
Oxidative Stress and Low-Grade Inflammation in Polycystic Ovary Syndrome: Controversies and New Insights


 ,
, 

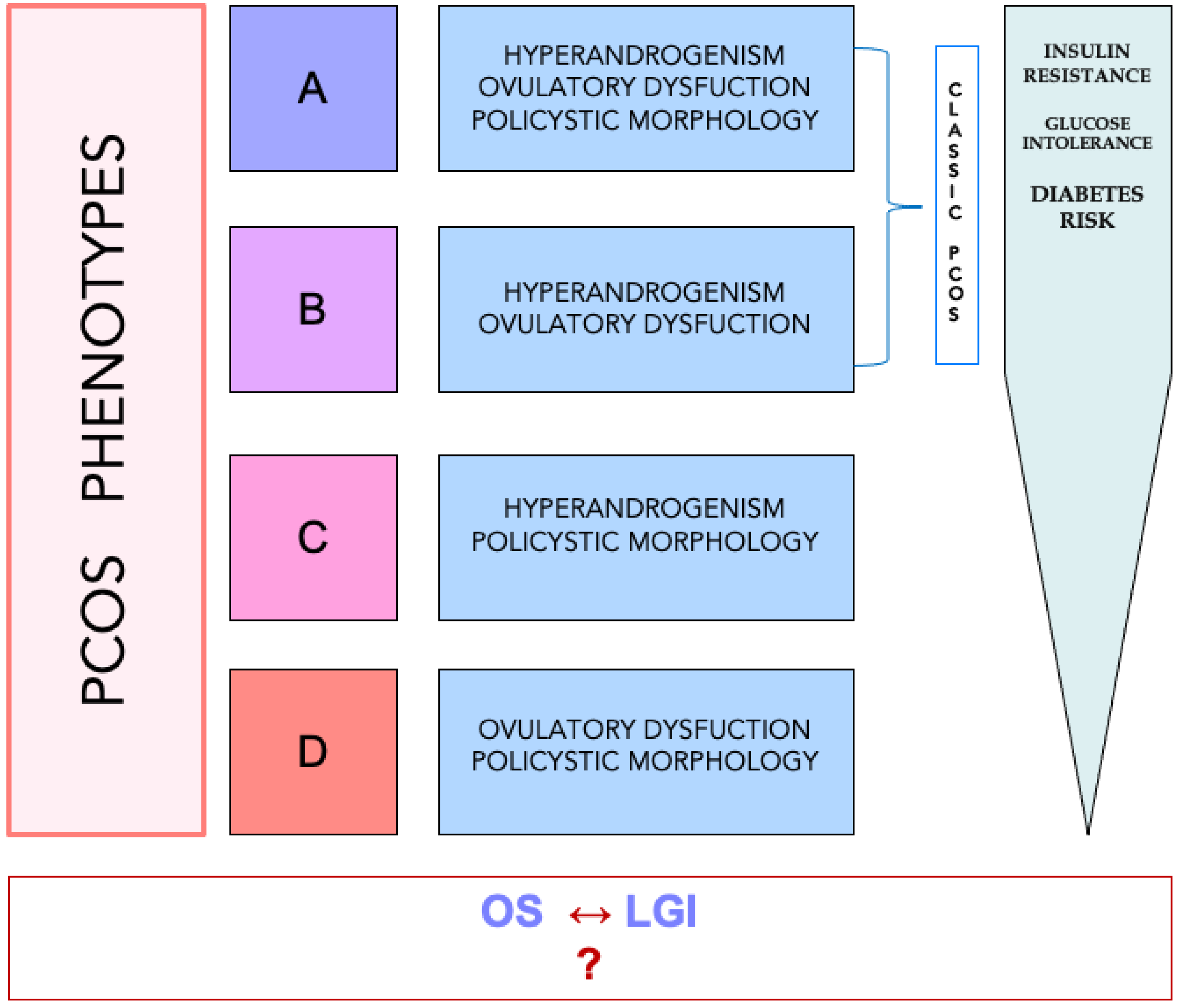{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mechanism of Oxidative Stress in PCOS
3. Role of Obesity: Pros
4. Role of Obesity: Cons
5. The Underestimated Role of GH/IGF-1 Axis
6. Other Implications Due to Oxidative Stress
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| CRP | C-reactive protein |
| DHT | Dihydrotestosterone |
| FLCs | Free Light Chains of immunoglobulins |
| GH | Growth Hormone |
| HY | Hyperandrogenism |
| IGF-1 | Insulin-like growth factor 1 |
| HA IR | Hyperandrogenism Insulin Resistance |
| LCN2 | Lipocalin-2 |
| LGI | Low-grade inflammation |
| MCP-1 | Monocytes chemoattractant protein 1 |
| MNC | Mononuclear cell |
| NIH | National Insitute of Health |
| NLR | Neutrophils/Linfocytes ratio |
| Nrf2 | Nuclear factor erythroid 2–related factor 2 |
| NW-PCOS | Normal weight PCOS |
| OB-POCS | Obese PCOS |
| OC | Osteocalcin |
| OD | Ovulatory Disfunction |
| OS | Oxidative Stress |
| PCOM | Polycystic Ovary Morphology |
| PCOS | Polycystic Ovary Syndrome |
| ROS | Reactive Oxygen Species |
| T | Testosterone |
| TNF | Tumor Necrosis Factor |
| u-OC | Undercarboxylated Osteocalcin |
References
- Murri, M.; Luque-ramírez, M.; Insenser, M.; Ojeda-ojeda, M.; Escobar-morreale, H.F. Circulating markers of oxidative stress and polycystic ovary syndrome (pcos): A systematic review and meta-analysis. Hum. Reprod. Update 2013, 19, 268–288. [Google Scholar] [CrossRef] [PubMed]
- Fauser, B.C.J.M. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar]
- Ladrón de Guevara, A.; Fux-Otta, C.; Crisosto, N.; Szafryk de Mereshian, P.; Echiburú, B.; Iraci, G.; Perez-Bravo, F.; Sir-Petermann, T. Metabolic profile of the different phenotypes of polycystic ovary syndrome in two Latin American populations. Fertil. Steril. 2014, 101, 1732–1739.e92. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The Androgen Excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. Position statement: Criteria for defining polycystic ovary syndrome as a predominantly hyperandrogenic syndrome: An androgen excess society guideline. J. Clin. Endocrinol. Metab. 2006, 91, 4237–4245. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.R.B.; Kaplan, L.K.; Ouyang, P.; Rizza, R.A. Evidence-Based Methodology Workshop on Polycystic Ovary Syndrome; National Institues of Health: Rockville Pike, MD, USA, 2012. [Google Scholar]
- Lizneva, D.; Suturina, L.; Walker, W.; Brakta, S.; Gavrilova-Jordan, L.; Azziz, R. Criteria, prevalence, and phenotypes of polycystic ovary syndrome. Fertil. Steril. 2016, 106, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Azziz, R. Introduction: Determinants of polycystic ovary syndrome. Fertil. Steril. 2016, 106, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Diamanti-Kandarakis, E.; Panidis, D. Unravelling the phenotypic map of polycystic ovary syndrome (PCOS): A prospective study of 634 women with PCOS. Clin. Endocrinol. 2007, 67, 735–742. [Google Scholar] [CrossRef]
- Goverde, A.J.; Van Koert, A.J.B.; Eijkemans, M.J.; Knauff, E.A.H.; Westerveld, H.E.; Fauser, B.C.J.M.; Broekmans, F.J. Indicators for metabolic disturbances in anovulatory women with polycystic ovary syndrome diagnosed according to the Rotterdam consensus criteria. Hum. Reprod. 2009, 24, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Mehrabian, F.; Khani, B.; Kelishadi, R.; Kermani, N. The prevalence of metabolic syndrome and insulin resistance according to the phenotypic subgroups of polycystic ovary syndrome in a representative sample of Iranian females. J. Res. Med. Sci. 2011, 16, 763. [Google Scholar] [PubMed]
- Welt, C.K.; Gudmundsson, J.A.; Arason, G.; Adams, J.; Palsdottir, H.; Gudlaugsdottir, G.; Ingadottir, G.; Crowley, W.F. Characterizing discrete subsets of polycystic ovary syndrome as defined by the Rotterdam criteria: The impact of weight on phenotype and metabolic features. J. Clin. Endocrinol. Metab. 2006, 91, 4842–4848. [Google Scholar] [CrossRef] [Green Version]
- Shorakae, S.; Teede, H.; de Courten, B.; Lambert, G.; Boyle, J.; Moran, L. The Emerging Role of Chronic Low-Grade Inflammation in the Pathophysiology of Polycystic Ovary Syndrome. Semin. Reprod. Med. 2015, 33, 257–269. [Google Scholar] [CrossRef]
- Jones, H.; Sprung, V.S.; Pugh, C.J.A.; Daousi, C.; Irwin, A.; Aziz, N.; Adams, V.L.; Thomas, E.L.; Bell, J.D.; Kemp, G.J.; et al. Polycystic ovary syndrome with hyperandrogenism is characterized by an increased risk of hepatic steatosis compared to nonhyperandrogenic PCOS phenotypes and healthy controls, independent of obesity and insulin resistance. J. Clin. Endocrinol. Metab. 2012, 97, 3709–3716. [Google Scholar] [CrossRef] [PubMed]
- Jamil, A.S.; Alalaf, S.K.; Al-Tawil, N.G.; Al-Shawaf, T. A case-control observational study of insulin resistance and metabolic syndrome among the four phenotypes of polycystic ovary syndrome based on Rotterdam criteria Female Fertility. Reprod. Health 2015, 12, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Hwang, K.R.; Choi, Y.M.; Moon, S.Y.; Chae, S.J.; Park, C.W.; Kim, H.O.; Choi, D.S.; Kwon, H.C.; Kang, B.M.; et al. Complete phenotypic and metabolic profiles of a large consecutive cohort of untreated Korean women with polycystic ovary syndrome. Fertil. Steril. 2014, 101, 1424–1430. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Eustress and distress in redox homeostasis. In Stress: Physiology, Biochemistry, and Pathology Handbook of Stress Series; Academic Press: Cambridge, MA, USA, 2019; pp. 153–163. [Google Scholar]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Giacchi, E.; Raimondo, S.; Di, C.; Silvestrini, A.; Meucci, E. Hypothyroidism, Oxidative Stress and Reproduction. In Hypothyroidism Influences and Treatments; InTech: Shangai, China, 2012. [Google Scholar]
