
Viral Infection and Cardiovascular Disease: Implications for the Molecular Basis of COVID-19 Pathogenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
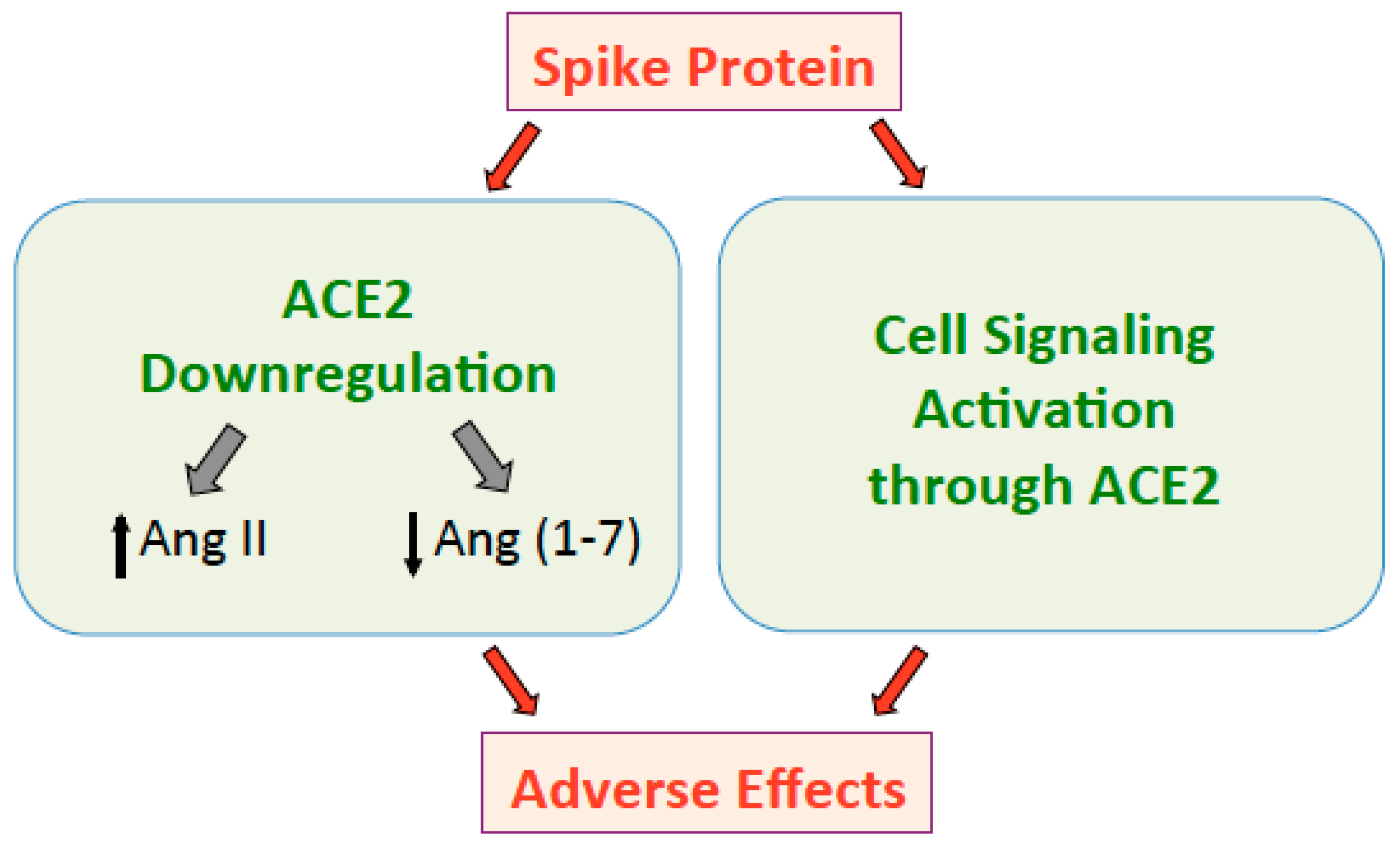
2. SARS-CoV-2: The Global Problem
3. Effects of Other Respiratory Viruses on the Cardiovascular System
3.1. SARS-CoV-1
3.2. Middle East Respiratory Syndrome-Related Coronavirus (MERS-CoV)
3.3. Influenza
4. Effects of Non-Respiratory Viruses on the Cardiovascular System
4.1. Dengue Virus
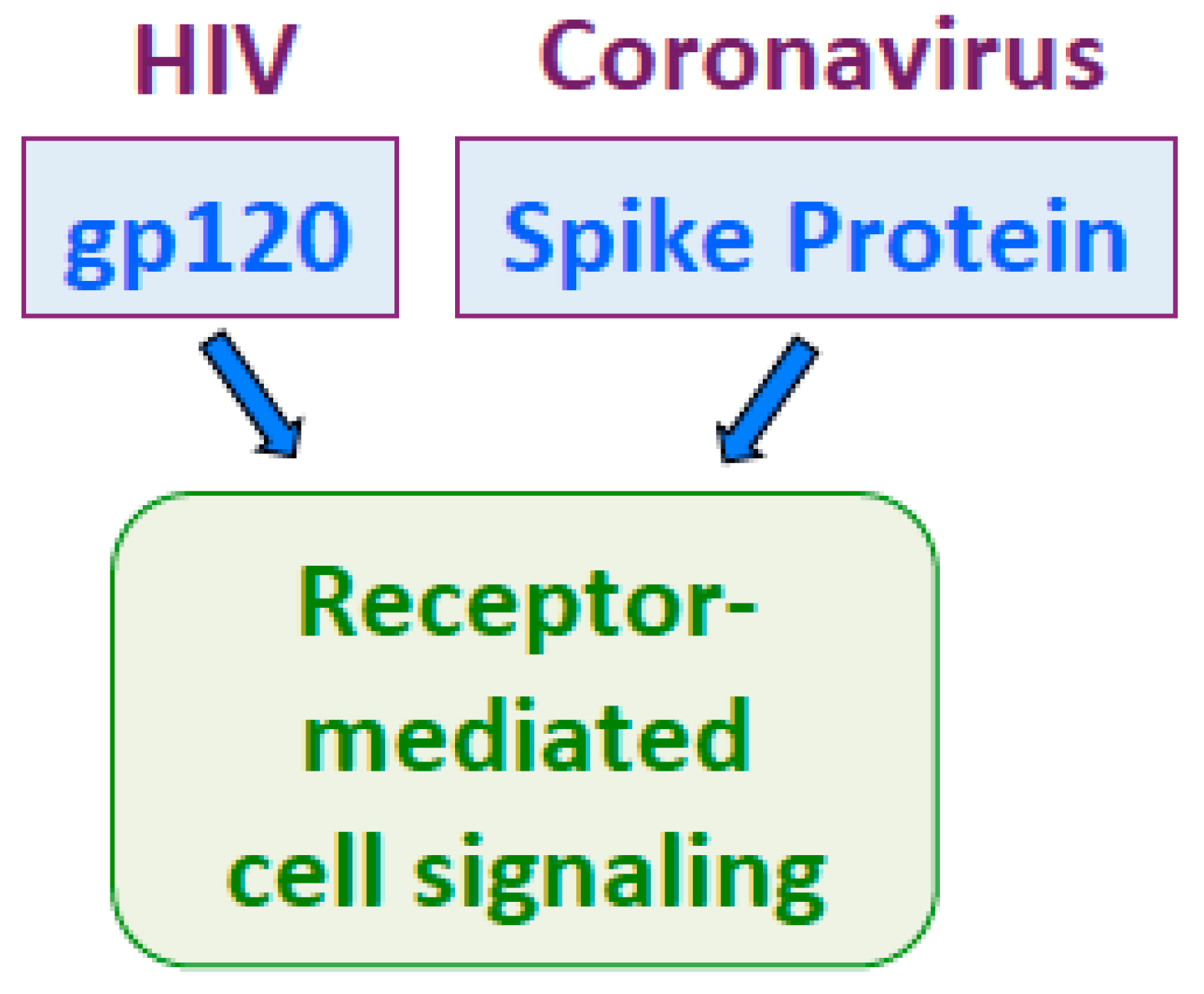
4.2. Human Immunodeficiency Virus (HIV)
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk factors associated with acute respiratory distress syndrome and death in patients with coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, e200994. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: Celebrating the 20th anniversary of the discovery of ACE. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Warner, F.J.; Smith, A.I.; Hooper, N.M.; Turner, A.J. Angiotensin-converting enzyme-2: A molecular and cellular perspective. Cell. Mol. Life Sci. 2004, 61, 2704–2713. [Google Scholar] [CrossRef]
- Pacurari, M.; Kafoury, R.; Tchounwou, P.B.; Ndebele, K. The renin-angiotensin-aldosterone system in vascular inflammation and remodeling. Int. J. Inflam. 2014, 2014, 689360. [Google Scholar] [CrossRef]
- Santos, R.A.S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/Angiotensin-(1-7)/MAS axis of the renin-angiotensin system: Focus on Angiotensin-(1-7). Physiol. Rev. 2018, 98, 505–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Mendoza-Torres, E.; Oyarzún, A.; Mondaca-Ruff, D.; Azocar, A.; Castro, P.F.; Jalil, J.E.; Chiong, M.; Lavandero, S.; Paz Ocaranza, M. ACE2 and vasoactive peptides: Novel players in cardiovascular/renal remodeling and hypertension. Adv. Cardiovasc. Dis. 2015, 9, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Crackower, M.A.; Sarao, R.; Oudit, G.Y.; Yagil, C.; Kozieradzki, I.; Scanga, S.E.; Oliveira-dos-Santos, A.J.; da Costa, J.; Zhang, L.; Pei, Y.; et al. Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature 2002, 417, 822–828. [Google Scholar] [CrossRef]
- Gurley, S.B.; Allred, A.; Le, T.H.; Griffiths, R.; Mao, L.; Philip, N.; Haystead, T.A.; Donoghue, M.; Breitbart, R.E.; Acton, S.L.; et al. Altered blood pressure responses and normal cardiac phenotype in ACE2-null mice. J. Clin. Investig. 2006, 116, 2218–2225. [Google Scholar] [CrossRef] [Green Version]
- Sriramula, S.; Cardinale, J.P.; Lazartigues, E.; Francis, J. ACE2 overexpression in the paraventricular nucleus attenuates angiotensin II-induced hypertension. Cardiovasc. Res. 2011, 92, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raizada, M.K.; Ferreira, A.J. ACE2: A new target for cardiovascular disease therapeutics. J. Cardiovasc. Pharm. 2007, 50, 112–119. [Google Scholar] [CrossRef]
- Úri, K.; Fagyas, M.; Kertész, A.; Borbély, A.; Jenei, C.; Bene, O.; Csanádi, Z.; Paulus, W.J.; Édes, I.; Papp, Z.; et al. Circulating ACE2 activity correlates with cardiovascular disease development. J. Renin Angiotensin Aldosterone Syst. 2016, 17, 1470320316668435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Freire, C.; Vázquez, J.; Correa de Adjounian, M.F.; Ferrari, M.F.R.; Yuan, L.; Silver, X.; Torres, R.; Raizada, M.K. ACE2 gene transfer attenuates hypertension-linked pathophysiological changes in the SHR. Physiol. Genom. 2006, 27, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Nishiga, M.; Wang, D.W.; Han, Y.; Lewis, D.B.; Wu, J.C. COVID-19 and cardiovascular disease: From basic mechanisms to clinical perspectives. Nat. Rev. Cardiol. 2020, 17, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Clerkin, K.J.; Fried, J.A.; Raikhelkar, J.; Sayer, G.; Griffin, J.M.; Masoumi, A.; Jain, S.S.; Burkhoff, D.; Kumaraiah, D.; Rabbani, L.; et al. COVID-19 and cardiovascular disease. Circulation 2020, 141, 1648–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, M. Cardiovascular disease and COVID-19. Diabetes Metab. Syndr. 2020, 14, 247–250. [Google Scholar] [CrossRef]
