Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
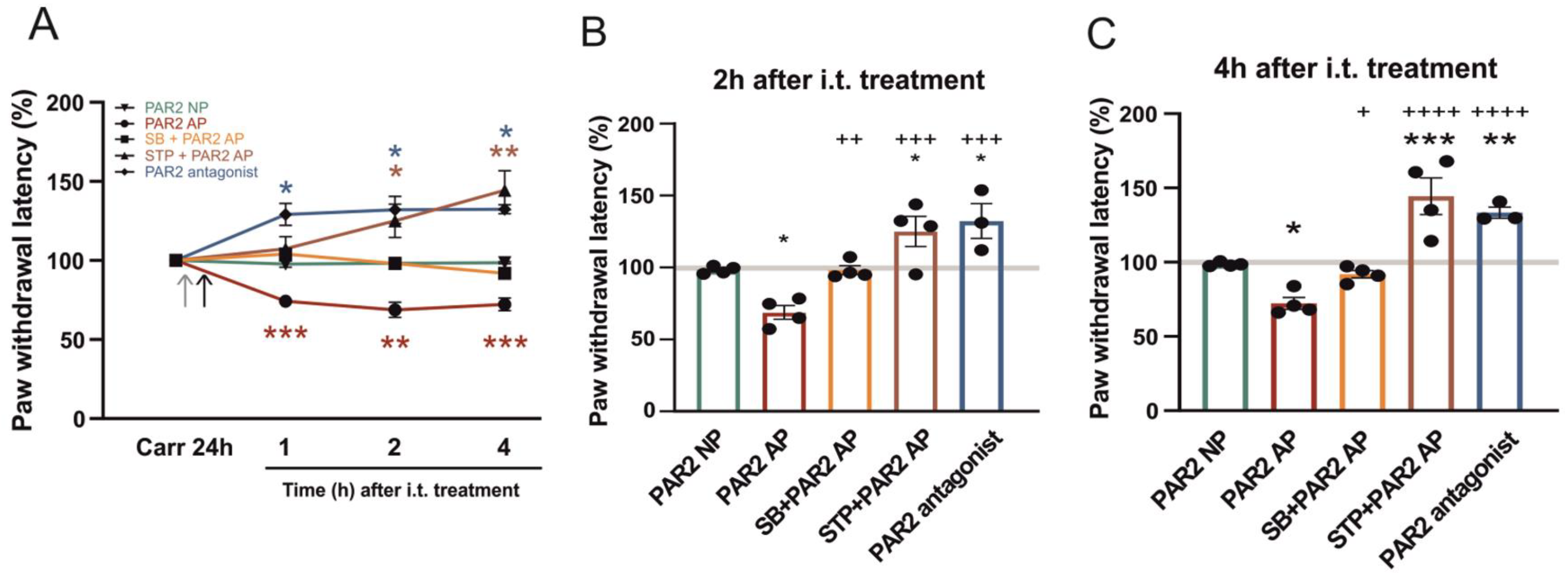
2.1. Activation of Spinal PAR2 in Thermal Hypersensitivity Induced by Peripheral Inflammation
2.2. Modulation of Miniature Excitatory Postsynaptic Currents (mEPSCs) in Dorsal Horn Neurons by PAR2 under the Inflammatory Conditions
2.3. Modulation of Spontaneous Excitatory Postsynaptic Currents (sEPSCs) in Dorsal Horn Neurons by PAR2 under Inflammatory Conditions
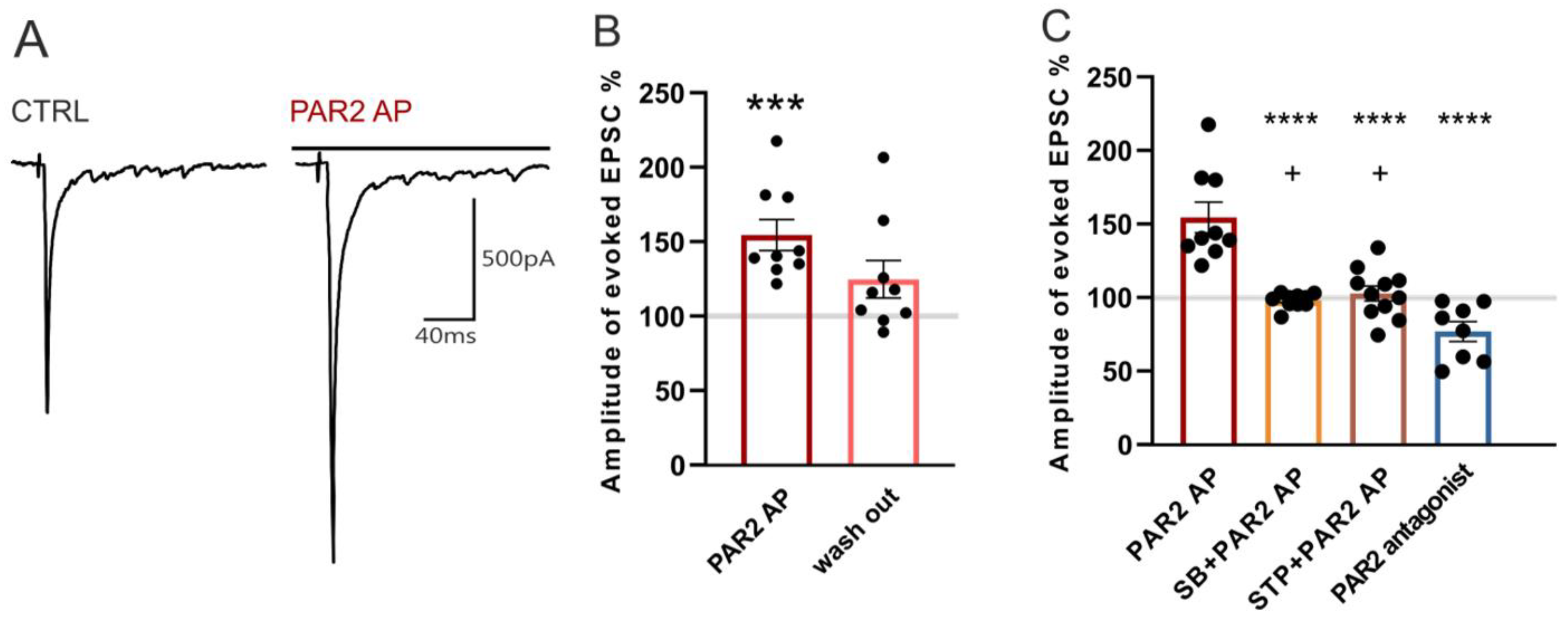
2.4. Modulation of Dorsal Root Stimulation-Evoked EPSC by PAR2 under the Inflammatory Conditions
3. Discussion
3.1. Mechanisms of Spinal PAR2-Mediated Thermal Hyperalgesia
3.2. The Effect of Carrageenan on the Basal Spontaneous EPSC Frequency and Synaptic Transmission
3.3. Activation of Spinal PAR2 after Peripheral Inflammation
4. Methods
4.1. Statement of Ethical Considerations
4.2. Animal Care and Utilization
4.3. Spinal Cord Slice Preparation
4.4. Patch-Clamp Recordings
4.5. Drug Treatment
4.6. Intrathecal Catheter Implantation
4.7. Carrageenan Model of Peripheral Inflammation
4.8. Behavioral Tests
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nystedt, S.; Emilsson, K.; Wahlestedt, C.; Sundelin, J. Molecular cloning of a potential proteinase activated receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 9208–9212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nystedt, S.; Ramakrishnan, V.; Sundelin, J. The proteinase-activated receptor 2 is induced by inflammatory mediators in human endothelial cells. Comparison with the thrombin receptor. J. Biol. Chem. 1996, 271, 14910–14915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelso, E.B.; Lockhart, J.C.; Hembrough, T.; Dunning, L.; Plevin, R.; Hollenberg, M.D.; Sommerhoff, C.P.; McLean, J.S.; Ferrell, W.R. Therapeutic promise of proteinase-activated receptor-2 antagonism in joint inflammation. J. Pharmacol. Exp. Ther. 2006, 316, 1017–1024. [Google Scholar] [CrossRef]
- Vergnolle, N.; Hollenberg, M.D.; Sharkey, K.A.; Wallace, J.L. Characterization of the inflammatory response to proteinase-activated receptor-2 (PAR2)-activating peptides in the rat paw. Br. J. Pharmacol. 1999, 127, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhoff, M.; Vergnolle, N.; Young, S.H.; Tognetto, M.; Amadesi, S.; Ennes, H.S.; Trevisani, M.; Hollenberg, M.D.; Wallace, J.L.; Caughey, G.H.; et al. Agonists of proteinase-activated receptor 2 induce inflammation by a neurogenic mechanism. Nat. Med. 2000, 6, 151–158. [Google Scholar] [CrossRef]
- Bunnett, N.W. Protease-activated receptors: How proteases signal to cells to cause inflammation and pain. Semin Thromb Hemost 2006, 32 (Suppl. 1), 39–48. [Google Scholar] [CrossRef]
- Jimenez-Vargas, N.N.; Pattison, L.A.; Zhao, P.; Lieu, T.; Latorre, R.; Jensen, D.D.; Castro, J.; Aurelio, L.; Le, G.T.; Flynn, B.; et al. Protease-activated receptor-2 in endosomes signals persistent pain of irritable bowel syndrome. Proc. Natl. Acad. Sci. USA 2018, 115, E7438–E7447. [Google Scholar] [CrossRef] [Green Version]