- Zhang, R.; Liu, H.; Bai, H.; Zhang, Y.; Liu, Q.; Guan, L.; Fan, P. Oxidative stress status in Chinese women with different clinical phenotypes of polycystic ovary syndrome. Clin. Endocrinol. 2017, 86, 88–96. [Google Scholar] [CrossRef]
- Abella, V.; Scotece, M.; Conde, J.; Gómez, R.; Lois, A.; Pino, J.; Gómez-Reino, J.J.; Lago, F.; Mobasheri, A.; Gualillo, O. The potential of lipocalin-2/NGAL as biomarker for inflammatory and metabolic diseases. Biomarkers 2015, 20, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Moschen, A.R.; Gerner, R.R.; Wang, J.; Klepsch, V.; Adolph, T.E.; Reider, S.J.; Hackl, H.; Pfister, A.; Schilling, J.; Moser, P.L.; et al. Lipocalin 2 Protects from Inflammation and Tumorigenesis Associated with Gut Microbiota Alterations. Cell Host Microbe 2016, 19, 455–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolie syndrome and altered gut microbiota in mice lacking toll-like receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef] [Green Version]
- van Greevenbroek, M.M.J.; Schalkwijk, C.G.; Stehouwer, C.D.A. Obesity-associated low-grade inflammation in type 2 diabetes mellitus: Causes and consequences. Neth. J. Med. 2013, 71, 174–187. [Google Scholar]
- Margioris, A.N.; Dermitzaki, E.; Venihaki, M.; Tsatsanis, C. Chronic low-grade inflammation. In Diet, Immunity and Inflammation; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780857090379. [Google Scholar]
- Masharani, U.; Gjerde, C.; Evans, J.L.; Youngren, J.F.; Goldfine, I.D. Effects of controlled-release alpha lipoic acid in lean, nondiabeticpatients with polycystic ovary syndrome. J. Diabetes Sci. Technol. 2010, 4, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Rzepczynska, I.J.; Foyouzi, N.; Piotrowski, P.C.; Celik-Ozenci, C.; Cress, A.; Duleba, A.J. Antioxidants induce apoptosis of rat ovarian theca-interstitial cells. Biol. Reprod. 2011, 84, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Moro, F.; Morciano, A.; Tropea, A.; Sagnella, F.; Palla, C.; Scarinci, E.; Cosentino, N.; Niccoli, G.; Liuzzo, G.; Crea, F.; et al. CD4+CD28null T lymphocyte frequency, a new marker of cardiovascular risk: Relationship with polycystic ovary syndrome phenotypes. Fertil. Steril. 2012, 98, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- González, F. Inflammation in Polycystic Ovary Syndrome: Underpinning of insulin resistance and ovarian dysfunction. Steroids 2012, 77, 300–305. [Google Scholar] [CrossRef] [Green Version]
- Ehrmann, D.A. Polycystic Ovary Syndrome. N. Engl. J. Med. 2005, 352, 1223–1236. [Google Scholar] [CrossRef]
- Kirchengast, S.; Huber, J. Body composition characteristics and body fat distribution in lean women with polycystic ovary syndrome. Hum. Reprod. 2001, 16, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Ganie, M.; Vasudevan, V.; Wani, I.; Baba, M.; Arif, T.; Rashid, A. Epidemiology, pathogenesis, genetics & management of polycystic ovary syndrome in India. Indian J. Med. Res. 2019, 150, 333. [Google Scholar] [PubMed]
- Zhang, J.; Bao, Y.; Zhou, X.; Zheng, L. Polycystic ovary syndrome and mitochondrial dysfunction. Reprod. Biol. Endocrinol. 2019, 17, 1–15. [Google Scholar] [CrossRef]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Chung, D.J.; Lee, H.S.; Kim, T.J.; Kim, M.H.; Jeong, H.J.; Im, J.A.; Lee, D.C.; Lee, J.W. Mitochondrial DNA copy number in peripheral blood in polycystic ovary syndrome. Metabolism 2011, 60, 1677–1682. [Google Scholar] [CrossRef]
- Lee, H.K.; Song, J.H.; Shin, C.S.; Park, D.J.; Park, K.S.; Lee, K.U.; Koh, C.S. Decreased mitochondrial DNA content in peripheral blood precedes the development of non-insulin-dependent diabetes mellitus. Diabetes Res. Clin. Pract. 1998, 42, 161–167. [Google Scholar] [CrossRef]
- Zhuo, G.; Ding, Y.; Feng, G.; Yu, L.; Jiang, Y. Analysis of mitochondrial DNA sequence variants in patients with polycystic ovary syndrome. Arch. Gynecol. Obs. 2012, 286, 653–659. [Google Scholar] [CrossRef]
- Wilson, F.H.; Hariri, A.; Farhi, A.; Zhao, H.; Petersen, K.F.; Toka, H.R.; Nelson-Williams, C.; Raja, K.M.; Kashgarian, M.; Shulman, G.I.; et al. A cluster of metabolic defects caused by mutation in a mitochondrial tRNA. Science 2004, 306, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Reddy, T.V.; Govatati, S.; Deenadayal, M.; Sisinthy, S.; Bhanoori, M. Impact of mitochondrial DNA copy number and displacement loop alterations on polycystic ovary syndrome risk in south Indian women. Mitochondrion 2019, 44, 35–40. [Google Scholar] [CrossRef]
- Liu, X.; Trakooljul, N.; Hadlich, F.; Murani, E.; Wimmers, K.; Ponsuksili, S. Mitochondrial-nuclear crosstalk, haplotype and copy number variation distinct in muscle fiber type, mitochondrial respiratory and metabolic enzyme activities. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Reddy, T.V.; Govatati, S.; Deenadayal, M.; Shivaji, S.; Bhanoori, M. Polymorphisms in the TFAM and PGC1-α genes and their association with polycystic ovary syndrome among South Indian women. Gene 2018, 641, 129–136. [Google Scholar] [CrossRef]
- González, F.; Rote, N.S.; Minium, J.; Kirwan, J.P. Increased activation of nuclear factor κB triggers inflammation and insulin resistance in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- González, F.; Rote, N.S.; Minium, J.; Kirwan, J.P. Reactive oxygen species-induced oxidative stress in the development of insulin resistance and hyperandrogenism in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 336–340. [Google Scholar] [CrossRef] [Green Version]
- González, F.; Minium, J.; Rote, N.S.; Kirwan, J.P. Hyperglycemia alters tumor necrosis factor-α release from mononuclear cells in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2005, 90, 5336–5342. [Google Scholar] [CrossRef] [Green Version]