- Wang, L.F.; Shi, Z.; Zhang, S.; Field, H.; Daszak, P.; Eaton, B.T. Review of bats and SARS. Emerg. Infect. Dis. 2006, 12, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 224. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.J.; Hiscox, J.A.; Hooper, N.M. ACE2: From vasopeptidase to SARS virus receptor. Trends Pharmacol. Sci. 2004, 25, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, A.; Karpinski, T.M. SARS-CoV, MERS-CoV, SARS-CoV-2 comparison of three emerging coronaviruses. Jundishapur. J. Microbiol. 2020, 13, e103744. [Google Scholar] [CrossRef]
- Rossi, G.A.; Sacco, O.; Mancino, E.; Cristiani, L.; Midulla, F. Differences and similarities between SARS-CoV and SARS-CoV-2: Spike receptor-binding domain recognition and host cell infection with support of cellular serine proteases. Infection 2020, 48, 665–669. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, H.; Shen, H.; Li, Z.; Geng, J.; Han, H.; Cai, J.; Li, X.; Kang, W.; Weng, D.; et al. The clinical pathology of severe acute respiratory syndrome (SARS): A report from China. J. Pathol. 2003, 200, 282–289. [Google Scholar] [CrossRef]
- Hwang, D.M.; Chamberlain, D.W.; Poutanen, S.M.; Low, D.E.; Asa, S.L.; Butany, J. Pulmonary pathology of severe acute respiratory syndrome in Toronto. Mod. Pathol. 2005, 18, 1–10. [Google Scholar] [CrossRef]
- Xiang-Hua, Y.; Le-Min, W.; Ai-Bin, L.; Zhu, G.; Riquan, L.; Xu-You, Z.; Wei-Wei, R.; Ye-Nan, W. Severe acute respiratory syndrome and venous thromboembolism in multiple organs. Am. J. Respir. Crit. Care Med. 2010, 182, 436–437. [Google Scholar] [CrossRef]
- Liu, Z.H.; Wei, R.; Wu, Y.P.; Lisman, T.; Wang, Z.X.; Han, J.J.; Ren, D.L.; Chen, B.; Xia, Z.L.; Chen, B.; et al. Elevated plasma tissue-type plasminogen activator (t-PA) and soluble thrombomodulin in patients suffering from severe acute respiratory syndrome (SARS) as a possible index for prognosis and treatment strategy. Biomed. Environ. Sci. 2005, 18, 260–264. [Google Scholar]
- Li, S.S.; Cheng, C.W.; Fu, C.L.; Chan, Y.H.; Lee, M.P.; Chan, J.W.; Yiu, S.F. Left ventricular performance in patients with severe acute respiratory syndrome: A 30-day echocardiographic follow-up study. Circulation 2003, 108, 1798–1803. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Kassiri, Z.; Jiang, C.; Liu, P.P.; Poutanen, S.M.; Penninger, J.M.; Butany, J. SARS-coronavirus modulation of myocardial ACE2 expression and inflammation in patients with SARS. Eur. J. Clin. Investig. 2009, 39, 618–625. [Google Scholar] [CrossRef]
- Yu, C.-M.; Wong, R.S.-M.; Wu, E.B.; Kong, S.-L.; Wong, J.; Yip, G.W.-K.; Soo, Y.O.Y.; Chiu, M.L.S.; Chan, Y.-S.; Hui, D.; et al. Cardiovascular complications of severe acute respiratory syndrome. Postgrad. Med. J. 2006, 82, 140–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Rao, S.; Jiang, C.; Penninger, J.M. Lessons from SARS: Control of acute lung failure by the SARS receptor ACE2. J. Mol. Med. 2006, 84, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Kuba, K.; Penninger, J.M. Angiotensin-converting enzyme 2 in acute respiratory distress syndrome. Cell. Mol. Life Sci. 2007, 64, 2006–2012. [Google Scholar] [CrossRef]
