
A Novel Triplet-Primed PCR Assay to Detect the Full Range of Trinucleotide CAG Repeats in the Huntingtin Gene (HTT)

 , , and
, , and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. AmplideX PCR/CE HTT Kit
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Squitieri, F. Neurodegenerative disease: ‘Fifty shades of grey’ in the Huntington disease gene. Nat. Rev. Neurol. 2013, 9, 421–422. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marder, K.; Zhao, H.; Myers, R.J.; Cudkowics, M.; Kayson, E.; Kieburtz, K.; Orme, C.; Paulsen, J.; Penney, J.B.; Siemers, E.; et al. Rate of functional decline in Huntington’s disease. Huntington Study Group. Neurology 2000, 54, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Keum, J.W.; Shin, A.; Gillis, T.; Mysore, J.S.; Elneel, K.A.; Lucente, D.; Hadzi, T.; Holmans, P.; Jones, L.; Orth, M.; et al. The HTT CAG-Expansion Mutation Determines Age at Death but Not Disease Duration in Huntington Disease. Am. J. Hum. Genet. 2016, 98, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Kremer, B.; Goldberg, P.; Andrew, S.E.; Theilmann, J.; Telenius, H.; Zeisler, J.; Squitieri, F.; Lin, B.; Bassett, A.; Almqvist, E.; et al. A worldwide study of the Huntington’s disease mutation: The sensitivity and specificity of measuring CAG repeats. N. Engl. J. Med. 1994, 330, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Leggo, J.; Coles, R.; Almqvist, E.; Biancalana, V.; Cassiman, J.J.; Chotai, K.; Connarty, M.; Craufurd, D.; Curtis, A.; et al. Phenotypic characterization of individuals with 30–40 CAG repeats in the Huntington disease (HD) gene reveals HD cases with 36 repeats and apparently normal elderly individuals with 36–39 repeats. Am. J. Hum. Genet. 1996, 59, 16–22. [Google Scholar]
- Quarrell, O.W.J.; Brewer, H.M.; Squitieri, F.; Barker, R.A.; Nance, M.A.; Landwehrmeyer, G.B. Juvenile Huntington’s Disease; Quarrell, O.W.J., Brewer, H.M., Squitieri, F., Barker, R.A., Nance, M.A., Landwehrmeyer, G.B., Eds.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Fusilli, C.; Migliore, S.; Mazza, T.; Consoli, F.; De Luca, A.; Barbagallo, G.; Ciammola, A.; Gatto, E.M.; Cesarini, M.; Etcheverry, J.L.; et al. Biological and clinical manifestations of juvenile Huntington’s disease: A retrospective analysis. Lancet Neurol. 2018, 17, 986–993. [Google Scholar] [CrossRef]
- Pearson, C.E.; Edamura, K.N.; Cleary, J.D. Repeat instability: Mechanisms of dynamic mutations. Nat. Rev. Genet. 2005, 6, 729–742. [Google Scholar] [CrossRef]
- Telenius, H.; Kremer, B.; Goldberg, Y.P.; Theilmann, J.; Andrew, S.E.; Zeisler, J.; Adam, S.; Greenberg, C.; Ives, E.J.; Clarke, L.A.; et al. Somatic and gonadal mosaicism of the Huntington disease gene CAG repeat in brain and sperm. Nat. Genet. 1994, 6, 409–414. [Google Scholar] [CrossRef]
- Kennedy, L.; Evans, E.; Chen, C.M.; Craven, L.; Detloff, P.J.; Ennis, M.; Shelbourne, P.F. Dramatic tissue-specific mutation length increases are an early molecular event in Huntington disease pathogenesis. Hum. Mol. Genet. 2003, 12, 3359–3367. [Google Scholar] [CrossRef] [PubMed]