- Ossovskaya, V.S.; Bunnett, N.W. Protease-activated receptors: Contribution to physiology and disease. Physiol. Rev. 2004, 84, 579–621. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Lieu, T.; Barlow, N.; Metcalf, M.; Veldhuis, N.A.; Jensen, D.D.; Kocan, M.; Sostegni, S.; Haerteis, S.; Baraznenok, V.; et al. Cathepsin S causes inflammatory pain via biased agonism of PAR2 and TRPV4. J. Biol. Chem. 2014, 289, 27215–27234. [Google Scholar] [CrossRef] [Green Version]
- Lieu, T.; Savage, E.; Zhao, P.; Edgington-Mitchell, L.; Barlow, N.; Bron, R.; Poole, D.P.; McLean, P.; Lohman, R.J.; Fairlie, D.P.; et al. Antagonism of the proinflammatory and pronociceptive actions of canonical and biased agonists of protease-activated receptor-2. Br. J. Pharmacol. 2016, 173, 2752–2765. [Google Scholar] [CrossRef] [Green Version]
- Bohm, S.K.; Kong, W.; Bromme, D.; Smeekens, S.P.; Anderson, D.C.; Connolly, A.; Kahn, M.; Nelken, N.A.; Coughlin, S.R.; Payan, D.G.; et al. Molecular cloning, expression and potential functions of the human proteinase-activated receptor-2. Biochem. J. 1996, 314, 1009–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanke, T.; Ishiwata, H.; Kabeya, M.; Saka, M.; Doi, T.; Hattori, Y.; Kawabata, A.; Plevin, R. Binding of a highly potent protease-activated receptor-2 (PAR2) activating peptide, [3H]2-furoyl-LIGRL-NH2, to human PAR2. Br. J. Pharmacol. 2005, 145, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricks, T.K.; Trejo, J. Phosphorylation of protease-activated receptor-2 differentially regulates desensitization and internalization. J. Biol. Chem. 2009, 284, 34444–34457. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Lau, C.S.; Mathur, M.; Wang, P.; DeFea, K.A. Differential effects of beta-arrestins on the internalization, desensitization and ERK1/2 activation downstream of protease activated receptor-2. Am. J. Physiol. Cell Physiol. 2007, 293, C346–C357. [Google Scholar] [CrossRef] [PubMed]
- Amadesi, S.; Nie, J.; Vergnolle, N.; Cottrell, G.S.; Grady, E.F.; Trevisani, M.; Manni, C.; Geppetti, P.; McRoberts, J.A.; Ennes, H.; et al. Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia. J. Neurosci. 2004, 24, 4300–4312. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Moriyama, T.; Higashi, T.; Togashi, K.; Kobayashi, K.; Yamanaka, H.; Tominaga, M.; Noguchi, K. Proteinase-activated receptor 2-mediated potentiation of transient receptor potential vanilloid subfamily 1 activity reveals a mechanism for proteinase-induced inflammatory pain. J. Neurosci. 2004, 24, 4293–4299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzer, I.; Ray, S.; Schicker, K.; Boehm, S. Nociceptor Signalling through ion Channel Regulation via GPCRs. Int. J. Mol. Sci. 2019, 20, 2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFea, K.A.; Zalevsky, J.; Thoma, M.S.; Déry, O.; Mullins, R.D.; Bunnett, N.W. beta-arrestin-dependent endocytosis of proteinase-activated receptor 2 is required for intracellular targeting of activated ERK1/2. J. Cell Biol. 2000, 148, 1267–1281. [Google Scholar] [CrossRef]