- Chen, P.I.; Chang, S.C.; Wu, H.Y.; Yu, T.C.; Wei, W.C.; Lin, S.; Chien, C.L.; Chang, M.F. Upregulation of the chemokine (C-C motif) ligand 2 via a severe acute respiratory syndrome coronavirus spike-ACE2 signaling. J. Virol. 2010, 84, 7703–7712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.J.; Nikolaienko, S.I.; Dibrova, V.A.; Dibrova, Y.V.; Vasylyk, V.M.; Novikov, M.Y.; Shults, N.V.; Gychka, S.G. SARS-CoV-2 spike protein-mediated cell signaling in lung vascular cells. Vascul. Pharmacol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rabaan, A.A.; Al-Ahmed, S.H.; Sah, R.; Alqumber, M.A.; Haque, S.; Patel, S.K.; Pathak, M.; Tiwari, R.; Yatoo, M.I.; Haq, A.U.; et al. MERS-CoV: Epidemiology, molecular dynamics, therapeutics, and future challenges. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 8. [Google Scholar] [CrossRef]
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F.; et al. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, R.; Nishiura, H.; Kutsuna, S.; Hayakawa, K.; Ohmagari, N. Clinical determinants of the severity of Middle East respiratory syndrome (MERS): A systematic review and meta-analysis. BMC Public Health 2016, 16, 1203. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, N.; Shaib, H. Middle East respiratory syndrome coronavirus (MERS-CoV): A review. Germs 2019, 9, 35–42. [Google Scholar] [CrossRef]
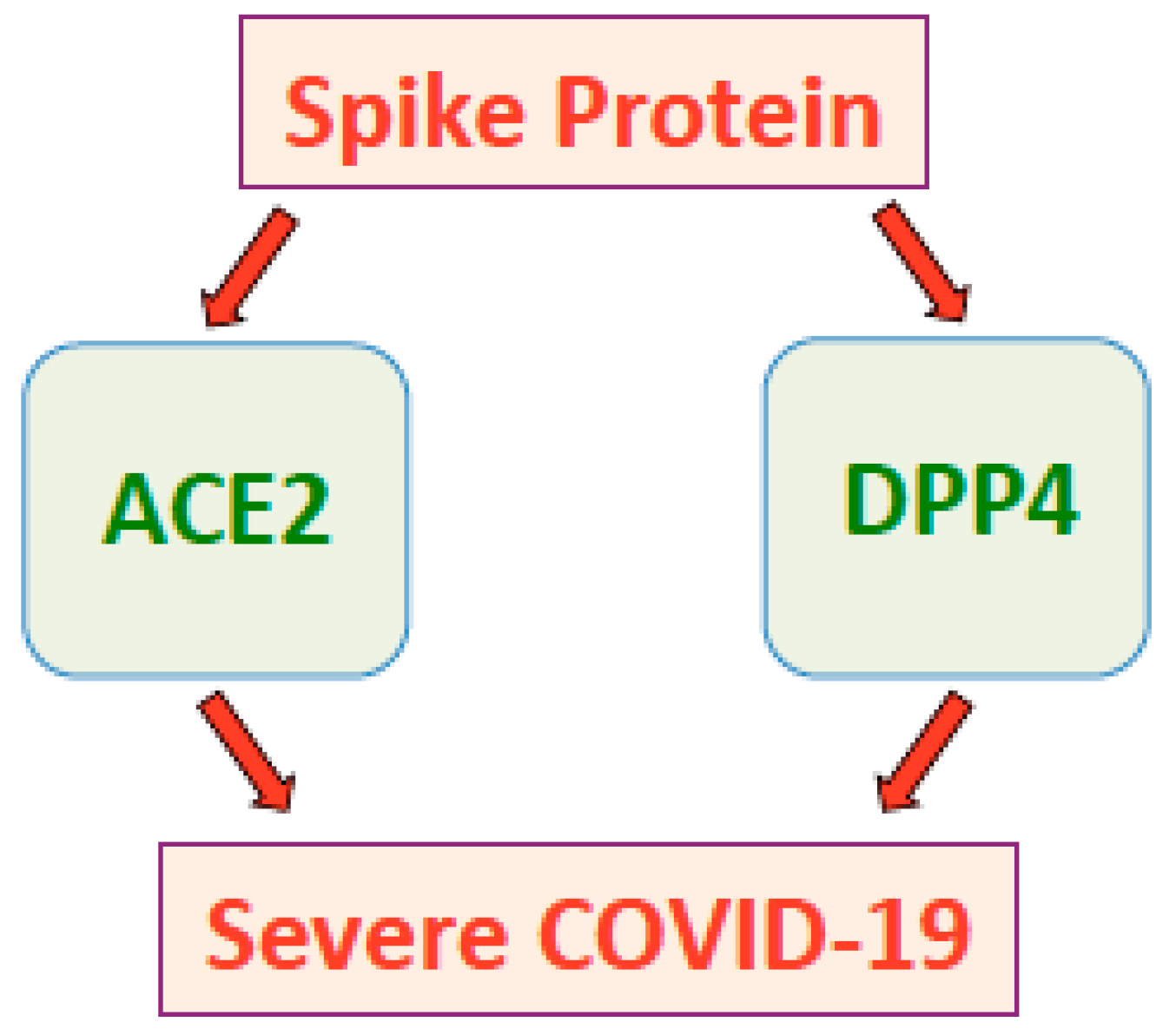
- Bassendine, M.F.; Bridge, S.H.; McCaughan, G.W.; Gorrell, M.D. COVID-19 and comorbidities: A role for dipeptidyl peptidase 4 (DPP4) in disease severity? J. Diabetes 2020, 12, 649–658. [Google Scholar] [CrossRef]
- Kosik, I.; Yewdell, J.W. Influenza hemagglutinin and neuraminidase: Yin–Yang proteins coevolving to thwart immunity. Viruses 2019, 11, 346. [Google Scholar] [CrossRef] [Green Version]
- Kalil, A.C.; Thomas, P.G. Influenza virus-related critical illness: Pathophysiology and epidemiology. Crit. Care 2019, 23, 258. [Google Scholar] [CrossRef] [Green Version]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Bronchoalveolar interferon-α, tumor necrosis factor-α, interleukin-1, and inflammation during acute influenza in pigs: A possible model for humans? J. Infect. Dis. 1998, 177, 1076–1079. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Le, T.Q.; Kurihara, N.; Chida, J.; Cisse, Y.; Yano, M.; Kido, H. Influenza virus-cytokine-protease cycle in the pathogenesis of vascular hyperpermeability in severe influenza. J. Infect. Dis. 2010, 202, 991–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.A.; Phoon, M.C.; van Rooijen, N.; Chow, V.T. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. Am. J. Pathol. 2011, 179, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Schmolke, M.; Viemann, D.; Roth, J.; Ludwig, S. Essential impact of NF-κB signaling on the H5N1 influenza A virus-induced transcriptome. J. Immunol. 2009, 183, 5180–5189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijaro, J.R.; Walsh, K.B.; Cahalan, S.; Fremgen, D.M.; Roberts, E.; Scott, F.; Martinborough, E.; Peach, R.; Oldstone, M.B.; Rosen, H. Endothelial cells are central orchestrators of cytokine amplification during influenza virus infection. Cell 2011, 146, 980–991. [Google Scholar] [CrossRef] [Green Version]