- González, F.; Rote, N.S.; Minium, J.; Kirwan, J.P. In vitro evidence that hyperglycemia stimulates tumor necrosis factor-α release in obese women with polycystic ovary syndrome. J. Endocrinol. 2006, 188, 521–529. [Google Scholar] [CrossRef]
- Liu, S.; Navarro, G.; Mauvais-Jarvis, F. Androgen excess produces systemic oxidative stress and predisposes to β-cell failure in female mice. PLoS ONE 2010, 5, e11302. [Google Scholar] [CrossRef]
- Siemienowicz, K.J.; Coukan, F.; Franks, S.; Rae, M.T.; Duncan, W.C. Aberrant subcutaneous adipogenesis precedes adult metabolic dysfunction in an ovine model of polycystic ovary syndrome (PCOS). Mol. Cell. Endocrinol. 2021, 519, 111042. [Google Scholar] [CrossRef] [PubMed]
- de Zegher, F.; Lopez-Bermejo, A.; Ibáñez, L. Adipose tissue expandability and the early origins of PCOS. Trends Endocrinol. Metab. 2009, 20, 418–423. [Google Scholar] [CrossRef] [PubMed]
- de Zegher, F.; López-Bermejo, A.; Ibáñez, L. Central Obesity, Faster Maturation, and ‘PCOS’ in Girls. Trends Endocrinol. Metab. 2018, 29, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Stener-Victorin, E.; Padmanabhan, V.; Walters, K.A.; Campbell, R.E.; Benrick, A.; Giacobini, P.; Dumesic, D.A.; Abbott, D.H. Animal Models to Understand the Etiology and Pathophysiology of Polycystic Ovary Syndrome. Endocr. Rev. 2020, 41, 538–576. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, T.A.; Kajimoto, Y.; Watada, H.; Kaneto, H.; Kishimoto, M.; Umayahara, Y.; Fujitani, Y.; Kamada, T.; Kawamori, R.; Yamasaki, Y. Glycation-dependent, reactive oxygen species-mediated suppression of the insulin gene promoter activity in HIT cells. J. Clin. Investig. 1997, 99, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Rudich, A.; Tlrosh, A.; Potashnik, R.; Hemi, R.; Kanety, H.; Bashan, N. Prolonged oxidative stress impairs insulin-induced GLUT4 translocation in 3T3-L1 adipocytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef]
- Takeda, E.; Arai, H.; Yamamoto, H.; Okumura, H.; Taketani, Y. Control of oxidative stress and metabolic homeostasis by the suppression of postprandial hyperglycemia. J. Med. Investig. 2005, 52, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, V.; Uchida, T.; Yenush, L.; Davis, R.; White, M.F. The c-Jun NH2-terminal kinase promotes insulin resistance during association with insulin receptor substrate-1 and phosphorylation of Ser307. J. Biol. Chem. 2000, 275, 9047–9054. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.A.; Satoh, H.; Favelyukis, S.; Babendure, J.L.; Imamura, T.; Sbodio, J.I.; Zalevsky, J.; Dahiyat, B.I.; Chi, N.W.; Olefsky, J.M. JNK and tumor necrosis factor-α mediate free fatty acid-induced insulin resistance in 3T3-L1 adipocytes. J. Biol. Chem. 2005, 280, 35361–35371. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Nong, S.; Huang, X.; Lu, Y.; Zhao, H.; Lin, Y.; Man, Y.; Wang, S.; Yang, J.; Li, J. The effects of palmitate on hepatic insulin resistance are mediated by NADPH oxidase 3-derived reactive oxygen species through JNK and p38 MAPK pathways. J. Biol. Chem. 2010, 285, 29965–29973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuncman, G.; Hirosumi, J.; Solinas, G.; Chang, L.; Karin, M.; Hotamisligil, G.S. Functional in vivo interactions between JNK1 and JNK2 isoforms in obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2006, 103, 10741–10746. [Google Scholar] [CrossRef] [Green Version]
- Masharani, U.B.; Maddux, B.A.; Li, X.; Sakkas, G.K.; Mulligan, K.; Schambelan, M.; Goldfine, I.D.; Youngren, J.F. Insulin resistance in non-obese subjects is associated with activation of the JNK pathway and impaired insulin signaling in skeletal muscle. PLoS ONE 2011, 6, e19878. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Martorana, G.E.; Magini, M.; Festa, R.; Raimondo, S.; Silvestrini, A.; Nicolotti, N.; Mordente, A.; Mele, M.C.; Miggiano, G.A.D.; et al. Oxidative stress and metabolic syndrome: Effects of a natural antioxidants enriched diet on insulin resistance. Clin. Nutr. Espen 2015, 10, e52–e60. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Horvath, T.L.; Andrews, Z.B.; Diano, S. Fuel utilization by hypothalamic neurons: Roles for ROS. Trends Endocrinol. Metab. 2009, 20, 78–87. [Google Scholar] [CrossRef]
- Best, C.L.; Pudney, J.; Welch, W.R.; Burger, N.; Hill, J.A. Localization and characterization of white blood cell populations within the human ovary throughout the menstrual cycle and menopause. Hum. Reprod. 1996, 11, 790–797. [Google Scholar] [CrossRef] [Green Version]
- Ortega, I.; Villanueva, J.A.; Wong, D.H.; Cress, A.B.; Sokalska, A.; Stanley, S.D.; Duleba, A.J. Resveratrol potentiates effects of simvastatin on inhibition of rat ovarian theca-interstitial cells steroidogenesis. J. Ovarian Res. 2014, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Festa, R.; Di Donna, V.; Leone, E.; Littarru, G.P.; Silvestrini, A.; Meucci, E.; Pontecorvi, A. Hormones and antioxidant systems: Role of pituitary and pituitary-dependent axes. J. Endocrinol. Investig. 2010, 33, 422–433. [Google Scholar] [CrossRef]
- Tsai-Turton, M.; Luong, B.T.; Tan, Y.; Luderer, U. Cyclophosphamide-induced apoptosis in COV434 human granulosa cells involves oxidative stress and glutathione depletion. Toxicol. Sci. 2007, 98, 216–230. [Google Scholar] [CrossRef] [Green Version]
- Hannon, E.; Knox, O.; Sugden, K.; Burrage, J.; Wong, C.C.Y.; Belsky, D.W.; Corcoran, D.L.; Arseneault, L.; Moffitt, T.E.; Caspi, A.; et al. Characterizing genetic and environmental influences on variable DNA methylation using monozygotic and dizygotic twins. PLoS Genet. 2018, 14, e1007544. [Google Scholar] [CrossRef]
