
hiPSC-Derived Epidermal Keratinocytes from Ichthyosis Patients Show Altered Expression of Cornification Markers


 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
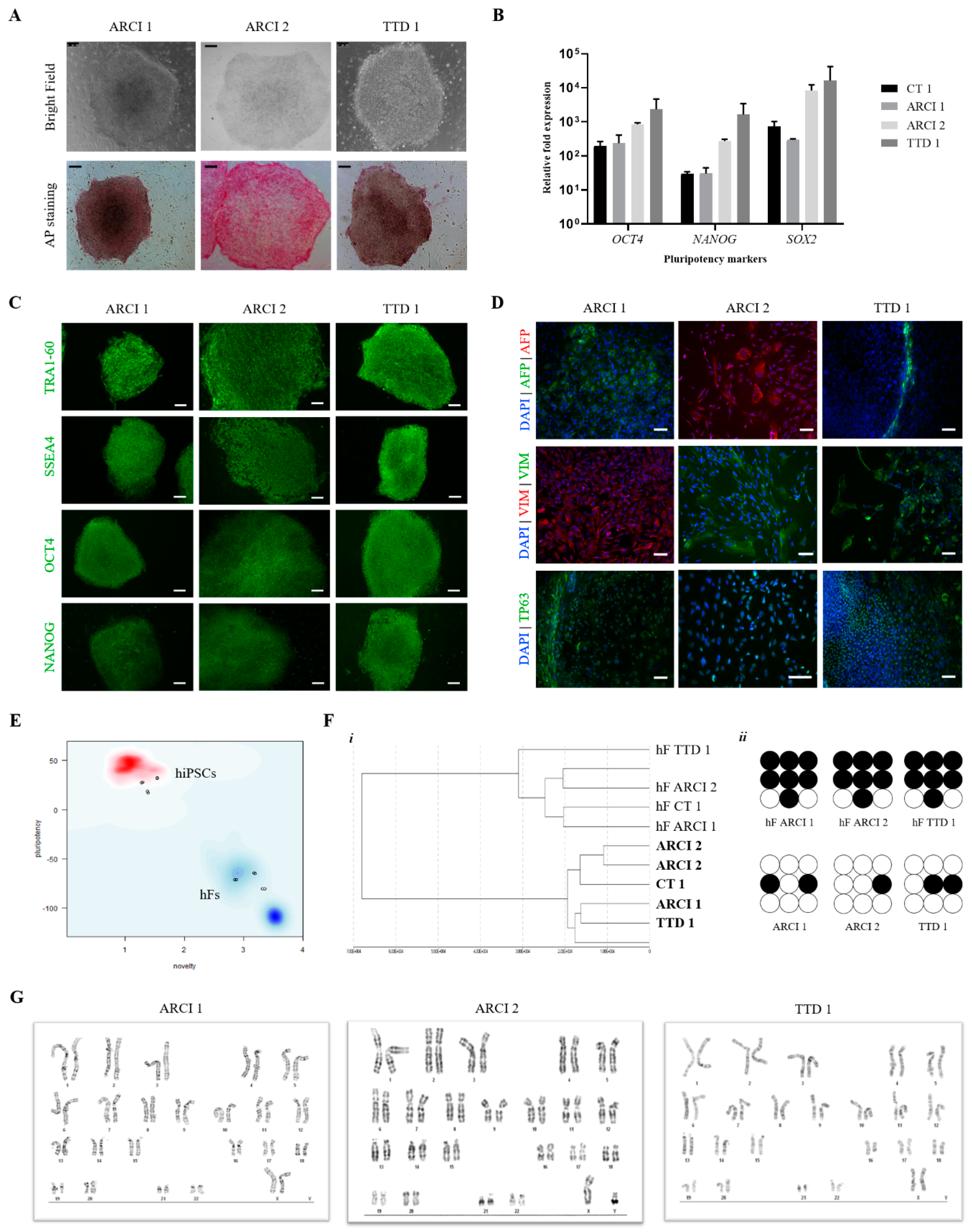
2.1. Generation of Induced Pluripotent Stem Cells (iPSCs)
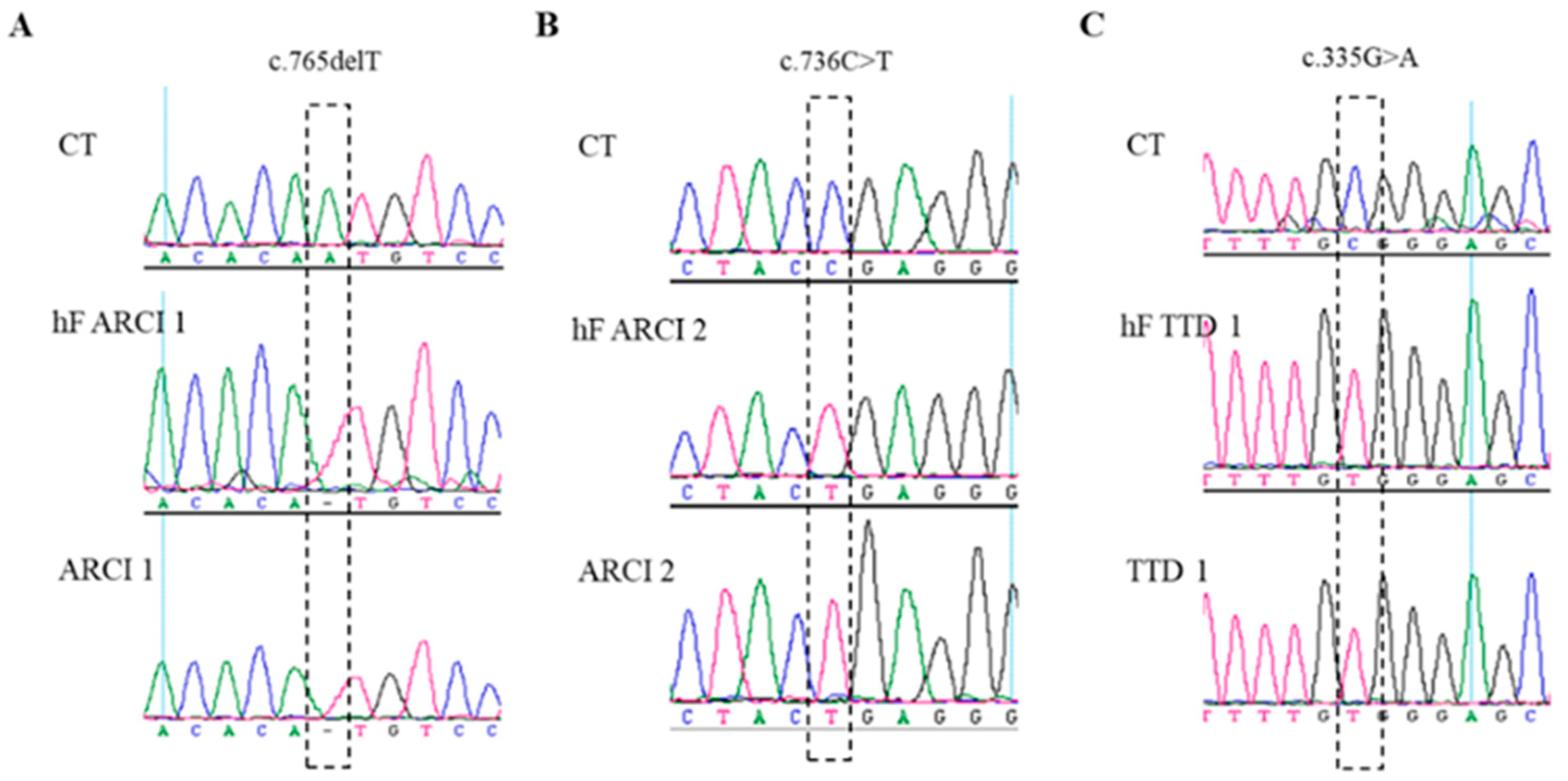
2.2. Genetic Analysis of Patient-Derived hiPSCs
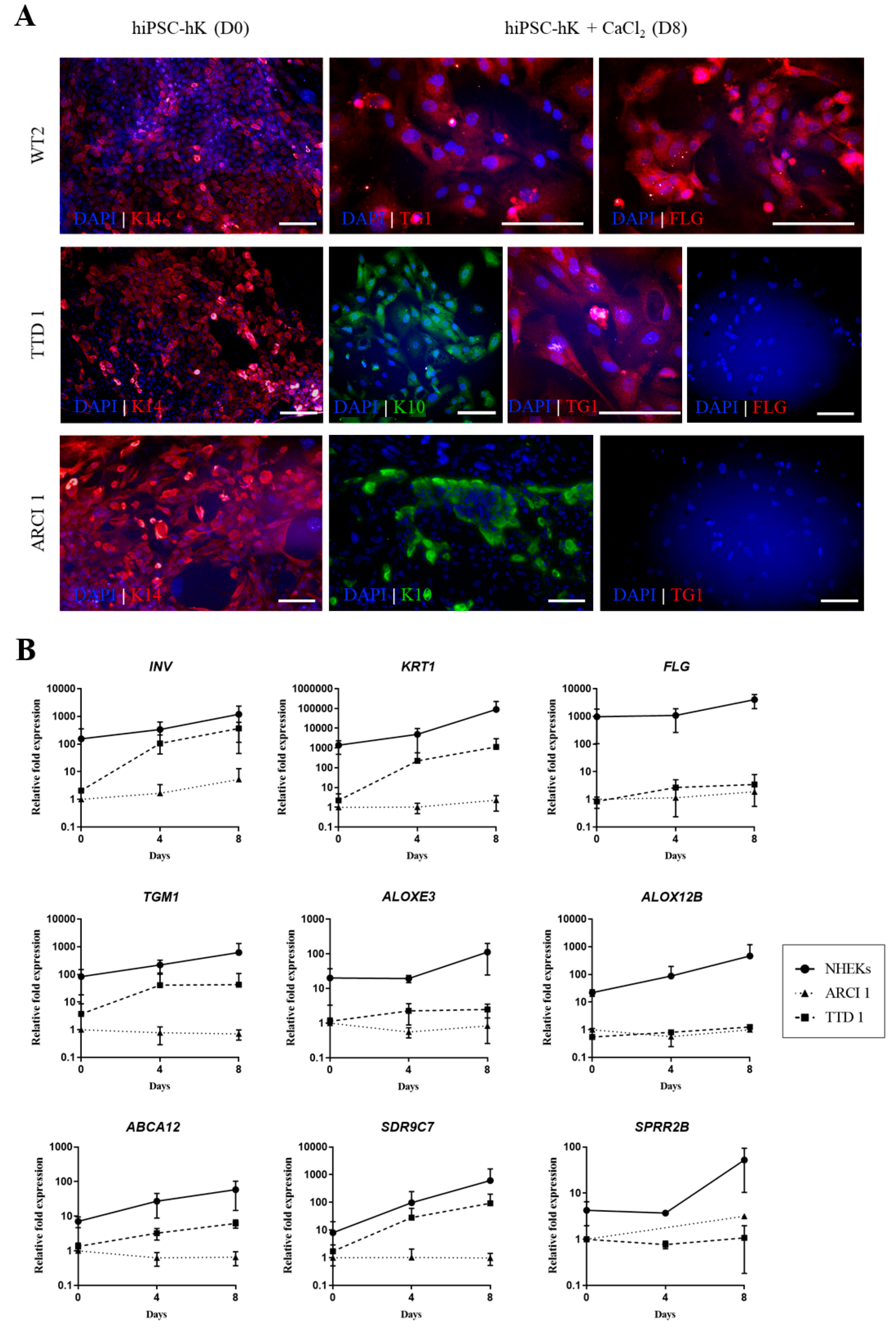
2.3. Differentiation of hiPSCs into Basal Epidermal Keratinocytes (hiPSC-bKs)
2.4. Expansion and Terminal Differentiation of hiPSC-bKs
3. Discussion
4. Materials and Methods
4.1. Ethical Approvals
4.2. Donor and Patient Description
4.3. Variant Analysis
4.4. Induced Pluripotent Stem Cells (iPSC) Generation
4.5. Cell Culturing
4.6. Differentiation of hiPSCs into Keratinocytes (iPSC-hKs)
4.7. Expansion and Terminal Differentiation of hiPSC-bKs
4.8. Alkaline Phosphatase Staining
4.9. RT-qPCR
4.10. Immunofluorescence
4.11. Karyotype Analysis
4.12. Short Tandem Repeat (STR) Analysis
4.13. Whole Gene Expression Profiling
4.14. Global DNA Methylation Array
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARCI | Autosomal recessive congenital ichthyosis |
| ATRA | All-trans retinoid acid |
| CLE | Cornified lipid envelope |
| HES | Human embryonic stem cell |
| hF | Human dermal fibroblast |
| hK | Human epidermal keratinocyte |
| hiPSC | Human induced pluripotent stem cell |
| hiPSC-bK | hiPSC-derived basal keratinocyte |
| KCM | Keratinocyte culture medium |
| LI | Lamellar ichthyosis |
| MEF | Mouse embryonic fibroblast |
| NHEK | Normal human epidermal keratinocytes |
| STR | Short tandem repeat |
| TTD | Trichothiodystrophy |
References
- Takeichi, T.; Akiyama, M. Inherited ichthyosis: Non-syndromic forms. J. Dermatol. 2016, 43, 242–251. [Google Scholar] [CrossRef]
- Yoneda, K. Inherited ichthyosis: Syndromic forms. J. Dermatol. 2016, 43, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Vahlquist, A.; Fischer, J.; Törmä, H. Inherited Nonsyndromic Ichthyoses: An Update on Pathophysiology, Diagnosis and Treatment. Am. J. Clin. Dermatol. 2018, 19, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Compe, E.; Egly, J.M. TFIIH: When transcription met DNA repair. Nat. Rev. Mol. Cell Biol. 2012, 13, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Salazar, E.P.; Broughton, B.C.; Lehmann, A.R.; Sarasin, A.; Thompson, L.H.; Weber, C.A. Defects in the DNA repair and transcription gene ERCC2(XPD) in trichothiodystrophy. Am. J. Hum. Genet. 1996, 58, 263–270. [Google Scholar] [PubMed]