- Bunce, P.E.; High, S.M.; Nadjafi, M.; Stanley, K.; Liles, W.C.; Christian, M.D. Pandemic H1N1 influenza infection and vascular thrombosis. Clin. Infect. Dis. 2011, 52, e14–e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinsky, D.J.; Naka, Y.; Liao, H.; Oz, M.C.; Wagner, D.D.; Mayadas, T.N.; Johnson, R.C.; Hynes, R.O.; Heath, M.; Lawson, C.A.; et al. Hypoxia-induced exocytosis of endothelial cell Weibel-Palade bodies. A mechanism for rapid neutrophil recruitment after cardiac preservation. J. Clin. Investig. 1996, 97, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M.G.; Gamage, A.; Zyla, R.; Armstrong, S.M.; Advani, S.; Advani, A.; Wang, C.; Lee, W.L. Influenza virus infection induces platelet-endothelial adhesion which contributes to lung injury. J. Virol. 2015, 90, 1812–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madjid, M.; Naghavi, M.; Litovsky, S.; Casscells, S.W. Influenza and cardiovascular disease: A new opportunity for prevention and the need for further studies. Circulation 2003, 108, 2730–2736. [Google Scholar] [CrossRef] [Green Version]
- Kodama, M. Influenza myocarditis. Circ. J. 2010, 74, 2060–2061. [Google Scholar] [CrossRef] [PubMed]
- Sellers, S.A.; Hagan, R.S.; Hayden, F.G.; Fischer, W.A. The hidden burden of influenza: A review of the extra-pulmonary complications of influenza infection. Influenza Other Respir. Viruses 2017, 11, 372–393. [Google Scholar] [CrossRef]
- Brown, S.M.; Pittman, J.; Miller Iii, R.R.; Horton, K.D.; Markewitz, B.; Hirshberg, E.; Jones, J.; Grissom, C.K. Right and left heart failure in severe H1N1 influenza A infection. Eur. Respir. J. 2011, 37, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, A.; Lucero-Obusan, C.; Schirmer, P.; Winston, C.; Holodniy, M. Acute cardiac injury events ≤30 days after laboratory-confirmed influenza virus infection among U.S. veterans, 2010–2012. BMC Cardiovasc. Disord. 2015, 15, 109. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, N.; Tang, J.; Liu, S.; Luo, D.; Duan, Q.; Wang, X. Downregulation of angiotensin-converting enzyme 2 by the neuraminidase protein of influenza A (H1N1) virus. Virus Res. 2014, 185, 64–71. [Google Scholar] [CrossRef]
- Zou, Z.; Yan, Y.; Shu, Y.; Gao, R.; Sun, Y.; Li, X.; Ju, X.; Liang, Z.; Liu, Q.; Zhao, Y.; et al. Angiotensin-converting enzyme 2 protects from lethal avian influenza A H5N1 infections. Nat. Commun. 2014, 5, 3594. [Google Scholar] [CrossRef]
- Yang, P.; Gu, H.; Zhao, Z.; Wang, W.; Cao, B.; Lai, C.; Yang, X.; Zhang, L.; Duan, Y.; Zhang, S.; et al. Angiotensin-converting enzyme 2 (ACE2) mediates influenza H7N9 virus-induced acute lung injury. Sci. Rep. 2014, 4, 7027. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Guo, J.; Zou, Z.; Liu, J.; Cao, B.; Zhang, S.; Li, H.; Wang, W.; Sheng, M.; Liu, S.; et al. Angiotensin II plasma levels are linked to disease severity and predict fatal outcomes in H7N9-infected patients. Nat. Commun. 2014, 5, 3595. [Google Scholar] [CrossRef]
- Suzuki, Y.J. The viral protein fragment theory of COVID-19 pathogenesis. Med. Hypotheses 2020, 144, 110267. [Google Scholar] [CrossRef]
- Chen, H.-R.; Lai, Y.-C.; Yeh, T.-M. Dengue virus non-structural protein 1: A pathogenic factor, therapeutic target, and vaccine candidate. J. Biomed. Sci. 2018, 25, 58. [Google Scholar] [CrossRef] [PubMed]
- Henrina, J.; Putra, I.C.S.; Lawrensia, S.; Handoyono, Q.F.; Cahyadi, A. Coronavirus Disease of 2019: A mimicker of dengue infection? SN Compr. Clin. Med. 2020, 2, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-K.; Lee, W.-H.; Liu, J.-W.; Yang, K.D. Acute myocarditis in dengue hemorrhagic fever: A case report and review of cardiac complications in dengue-affected patients. Int. J. Infect. Dis. 2010, 14, e919–e922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-M.; Huang, Y.-K.; Su, Y.-C.; Kao, C.-H. Risk of stroke in patients with dengue fever: A population-based cohort study. CMAJ 2018, 190, E285–E290. [Google Scholar] [CrossRef] [Green Version]