- Genetic Modifiers of Huntington’s Disease (GeM-HD) Consortium. Identification of Genetic Factors that Modify Clinical Onset of Huntington’s Disease. Cell 2015, 162, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensman Moss, D.J.; Pardiñas, A.F.; Langbehn, D.; Lo, K.; Leavitt, B.R.; Roos, R.A.C.; Durr, A.; Mead, S. Identification of genetic variants associated with Huntington’s disease progression. A genome-wide association study. Lancet Neurol. 2017, 16, 701–711. [Google Scholar] [CrossRef]
- Ciosi, M.; Maxwell, A.; Cumming, S.A.; Hensman Moss, D.J.; Alshammari, A.M.; Flower, M.D.; Durr, A.; Leavitt, B.R.; Roos, R.A.C.; The Track-HD Team; et al. A genetic association study of glutamine-encoding DNA sequence structures, somatic CAG expansion, and DNA repair gene variants, with Huntington disease clinical outcomes. EBioMedicine 2019, 48, 568–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, G.E.B.; Collins, J.A.; Kay, C.; McDonald, C.; Dolzhenko, E.; Xia, Q.; Becanovic, K.; Drogemoller, B.I.; Semaka, A.; Nguyen, C.M.; et al. Length of Uninterrupted CAG, Independent of Polyglutamine Size, Results in Increased Somatic Instability, Hastening Onset of Huntington Disease. Am. J. Hum. Genet. 2019, 104, 1116–1126. [Google Scholar] [CrossRef] [Green Version]
- Black, H.F.; Wright, G.E.B.; Collins, J.A.; Caron, N.; Kay, C.; Xia, Q.; Arning, L.; Bijlsma, E.K.; Squitieri, F.; Nguyen, H.P.; et al. Frequency of the loss of CAA interruption in the HTT CAG tract and implications for Huntington disease in the reduced penetrance range. Genet. Med. 2020, 22, 2108–2113. [Google Scholar] [CrossRef]
- Wexler, N.S. The Venezuela Collaborative Research Project: Venezuelan kindreds reveal that genetic and environmental factors modulate Huntington’s disease age of onset. Proc. Natl. Acad. Sci. USA 2004, 101, 3498–3503. [Google Scholar]
- Goldberg, Y.P.; McMurray, C.T.; Zeisier, J.; Almqvist, E.; Sillence, D.; Richards, F.; Gacy, A.M.; Buchanan, J.; Telenius, H.; Hayden, M.R. Increased instability of intermediate alleles in families with sporadic Huntington disease compared to similar sized intermediate alleles in the general population. Hum. Mol. Genet. 1995, 4, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Suarez, A.; Gandia-Pla, S.; Gòmez-Llorente, C.; Antùnez, A.; Gòmez-Capilla, J.A.; Fàrez-Vidal, M.E. Use of capillary electrophoresis for accurate determination of CAG repeats causing Huntington disease. An oligonucleotide design avoiding shadow bands. Scand. J. Clin. Lab. Investig. 2009, 68, 577–584. [Google Scholar] [CrossRef]
- Andrew, S.E.; Goldberg, Y.P.; Theilmann, J.; Zeisler, J.; Hayden, M.R. A CCG repeat polymorphism adjacent to the CAG repeat in the Huntington disease gene: Implications for diagnostic accuracy and predictive testing. Hum. Mol. Genet. 1994, 3, 65–67. [Google Scholar] [CrossRef]
- Tóth, T.; Findlay, I.; Papp, C.; Tóth-Pál, E.; Marton, T.; Nagy, B.; Quirke, P.; Papp, Z. Prenatal detection of trisomy 21 and 18 from amniotic fluid by quantitative fluorescent polymerase chain reaction. J. Med. Genet. 1998, 35, 126–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guida, M.; Fenwick, R.G.; Papp, A.C.; Snyder, P.J.; Sedra, M.; Prior, T.W. Southern transfer protocol for confirmation of Huntington disease. Clin. Chem. 1996, 42, 1711–1712. [Google Scholar] [CrossRef] [Green Version]