- Özer, A.; Bakacak, M.; Kiran, H.; Ercan, Ö.; Köstü, B.; Kanat-Pektaş, M.; Kilinç, M.; Aslan, F. Increased oxidative stress is associated with insulin resistance and infertility in polycystic ovary syndrome. Ginekol. Pol. 2016, 87, 733–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharm. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Škrha, J.; Šindelka, G.; Kvasnička, J.; Hilgertová, J. Insulin action and fibrinolysis influenced by vitamin E in obese Type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 1999, 44, 27–33. [Google Scholar] [CrossRef]
- Dandona, P.; Mohanty, P.; Ghanim, H.; Aljada, A.; Browne, R.; Hamouda, W.; Prabhala, A.; Afzal, A.; Garg, R. The suppressive effect of dietary restriction and weight loss in the obese on the generation of reactive oxygen species by leukocytes, lipid peroxidation, and protein carbonylation. J. Clin. Endocrinol. Metab. 2001, 86, 355–362. [Google Scholar]
- Block, G.; Dietrich, M.; Norkus, E.P.; Morrow, J.D.; Hudes, M.; Caan, B.; Packer, L. Factors associated with oxidative stress in human populations. Am. J. Epidemiol. 2002, 156, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Konukoǧlu, D.; Serin, Ö.; Ercan, M.; Turhan, M.S. Plasma homocysteine levels in obese and non-obese subjects with or without hypertension; its relationship with oxidative stress and copper. Clin. Biochem. 2003, 36, 405–408. [Google Scholar] [CrossRef]
- Festa, A.; D’Agostino, R.; Williams, K.; Karter, A.J.; Mayer-Davis, E.J.; Tracy, R.P.; Haffner, S.M. The relation of body fat mass and distribution to markers of chronic inflammation. Int. J. Obes. 2001, 25, 1407–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancello, R.; Tordjman, J.; Poitou, C.; Guilhem, G.; Bouillot, J.L.; Hugol, D.; Coussieu, C.; Basdevant, A.; Hen, A.B.; Bedossa, P.; et al. Increased infiltration of macrophages in omental adipose tissue is associated with marked hepatic lesions in morbid human obesity. Diabetes 2006, 55, 1554–1561. [Google Scholar] [CrossRef] [Green Version]
- ClÉment, K.; Viguerie, N.; Poitou, C.; Carette, C.; Pelloux, V.; Curat, C.A.; Sicard, A.; Rome, S.; Benis, A.; Zucker, J.; et al. Weight loss regulates inflammation-related genes in white adipose tissue of obese subjects. FASEB J. 2004, 18, 1657–1669. [Google Scholar]
- Anna Diehl, M.A.E. Nonalcoholic steatosis and steatohepatitis IV. Nonalcoholic fatty liver disease abnormalities in macrophage function and cytokines. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G1–G5. [Google Scholar] [CrossRef]
- Arkan, M.C.; Hevener, A.L.; Greten, F.R.; Maeda, S.; Li, Z.W.; Long, J.M.; Wynshaw-Boris, A.; Poli, G.; Olefsky, J.; Karin, M. IKK-β links inflammation to obesity-induced insulin resistance. Nat. Med. 2005, 11, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Schünbeck, U.; Mach, F.; Sukhova, G.K.; Herman, M.; Graber, P.; Kehry, M.R.; Libby, P. CD40 ligation induces tissue factor expression in human vascular smooth muscle cells. Am. J. Pathol. 2000, 156, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Reardon, C.A.; Getz, G.S. Mouse models of atherosclerosis. Curr. Opin. Lipidol. 2001, 12, 167–173. [Google Scholar] [CrossRef]
- Michelsen, K.S.; Wong, M.H.; Shah, P.K.; Zhang, W.; Yano, J.; Doherty, T.M.; Akira, S.; Rajavashisth, T.B.; Arditi, M. Lack of toll-like receptor 4 or myeloid differentiation factor 88 reduces atherosclerosis and alters plaque phenotype in mice deficient in apolipoprotein E. Proc. Natl. Acad. Sci. USA 2004, 101, 10679–10684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Luck, H.; Tsai, S.; Chung, J.; Clemente-Casares, X.; Ghazarian, M.; Revelo, X.S.; Lei, H.; Luk, C.T.; Shi, S.Y.; Surendra, A.; et al. Regulation of obesity-related insulin resistance with gut anti-inflammatory agents. Cell Metab. 2015, 21, 527–542. [Google Scholar] [CrossRef] [Green Version]
- Garidou, L.; Pomié, C.; Klopp, P.; Waget, A.; Charpentier, J.; Aloulou, M.; Giry, A.; Serino, M.; Stenman, L.; Lahtinen, S.; et al. The Gut Microbiota Regulates Intestinal CD4 T Cells Expressing RORγt and Controls Metabolic Disease. Cell Metab. 2015, 22, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Silva, C.; Rotellar, F.; Gil, M.J.; Cienfuegos, J.A.; Salvador, J.; Frühbeck, G. Increased adipose tissue expression of lipocalin-2 in obesity is related to inflammation and matrix metalloproteinase-2 and metalloproteinase-9 activities in humans. J. Mol. Med. 2009, 87, 803–813. [Google Scholar] [CrossRef]
- Auguet, T.; Quintero, Y.; Terra, X.; Martínez, S.; Lucas, A.; Pellitero, S.; Aguilar, C.; Hernández, M.; del Castillo, D.; Richart, C. Upregulation of lipocalin 2 in adipose tissues of severely obese women: Positive relationship with proinflammatory cytokines. Obesity 2011, 19, 2295–2300. [Google Scholar] [CrossRef]
- Yan, Q.W.; Yang, Q.; Mody, N.; Graham, T.E.; Hsu, C.H.; Xu, Z.; Houstis, N.E.; Kahn, B.B.; Rosen, E.D. The adipokine lipocalin 2 is regulated by obesity and promotes insulin resistance. Diabetes 2007, 56, 2533–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.Y.; Jo, Y.H.; Kim, M.J. The level of nitric oxide regulates lipocalin-2 expression under inflammatory condition in RINm5F beta-cells. Atherosclerosis 2016, 252, e181. [Google Scholar] [CrossRef]