- da Costa, P.S.; Ribeiro, G.M.; Junior, C.S.; da Costa Campos, L. Severe thrombotic events associated with dengue fever, Brazil. Am. J. Trop. Med. Hyg. 2012, 87, 741–742. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Murao, L.A.; Lan, N.T.P.; Huy, N.T.; Huong, V.T.Q.; Thuy, T.T.; Tham, V.D.; Nga, C.T.P.; Ha, T.T.N.; Ohmoto, Y.; et al. Association of mast cell-derived VEGF and proteases in Dengue shock syndrome. PLoS Negl. Trop. Dis. 2012, 6, e1505. [Google Scholar] [CrossRef] [Green Version]
- St John, A.L.; Rathore, A.P.; Raghavan, B.; Ng, M.-L.; Abraham, S.N. Contributions of mast cells and vasoactive products, leukotrienes and chymase, to dengue virus-induced vascular leakage. eLife 2013, 2, e00481. [Google Scholar] [CrossRef]
- Brown, M.G.; Hermann, L.L.; Issekutz, A.C.; Marshall, J.S.; Rowter, D.; Al-Afif, A.; Anderson, R. Dengue virus infection of mast cells triggers endothelial cell activation. J. Virol. 2011, 85, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Jeewandara, C.; Gomes, L.; Wickramasinghe, N.; Gutowska-Owsiak, D.; Waithe, D.; Paranavitane, S.A.; Shyamali, N.L.; Ogg, G.S.; Malavige, G.N. Platelet activating factor contributes to vascular leak in acute dengue infection. PLoS Negl. Trop. Dis. 2015, 9, e0003459. [Google Scholar] [CrossRef]
- Michels, M.; van der Ven, A.J.; Djamiatun, K.; Fijnheer, R.; de Groot, P.G.; Griffioen, A.W.; Sebastian, S.; Faradz, S.M.; de Mast, Q. Imbalance of angiopoietin-1 and angiopoetin-2 in severe dengue and relationship with thrombocytopenia, endothelial activation, and vascular stability. Am. J. Trop. Med. Hyg. 2012, 87, 943–946. [Google Scholar] [CrossRef] [Green Version]
- Van de Weg, C.A.; Pannuti, C.S.; van den Ham, H.J.; de Araújo, E.S.; Boas, L.S.; Felix, A.C.; Carvalho, K.I.; Levi, J.E.; Romano, C.M.; Centrone, C.C.; et al. Serum angiopoietin-2 and soluble VEGF receptor 2 are surrogate markers for plasma leakage in patients with acute dengue virus infection. J. Clin. Virol. 2014, 60, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.; Zhang, Q.; Liu, W.; Wang, W.; Yu, Z.; Lao, Z.; Zhang, W.; Shen, M.; Wan, P.; Xiao, F.; et al. Dengue virus infection activates interleukin-1β to induce tissue injury and vascular leakage. Front Microbiol. 2019, 10, 2637. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, D.F.; te Velde, A.A. Severe COVID-19: NLRP3 inflammasome dysregulated. Front Immunol. 2020, 11, 1580. [Google Scholar] [CrossRef]
- Libraty, D.H.; Young, P.R.; Pickering, D.; Endy, T.P.; Kalayanarooj, S.; Green, S.; Vaughn, D.W.; Nisalak, A.; Ennis, F.A.; Rothman, A.L. High circulating levels of the dengue virus nonstructural protein NS1 early in dengue illness correlate with the development of dengue hemorrhagic fever. J. Infect. Dis. 2002, 186, 1165–1168. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.R.; Puerta-Guardo, H.; Killingbeck, S.S.; Glasner, D.R.; Hopkins, K.; Harris, E. Dengue virus NS1 triggers endothelial permeability and vascular leak that is prevented by NS1 vaccination. Sci. Transl. Med. 2015, 7, 304ra141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, Y.P.; Mastrullo, V.; Maselli, D.; Butsabong, T.; Madeddu, P.; Maringer, K.; Campagnolo, P. A critical role for perivascular cells in amplifying vascular leakage induced by dengue virus nonstructural protein 1. mSphere 2020, 5, e00258-20. [Google Scholar] [CrossRef] [PubMed]