- Gellera, C.; Meoni, C.; Castellotti, B.; Zappacosta, B.; Girotti, F.; Taroni, F.; Di Donato, S. Errors in Huntington disease diagnostic test caused by trinucleotide deletion in the IT15 gene. Am. J. Hum. Genet. 1996, 59, 475–477. [Google Scholar]
- Magri, S.; Nanetti, L.; Mongelli, A.; Rizzo, E.; Taroni, F.; Mariotti, C.; Gellera, C. Missing the pathological expansion in Huntington disease: De novo c.51C > G variant on the expanded allele causing intrafamilial allele dropout. Am. J. Med. Genet. A 2020. Online ahead of print. [Google Scholar] [CrossRef]
- Warner, J.P.; Barron, L.H.; Goudie, D.; Kelly, K.; Dow, D.; Fitzpatrick, D.R.; Brock, D.J.H. A general method for the detection of large CAG repeat expansions by fluorescent PCR. J. Med. Genet. 1996, 33, 1022–1026. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Lee, C.G.; Law, H.Y.; Chong, S.S. Enhanced Detection and Sizing of the HTT CAG Repeat Expansion in Huntington Disease Using an Improved Triplet-Primed PCR Assay. Neurodegener. Dis. 2016, 16, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Jama, M.; Millson, A.; Miller, C.E.; Lyon, E. Triplet Repeat Primed PCR Simplifies Testing for Huntington Disease. J. Mol. Diagn. 2013, 15, 255–262. [Google Scholar] [CrossRef]
- Bean, L.; Bayrak-Toydemir, P. American College of Medical Genetics and Genomics Standards and Guidelines for Clinical Genetics Laboratories, 2014 edition: Technical standards and guidelines for Huntington disease. Genet. Med. 2014, 16, e2. [Google Scholar] [CrossRef] [Green Version]
- Kalman, L.; Johnson, M.A.; Beck, J.; Berry-Kravis, E.; Buller, A.; Casey, B.; Feldman, G.L.; Handsfield, J.; Jakupciak, J.P.; Maragh, S.; et al. Development of genomic reference materials for Huntington disease genetic testing. Genet. Med. 2007, 9, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Losekoot, M.; Van Belzen, M.J.; Seneca, S.; Bauer, P.; Stenhouse, S.A.; Barton, D.E. European Molecular Genetic Quality Network (EMQN). EMQN/CMGS best practice guidelines for the molecular genetic testing of Huntington disease. Eur. J. Hum. Genet. 2013, 21, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Cheah, F.S.H.; Chen, M.; Lee, C.G.; Law, H.Y.; Chong, S.S. Improved high sensitivity screen for Huntington disease using a one-step triplet-primed PCR and melting curve assay. PLoS ONE 2017, 12, e0180984. [Google Scholar] [CrossRef]
- Squitieri, F.; Mazza, T.; Maffi, S.; De Luca, A.; AlSalmi, Q.; AlHarasi, S.; Collins, J.A.; Kay, C.; Baine-Savanhu, F.; Landwhermeyer, B.G.; et al. Tracing the mutated HTT and haplotype of the African ancestor who spread Huntington disease into the Middle East. Genet. Med. 2020, 22, 1903–1908. [Google Scholar] [CrossRef]
- European Medicines Agency EMA decision CW/0001/2015. Available online: http://www.ema.europa.eu/docs/en_GB/document_library/Other/2015/07/WC500190385pdf (accessed on 21 December 2018).