- Stalheim, L.; Ding, Y.; Gullapalli, A.; Paing, M.M.; Wolfe, B.L.; Morris, D.R.; Trejo, J. Multiple independent functions of arrestins in the regulation of protease-activated receptor-2 signaling and trafficking. Mol. Pharmacol. 2005, 67, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Metcalf, M.; Bunnett, N.W. Biased signaling of protease-activated receptors. Front. Endocrinol. 2014, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Alier, K.A.; Endicott, J.A.; Stemkowski, P.L.; Cenac, N.; Cellars, L.; Chapman, K.; Andrade-Gordon, P.; Vergnolle, N.; Smith, P.A. Intrathecal administration of proteinase-activated receptor-2 agonists produces hyperalgesia by exciting the cell bodies of primary sensory neurons. J. Pharmacol. Exp. Ther. 2008, 324, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Spicarova, D.; Nerandzic, V.; Palecek, J. Update on the role of spinal cord TRPV1 receptors in pain modulation. Physiol. Res. 2014, 63, S225–S236. [Google Scholar] [CrossRef] [PubMed]
- Amadesi, S.; Cottrell, G.S.; Divino, L.; Chapman, K.; Grady, E.F.; Bautista, F.; Karanjia, R.; Barajas-Lopez, C.; Vanner, S.; Vergnolle, N.; et al. Protease-activated receptor 2 sensitizes TRPV1 by protein kinase Cepsilon- and A-dependent mechanisms in rats and mice. J. Physiol. 2006, 575, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Spicarova, D.; Palecek, J. The role of the TRPV1 endogenous agonist N-Oleoyldopamine in modulation of nociceptive signaling at the spinal cord level. J. Neurophysiol. 2009, 102, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicarova, D.; Palecek, J. Tumor necrosis factor alpha sensitizes spinal cord TRPV1 receptors to the endogenous agonist N-oleoyldopamine. J. Neuroinflamm. 2010, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicarova, D.; Adamek, P.; Kalynovska, N.; Mrozkova, P.; Palecek, J. TRPV1 receptor inhibition decreases CCL2-induced hyperalgesia. Neuropharmacology 2014, 81, 75–84. [Google Scholar] [CrossRef]
- Koetzner, L.; Gregory, J.A.; Yaksh, T.L. Intrathecal protease-activated receptor stimulation produces thermal hyperalgesia through spinal cyclooxygenase activity. J. Pharmacol. Exp. Ther. 2004, 311, 356–363. [Google Scholar] [CrossRef]
- Grant, A.D.; Cottrell, G.S.; Amadesi, S.; Trevisani, M.; Nicoletti, P.; Materazzi, S.; Altier, C.; Cenac, N.; Zamponi, G.W.; Bautista-Cruz, F.; et al. Protease-activated receptor 2 sensitizes the transient receptor potential vanilloid 4 ion channel to cause mechanical hyperalgesia in mice. J. Physiol. 2007, 578, 715–733. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, S.; Tominaga, M.; Yamamoto, S.; Fukuoka, T.; Higashi, T.; Kobayashi, K.; Obata, K.; Yamanaka, H.; Noguchi, K. Sensitization of TRPA1 by PAR2 contributes to the sensation of inflammatory pain. J. Clin. Investig. 2007, 117, 1979–1987. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, C.; Wang, Z.J. Proteinase-activated receptor 2 sensitizes transient receptor potential vanilloid 1, transient receptor potential vanilloid 4, and transient receptor potential ankyrin 1 in paclitaxel-induced neuropathic pain. Neuroscience 2011, 193, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Linley, J.E.; Rose, K.; Patil, M.; Robertson, B.; Akopian, A.N.; Gamper, N. Inhibition of M current in sensory neurons by exogenous proteases: A signaling pathway mediating inflammatory nociception. J. Neurosci. 2008, 28, 11240–11249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Shah, S.; Liu, Y.; Zhang, H.; Lees, M.; Fu, Z.; Lippiat, J.D.; Beech, D.J.; Sivaprasadarao, A.; Baldwin, S.A.; et al. Activation of the Cl− channel ANO1 by localized calcium signals in nociceptive sensory neurons requires coupling with the IP3 receptor. Sci. Signal. 2013, 6, ra73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrozkova, P.; Spicarova, D.; Palecek, J. Hypersensitivity Induced by Activation of Spinal Cord PAR2 Receptors Is Partially Mediated by TRPV1 Receptors. PLoS ONE 2016, 11, e0163991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrozkova, P.; Palecek, J.; Spicarova, D. The role of protease-activated receptor type 2 in nociceptive signaling and pain. Physiol. Res. 2016, 65, 357–367. [Google Scholar] [CrossRef]
- Huang, Z.; Tao, K.; Zhu, H.; Miao, X.; Wang, Z.; Yu, W.; Lu, Z. Acute PAR2 activation reduces GABAergic inhibition in the spinal dorsal horn. Brain Res. 2011, 1425, 20–26. [Google Scholar] [CrossRef]
- Noorbakhsh, F.; Tsutsui, S.; Vergnolle, N.; Boven, L.A.; Shariat, N.; Vodjgani, M.; Warren, K.G.; Andrade-Gordon, P.; Hollenberg, M.D.; Power, C. Proteinase-activated receptor 2 modulates neuroinflammation in experimental autoimmune encephalomyelitis and multiple sclerosis. J. Exp. Med. 2006, 203, 425–435. [Google Scholar] [CrossRef]
- Cenac, N.; Coelho, A.M.; Nguyen, C.; Compton, S.; Andrade-Gordon, P.; MacNaughton, W.K.; Wallace, J.L.; Hollenberg, M.D.; Bunnett, N.W.; Garcia-Villar, R.; et al. Induction of intestinal inflammation in mouse by activation of proteinase-activated receptor-2. Am. J. Pathol. 2002, 161, 1903–1915. [Google Scholar] [CrossRef] [Green Version]
- Radulovic, M.; Yoon, H.; Wu, J.; Mustafa, K.; Fehlings, M.G.; Scarisbrick, I.A. Genetic targeting of protease activated receptor 2 reduces inflammatory astrogliosis and improves recovery of function after spinal cord injury. Neurobiol. Dis. 2015, 83, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Ferrell, W.R.; Lockhart, J.C.; Kelso, E.B.; Dunning, L.; Plevin, R.; Meek, S.E.; Smith, A.J.; Hunter, G.D.; McLean, J.S.; McGarry, F.; et al. Essential role for proteinase-activated receptor-2 in arthritis. J. Clin. Investig. 