- Barbachano-Guerrero, A.; Endy, T.P.; King, C.A. Dengue virus non-structural protein 1 activates the p38 MAPK pathway to decrease barrier integrity in primary human endothelial cells. J. Gen. Virol. 2020, 101, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; De Clercq, E. HIV genome-wide protein associations: A review of 30 years of research. Microbiol. Mol. Biol. Rev. 2016, 80, 679–731. [Google Scholar] [CrossRef] [Green Version]
- Escaut, L.; Monsuez, J.J.; Chironi, G.; Merad, M.; Teicher, E.; Smadja, D.; Simon, A.; Vittecoq, D. Coronary artery disease in HIV infected patients. Intensive Care Med. 2003, 29, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Periard, D.; Cavassini, M.; Taffé, P.; Chevalley, M.; Senn, L.; Chapuis-Taillard, C.; de Vallière, S.; Hayoz, D.; Tarr, P.E. Swiss HIV Cohort Study. High prevalence of peripheral arterial disease in HIV-infected persons. Clin. Infect. Dis. 2008, 46, 761–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenstein, K.A.; Armon, C.; Buchacz, K.; Chmiel, J.S.; Buckner, K.; Tedaldi, E.M.; Wood, K.; Holmberg, S.D.; Brooks, J.T. HIV Outpatient Study (HOPS) Investigators. Low CD4+ T cell count is a risk factor for cardiovascular disease events in the HIV outpatient study. Clin. Infect. Dis. 2010, 51, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triant, V.A.; Lee, H.; Hadigan, C.; Grinspoon, S.K. Increased acute myocardial infarction rates and cardiovascular risk factors among patients with human immunodeficiency virus disease. J. Clin. Endocrinol. Metab. 2007, 92, 2506–2512. [Google Scholar] [CrossRef]
- Post, W.S.; Budoff, M.; Kingsley, L.; Palella, F.J., Jr.; Witt, M.D.; Li, X.; George, R.T.; Brown, T.T.; Jacobson, L.P. Associations between HIV infection and subclinical coronary atherosclerosis. Ann. Intern. Med. 2014, 160, 458–467. [Google Scholar] [CrossRef]
- Hsia, J.; Ross, A.M. Pericardial effusion and pericardiocentesis in human immunodeficiency virus infection. Am. J. Cardiol. 1994, 74, 94–96. [Google Scholar] [CrossRef]
- Currie, P.F.; Goldman, J.H.; Caforio, A.L.; Jacob, A.J.; Baig, M.K.; Brettle, R.P.; Haven, A.J.; Boon, N.A.; McKenna, W.J. Cardiac autoimmunity in HIV related heart muscle disease. Heart 1998, 79, 599–604. [Google Scholar] [CrossRef]
- Levy, W.S.; Simon, G.L.; Rios, J.C.; Ross, A.M. Prevalence of cardiac abnormalities in human immunodeficiency virus infection. Am. J. Cardiol. 1989, 63, 86–89. [Google Scholar] [CrossRef]
- Currie, P.F.; Jacob, A.J.; Foreman, A.R.; Elton, R.A.; Brettle, R.P.; Boon, N.A. Heart muscle disease related to HIV infection: Prognostic implications. BMJ 1994, 309, 1605–1607. [Google Scholar] [CrossRef] [Green Version]
- Paton, P.; Tabib, A.; Loire, R.; Tete, R. Coronary artery lesions and human immunodeficiency virus infection. Res. Virol. 1993, 144, 225–231. [Google Scholar] [CrossRef]