- Migliore, S.; Jankovic, J.; Squitieri, F. Genetic Counseling in Huntington’s Disease: Potential New Challenges on Horizon? Front. Neurol. 2019, 30, 453. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Hayeems, R.Z.; Shuman, C.; Szego, M.J.; Monfared, N.; Bowdin, S.; Zlotnik Shaul, R.; Mein, M.S. Predictive genetic testing for adult-onset disorders in minors: A critical analysis of the arguments for and against the 2013 ACMG guidelines. Clin. Genet. 2015, 87, 301–310. [Google Scholar] [CrossRef]
- Dolzhenko, E.; Deshpande, V.; Shlesinger, F.; Krusche, P.; Petrovski, R.; Chen, S.; Emig-Agius, D.; Gross, A.; Narzisi, G.; Bowman, B.; et al. ExpansionHunter: A sequence-graph-based tool to analyze variation in short tandem repeat regions. Bioinformatics 2019, 35, 4754–4756. [Google Scholar] [CrossRef] [Green Version]
- Filipovic-Sadic, S.; Sah, S.; Chen, L.; Krosting, J.; Sekinger, E.; Zhang, W.; Hagerman, P.J.; Stenzel, T.T.; Hadd, A.G.; Latham, G.J.; et al. A novel FMR1 PCR method for the routine detection of low abundance expanded alleles and full mutations in fragile X syndrome. Clin. Chem. 2010, 56, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bram, E.; Javanmardi, K.; Nicholson, K.; Culp, K.; Thibert, J.R.; Kemppainen, J.; Le, V.; Schlageter, A.; Hadd, A.; Latham, G.J. Comprehensive genotyping of the C9orf72 hexanucleotide repeat region in 2095 ALS samples from the NINDS collection using a two-mode, long-read PCR assay. Amyotroph. Lateral Scler. Front. Degener. 