- Eckl, K.M.; Krieg, P.; Küster, W.; Traupe, H.; André, F.; Wittstruck, N.; Fürstenberger, G.; Hennies, H.C. Mutation spectrum and functional analysis of epidermis-type lipoxygenases in patients with autosomal recessive congenital ichthyosis. Hum. Mutat. 2005, 26, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Eckl, K.M.; Tidhar, R.; Thiele, H.; Oji, V.; Hausser, I.; Brodesser, S.; Preil, M.L.; Onal-Akan, A.; Stock, F.; Müller, D.; et al. Impaired epidermal ceramide synthesis causes autosomal recessive congenital ichthyosis and reveals the importance of ceramide acyl chain length. J. Investig. Dermatol. 2013, 133, 2202–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichery, M.; Huchenq, A.; Sandhoff, R.; Severino-Freire, M.; Zaafouri, S.; Opálka, L.; Levade, T.; Soldan, V.; Bertrand-Michel, J.; Lhuillier, E.; et al. PNPLA1 defects in patients with autosomal recessive congenital ichthyosis and KO mice sustain PNPLA1 irreplaceable function in epidermal omega-O-acylceramide synthesis and skin permeability barrier. Hum. Mol. Genet. 2017, 26, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Loriè, E.P.; Fischer, J.; Vahlquist, A.; Törmä, H. The expression of epidermal lipoxygenases and transglutaminase-1 is perturbed by NIPAL4 mutations: Indications of a common metabolic pathway essential for skin barrier homeostasis. J. Investig. Dermatol. 2012, 132, 2368–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, Y.; Nakamichi, S.; Ohkuni, A.; Kamiyama, N.; Naoe, A.; Tsujimura, H.; Yokose, U.; Sugiura, K.; Ishikawa, J.; Akiyama, M.; et al. Essential role of the cytochrome P450 CYP4F22 in the production of acylceramide, the key lipid for skin permeability barrier formation. Proc. Natl. Acad. Sci. USA 2015, 112, 7707–7712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, P.; Fürstenberger, G. The role of lipoxygenases in epidermis. Biochim. Biophys. Acta 2014, 1841, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Rabionet, M.; Gorgas, K.; Sandhoff, R. Ceramide synthesis in the epidermis. Biochim. Biophys. Acta 2014, 1841, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Backendorf, C.; de Wit, J.; van