
The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies

 ,
, 
 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Behavioural Studies
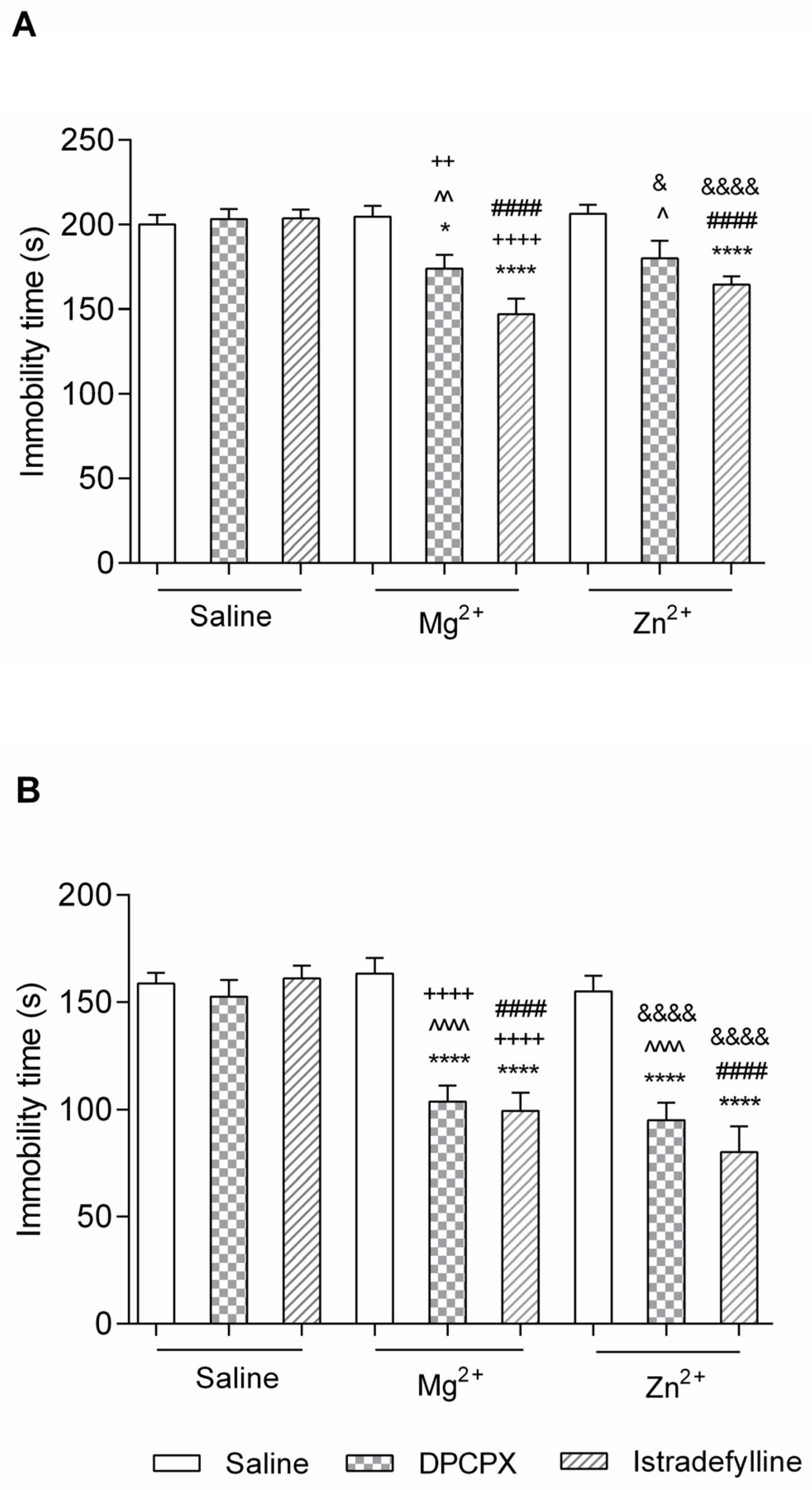
2.1.1. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc in the FST
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.1.2. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium in the TST
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.1.3. Spontaneous Locomotor Motility
2.2. Biochemical and Molecular Studies
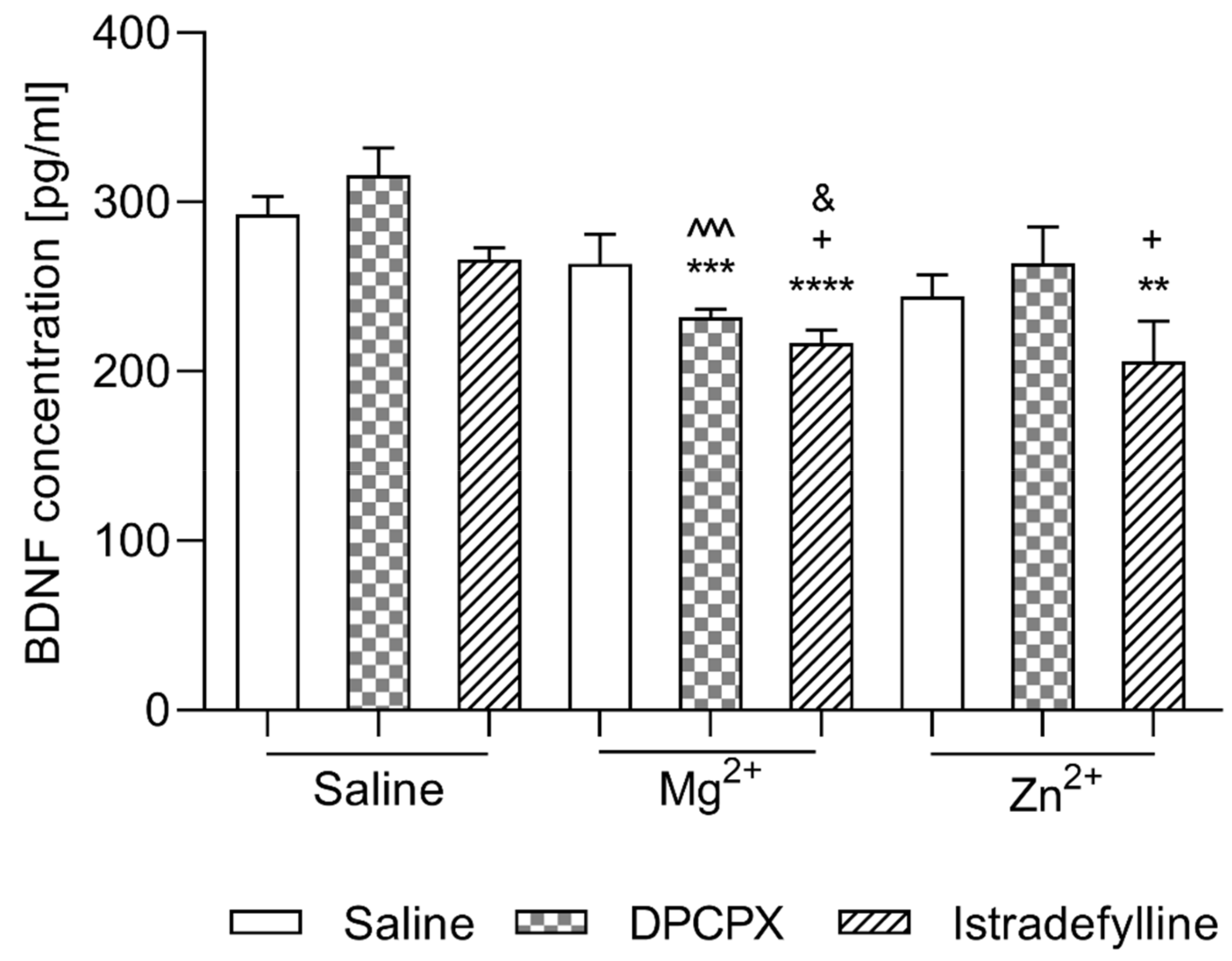
2.2.1. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on BDNF Concentration
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
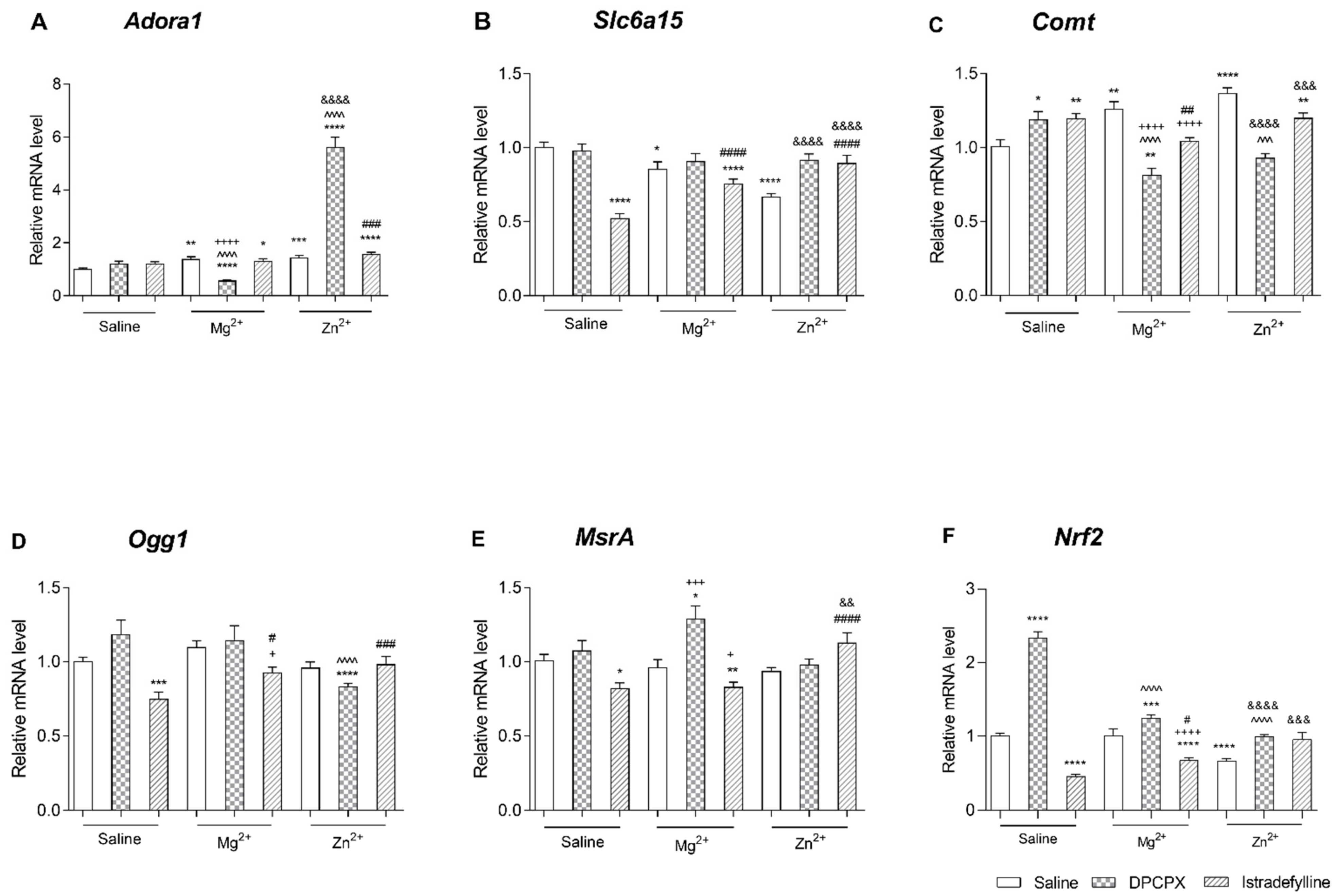
2.2.2. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Adora1 Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.2.3. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Slc6a15 Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.2.4. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Comt Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.2.5. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Ogg1 Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.2.6. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Msra Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
2.2.7. Effect of Co-Administration of Selective Adenosine Receptor Antagonists and Magnesium or Zinc on Nrf2 Expression
- (1)
- DPCPX and Magnesium or Zinc
- (2)
- Istradefylline and Magnesium or Zinc
3. Discussion
3.1. Behavioural Studies
3.2. BDNF Level Analysis
3.3. Gene Expression Analysis
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Treatment Schedule
4.4. Behavioural Studies
4.4.1. Forced Swim Test (FST)
4.4.2. Tail Suspension Test (TST)
4.4.3. Spontaneous Locomotor Motility
4.5. Biochemical and Molecular Studies
4.5.1. BDNF Levels Analysis
- (1)
- Collection of Blood
- (2)
- Determination of BDNF Concentration
4.5.2. Gene Expression Analysis
- (1)
- Collection of Prefrontal Cortex
- (2)
- RNA Isolation
- (3)
- cDNA Synthesis
- (4)
- Real-Time PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Takeda, A. Movement of zinc and its functional significance in the brain. Brain Res. Rev. 2000, 34, 137–148. [Google Scholar] [CrossRef]
- Eby, G.A.; Eby, K.L.; Murk, H. Magnesium and major depression. In Magnesium in the Central Nervous System; Vink, R., Nechifor, M., Eds.; University of Adelaide Press: Adelaide, Australia, 2011; pp. 313–330. [Google Scholar]
- Sandstead, H.H. Subclinical zinc deficiency impairs human brain function. J. Trace Elem Med. Biol. 2012, 26, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, K.; Klenk-Majewska, B.; Danilczuk, Z.; Wróbel, A.; Łupina, T.; Ossowska, G. Influence of zinc supplementation on imipramine effect in a chronic unpredictable stress (CUS) model in rats. Pharmacol. Rep. 2007, 59, 46–52. [Google Scholar] [PubMed]
- Cunha, M.P.; Machado, D.G.; Bettio, L.E.; Capra, J.C.; Rodrigues, A.L. Interaction of zinc with antidepressants in the tail suspension test. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Młyniec, K. Zinc in the glutamatergic theory of depression. Curr. Neuropharmacol. 2015, 13, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Salari, S.; Khomand, P.; Arasteh, M.; Yousefzamani, B.; Hassanzadeh, K. Zinc sulphate: A reasonable choice for depression management in patients with multiple sclerosis: A randomized, double-blind, placebo-controlled clinical trial. Pharmacol. Rep. 2015, 67, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B.; Pochwat, B.; Rafało, A.; Palucha-Poniewiera, A.; Domin, H.; Nowak, G. Activation of mTOR dependent signaling pathway is a necessary mechanism of antidepressant-like activity of zinc. Neuropharmacology 2015, 99, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Poleszak, E.; Wlaź, P.; Kędzierska, E.; Nieoczym, D.; Wyska, E.; Szymura-Oleksiak, J.; Fidecka, S.; Radziwoń-Zaleska, M.; Nowak, G. Immobility stress induces depression-like behavior in the forced swim test in mice: Effect of magnesium and imipramine. Pharmacol. Rep. 2006, 58, 746–752. [Google Scholar]