2003, 111, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Vergnolle, N.; Bunnett, N.W.; Sharkey, K.A.; Brussee, V.; Compton, S.J.; Grady, E.F.; Cirino, G.; Gerard, N.; Basbaum, A.I.; Andrade-Gordon, P.; et al. Proteinase-activated receptor-2 and hyperalgesia: A novel pain pathway. Nat. Med. 2001, 7, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N. Protease-activated receptors as therapeutic targets in visceral pain. Curr. Neuropharmacol. 2013, 11, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, K.T.; Mohammad, H.; Sweitzer, S.M. Protein kinase C in pain: Involvement of multiple isoforms. Pharmacol. Res. 2007, 55, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honore, P.; Wismer, C.T.; Mikusa, J.; Zhu, C.Z.; Zhong, C.; Gauvin, D.M.; Gomtsyan, A.; El Kouhen, R.; Lee, C.H.; Marsh, K.; et al. A-425619 [1-isoquinolin-5-yl-3-(4-trifluoromethyl-benzyl)-urea], a novel transient receptor potential type V1 receptor antagonist, relieves pathophysiological pain associated with inflammation and tissue injury in rats. J. Pharmacol. Exp. Ther. 2005, 314, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Honore, P.; Zhong, C.; Gauvin, D.; Mikusa, J.; Hernandez, G.; Chandran, P.; Gomtsyan, A.; Brown, B.; Bayburt, E.K.; et al. TRPV1 receptors in the CNS play a key role in broad-spectrum analgesia of TRPV1 antagonists. J. Neurosci. 2006, 26, 9385–9393. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Yang, F.; Luo, H.; Liu, F.Y.; Han, J.S.; Xing, G.G.; Wan, Y. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund’s adjuvant. Mol. Pain 2008, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chen, S.R.; Chen, H.; Pan, H.L. Endogenous transient receptor potential ankyrin 1 and vanilloid 1 activity potentiates glutamatergic input to spinal lamina I neurons in inflammatory pain. J. Neurochem. 2019, 149, 381–398. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, Y.; Tian, Q.; Deng, Q.; Zhou, T.; Liu, Q.; Mei, K.; Liu, H.; Ma, R.; Ding, Y.; et al. TRPV1 SUMOylation regulates nociceptive signaling in models of inflammatory pain. Nat. Commun. 2018, 9, 1529. [Google Scholar] [CrossRef]
- Gregus, A.M.; Doolen, S.; Dumlao, D.S.; Buczynski, M.W.; Takasusuki, T.; Fitzsimmons, B.L.; Hua, X.Y.; Taylor, B.K.; Dennis, E.A.; Yaksh, T.L. Spinal 12-lipoxygenase-derived hepoxilin A3 contributes to inflammatory hyperalgesia via activation of TRPV1 and TRPA1 receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 6721–6726. [Google Scholar] [CrossRef] [Green Version]
- Tohda, C.; Sasaki, M.; Konemura, T.; Sasamura, T.; Itoh, M.; Kuraishi, Y. Axonal transport of VR1 capsaicin receptor mRNA in primary afferents and its participation in inflammation-induced increase in capsaicin sensitivity. J. Neurochem. 2001, 76, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Abooj, M.; Bishnoi, M.; Bosgraaf, C.A.; Premkumar, L.S. Changes in Spinal Cord Following Inflammatory and Neuropathic Pain and the Effectiveness of Resiniferatoxin. Open Pain J. 2016, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ren, K.; Dubner, R. Inflammatory Models of Pain and Hyperalgesia. ILAR J. 1999, 40, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Soh, U.J.; Dores, M.R.; Chen, B.; Trejo, J. Signal transduction by protease-activated receptors. Br. J. Pharmacol. 2010, 160, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesare, P.; Moriondo, A.; Vellani, V.; McNaughton, P.A. Ion channels gated by heat. Proc. Natl. Acad. Sci. USA 1999, 96, 7658–7663. [Google Scholar] [CrossRef] [Green Version]