Oosten, M.; Stout, G.J.; Mitchell, J.R.; Borgstein, A.M.; van der Horst, G.T.; de Gruijl, F.R.; Brouwer, J.; Mullenders, L.H.; et al. Repair characteristics and differentiation propensity of long-term cultures of epidermal keratinocytes derived from normal and NER-deficient mice. DNA Repair 2005, 4, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.; de Wit, J.; van Steeg, H.; Berg, R.J.; Morreau, H.; Visser, P.; Lehmann, A.R.; Duran, M.; Hoeijmakers, J.H.; Weeda, G. A mouse model for the basal transcription/DNA repair syndrome trichothiodystrophy. Mol. Cell 1998, 1, 981–990. [Google Scholar] [CrossRef]
- Stefanini, M.; Botta, E.; Lanzafame, M.; Orioli, D. Trichothiodystrophy: From basic mechanisms to clinical implications. DNA Repair 2010, 9, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Takanari, H.; Compe, E.; Egly, J.M. Dysregulation of LXR responsive genes contribute to ichthyosis in trichothiodystrophy. J. Dermatol. Sci. 2020, 97, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M. The roles of ABCA12 in epidermal lipid barrier formation and keratinocyte differentiation. Biochim. Biophys. Acta 2014, 1841, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, M.; Yamashita, F.; Ishida-Yamamoto, A.; Yamada, K.; Kinoshita, C.; Fushiki, S.; Ueda, E.; Morishima, Y.; Tabata, K.; Yasuno, H.; et al. Defective stratum corneum and early neonatal death in mice lacking the gene for transglutaminase 1 (keratinocyte transglutaminase). Proc. Natl. Acad. Sci. USA 1998, 95, 1044–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, P.; Rosenberger, S.; de Juanes, S.; Latzko, S.; Hou, J.; Dick, A.; Kloz, U.; van der Hoeven, F.; Hausser, I.; Esposito, I.; et al. Aloxe3 knockout mice reveal a function of epidermal lipoxygenase-3 as hepoxilin synthase and its pivotal role in barrier formation. J. Investig. Dermatol. 2013, 133, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grond, S.; Eichmann, T.O.; Dubrac, S.; Kolb, D.; Schmuth, M.; Fischer, J.; Crumrine, D.; Elias, P.M.; Haemmerle, G.; Zechner, R.; et al. PNPLA1 Deficiency in Mice and Humans Leads to a Defect in the Synthesis of Omega-O-Acylceramides. J. Investig. Dermatol. 