- Poleszak, E.; Wlaź, P.; Szewczyk, B.; Kędzierska, E.; Wyska, E.; Librowski, T.; Szymura-Oleksiak, J.; Fidecka, S.; Pilc, A.; Nowak, G. Enhancement of antidepressant-like activity by joint administration of imipramine and magnesium in the forced swim test: Behavioral and pharmacokinetic studies in mice. Pharmacol. Biochem. Behav. 2005, 81, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Whittle, N.; Lubec, G.; Singewald, N. Zinc deficiency induces enhanced depression-like behaviour and altered limbic activation reversed by antidepressant treatment in mice. Amino Acids 2009, 36, 147–158. [Google Scholar] [CrossRef]
- Szewczyk, B.; Poleszak, E.; Wlaź, P.; Wróbel, A.; Blicharska, E.; Cichy, A.; Dybała, M.; Siwek, A.; Pomierny-Chamioło, L.; Piotrowska, A.; et al. The involvement of serotonergic system in the antidepressant effect of zinc in the forced swim test. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Besser, L.; Chorin, E.; Sekler, I.; Silverman, W.F.; Atkin, S.; Russell, J.T.; Hershfinkel, M. Synaptically released zinc triggers metabotropic signaling via a zinc-sensing receptor in the hippocampus. J. Neurosci. 2009, 29, 2890–2901. [Google Scholar] [CrossRef] [Green Version]
- Poleszak, E.; Wlaź, P.; Kędzierska, E.; Nieoczym, D.; Wróbel, A.; Fidecka, S.; Pilc, A.; Nowak, G. NMDA/glutamate mechanism of antidepressant-like action of magnesium in forced swim test in mice. Pharmacol. Biochem. Behav. 2007, 88, 158–164. [Google Scholar] [CrossRef]
- Doboszewska, U.; Wlaź, P.; Nowak, G.; Radziwoń-Zaleska, M.; Cui, R.; Młyniec, K. Zinc in the monoaminergic theory of depression: Its relationship to neural plasticity. Neural Plast. 2017, 2017, 3682752. [Google Scholar] [CrossRef]
- Paoletti, P.; Vergnano, A.M.; Barbour, B.; Casado, M. Zinc at glutamatergic synapses. Neuroscience 2009, 158, 126–136. [Google Scholar] [CrossRef]
- Szewczyk, B.; Kubera, M.; Nowak, G. The role of zinc in neurodegenerative inflammatory pathways in depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 693–701. [Google Scholar] [CrossRef]
- Fuxe, K.; Dahlström, A.B.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Linden, J.; Müller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Fuxe, K. Adenosine heteroreceptor complexes in the basal ganglia are implicated in Parkinson’s disease and its treatment. J. Neural Transm. 2019, 126, 455–471. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.V.; Kaster, M.P.; Tomé, A.R.; Agostinho, P.M.; Cunha, R.A. Adenosine receptors and brain diseases: Neuroprotection and neurodegeneration. Biochim. Biophys. Acta 2011, 1808, 1380–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciruela, F.; Casadó, V.; Rodrigues, R.J.; Luján, R.; Burgueño, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef] [Green Version]
- Fredholm, B.B.; Chen, J.F.; Cunha, R.A.; Svenningsson, P.; Vaugeois, J.M. Adenosine and brain function. Int. Rev. Neurobiol. 2005, 63, 191–270. [Google Scholar]
- Wardas, J. Neuroprotective role of adenosine in the CNS. Pol. J. Pharmacol. 2002, 54, 313–326. [Google Scholar] [PubMed]