2018, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Zhang, A.; Dlouhy, S.; Bai, S. Detection of large expansions in myotonic dystrophy type 1 using triplet primed PCR. Front. Genet. 2014, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Thibert, J.R.; Nicholson, K.; Wisotsky, J.; Le, V.; Latham, G.J.; Statt, S. A Robust PCR/CE Assay Using AmplideX Technology for Rapid and Accurate Genotyping of CAG Repeat Expansions in HTT. Poster 2018. In Proceedings of the ACMG Annual Clinical Genetics Meeting, Charlotte, NC, USA, 10–14 April 2018. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vendor ID | Input (ng) | Categorical Bin | Published (Kalman et al., 2007) | Measured AmplideX PCR/CE HTT Kit | ||
|---|---|---|---|---|---|---|
| Allele 1 | Allele 2 | Allele 1 | Allele 2 | |||
| NA20245 | 20 | Normal | 15 | 15 | 15 | 15 |
| NA20206 | 20 | Normal | 17 | 18 | 17 | 18 |
| NA20207 | 20 | Normal | 19 | 21 | 19 | 21 |
| NA20246 | 20 | Normal | 15 | 24 | 15 | 24 |
| NA20247 | 20 | Intermediate | 15 | 29 | 15 | 29 |
| NA20248 | 20 | Reduced Penetrance | 17 | 36 | 17 | 36 |
| NA20249 | 20 | Reduced Penetrance | 22 | 39 | 22 | 39 |
| NA20208 | 20 | Expanded | 35 | 45 | 35 | 45 |
| NA20209 | 20 | Expanded | 45 | 47 | 45 | 47 |
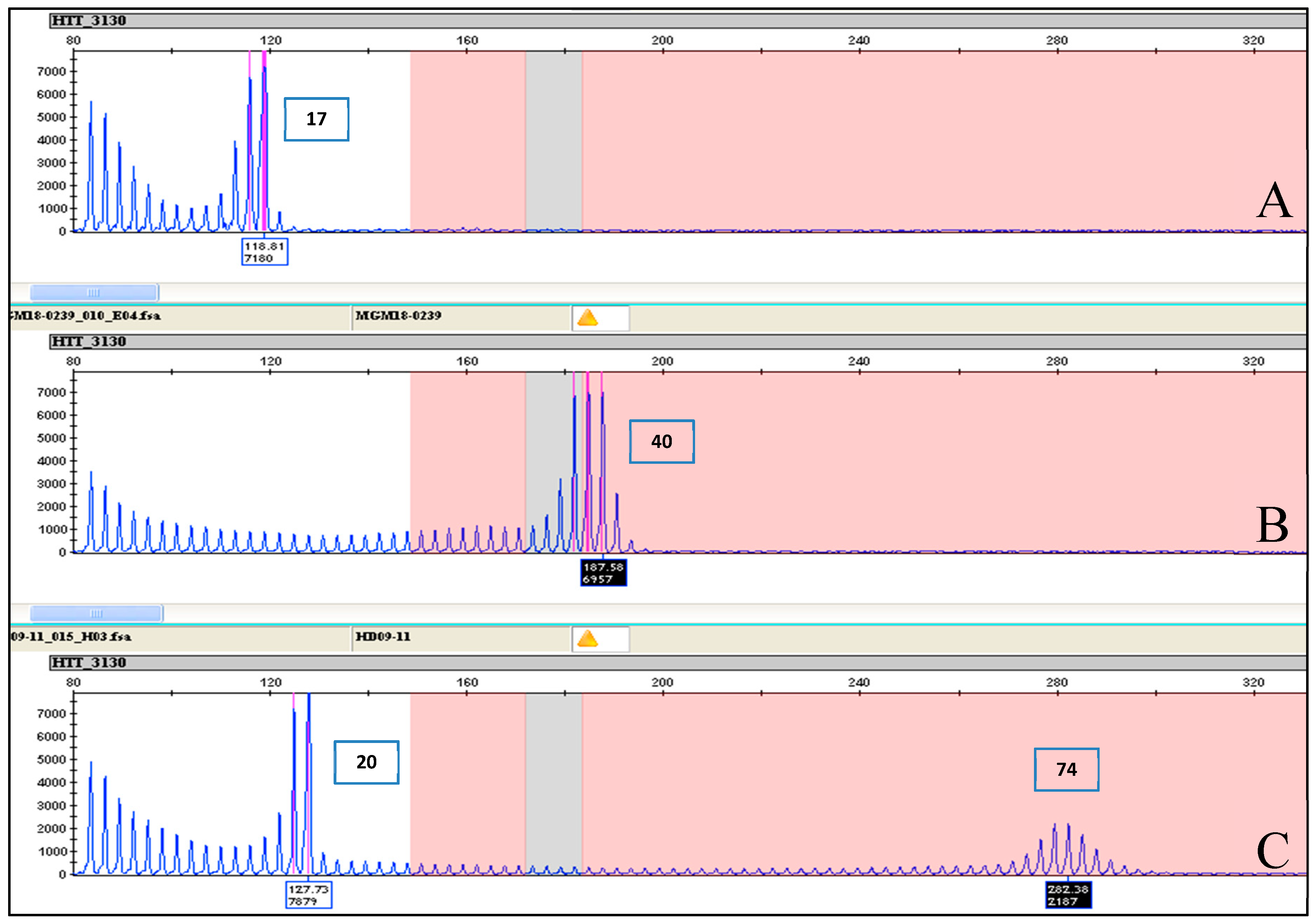