- Gherardi, R.; Belec, L.; Mhiri, C.; Gray, F.; Lescs, M.C.; Sobel, A.; Guillevin, L.; Wechsler, J. The spectrum of vasculitis in human immunodeficiency virus–infected patients. A clinicopathologic evaluation. Arthritis Rheum. 1993, 36, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Aoun, S.; Ramos, E. Hypertension in the HIV-infected patient. Curr. Hypertens. Rep. 2000, 2, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Isasti, G.; Moreno, T.; Pérez, I.; Cabrera, F.; Palacios, R.; Santos, J. High prevalence of pulmonary arterial hypertension in a cohort of asymptomatic HIV-infected patients. AIDS Res. Hum. Retrovir. 2013, 29, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.J.; Khan, I.A.; Mehta, R.N.; Sepkowitz, D.A. HIV-Related pulmonary hypertension: Analytic review of 131 cases. Chest 2000, 118, 1133–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellicelli, A.; Barbaro, G.; Palmieri, F.; Girardi, E.; D’Ambrosio, C.; Rianda, A.; Barbarini, G.; Frigiotti, D.; Borgia, M.C.; Petrosillo, N. Primary pulmonary hypertension in HIV patients: A systematic review. Angiology 2001, 52, 31–41. [Google Scholar] [CrossRef]
- Sitbon, O.; Lascoux-Combe, C.; Delfraissy, J.-F.; Yeni, P.G.; Raffi, F.; De Zuttere, D.; Gressin, V.; Clerson, P.; Sereni, D.; Simonneau, G. Prevalence of HIV-related pulmonary arterial hypertension in the current antiretroviral therapy era. Am. J. Respir. Crit. Care Med. 2008, 177, 108–113. [Google Scholar] [CrossRef]
- Bugnone, A.N.; Viamonte, M.; Garcia, H. Imaging findings in human immunodeficiency virus–related pulmonary hypertension: Report of five cases and review of the literature. Radiology 2002, 223, 820–827. [Google Scholar] [CrossRef]
- Parikh, R.V.; Ma, Y.; Scherzer, R.; Heringer, A.S.; Macgregor, J.S.; Martin, J.N.; Deeks, S.G.; Ganz, P.; Hsue, P.Y. Endothelin-1 predicts hemodynamically assessed pulmonary arterial hypertension in HIV Infection. PLoS ONE 2016, 11, e0146355. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.B.; Khan, M.; Garcia-Barrio, M.; Powell, M.; Bond, V.C. Apoptotic effects in primary human umbilical vein endothelial cell cultures caused by exposure to virion-associated and cell membrane-associated HIV-1 gp120. J. Acquir. Immune Defic. Syndr. 2001, 27, 213–221. [Google Scholar] [CrossRef]
- Ullrich, C.K.; Groopman, J.E.; Ganju, R.K. HIV-1 gp120- and gp160-induced apoptosis in cultured endothelial cells is mediated by caspases. Blood 2000, 96, 1438–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.B.; Bond, V.C. Involvement of protein kinase C in HIV-1 gp120-induced apoptosis in primary endothelium. J. Acquir. Immune Defic. Syndr. 2000, 25, 375–389. [Google Scholar] [CrossRef]
- Khan, N.A.; Di Cello, F.; Stins, M.; Kim, K.S. Gp120-mediated cytotoxicity of human brain microvascular endothelial cells is dependent on p38 mitogen-activated protein kinase activation. J. Neurovirol. 2007, 13, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Price, T.O.; Uras, F.; Banks, W.A.; Ercal, N. A novel antioxidant N-acetylcysteine amide prevents gp120- and Tat-induced oxidative stress in brain endothelial cells. Exp. Neurol. 2006, 201, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kanmogne, G.D.; Primeaux, C.; Grammas, P. HIV-1 gp120 proteins alter tight junction protein expression and brain endothelial cell permeability: Implications for the pathogenesis of HIV-associated dementia. J. Neuropathol. Exp. Neurol. 2005, 64, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Akhter, S.; Chaudhuri, A.; Kanmogne, G.D. HIV-1 gp120 induces cytokine expression, leukocyte adhesion, and transmigration across the blood-brain barrier: Modulatory effects of STAT1 signaling. Microvasc. Res. 2009, 77, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Kanmogne, G.D.; Primeaux, C.; Grammas, P. Induction of apoptosis and endothelin-1 secretion in primary human lung endothelial cells by HIV-1 gp120 proteins. Biochem. Biophys. Res. Commun. 2005, 333, 1107–1115. [Google Scholar] [CrossRef]