- Brand, A.; Vissiennon, Z.; Eschke, D.; Nieber, K. Adenosine A(1) and A(3) receptors mediate inhibition of synaptic transmission in rat cortical neurons. Neuropharmacology 2001, 40, 85–95. [Google Scholar] [CrossRef]
- Scammell, T.E.; Arrigoni, E.; Thompson, M.A.; Ronan, P.J.; Saper, C.B.; Greene, R.W. Focal deletion of the adenosine A1 receptor in adult mice using an adeno-associated viral vector. J. Neurosci. 2003, 23, 5762–5770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotanus, S.M.; Fredholm, B.B.; Chergui, K. NMDA depresses glutamatergic synaptic transmission in the striatum through the activation of adenosine A1 receptors: Evidence from knockout mice. Neuropharmacology 2006, 51, 272–282. [Google Scholar] [CrossRef]
- Franco, R. Neurotransmitter receptor heteromers in neurodegenerative diseases and neural plasticity. J. Neural Transm. 2009, 116, 983–987. [Google Scholar] [CrossRef]
- Szopa, A.; Poleszak, E.; Wyska, E.; Serefko, A.; Wośko, S.; Wlaź, A.; Pieróg, M.; Wróbel, A.; Wlaź, P. Caffeine enhances the antidepressant-like activity of common antidepressant drugs in the forced swim test in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2016, 389, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serefko, A.; Szopa, A.; Wlaź, A.; Wośko, S.; Wlaź, P.; Poleszak, E. Synergistic antidepressant-like effect of the joint administration of caffeine and NMDA receptor ligands in the forced swim test in mice. J. Neural Transm. 2016, 123, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Poleszak, E.; Bogatko, K.; Wyska, E.; Wośko, S.; Doboszewska, U.; Świąder, K.; Wlaź, A.; Dudka, J.; Wróbel, A.; et al. DPCPX, a selective adenosine A1 receptor antagonist, enhances the antidepressant-like effects of imipramine, escitalopram, and reboxetine in mice behavioral tests. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 1361–1371. [Google Scholar] [CrossRef] [Green Version]
- Poleszak, E.; Szopa, A.; Bogatko, K.; Wyska, E.; Wośko, S.; Świąder, K.; Doboszewska, U.; Wlaź, A.; Wróbel, A.; Wlaź, P.; et al. Antidepressant-like activity of typical antidepressant drugs in the forced swim test and tail suspension test in mice is augmented by DMPX, an adenosine A(2A) receptor antagonist. Neurotox. Res. 2019, 35, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Herbet, M.; Szopa, A.; Serefko, A.; Wośko, S.; Gawrońska-Grzywacz, M.; Izdebska, M.; Piątkowska-Chmiel, I.; Betiuk, P.; Poleszak, E.; Dudka, J. 8-Cyclopentyl-1,3-dimethylxanthine enhances effectiveness of antidepressant in behavioral tests and modulates redox balance in the cerebral cortex of mice. Saudi Pharm. J. 2018, 26, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef] [PubMed]
- Petit-Demouliere, B.; Chenu, F.; Bourin, M. Forced swimming test in mice: A review of antidepressant activity. Psychopharmacology 2005, 177, 245–255. [Google Scholar] [CrossRef]
- Bai, F.; Li, X.; Clay, M.; Lindstrom, T.; Skolnick, P. Intra- and interstrain differences in models of “behavioral despair”. Pharmacol. Biochem. Behav. 2001, 70, 187–192. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Lenegre, A. Behavioral models of depression. In Experimental Approaches to Anxiety and Depression; Elliot, J.M., Heal, D.J., Marsden, C.A., Eds.; John Wiley & Sons: London, UK, 1992; pp. 73–85. [Google Scholar]
- Dalvi, A.; Lucki, I. Murine models of depression. Psychopharmacology 1999, 147, 14–16. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Maj, J.; Rogóż, Z.; Skuza, G.; Sowińska, H. Effects of MK-801 and antidepressant drugs in the forced swimming test in rats. Eur. Neuropsychopharmacol. 1992, 2, 37–41. [Google Scholar] [CrossRef]