| NA20210 | 20 | Expanded | 17 | 74 | 17 | 75 |
| NA20250 | 20 | Expanded | 15 | 40 | 15 | 40 |
| NA20251 | 20 | Expanded | 39 | 50 | 39 | 50 |
| NA20252 | 20 | Expanded | 22 | 65 & 66 | 22 | 66 * |
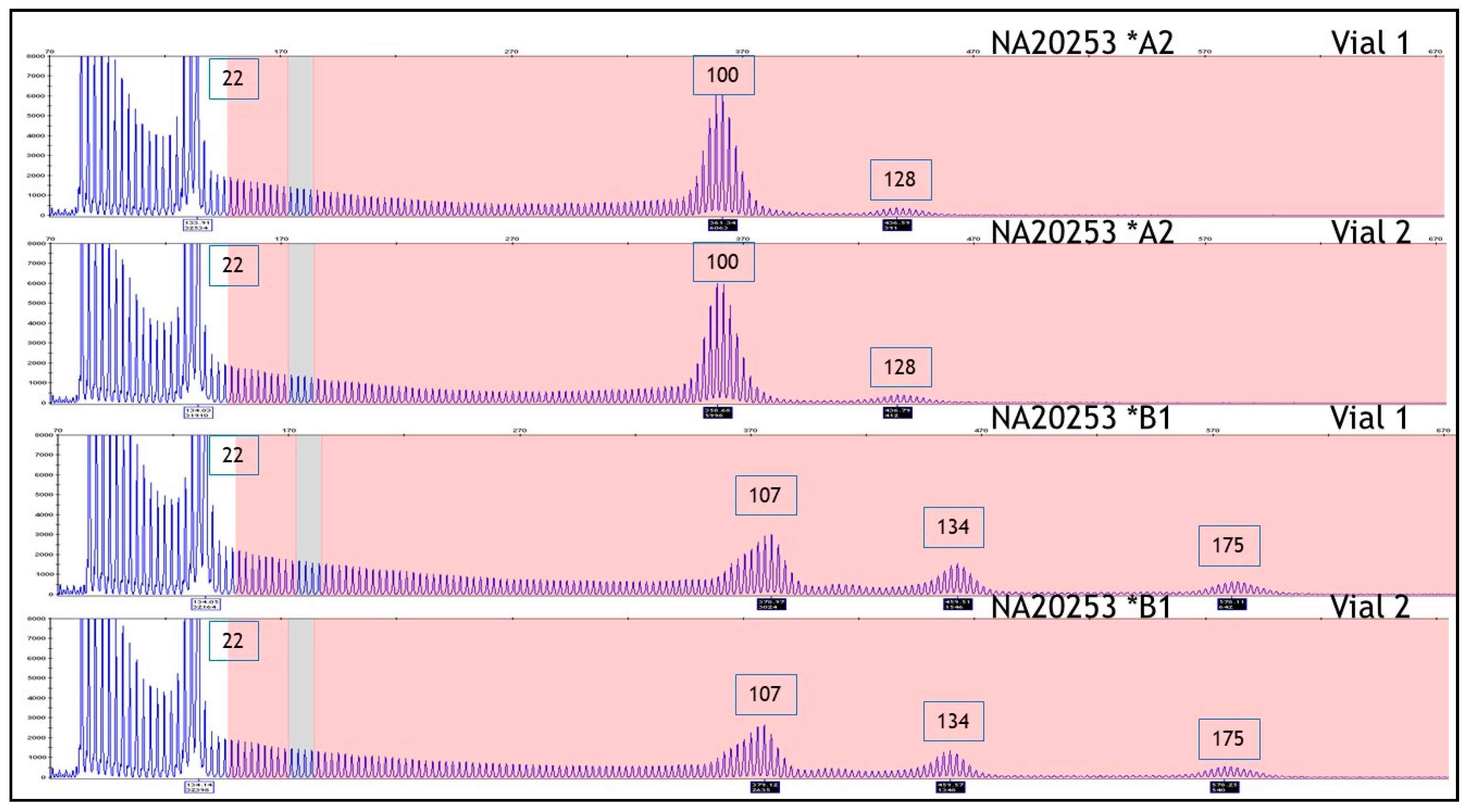
| NA20253 | 20 | Expanded | 22 | 100 | 22 | 100 ** |
| LAB ID | Categorical Bin | “In House” PCR | AmplideX PCR/CE HTT Kit | ||
|---|---|---|---|---|---|
| Allele 1 | Allele 2 | Allele 1 | Allele 2 | ||
| MGM17-1760 | Normal | 17 | 17 | 17 | 17 |
| MGM18-1250 | Normal | 17 | 17 | 17 | 17 |
| MGM17-0052 * | Intermediate | 15 | 29 | 15 | 29 |
| MGM17-0089 | Intermediate | 19 | 32 | 19 | 32 |
| MGM16-0689 | Reduced penetrance | 17 | 37 | 17 | 37 |
| HD157-02 | Reduced penetrance | 17 | 37 | 17 | 37 |
| MGM16-0828 | Reduced penetrance | 17 | 37 | 17 | 37 |
| MGM17-1738 | Reduced penetrance | 17 | 39 | 17 | 39 |
| MGM15-1651 | Reduced penetrance | 24 | 39 | 24 | 39 |
| HD23-05 | Reduced penetrance | 33 | 39 | 33 | 39 |
| MGM16-0607 | Full penetrance | 20 | 40 | 20 | 40 |
| MGM18-0177 | Full penetrance | 14 | 41 | 14 | 41 |
| CTRL-DNA_S0162119 * | Full penetrance | 17 | 41 | 17 | 41 |
| MGM16-0196 | Full penetrance | 17 | 41 | 17 | 41 |
| MGM18-0239 | Full penetrance | 40 | 40 | 40 | 40 |
| CTRL-DNA_S0162120 * | Full penetrance | 15 | 42 | 15 | 42 |
| MGM16-0086 | Full penetrance | 18 | 42 | 18 | 42 |