- Suresh, S.J.; Suzuki, Y.J. SARS-CoV-2 spike protein and lung vascular cells. J. Respir. 2021, 1, 40–48. [Google Scholar] [CrossRef]
- Amsellem, V.; Lipskaia, L.; Abid, S.; Poupel, L.; Houssaini, A.; Quarck, R.; Marcos, E.; Mouraret, N.; Parpaleix, A.; Bobe, R.; et al. CCR5 as a treatment target in pulmonary arterial hypertension. Circulation 2014, 130, 880–891. [Google Scholar] [CrossRef]
- Schecter, A.D.; Berman, A.B.; Yi, L.; Mosoian, A.; McManus, C.M.; Berman, J.W.; Klotman, M.E.; Taubman, M.B. HIV envelope gp120 activates human arterial smooth muscle cells. Proc. Natl. Acad. Sci. USA 2001, 98, 10142–10147. [Google Scholar] [CrossRef] [Green Version]
- András, I.E.; Pu, H.; Deli, M.A.; Nath, A.; Hennig, B.; Toborek, M. HIV-1 Tat protein alters tight junction protein expression and distribution in cultured brain endothelial cells. J. Neurosci. Res. 2003, 74, 255–265. [Google Scholar] [CrossRef]
- Albini, A.; Soldi, R.; Giunciuclio, D.; Giraudo, E.; Benelli, R.; Primo, L.; Noonan, D.; Salio, M.; Camussi, G.; Rockl, W.; et al. The angiogenesis induced by HIV–1 tat protein is mediated by the Flk–1/KDR receptor on vascular endothelial cells. Nat. Med. 1996, 2, 1371–1375. [Google Scholar] [CrossRef]
- Park, I.W.; Ullrich, C.K.; Schoenberger, E.; Ganju, R.K.; Groopman, J.E. HIV-1 Tat induces microvascular endothelial apoptosis through caspase activation. J. Immunol. 2001, 167, 2766–2771. [Google Scholar] [CrossRef]
- Matzen, K.; Dirkx, A.E.; oude Egbrink, M.G.; Speth, C.; Götte, M.; Ascherl, G.; Grimm, T.; Griffioen, A.W.; Stürzl, M. HIV-1 Tat increases the adhesion of monocytes and T-cells to the endothelium in vitro and in vivo: Implications for AIDS-associated vasculopathy. Virus Res. 2004, 104, 145–155. [Google Scholar] [CrossRef]
- Wang, T.; Green, L.A.; Gupta, S.K.; Kim, C.; Wang, L.; Almodovar, S.; Flores, S.C.; Prudovsky, I.A.; Jolicoeur, P.; Liu, Z.; et al. Transfer of intracellular HIV Nef to endothelium causes endothelial dysfunction. PLoS ONE 2014, 9, e91063. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Yao, S.; Hu, M.; Li, W.; Hao, T.; Zhou, F.; Zhu, X.; Lu, H.; Qin, D.; Yan, Q.; et al. HIV-1 Nef and KSHV oncogene K1 synergistically promote angiogenesis by inducing cellular miR-718 to regulate the PTEN/AKT/mTOR signaling pathway. Nucleic Acids Res. 2014, 42, 9862–9879. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.J.; Gychka, S.G. SARS-CoV-2 spike protein elicits cell signaling in human host cells: Implications for possible consequences of COVID-19 vaccines. Vaccines 2021, 9, 36. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seeherman, S.; Suzuki, Y.J. Viral Infection and Cardiovascular Disease: Implications for the Molecular Basis of COVID-19 Pathogenesis. Int. J. Mol. Sci. 2021, 22, 1659. https://doi.org/10.3390/ijms22041659
Seeherman S, Suzuki YJ. Viral Infection and Cardiovascular Disease: Implications for the Molecular Basis of COVID-19 Pathogenesis. International Journal of Molecular Sciences. 2021; 22(4):1659. https://doi.org/10.3390/ijms22041659
Chicago/Turabian StyleSeeherman, Sarah, and Yuichiro J. Suzuki. 2021. "Viral Infection and Cardiovascular Disease: Implications for the Molecular Basis of COVID-19 Pathogenesis" International Journal of Molecular Sciences 22, no. 4: 1659. https://doi.org/10.3390/ijms22041659
APA StyleSeeherman, S., & Suzuki, Y. J. (2021). Viral Infection and Cardiovascular Disease: Implications for the Molecular Basis of COVID-19 Pathogenesis. International Journal of Molecular Sciences, 22(4), 1659. https://doi.org/10.3390/ijms22041659