- Trullas, R.; Skolnick, P. Functional antagonists at the NMDA receptor complex exhibit antidepressant actions. Eur. J. Pharmacol 1990, 185, 1–10. [Google Scholar] [CrossRef]
- Li, X.; Tizzano, J.P.; Griffey, K.; Clay, M.; Lindstrom, T.; Skolnick, P. Antidepressant-like actions of an AMPA receptor potentiator (LY392098). Neuropharmacology 2001, 40, 1028–1033. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Chen, J.F.; Masino, S.A.; Vaugeois, J.M. Actions of adenosine at its receptors in the CNS: Insights from knockouts and drugs. Annu Rev. Pharmacol. Toxicol. 2005, 45, 385–412. [Google Scholar] [CrossRef] [PubMed]
- Górska, A.M.; Gołembiowska, K. The role of adenosine A1 and A2A receptors in the caffeine effect on MDMA-induced DA and 5-HT release in the mouse striatum. Neurotox. Res. 2015, 27, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Sperlágh, B.; Vizi, E.S. The role of extracellular adenosine in chemical neurotransmission in the hippocampus and Basal Ganglia: Pharmacological and clinical aspects. Curr. Top. Med. Chem. 2011, 11, 1034–1046. [Google Scholar] [CrossRef]
- Blum, D.; Galas, M.C.; Pintor, A.; Brouillet, E.; Ledent, C.; Müller, C.E.; Bantubungi, K.; Galluzzo, M.; Gall, D.; Cuvelier, L.; et al. A dual role of adenosine A2A receptors in 3-nitropropionic acid-induced striatal lesions: Implications for the neuroprotective potential of A2A antagonists. J. Neurosci. 2003, 23, 5361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsi, C.; Pinna, A.; Gianfriddo, M.; Melani, A.; Morelli, M.; Pedata, F. Adenosine A2A receptor antagonism increases striatal glutamate outflow in dopamine-denervated rats. Eur. J. Pharmacol. 2003, 464, 33–38. [Google Scholar] [CrossRef]
- Karcz-Kubicha, M.; Quarta, D.; Hope, B.T.; Antoniou, K.; Müller, C.E.; Morales, M.; Schindler, C.W.; Goldberg, S.R.; Ferré, S. Enabling role of adenosine A1 receptors in adenosine A2A receptor-mediated striatal expression of c-fos. Eur. J. Neurosci. 2003, 18, 296–302. [Google Scholar] [CrossRef]
- Pintor, A.; Quarta, D.; Pezzola, A.; Reggio, R.; Popoli, P. SCH 58261 (an adenosine A(2A) receptor antagonist) reduces, only at low doses, K(+)-evoked glutamate release in the striatum. Eur. J. Pharmacol 2001, 421, 177–180. [Google Scholar] [CrossRef]
- Quarta, D.; Ferré, S.; Solinas, M.; You, Z.B.; Hockemeyer, J.; Popoli, P.; Goldberg, S.R. Opposite modulatory roles for adenosine A1 and A2A receptors on glutamate and dopamine release in the shell of the nucleus accumbens. Effects of chronic caffeine exposure. J. Neurochem 2004, 88, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Solinas, M.; Ferré, S.; You, Z.B.; Karcz-Kubicha, M.; Popoli, P.; Goldberg, S.R. Caffeine induces dopamine and glutamate release in the shell of the nucleus accumbens. J. Neurosci. 2002, 22, 6321–6324. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Hinz, S.; Navarro, G.; Franco, R.; MĂĽller, C.E.; Fuxe, K. Understanding the Role of Adenosine A2AR Heteroreceptor Complexes in Neurodegeneration and Neuroinflammation. Front. Neurosci. 2018, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, A.; Dravolina, O.; Belozertseva, I.; Adamcio, B.; Zvartau, E. Lowered brain stimulation reward thresholds in rats treated with a combination of caffeine and N-methyl-D-aspartate but not alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate or metabotropic glutamate receptor-5 receptor antagonists. Behav. Pharmacol. 2006, 17, 295–302. [Google Scholar] [CrossRef]
