8 | 18.5 | 1.7 | Thin-layer chromatography and gas–liquid chromatography | Hamid et al. [19]/1980 | |
| 31.5 | 27.5 | - | 18.9 | - | 1.8 | 6.3 | 13.1 | 0.9 | Two-dimensional thin layer chromatography | Owen et al. [20]/1981 | |
| 18.9 | 34.8 | - | 18.8 | - | 11.8 | 13.2 | 16.0 | Thin layer and gas–liquid chromatography | In Italy, Dougherty et al. [21]/1987 | ||
| 22.2 | 38.2 | - | 25.8 | - | 6.5 | 10.6 | 13.2 | Thin layer and gas–liquid chromatography | In Finland, Dougherty et al. [21]/1987 | ||
| 19.3 | 31.1 | - | 23 | - | 10.6 | 12.0 | 16.5 | Thin layer and gas–liquid chromatography | In USA, Dougherty et al. [21]/1987 | ||
| 15,15 | 43 | - | 21.47 | - | 8.8 | 26 | - | thin-layer chromatography using a plate scanner with a flame ionization detector | Watanabe et al. [22]/1998 | ||
| 30 | 26 | - | 12 | - | 1 | 9 | 11 | 2 | Mass spectrometry | Leidl et al. [23]/2008 | |
| Lipid composition of whole stimulated platelet by thrombin | Unknown | 38.44 | - | 28.96 | - | 3.00 | 11.01 | 15.1 | 3.49 | Two-dimensional thin layer chromatography and gas chromatography | Broekman et al. [18]/1980 |
| Unknown | 36.2 | - | 29.4 | - | 1.6 | 9.3 | 23.6 | - | Gas–liquid chromatography | Skeaff et al. [14]/1985 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagoutte-Renosi, J.; Allemand, F.; Ramseyer, C.; Rabani, V.; Davani, S. Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite. Int. J. Mol. Sci. 2021, 22, 1432. https://doi.org/10.3390/ijms22031432
Lagoutte-Renosi J, Allemand F, Ramseyer C, Rabani V, Davani S. Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite. International Journal of Molecular Sciences. 2021; 22(3):1432. https://doi.org/10.3390/ijms22031432
Chicago/Turabian StyleLagoutte-Renosi, Jennifer, Florentin Allemand, Christophe Ramseyer, Vahideh Rabani, and Siamak Davani. 2021. "Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite" International Journal of Molecular Sciences 22, no. 3: 1432. https://doi.org/10.3390/ijms22031432
APA StyleLagoutte-Renosi, J., Allemand, F., Ramseyer, C., Rabani, V., & Davani, S. (2021). Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite. International Journal of Molecular Sciences, 22(3), 1432. https://doi.org/10.3390/ijms22031432