- Ding-Pfennigdorff, D.; Averbeck, B.; Michaelis, M. Stimulation of PAR-2 excites and sensitizes rat cutaneous C-nociceptors to heat. Neuroreport 2004, 15, 2071–2075. [Google Scholar] [CrossRef]
- Bao, Y.; Hou, W.; Yang, L.; Liu, R.; Gao, Y.; Kong, X.; Shi, Z.; Li, W.; Zheng, H.; Jiang, S.; et al. Increased expression of protease-activated receptor 2 and 4 within dorsal root ganglia in a rat model of bone cancer pain. J. Mol. Neurosci. 2015, 55, 706–714. [Google Scholar] [CrossRef]
- Bao, Y.; Hou, W.; Liu, R.; Gao, Y.; Kong, X.; Yang, L.; Shi, Z.; Li, W.; Zheng, H.; Jiang, S.; et al. PAR2-mediated upregulation of BDNF contributes to central sensitization in bone cancer pain. Mol. Pain 2014, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Zhang, Z.F.; Liao, M.F.; Yao, W.L.; Wang, J.; Wang, X.R. Blocking PAR2 attenuates oxaliplatin-induced neuropathic pain via TRPV1 and releases of substance P and CGRP in superficial dorsal horn of spinal cord. J. Neurol. Sci. 2015, 352, 62–67. [Google Scholar] [CrossRef]
- Chen, D.; Liu, N.; Li, M.; Liang, S. Blocking PAR2 Alleviates Bladder Pain and Hyperactivity via TRPA1 Signal. Transl. Neurosci. 2016, 7, 133–138. [Google Scholar] [CrossRef]
- Lucena, F.; McDougall, J.J. Pain responses to protease-activated receptor-2 stimulation in the spinal cord of naïve and arthritic rats. Neurosci. Lett. 2020, 739, 135391. [Google Scholar] [CrossRef] [PubMed]
- Spicarova, D.; Nerandzic, V.; Palecek, J. Modulation of spinal cord synaptic activity by tumor necrosis factor α in a model of peripheral neuropathy. J. Neuroinflamm. 2011, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchytilova, E.; Spicarova, D.; Palecek, J. TRPV1 antagonist attenuates postoperative hypersensitivity by central and peripheral mechanisms. Mol. Pain 2014, 10, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrozkova, P.; Spicarova, D.; Palecek, J. Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats. Int. J. Mol. Sci. 2021, 22, 991. https://doi.org/10.3390/ijms22030991
Mrozkova P, Spicarova D, Palecek J. Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats. International Journal of Molecular Sciences. 2021; 22(3):991. https://doi.org/10.3390/ijms22030991
Chicago/Turabian StyleMrozkova, Petra, Diana Spicarova, and Jiri Palecek. 2021. "Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats" International Journal of Molecular Sciences 22, no. 3: 991. https://doi.org/10.3390/ijms22030991
APA StyleMrozkova, P., Spicarova, D., & Palecek, J. (2021). Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats. International Journal of Molecular Sciences, 22(3), 991. https://doi.org/10.3390/ijms22030991