- de Oliveira, R.V.; Dall’Igna, O.P.; Tort, A.B.; Schuh, J.F.; Neto, P.F.; Santos Gomes, M.W.; Souza, D.O.; Lara, D.R. Effect of subchronic caffeine treatment on MK-801-induced changes in locomotion, cognition and ataxia in mice. Behav. Pharmacol. 2005, 16, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Lobato, K.R.; Binfaré, R.W.; Budni, J.; Rosa, A.O.; Santos, A.R.; Rodrigues, A.L. Involvement of the adenosine A1 and A2A receptors in the antidepressant-like effect of zinc in the forced swimming test. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 994–999. [Google Scholar] [CrossRef]
- Kreiner, G.; Chmielarz, P.; Roman, A.; Nalepa, I. Gender differences in genetic mouse models evaluated for depressive-like and antidepressant behavior. Pharmacol. Rep. 2013, 65, 1580–1590. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, J.M.; Su, W.J.; Wang, B.; Jiang, C.L. Sex differences in depressive-like behaviour may relate to imbalance of microglia activation in the hippocampus. Brain Behav. Immun. 2019, 81, 188–197. [Google Scholar] [CrossRef] [PubMed]
- McHenry, J.; Carrier, N.; Hull, E.; Kabbaj, M. Sex differences in anxiety and depression: Role of testosterone. Front. Neuroendocrinol. 2014, 35, 42–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaster, M.P.; Rosa, A.O.; Rosso, M.M.; Goulart, E.C.; Santos, A.R.; Rodrigues, A.L. Adenosine administration produces an antidepressant-like effect in mice: Evidence for the involvement of A1 and A2A receptors. Neurosci. Lett. 2004, 355, 21–24. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: A primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar]
- Browne, R.G.; Welch, W.M. Stereoselective antagonism of phencyclidine’s discriminative properties by adenosine receptor agonists. Science 1982, 217, 1157–1159. [Google Scholar] [CrossRef]
- Popoli, P.; Reggio, R.; Pezzola, A. Adenosine A1 and A2 receptor agonists significantly prevent the electroencephalographic effects induced by MK-801 in rats. Eur. J. Pharmacol. 1997, 333, 143–146. [Google Scholar] [CrossRef]
- Craig, C.G.; White, T.D. N-methyl-D-aspartate- and non-N-methyl-D-aspartate-evoked adenosine release from rat cortical slices: Distinct purinergic sources and mechanisms of release. J. Neurochem. 1993, 60, 1073–1080. [Google Scholar] [CrossRef]
- Manzoni, O.J.; Manabe, T.; Nicoll, R.A. Release of adenosine by activation of NMDA receptors in the hippocampus. Science 1994, 265, 2098–2101. [Google Scholar] [CrossRef] [PubMed]
- Melani, A.; Corsi, C.; Giménez-Llort, L.; Martínez, E.; Ogren, S.O.; Pedata, F.; Ferré, S. Effect of N-methyl-D-aspartate on motor activity and in vivo adenosine striatal outflow in the rat. Eur. J. Pharmacol. 1999, 385, 15–19. [Google Scholar] [CrossRef]
- Sebastiao, A.M.; Ribeiro, J.A. Fine-tuning neuromodulation by adenosine. Trends Pharmacol. Sci. 2000, 21, 341–346. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Z.Y. The role of BDNF in depression on the basis of its location in the neural circuitry. Acta Pharmacol. Sin. 2011, 32, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Malberg, J.; Nakagawa, S.; D’Sa, C. Neuronal plasticity and survival in mood disorders. Biol. Psychiatry 2000, 48, 732–739. [Google Scholar] [CrossRef]
- Altar, C.A. Neurotrophins and depression. Trends Pharmacol. Sci. 1999, 20, 59–61. [Google Scholar] [CrossRef]
- Orzelska-Górka, J.; Szewczyk, K.; Gawrońska-Grzywacz, M.; Kędzierska, E.; Głowacka, E.; Herbet, M.; Dudka, J.; Biała, G. Monoaminergic system is implicated in the antidepressant-like effect of hyperoside and protocatechuic acid isolated from Impatiens glandulifera Royle in mice. Neurochem. Int. 2019, 128, 206–214. [Google Scholar] [CrossRef]