- Law, I.K.M.; Xu, A.; Lam, K.S.L.; Berger, T.; Mak, T.W.; Vanhoutte, P.M.; Liu, J.T.C.; Sweeney, G.; Zhou, M.; Yang, B.; et al. Lipocalin-2 deficiency attenuates insulin resistance associated with aging and obesity. Diabetes 2010, 59, 872–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Jin, D.; Zhang, Y.; Wright, W.; Bazuine, M.; Brockman, D.A.; Bernlohr, D.A.; Chen, X. Lipocalin-2 deficiency impairs thermogenesis and potentiates diet-induced insulin resistance in mice. Diabetes 2010, 59, 1376–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, L.S.; Parker Siddall, C.; Rosen, E.D. A minor role for lipocalin 2 in high-fat diet-induced glucose intolerance. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E825–E835. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, Q.; Lv, C.; Qin, N.; Lei, S.; Yuan, Q.; Wang, G. The changes of serum sKlotho and NGAL levels and their correlation in type 2 diabetes mellitus patients with different stages of urinary albumin. Diabetes Res. Clin. Pract. 2014, 106, 343–350. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Manco, M.; Ibáñez, J.; García-Fuentes, E.; Ortega, F.; Gorostiaga, E.; Vendrell, J.; Izquierdo, M.; Martínez, C.; Nolfe, G.; et al. Metabolic endotoxemia and saturated fat contribute to circulating NGAL concentrations in subjects with insulin resistance. Int. J. Obes. 2010, 34, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hamnvik, O.-P.R.; Petrou, M.; Gong, H.; Chamberland, J.P.; Christophi, C.A.; Kales, S.N.; Christiani, D.C.; Mantzoros, C.S. Circulating lipocalin 2 is associated with body fat distribution at baseline but is not an independent predictor of insulin resistance: The prospective Cyprus Metabolism Study. Eur. J. Endocrinol. 2011, 165, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Currò, D.; Vergani, E.; Bruno, C.; Comi, S.; D’Abate, C.; Mancini, A. Plasmatic lipocalin-2 levels in chronic low-grade inflammation syndromes: Comparison between metabolic syndrome, total and partial adult growth hormone deficiency. BioFactors 2020, 46, 629–636. [Google Scholar] [CrossRef]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Obesity 2008, 16, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Schaeffler, A.; Gross, P.; Buettner, R.; Bollheimer, C.; Buechler, C.; Neumeier, M.; Kopp, A.; Schoelmerich, J.; Falk, W. Fatty acid-induced induction of Toll-like receptor-4/nuclear factor-κB pathway in adipocytes links nutritional signalling with innate immunity. Immunology 2009, 126, 233–245. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Villuendas, G.; Botella-Carretero, J.I.; Sancho, J.; San Millán, J.L. Obesity, and not insulin resistance, is the major determinant of serum inflammatory cardiovascular risk markers in pre-menopausal women. Diabetologia 2003, 46, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möhlig, M.; Spranger, J.; Osterhoff, M.; Ristow, M.; Pfeiffer, A.F.H.; Schill, T.; Schlösser, H.W.; Brabant, G.; Schöfl, C. The polycystic ovary syndrome per se is not associated with increased chronic inflammation. Eur. J. Endocrinol. 2004, 150, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puder, J.J.; Varga, S.; Kraenzlin, M.; De Geyter, C.; Keller, U.; Müller, B. Central fat excess in polycystic ovary syndrome: Relation to low-grade inflammation and insulin resistance. J. Clin. Endocrinol. Metab. 2005, 90, 6014–6021. [Google Scholar] [CrossRef] [Green Version]
- Stephens, J.M.; Butts, M.D.; Pekala, P.H. Regulation of transcription factor mRNA accumulation during 3T3-L1 preadipocyte differentiation by tumour necrosis factor-α. J. Mol. Endocrinol. 1992, 9, 61–72. [Google Scholar] [CrossRef]
- Kelly, C.J.G.; Lyall, H.; Petrie, J.R.; Gould, G.W.; Connell, J.M.C.; Rumley, A.; Lowe, G.D.O.; Sattar, N. A specific elevation in tissue plasminogen activator antigen in women with polycystic ovarian syndrome. J. Clin. Endocrinol. Metab. 2002, 87, 3287–3290. [Google Scholar] [CrossRef]
- Morris, R.S.; Gentschein, E.; Wong, I.L.; Paulson, R.J.; Hatch, I.E.; Lobo, R.A. Prorenin is elevated in polycystic ovary syndrome and may reflect hyperandrogenism. Fertil. Steril. 1995, 64, 1099–1103. [Google Scholar] [CrossRef]
- Jaatinen, T.A.; Matinlauri, I.; Anttila, L.; Koskinen, P.; Erkkola, R.; Irjala, K. Serum total renin is elevated in women with polycystic ovarian syndrome. Fertil. Steril. 1995, 63, 1000–1004. [Google Scholar] [CrossRef]
- Panidis, D.; Kourtis, A.; Farmakiotis, D.; Mouselech, T.; Rousso, D.; Koliakos, G. Serum adiponectin levels in women with polycystic ovary syndrome. Hum. Reprod. 2003, 18, 1790–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murri, M.; García-Fuentes, E.; García-Almeida, J.M.; Garrido-Sánchez, L.; Mayas, M.D.; Bernal, R.; Tinahones, F.J. Changes in oxidative stress and insulin resistance in morbidly obese patients after bariatric surgery. Obes. Surg. 2010, 20, 363–368. [Google Scholar] [CrossRef]
- Kisakol, G.; Guney, E.; Bayraktar, F.; Yilmaz, C.; Kabalak, T.; Özmen, D. Effect of surgical weight loss on free radical and antioxidant balance: A preliminary report. Obes. Surg. 2002, 12, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Uzun, H.; Konukoglu, D.; Gelisgen, R.; Zengin, K.; Taskin, M. Plasma protein carbonyl and thiol stress before and after laparoscopic gastric banding in morbidly obese patients. Obes. Surg. 2007, 17, 1367–1373. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Millán, J.L.S. Abdominal adiposity and the polycystic ovary syndrome. Trends Endocrinol. Metab. 2007, 18, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Attardi, G.; Schatz, G. Biogenesis of mitochondria. Annu. Rev. Cell Biol. 1988, 4, 289–333. [Google Scholar] [CrossRef]
- Keskin Kurt, R.; Okyay, A.G.; Hakverdi, A.U.; Gungoren, A.; Dolapcioglu, K.S.; Karateke, A.; Dogan, M.O. The effect of obesity on inflammatory markers in patients with PCOS: A BMI-matched case-control study. Arch. Gynecol. Obs. 2014, 290, 315–319. [Google Scholar] [CrossRef]
- Çakıroğlu, Y.; Vural, F.; Vural, B. The inflammatory markers in polycystic ovary syndrome: Association with obesity and IVF outcomes. J. Endocrinol. Invest. 2016, 39, 899–907. [Google Scholar] [CrossRef]
- Ray, K.K.; Morrow, D.A.; Sabatine, M.S.; Shui, A.; Rifai, N.; Cannon, C.P.; Braunwald, E. Long-term prognostic value of neopterin: A novel marker of monocyte activation in patients with acute coronary syndrome. Circulation 2007, 115, 3071–3078. [Google Scholar] [CrossRef]