| MGM17-1838 | Full penetrance | 17 | 43 | 17 | 43 |
| HD672-01 | Full penetrance | 17 | 44 | 17 | 44 |
| MGM16-0250 | Full penetrance | 17 | 46 | 16 | 46 |
| MGM18-0181 | Full penetrance | 40 | 47 | 40 | 47 |
| HD423-02 | Full penetrance | 22 | 50 | 22 | 50 |
| HD09-08 | Full penetrance | 18 | 59 | 18 | 59 |
| HD87-07 | Full penetrance | 17 | 62 | 17 | 62 |
| MGM17-1716 | Full penetrance | 17 | 63 | 17 | 63 |
| MGM17-0054 * | Full penetrance | 22 | 63 | 22 | 63 |
| MGM18-0054 | Full penetrance | 22 | 63 | 22 | 63 |
| MGM18_0054 | Full penetrance | 22 | 63 | 22 | 63 |
| HD438-01 | Full penetrance | 17 | 66 | 17 | 66 |
| MGM17-1387 | Full penetrance | 20 | 74 | 20 | 74 |
| HD09-11 | Full penetrance | 20 | 74 | 20 | 74 |
| MGM16-1030 | Full penetrance | 26 | 76 | 26 | 76 |
| HD315-01 | Full penetrance | 26 | 78 | 26 | 78 |
| MGM16-1033 | Full penetrance | 17 | 80 | 17 | 81 |
| MGM16-1032 | Full penetrance | 33 | 86 | 33 | 87 |
| HD636-01 | Full penetrance | 17 | 86 | 17 | 86 |
| HD09-12 | Full penetrance | 18 | 86 | 18 | 86 |
| MGM16-1031 | Full penetrance | 19 | 88 | 19 | 89 |
| HD379-05 | Full penetrance | 18 | 89 | 18 | 89 |
| MGM16-1034 | Full penetrance | 18 | 96 | 18 | 96 |
| HD130-05 | Full penetrance | 25 | 98 | 25 | 98 |
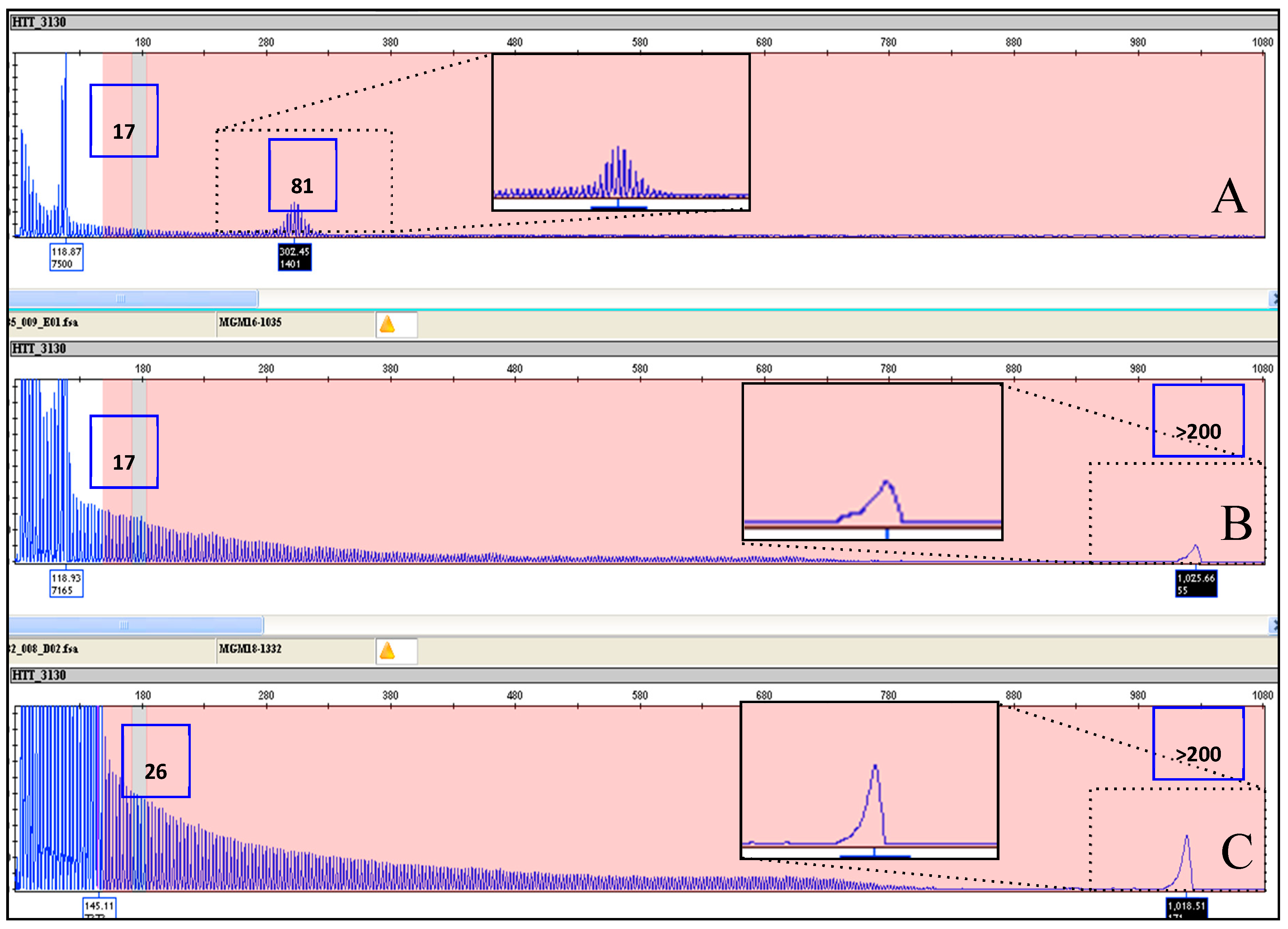
| MGM16-1035 | Full penetrance | 17 | 84 | 17 | >200 |