2017, 137, 394–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, A.; Celli, A.; Jacquet, L.; Dafou, D.; Crumrine, D.; Hupe, M.; Arno, M.; Hobbs, C.; Cvoro, A.; Karagiannis, P.; et al. 3D In vitro model of a functional epidermal permeability barrier from human embryonic stem cells and induced pluripotent stem cells. Stem Cell Rep. 2014, 2, 675–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, M.; Kiuru, M.; Cairo, M.S.; Christiano, A.M. Generation of keratinocytes from normal and recessive dystrophic epidermolysis bullosa-induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2011, 108, 8797–8802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalom-Feuerstein, R.; Serror, L.; Aberdam, E.; Müller, F.J.; van Bokhoven, H.; Wiman, K.G.; Zhou, H.; Aberdam, D.; Petit, I. Impaired epithelial differentiation of induced pluripotent stem cells from ectodermal dysplasia-related patients is rescued by the small compound APR-246/PRIMA-1MET. Proc. Natl. Acad. Sci. USA 2013, 110, 2152–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinella, J.; Koster, M.I.; Koch, P.J. Use of induced pluripotent stem cells in dermatological research. J. Investig. Dermatol. 2014, 134, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Metallo, C.M.; Ji, L.; de Pablo, J.J.; Palecek, S.P. Retinoic acid and bone morphogenetic protein signaling synergize to efficiently direct epithelial differentiation of human embryonic stem cells. Stem Cells 2008, 26, 372–380. [Google Scholar] [CrossRef]
- Soares, E.; Xu, Q.; Li, Q.; Qu, J.; Zheng, Y.; Raeven, H.H.M.; Brandao, K.O.; Petit, I.; van den Akker, W.M.R.; van Heeringen, S.J.; et al. Single-cell RNA-seq identifies a reversible mesodermal activation in abnormally specified epithelia of p63 EEC syndrome. Proc. Natl. Acad. Sci. USA 2019, 116, 17361–17370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botta, E.; Nardo, T.; Broughton, B.C.; Marinoni, S.; Lehmann, A.R.; Stefanini, M. Analysis of mutations in the XPD gene in Italian patients with trichothiodystrophy: Site of mutation correlates with repair deficiency, but gene dosage appears to determine clinical severity. Am. J. Hum. Genet. 1998, 63, 1036–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méjécase, C.; Harding, P.; Sarkar, H.; Eintracht, J.; Lima Cunha, D.; Toualbi, L.; Moosajee, M. Generation of two human control iPS cell lines (UCLi016-A and UCLi017-A) from healthy donors with no known ocular conditions. Stem Cell Res. 2020, 49, 102113. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E. Scratching the surface of skin development. Nature 2007, 445, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, H.D.; Gassmann, M.G.; Munz, B.; Steiling, H.; Engelhardt, F.; Bleuel, K.; Werner, S. Expression and function of keratinocyte growth factor and activin in skin morphogenesis and cutaneous wound repair. J. Investig. Dermatol. Symp. Proc. 2000, 5, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Richardson, G.D.; Bazzi, H.; Fantauzzo, K.A.; Waters, J.M.; Crawford, H.; Hynd, P.; Christiano, A.M.; Jahoda, C.A. KGF and EGF signalling block hair follicle induction and promote interfollicular epidermal fate in developing mouse skin. Development 2009, 136, 2153–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.J.; Liu, Y.; Dai, Z.M.; Zhang, X.; Yang, X.; Li, Y.; Qiu, M.; Fu, J.; Hsu, W.; Chen, Y.; et al. BMP-FGF signaling axis mediates Wnt-induced epidermal stratification in developing mammalian skin. PLoS Genet. 2014, 10, e1004687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, C.; Rubin, J.; Ron, D.; Faggioni, A.; Torrisi, M.R.; Messina, A.; Frati, L.; Aaronson, S.A. Human keratinocyte growth factor activity on proliferation and differentiation of human keratinocytes: Differentiation response distinguishes KGF from EGF family. J. Cell. Physiol. 1990, 144, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.D.; Sastalla, I.; Earland, N.J.; Mahnaz, M.; Moore, I.N.; Otaizo-Carrasquero, F.; Myers, T.G.; Myles, C.A.; Datta, S.K.; Myles, I.A. Prolonging culture of primary human keratinocytes isolated from suction blisters with the Rho kinase inhibitor Y-27632. PloS ONE 2018, 13, e0198862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croze, R.H.; Buchholz, D.E.; Radeke, M.J.; Thi, W.J.; Hu, Q.; Coffey, P.J.; Clegg, D.O. ROCK Inhibition Extends Passage of Pluripotent Stem Cell-Derived Retinal Pigmented Epithelium. Stem Cells Transl. Med. 2014, 3, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qin, J.; Xie, Z.; Liu, C.; Su, Y.; Chen, Z.; Zhou, Q.; Ma, C.; Liu, G.; Paus, R.; et al. Y-27632 preserves epidermal integrity in a human skin organ-culture (hSOC) system by regulating AKT and ERK signaling pathways. J. Dermatol. Sci. 2019, 96, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.J.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, E.; Egly, J.M. Trichothiodystrophy, a transcription syndrome. Trends Genet. 2001, 17, 279–286. [Google Scholar] [CrossRef]