- Abiad, F.; Khalife, D.; Safadi, B.; Alami, R.; Awwad, J.; Khalifeh, F.; Ghazeeri, G. The effect of bariatric surgery on inflammatory markers in women with polycystic ovarian syndrome. Diabetes Metab. Syndr. Clin. Res. Rev. 2018, 12, 999–1005. [Google Scholar] [CrossRef]
- Manousopoulou, A.; Al-Daghri, N.M.; Sabico, S.; Garay-Baquero, D.J.; Teng, J.; Alenad, A.; Alokail, M.S.; Athanasopoulos, N.; Deligeoroglou, E.; Chrousos, G.P.; et al. Polycystic Ovary Syndrome and Insulin Physiology: An Observational Quantitative Serum Proteomics Study in Adolescent, Normal-Weight Females. Proteom. Clin. Appl. 2019, 13, 1800184. [Google Scholar] [CrossRef]
- Kazer, R.R.; Unterman, T.G.; Glick, R.P. An Abnormality of the Growth Hormone/Insulin-Like Growth Factor-I Axis in Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1990, 71, 958–962. [Google Scholar] [CrossRef]
- Villa, P.; Soranna, L.; Mancini, A.; De Marinis, L.; Valle, D.; Mancuso, S.; Lanzone, A. Effect of feeding on growth hormone response to growth hormone-releasing hormone in polycystic ovarian syndrome: Relation with body weight and hyperinsulinism. Hum. Reprod. 2001, 16, 430–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltsas, T.; Pontikides, N.; Krassas, G.E.; Seferiadis, K.; Lolis, D.; Messinis, I.E. Effect of gonadotrophin-releasing hormone agonist treatment on growth hormone secretion in women with polycystic ovarian syndrome. Hum. Reprod. 1998, 13, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Prelević, G.M.; Wurzburger, M.I.; Balint-Perić, L.; Ginsburg, J. Twenty-four-hour serum growth hormone, insulin, C-peptide and blood glucose profiles and serum insulin-like growth factor-I concentrations in women with polycystic ovaries. Horm. Res. Paediatr. 1992, 37, 125–131. [Google Scholar]
- Lanzone, A.; Villa, P.; Fulghesu, A.M.; Pavone, V.; Caruso, A.; Mancuso, S. Endocrinology: The growth hormone response to growth hormone-releasing hormone is blunted in polycystic ovary syndrome: Relationship with obesity and hyperinsulinaemia. Hum. Reprod. 1995, 10, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Micić, D.; Šumarac-Dumanović, M.; Macut, D.; Kendereški, A.; Zorič, S.; Popovíc, V.; Cvijović, G.; Dieguez, C.; Casanueva, F.F. Growth-hormone response to combined stimulation with GHRH plus GH-releasing peptide-6 in obese patients with polycystic ovary syndrome before and after short-term fasting. J. Endocrinol. Investig. 2003, 26, 333–340. [Google Scholar]
- Lee, E.J.; Lee, B.S.; Lee, H.C.; Park, K.H.; Song, C.H.; Huh, K.B. Growth hormone response to L-dopa and pyridostigmine in women with polycystic ovarian syndrome. Fertil. Steril. 1993, 60, 53–57. [Google Scholar]
- Kadoura, S.; Alhalabi, M.; Nattouf, A.H. Effect of calcium and vitamin d supplements as an adjuvant therapy to metformin on menstrual cycle abnormalities, hormonal profile, and IGF-1 system in polycystic ovary syndrome patients: A randomized, placebo-controlled clinical trial. Adv. Pharm. Sci. 2019, 2019, 9680390. [Google Scholar] [CrossRef] [PubMed]
- Barreca, A.; Del Monte, P.; Ponzani, P.; Artini, P.G.; Genazzani, A.R.; Minuto, F. Intrafollicular insulin-like growth factor-II levels in normally ovulating women and in patients with polycystic ovary syndrome. Fertil. Steril. 1996, 65, 739–745. [Google Scholar] [CrossRef]
- Samoto, T.; Maruo, T.; Matsuo, H.; Katayama, K.; Mochizuki, M.; Barnea, E.R. Altered Expression of Insulin and Insulin-Like Growth Factor-l Receptors in Follicular and Stromal Compartments of Polycystic Ovaries. Endocr. J. 1993, 40, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucera, R.; Babuska, V.; Ulcova-Gallova, Z.; Kulda, V.; Topolcan, O. Follicular fluid levels of anti-Müllerian hormone, insulin-like growth factor 1 and leptin in women with fertility disorders. Syst. Biol. Reprod. Med. 2018, 64, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Vergani, E.; Bruno, C.; Olivieri, G.; Segni, C.D.I. Oxidative stress as a possible mechanism underlying multi-hormonal deficiency in chronic heart failure. Eur. Rev. Med. Pharmacol. 2018, 22, 3935–3960. [Google Scholar]
- Conti, E.; Pitocco, D.; Capoluongo, E.; Zuppi, C.; Ghirlanda, G.; Crea, F.; Andreotti, F.; Leinonen, E.S.; Leinonen, P.J.; Taskinen, M.R. IGF-1 and macrovascular complications of diabetes: Alternative interpretations of recently published data [21] (multiple letters). Diabetes Care 2003, 26, 1653–1655. [Google Scholar] [CrossRef] [Green Version]
- Panee, J. Monocyte Chemoattractant Protein 1 (MCP-1) in obesity and diabetes. Cytokine 2012, 60, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pouvreau, C.; Dayre, A.; Butkowski, E.; de Jong, B.; Jelinek, H.F. Inflammation and oxidative stress markers in diabetes and hypertension. J. Inflamm. Res. 2018, 11, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Juul, A.; Scheike, T.; Davidsen, M.; Gyllenborg, J.; Jørgensen, T. Low serum insulin-like growth factor I is associated with increased risk of ischemic heart disease: A population-based case-control study. Circulation 2002, 106, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Shai, S.Y.; Sukhanov, S.; Higashi, Y.; Vaughn, C.; Rosen, C.J.; Delafontaine, P. Low circulating insulin-like growth factor I increases atherosclerosis in ApoE-deficient mice. Am. J. Physiol. Hear. Circ. Physiol. 2011, 300, H1898–H1906. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, G.A.; Barrett-Connor, E.; Criqui, M.H.; Kritz-Silverstein, D. The Prospective Association of Serum Insulin-Like Growth Factor I (IGF-I) and IGF-Binding Protein-1 Levels with All Cause and Cardiovascular Disease Mortality in Older Adults: The Rancho Bernardo Study. J. Clin. Endocrinol. Metab. 2004, 89, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiGirolamo, M.; Edén, S.; Enberg, G.; Isaksson, O.; Lönnroth, P.; Hall, K.; Smith, U. Specific binding of human growth hormone but not insulin-like growth factors by human adipocytes. Febs Lett. 1986, 205, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Ita, J.R.; Castilla-Cortázar, I.; Aguirre, G.A.; Sánchez-Yago, C.; Santos-Ruiz, M.O.; Guerra-Menéndez, L.; Martín-Estal, I.; García-Magariño, M.; Lara-Díaz, V.J.; Puche, J.E.; et al. Altered liver expression of genes involved in lipid and glucose metabolism in mice with partial IGF-1 deficiency: An experimental approach to metabolic syndrome. J. Transl. Med. 2015, 13, 1–13. [Google Scholar]