- Kozisek, M.E.; Middlemas, D.; Bylund, D.B. Brain-derived neurotrophic factor and its receptor tropomyosin-related kinase B in the mechanism of action of antidepressant therapies. Pharmacol. Ther. 2008, 117, 30–51. [Google Scholar] [CrossRef]
- Hennigan, A.; Callaghan, C.K.; Kealy, J.; Rouine, J.; Kelly, A.M. Deficits in LTP and recognition memory in the genetically hypertensive rat are associated with decreased expression of neurotrophic factors and their receptors in the dentate gyrus. Behav. Brain Res. 2009, 197, 371–377. [Google Scholar] [CrossRef]
- Paizanis, E.; Kelai, S.; Renoir, T.; Hamon, M.; Lanfumey, L. Life-long hippocampal neurogenesis: Environmental, pharmacological and neurochemical modulations. Neurochem. Res. 2007, 32, 1762–1771. [Google Scholar] [CrossRef]
- Phillips, C. Brain-derived neurotrophic factor, depression, and physical activity: Making the neuroplastic connection. Neural Plast. 2017, 2017, 7260130. [Google Scholar] [CrossRef]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2011, 14, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartorius, A.; Hellweg, R.; Litzke, J.; Vogt, M.; Dormann, C.; Vollmayr, B.; Danker-Hopfe, H.; Gass, P. Correlations and discrepancies between serum and brain tissue levels of neurotrophins after electroconvulsive treatment in rats. Pharmacopsychiatry 2009, 42, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Gejl, A.K.; Enevold, C.; Bugge, A.; Andersen, M.S.; Nielsen, C.H.; Andersen, L.B. Associations between serum and plasma brain-derived neurotrophic factor and influence of storage time and centrifugation strategy. Sci. Rep. 2019, 9, 9655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karege, F.; Schwald, M.; Cisse, M. Postnatal developmental profile of brain-derived neurotrophic factor in rat brain and platelets. Neurosci. Lett. 2002, 328, 261–264. [Google Scholar] [CrossRef]
- Russo-Neustadt, A.; Beard, R.C.; Cotman, C.W. Exercise, antidepressant medications, and enhanced brain derived neurotrophic factor expression. Neuropsychopharmacology 1999, 21, 679–682. [Google Scholar] [CrossRef] [Green Version]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.H.; Lee, I.H.; Chi, M.H.; Lin, S.H.; Chen, K.C.; Chen, P.S.; Chiu, N.T.; Yao, W.J.; Yang, Y.K. Prefrontal cortex modulates the correlations between brain-derived neurotrophic factor level, serotonin, and the autonomic nervous system. Sci. Rep. 2018, 8, 2558. [Google Scholar] [CrossRef] [Green Version]
- Serchov, T.; Clement, H.W.; Schwarz, M.K.; Iasevoli, F.; Tosh, D.K.; Idzko, M.; Jacobson, K.A.; de Bartolomeis, A.; Normann, C.; Biber, K.; et al. Increased signaling via adenosine A1 receptors, sleep deprivation, imipramine, and ketamine inhibit depressive-like behavior via induction of Homer1a. Neuron 2015, 87, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Han, K.M.; Kang, J.; Won, E.; Chang, H.S.; Tae, W.S.; Son, K.R.; Kim, S.J.; Lee, M.S.; Ham, B.J. Effects of a polymorphism of the neuronal amino acid transporter SLC6A15 gene on structural integrity of white matter tracts in major depressive disorder. PLoS ONE 2016, 11, e0164301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhofer, G.; Kopin, I.J.; Goldstein, D.S. Catecholamine metabolism: A contemporary view with implications for physiology and medicine. Pharmacol. Rev. 2004, 56, 331–349. [Google Scholar] [CrossRef]
- Coelho, J.E.; de Mendonça, A.; Ribeiro, J.A. Presynaptic inhibitory receptors mediate the depression of synaptic transmission upon hypoxia in rat hippocampal slices. Brain Res. 2000, 869, 158–165. [Google Scholar] [CrossRef]