| MGM18-1332 | Full penetrance | 26 | 95 | 26 | >200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, A.; Morella, A.; Consoli, F.; Fanelli, S.; Thibert, J.R.; Statt, S.; Latham, G.J.; Squitieri, F. A Novel Triplet-Primed PCR Assay to Detect the Full Range of Trinucleotide CAG Repeats in the Huntingtin Gene (HTT). Int. J. Mol. Sci. 2021, 22, 1689. https://doi.org/10.3390/ijms22041689
De Luca A, Morella A, Consoli F, Fanelli S, Thibert JR, Statt S, Latham GJ, Squitieri F. A Novel Triplet-Primed PCR Assay to Detect the Full Range of Trinucleotide CAG Repeats in the Huntingtin Gene (HTT). International Journal of Molecular Sciences. 2021; 22(4):1689. https://doi.org/10.3390/ijms22041689
Chicago/Turabian StyleDe Luca, Alessandro, Annunziata Morella, Federica Consoli, Sergio Fanelli, Julie R. Thibert, Sarah Statt, Gary J. Latham, and Ferdinando Squitieri. 2021. "A Novel Triplet-Primed PCR Assay to Detect the Full Range of Trinucleotide CAG Repeats in the Huntingtin Gene (HTT)" International Journal of Molecular Sciences 22, no. 4: 1689. https://doi.org/10.3390/ijms22041689
APA StyleDe Luca, A., Morella, A., Consoli, F., Fanelli, S., Thibert, J. R., Statt, S., Latham, G. J., & Squitieri, F. (2021). A Novel Triplet-Primed PCR Assay to Detect the Full Range of Trinucleotide CAG Repeats in the Huntingtin Gene (HTT). International Journal of Molecular Sciences, 22(4), 1689. https://doi.org/10.3390/ijms22041689