- Succurro, E.; Andreozzi, F.; Marini, M.A.; Lauro, R.; Hribal, M.L.; Perticone, F.; Sesti, G. Low plasma insulin-like growth factor-1 levels are associated with reduced insulin sensitivity and increased insulin secretion in nondiabetic subjects. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 713–719. [Google Scholar] [CrossRef]
- Sesti, G.; Sciacqua, A.; Cardellini, M.; Marini, M.A.; Maio, R.; Vatrano, M.; Succurro, E.; Lauro, R.; Federici, M.; Perticone, F. Plasma concentration of IGF-I is independently associated with insulin sensitivity in subjects with different degrees of glucose tolerance. Diabetes Care 2005, 28, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.H.; Frystyk, J.; Andersen, T.; Breum, L.; Christiansen, J.S.; Hilsted, J. The impact of obesity, fat distribution, and energy restriction on insulin-like growth factor-1 (IGF-1), IGF-binding protein-3, insulin, and growth hormone. Metabolism 1994, 43, 315–319. [Google Scholar] [CrossRef]
- De Pergola, G. Divergent effects of short-term, very-low-calorie diet on insulin-like growth factor-I and insulin-like growth factor binding protein-3 serum concentrations in premenopausal women with obesity. Obes. Res. 1998, 6, 408–415. [Google Scholar] [CrossRef]
- Kunitomi, M.; Wada, J.; Takahashi, K.; Tsuchiyama, Y.; Mimura, Y.; Hida, K.; Miyatake, N.; Fujii, M.; Kira, S.; Shikata, K.; et al. Relationship between reduced serum IGF-I levels and accumulation of visceral fat in Japanese men. Int. J. Obes. 2002, 26, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Marin, P.; Kvist, H.; Lindstedt, G.; Sjostrom, L.; Bjorntorp, P. Low concentrations of insulin-like growth factor-I in abdominal obesity. Int. J. Obes. 1993, 17, 83–89. [Google Scholar]
- Rasmussen, M.H.; Hvidberg, A.; Juul, A.; Main, K.M.; Gotfredsen, A.; Skakkebaek, N.E.; Hilsted, J. Massive weight loss restores 24-hour growth hormone release profiles and serum insulin-like growth factor-I levels in obese subjects. J. Clin. Endocrinol. Metab. 1995, 80, 1407–1415. [Google Scholar] [CrossRef]
- Zhang, Y.; Yuan, M.; Bradley, K.M.; Dong, F.; Anversa, P.; Ren, J. Insulin-like growth factor 1 alleviates high-fat diet-induced myocardial contractile dysfunction: Role of insulin signaling and mitochondrial function. Hypertension 2012, 59, 680–693. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Lin, Z.; Robb, S.W.; Ezeamama, A.E. Serum vitamin d levels and polycystic ovary syndrome: A systematic review and meta-analysis. Nutrients 2015, 7, 4555–4577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimalawansa, S.J. Vitamin D deficiency: Effects on oxidative stress, epigenetics, gene regulation, and aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Ryan, Z.C.; Craig, T.A.; Folmes, C.D.; Wang, X.; Lanza, I.R.; Schaible, N.S.; Salisbury, J.L.; Nair, K.S.; Terzic, A.; Sieck, G.C.; et al. 1α,25-dihydroxyvitamin D3 regulates mitochondrial oxygen consumption and dynamics in human skeletal muscle cells. J. Biol. Chem. 2016, 291, 1514–1528. [Google Scholar] [CrossRef] [Green Version]
- Bouillon, R.; Verstuyf, A. Vitamin D, mitochondria, and muscle. J. Clin. Endocrinol. Metab. 2013, 98, 961–963. [Google Scholar] [CrossRef]
- Wang, L.; Lewis, T.; Zhang, Y.-L.; Khodier, C.; Magesh, S.; Chen, L.; Inoyama, D.; Chen, Y.; Zhen, J.; Hu, L.; et al. The Identification and Characterization of Non-Reactive Inhibitor of Keap1-Nrf2 Interaction through HTS Using a Fluorescence Polarization Assay. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information: Bethesda, MD, USA, 2010. [Google Scholar]
- Song, C.; Fu, B.; Zhang, J.; Zhao, J.; Yuan, M.; Peng, W.; Zhang, Y.; Wu, H. Sodium fluoride induces nephrotoxicity via oxidative stress-regulated mitochondrial SIRT3 signaling pathway. Sci. Rep. 2017, 7, 1–15. [Google Scholar]
- Wei, R.; Christakos, S. Mechanisms underlying the regulation of innate and adaptive immunity by vitamin D. Nutrients 2015, 7, 8251–8260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beilfuss, J.; Berg, V.; Sneve, M.; Jorde, R.; Kamycheva, E. Effects of a 1-year supplementation with cholecalciferol on interleukin-6, tumor necrosis factor-alpha and insulin resistance in overweight and obese subjects. Cytokine 2012, 60, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Calton, E.K.; Keane, K.N.; Soares, M.J.; Rowlands, J.; Newsholme, P. Prevailing vitamin D status influences mitochondrial and glycolytic bioenergetics in peripheral blood mononuclear cells obtained from adults. Redox Biol. 2016, 10, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hyde, A.S.; Simpson, M.A.; Barycki, J.J. Emerging regulatory paradigms in glutathione metabolism. Adv. Cancer Res. 2014, 122, 69–101. [Google Scholar] [PubMed] [Green Version]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Heydari, S.T.; Kavari, S.H.; Mirhosseini, N.; Mafi, A.; Dastorani, M.; et al. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress among Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm. Metab. Res. 2018, 50, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Foroozanfard, F.; Jamilian, M.; Bahmani, F.; Talaee, R.; Talaee, N.; Hashemi, T.; Nasri, K.; Asemi, Z.; Esmaillzadeh, A. Calcium plus vitamin D supplementation influences biomarkers of inflammation and oxidative stress in overweight and vitamin D-deficient women with polycystic ovary syndrome: A randomized double-blind placebo-controlled clinical trial. Clin. Endocrinol. 2015, 83, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Razavi, M.; Jamilian, M.; Karamali, M.; Bahmani, F.; Aghadavod, E.; Asemi, Z. The Effects of Vitamin D-K-Calcium Co-Supplementation on Endocrine, Inflammation, and Oxidative Stress Biomarkers in Vitamin D-Deficient Women with Polycystic Ovary Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Horm. Metab. Res. 2016, 48, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, F.; Mohammadzadeh, A.; Sadeghi, M.R.; Akhondi, M.M.; Mohammadmoradi, S.; Kamali, K.; Lackpour, N.; Jouhari, S.; Zafadoust, S.; Mokhtar, S.; et al. Role of vitamin E and D3 supplementation in Intra-Cytoplasmic Sperm Injection outcomes of women with polycystic ovarian syndrome: A double blinded randomized placebo-controlled trial. Clin. Nutr. Espen 2017, 18, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.S. FGF23, klotho and vitamin D interactions: What have we learned from in vivo mouse genetics studies? Adv. Exp. Med. Biol. 2012, 728, 84–91. [Google Scholar]
- Jain, S.K.; Parsanathan, R.; Achari, A.E.; Kanikarla-Marie, P.; Bocchini, J.A. Glutathione Stimulates Vitamin D Regulatory and Glucose-Metabolism Genes, Lowers Oxidative Stress and Inflammation, and Increases 25-Hydroxy-Vitamin D Levels in Blood: A Novel Approach to Treat 25-Hydroxyvitamin D Deficiency. Antioxid. Redox Signal. 2018, 29, 1792–1807. [Google Scholar] [CrossRef]
- Crivello, J.F. Oxidative stress limits vitamin D metabolism by bovine proximal tubule cellsin vitro. Arch. Biochem. Biophys. 1988, 262, 471–480. [Google Scholar] [CrossRef]
- Erkan, G.; Tayyar, A.T.; Açmaz, G.; Müderris, İ.İ.; Başkol, G.; Bayram, F. Polikistik over sendromlu hastalarda insülin direncinde osteokalsin, tümör nekroz faktörü-alfa ve adiponektinin rolü. Turk Jinekoloji Ve Obs. Dern. Derg. 2017, 14, 89–93. [Google Scholar]
- Lingaiah, S.; Morin-Papunen, L.; Piltonen, T.; Puurunen, J.; Sundström-Poromaa, I.; Stener-Victorin, E.; Bloigu, R.; Risteli, J.; Tapanainen, J.S. Bone markers in polycystic ovary syndrome: A multicentre study. Clin. Endocrinol. 2017, 87, 673–679. [Google Scholar] [CrossRef]
- Pepene, C.E. Serum under-carboxylated osteocalcin levels in women with polycystic ovary syndrome: Weight-dependent relationships with endocrine and metabolic traits. J. Ovarian Res. 2013, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Razny, U.; Goralska, J.; Zdzienicka, A.; Fedak, D.; Masania, J.; Rabbani, N.; Thornalley, P.; Pawlica-Gosiewska, D.; Gawlik, K.; Dembinska-Kiec, A.; et al. Relation of the protein glycation, oxidation and nitration to the osteocalcin level in obese subjects. Acta Biochim. Pol. 2017, 64, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamal, S.M.; Sadek, N.B.; Rashed, L.A.; Shawky, H.M.; Gamal El-Din, M.M. Effect of gamma-carboxylase inhibition on serum osteocalcin may be partially protective against developing diabetic cardiomyopathy in type 2 diabetic rats. Diabetes Vasc. Dis. Res. 2016, 13, 405–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Lu, P.; Guo, X.; Song, K.; Lyu, Y.; Bothwell, J.; Wu, J.; Hawkins, O.; Clarke, S.L.; Lucas, E.A.; et al. β-carotene oxygenase 2 deficiency-triggered mitochondrial oxidative stress promotes low-grade inflammation and metabolic dysfunction. Free Radic. Biol. Med. 2021, 164, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, S.; Siejka, A. The pathogenesis and treatment of polycystic ovary syndrome: What’s new? Adv. Clin. Exp. Med. 2017, 26, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Brunetti, A.; Bruno, C.; Vergani, E.; Pocino, K.; Napodano, C.; Gulli, F.; Santini, S.A.; Basile, U. Plasmatic free light chains in polycystic ovary syndrome. Gynecol. Endocrinol. 2019, 35, 710–713. [Google Scholar] [CrossRef]
- Nasri, F.; Doroudchi, M.; Jahromi, B.N.; Gharesi-Fard, B. T helper cells profile and cd4+cd25+foxp3+regulatory t cells in polycystic ovary syndrome. Iran. J. Immunol. 2018, 15, 175–185. [Google Scholar]
- Dumitriu, I.E.; Araguás, E.T.; Baboonian, C.; Kaski, J.C. CD4+CD28null T cells in coronary artery disease: When helpers become killers. Cardiovasc. Res. 2009, 81, 11–19. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancini, A.; Bruno, C.; Vergani, E.; d’Abate, C.; Giacchi, E.; Silvestrini, A. Oxidative Stress and Low-Grade Inflammation in Polycystic Ovary Syndrome: Controversies and New Insights. Int. J. Mol. Sci. 2021, 22, 1667. https://doi.org/10.3390/ijms22041667
Mancini A, Bruno C, Vergani E, d’Abate C, Giacchi E, Silvestrini A. Oxidative Stress and Low-Grade Inflammation in Polycystic Ovary Syndrome: Controversies and New Insights. International Journal of Molecular Sciences. 2021; 22(4):1667. https://doi.org/10.3390/ijms22041667
Chicago/Turabian StyleMancini, Antonio, Carmine Bruno, Edoardo Vergani, Claudia d’Abate, Elena Giacchi, and Andrea Silvestrini. 2021. "Oxidative Stress and Low-Grade Inflammation in Polycystic Ovary Syndrome: Controversies and New Insights" International Journal of Molecular Sciences 22, no. 4: 1667. https://doi.org/10.3390/ijms22041667
APA StyleMancini, A., Bruno, C., Vergani, E., d’Abate, C., Giacchi, E., & Silvestrini, A. (2021). Oxidative Stress and Low-Grade Inflammation in Polycystic Ovary Syndrome: Controversies and New Insights. International Journal of Molecular Sciences, 22(4), 1667. https://doi.org/10.3390/ijms22041667