- Stone, T.W.; Newby, A.C.; Lloyd, H.G.E. Adenosine release. In Adenosine and Adenosine Receptors; Williams, M., Ed.; The Humana Press: Totowa, NJ, USA, 1990; pp. 173–223. [Google Scholar]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Bertorelli, R.; Ongini, E.; Costentin, J.; Vaugeois, J.M. Adenosine A2A receptor antagonists are potential antidepressants: Evidence based on pharmacology and A2A receptor knockout mice. Br. J. Pharmacol. 2001, 134, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, M.A.; Lucae, S.; Saemann, P.G.; Schmidt, M.V.; Demirkan, A.; Hek, K.; Czamara, D.; Alexander, M.; Salyakina, D.; Ripke, S.; et al. The neuronal transporter gene SLC6A15 confers risk to major depression. Neuron 2011, 70, 252–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarelli, S.; Wagner, K.V.; Labermaier, C.; Uribe, A.; Dournes, C.; Balsevich, G.; Hartmann, J.; Masana, M.; Holsboer, F.; Chen, A.; et al. SLC6A15, a novel stress vulnerability candidate, modulates anxiety and depressive-like behavior: Involvement of the glutamatergic system. Stress 2016, 19, 83–90. [Google Scholar] [CrossRef]
- Gogos, J.A.; Morgan, M.; Luine, V.; Santha, M.; Ogawa, S.; Pfaff, D.; Karayiorgou, M. Catechol-O-methyltransferase-deficient mice exhibit sexually dimorphic changes in catecholamine levels and behavior. Proc. Natl. Acad. Sci. USA 1998, 95, 9991–9996. [Google Scholar] [CrossRef] [Green Version]
- Antypa, N.; Drago, A.; Serretti, A. The role of COMT gene variants in depression: Bridging neuropsychological, behavioral and clinical phenotypes. Neurosci. BioBehav. Rev. 2013, 37, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Fukae, J.; Takanashi, M.; Kubo, S.; Nishioka, K.; Nakabeppu, Y.; Mori, H.; Mizuno, Y.; Hattori, N. Expression of 8-oxoguanine DNA glycosylase (OGG1) in Parkinson’s disease and related neurodegenerative disorders. Acta Neuropathol. 2005, 109, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Vaváková, M.; Ďuračková, Z.; Trebatická, J. Markers of oxidative stress and neuroprogression in depression disorder. Oxid. Med. Cell. Longev. 2015, 2015, 898393. [Google Scholar] [CrossRef] [Green Version]
- Ceylan, D.; Yilmaz, S.; Tuna, G.; Kant, M.; Er, A.; Ildi, A.; Verim, B.; Akis, M.; Akan, P.; Islekel, H.; et al. Alterations in levels of 8-Oxo-2′-deoxyguanosine and 8-Oxoguanine DNA glycosylase 1 during a current episode and after remission in unipolar and bipolar depression. Psychoneuroendocrinology 2020, 114, 104600. [Google Scholar] [CrossRef]
- Szebeni, A.; Szebeni, K.; DiPeri, T.P.; Johnson, L.A.; Stockmeier, C.A.; Crawford, J.D.; Chandley, M.J.; Hernandez, L.J.; Burgess, K.C.; Brown, R.W.; et al. Elevated DNA oxidation and DNA repair enzyme expression in brain white matter in major depressive disorder. Int. J. Neuropsychopharmacol. 2017, 20, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskovitz, J.; Bar-Noy, S.; Williams, W.M.; Requena, J.; Berlett, B.S.; Stadtman, E.R. Methionine sulfoxide reductase (MsrA) is a regulator of antioxidant defense and lifespan in mammals. Proc. Natl. Acad. Sci. USA 2001, 98, 12920–12925. [Google Scholar] [CrossRef] [Green Version]
- Moskovitz, J.; Flescher, E.; Berlett, B.S.; Azare, J.; Poston, J.M.; Stadtman, E.R. Overexpression of peptide-methionine sulfoxide reductase in Saccharomyces cerevisiae and human T cells provides them with high resistance to oxidative stress. Proc. Natl. Acad. Sci. USA 1998, 95, 14071–14075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomobe, K.; Shinozuka, T.; Kuroiwa, M.; Nomura, Y. Age-related changes of Nrf2 and phosphorylated GSK-3b˛ in a mouse model of accelerated aging (SAMP8). Arch. Gerontol. Geriatr. 2012, 54, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Qu, L.Y.; Quan, K.; Jiang, Y.L.; Tang, X.W. Effect of tBHQ and sulforaphane on Nrf2-ARE signaling pathway of Caco2 cells. Zhejiang Da Xue Xue Bao Yi Xue Ban 2010, 39, 17–23. [Google Scholar] [PubMed]