- Takeichi, T.; Hirabayashi, T.; Miyasaka, Y.; Kawamoto, A.; Okuno, Y.; Taguchi, S.; Tanahashi, K.; Murase, C.; Takama, H.; Tanaka, K.; et al. SDR9C7 catalyzes critical dehydrogenation of acylceramides for skin barrier formation. J. Clin. Investig. 2020, 130, 890–903. [Google Scholar] [CrossRef]
- Zhang, H.; Ericsson, M.; Weström, S.; Vahlquist, A.; Virtanen, M.; Törmä, H. Patients with congenital ichthyosis and TGM1 mutations overexpress other ARCI genes in the skin: Part of a barrier repair response? Exp. Dermatol. 2019, 28, 1164–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortmann, D.; Vallier, L. Variability of human pluripotent stem cell lines. Curr. Opin. Genet. Dev. 2017, 46, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Volpato, V.; Webber, C. Addressing variability in iPSC-derived models of human disease: Guidelines to promote reproducibility. Dis. Models Mech. 2020, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aufenvenne, K.; Larcher, F.; Hausser, I.; Duarte, B.; Oji, V.; Nikolenko, H.; Del Rio, M.; Dathe, M.; Traupe, H. Topical enzyme-replacement therapy restores transglutaminase 1 activity and corrects architecture of transglutaminase-1-deficient skin grafts. Am. J. Hum. Genet. 2013, 93, 620–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plank, R.; Yealland, G.; Miceli, E.; Lima Cunha, D.; Graff, P.; Thomforde, S.; Gruber, R.; Moosbrugger-Martinz, V.; Eckl, K.; Calderón, M.; et al. Transglutaminase 1 Replacement Therapy Successfully Mitigates the Autosomal Recessive Congenital Ichthyosis Phenotype in Full-Thickness Skin Disease Equivalents. J. Investig. Dermatol. 2019, 139, 1191–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, J.M.; Cooper, D.N.; Schuelke, M.; Seelow, D. MutationTaster2: Mutation prediction for the deep-sequencing age. Nat. Methods 2014, 11, 361–362. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Desmet, F.O.; Hamroun, D.; Lalande, M.; Collod-Béroud, G.; Claustres, M.; Béroud, C. Human Splicing Finder: An online bioinformatics tool to predict splicing signals. Nucleic Acids Res. 2009, 37, e67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiscornia, G.; Singer, O.; Verma, I.M. Production and purification of lentiviral vectors. Nat. Protoc. 2006, 1, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Berggren, W.T.; Lutz, M.; Modesto, V. General Spinfection Protocol. In StemBook; Harvard Stem Cell Institute: Cambridge, MA, USA, 2008. [Google Scholar]