- Haskew-Layton, R.E.; Payappilly, J.B.; Smirnova, N.A.; Ma, T.C.; Chan, K.K.; Murphy, T.H.; Guo, H.; Langley, B.; Sultana, R.; Butterfield, D.A.; et al. Controlled enzymatic production of astrocytic hydrogen peroxide protects neurons from oxidative stress via an Nrf2-independent pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 17385–17390. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Kobayashi, M.; Mori, A.; Jenner, P.; Kanda, T. Antidepressant-like activity of the adenosine A(2A) receptor antagonist, istradefylline (KW-6002), in the forced swim test and the tail suspension test in rodents. Pharmacol. Biochem. Behav. 2013, 114–115, 23–30. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment (mg/kg) | Distance (cm) between the 2nd and the 6th Minute |
|---|---|
| saline + saline | 1196.5 ± 58.3 |
| DPCPX 1 + saline | 1111.8 ± 57.1 |
| istradefylline 0.5 + saline | 1093.2 ± 54.7 |
| Mg2+ 10 + saline | 1028.9 ± 73.2 |
| DPCPX 1 + Mg2+ 10 | 905.40 ± 93.5 |
| istradefylline 0.5 + Mg2+ 10 | 1150.9 ± 37.3 |
| Zn2+ 2.5 + saline | 986.90 ± 38.2 * |
| DPCPX 1 + Zn2+ 2.5 | 766.40 ± 89.7 ***,^^^,& |
| istradefylline 0.5 + Zn2+ 2.5 | 1186.8 ± 90.9 |
| Gene Symbol | Gene Name | Ref. Seq | Assay ID | Amplicon Length |
|---|---|---|---|---|
| Ogg1 | 8–Oxoguanine DNA–glycosylase 1 | NM_010957.4 | Mm00501784_m1 | 90 |
| Msra | Methionine sulfoxide reductase A | NM_001253712.1 NM_001253714.1 NM_001253716.1 NM_026322.4 | Mm00452737_m1 | 69 |
| Nfe2l2 | Nuclear factor, erythroid derived 2, like 2 | NM_010902.3 | Mm00477784_m1 | 61 |
| Adora1 | Adenosine A1 receptor | NM_001008533.3 NM_001039510.2 NM_001282945.1 | Mm01308023_m1 | 58 |
| Slc6a15 | Solute carrier family 6 (neurotransmitter transporter), member 15 | NM_001252330.1 NM_175328.3 | Mm00558415_m1 | 84 |
| Comt | Catechol–O–methyl– transferase | NM_001111062.1 NM_001111063.1 NM_007744.3 | Mm00514377_m1 | 97 |
| Hprt | Hypoxanthine guanine phosphoribosyl transferase | NM_013556.2 | Mm00446968_m1 | 65 |
| Tbp | TATA box binding protein | NM_013684.3 | Mm00446974_m1 | 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szopa, A.; Bogatko, K.; Herbet, M.; Serefko, A.; Ostrowska, M.; Wośko, S.; Świąder, K.; Szewczyk, B.; Wlaź, A.; Skałecki, P.; et al. The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies. Int. J. Mol. Sci. 2021, 22, 1840. https://doi.org/10.3390/ijms22041840
Szopa A, Bogatko K, Herbet M, Serefko A, Ostrowska M, Wośko S, Świąder K, Szewczyk B, Wlaź A, Skałecki P, et al. The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies. International Journal of Molecular Sciences. 2021; 22(4):1840. https://doi.org/10.3390/ijms22041840
Chicago/Turabian StyleSzopa, Aleksandra, Karolina Bogatko, Mariola Herbet, Anna Serefko, Marta Ostrowska, Sylwia Wośko, Katarzyna Świąder, Bernadeta Szewczyk, Aleksandra Wlaź, Piotr Skałecki, and et al. 2021. "The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies" International Journal of Molecular Sciences 22, no. 4: 1840. https://doi.org/10.3390/ijms22041840
APA StyleSzopa, A., Bogatko, K., Herbet, M., Serefko, A., Ostrowska, M., Wośko, S., Świąder, K., Szewczyk, B., Wlaź, A., Skałecki, P., Wróbel, A., Mandziuk, S., Pochodyła, A., Kudela, A., Dudka, J., Radziwoń-Zaleska, M., Wlaź, P., & Poleszak, E. (2021). The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies. International Journal of Molecular Sciences, 22(4), 1840. https://doi.org/10.3390/ijms22041840