- Eckl, K.M.; Alef, T.; Torres, S.; Hennies, H.C. Full-thickness human skin models for congenital ichthyosis and related keratinization disorders. J. Investig. Dermatol. 2011, 131, 1938–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, I.; Watt, F. Keratinocyte Methods; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Müller, F.-J.; Schuldt, B.M.; Williams, R.; Mason, D.; Altun, G.; Papapetrou, E.P.; Danner, S.; Goldmann, J.E.; Herbst, A.; Schmidt, N.O.; et al. A bioinformatic assay for pluripotency in human cells. Nat. Methods 2011, 8, 315–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima Cunha, D.; Oram, A.; Gruber, R.; Plank, R.; Lingenhel, A.; Gupta, M.K.; Altmüller, J.; Nürnberg, P.; Schmuth, M.; Zschocke, J.; et al. hiPSC-Derived Epidermal Keratinocytes from Ichthyosis Patients Show Altered Expression of Cornification Markers. Int. J. Mol. Sci. 2021, 22, 1785. https://doi.org/10.3390/ijms22041785
Lima Cunha D, Oram A, Gruber R, Plank R, Lingenhel A, Gupta MK, Altmüller J, Nürnberg P, Schmuth M, Zschocke J, et al. hiPSC-Derived Epidermal Keratinocytes from Ichthyosis Patients Show Altered Expression of Cornification Markers. International Journal of Molecular Sciences. 2021; 22(4):1785. https://doi.org/10.3390/ijms22041785
Chicago/Turabian StyleLima Cunha, Dulce, Amanda Oram, Robert Gruber, Roswitha Plank, Arno Lingenhel, Manoj K. Gupta, Janine Altmüller, Peter Nürnberg, Matthias Schmuth, Johannes Zschocke, and et al. 2021. "hiPSC-Derived Epidermal Keratinocytes from Ichthyosis Patients Show Altered Expression of Cornification Markers" International Journal of Molecular Sciences 22, no. 4: 1785. https://doi.org/10.3390/ijms22041785
APA StyleLima Cunha, D., Oram, A., Gruber, R., Plank, R., Lingenhel, A., Gupta, M. K., Altmüller, J., Nürnberg, P., Schmuth, M., Zschocke, J., Šarić, T., Eckl, K. M., & Hennies, H. C. (2021). hiPSC-Derived Epidermal Keratinocytes from Ichthyosis Patients Show Altered Expression of Cornification Markers. International Journal of Molecular Sciences, 22(4), 1785. https://doi.org/10.3390/ijms22041785