
Potassium Channels Kv1.3 and Kir2.1 But Not Kv1.5 Contribute to BV2 Cell Line and Primary Microglial Migration

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
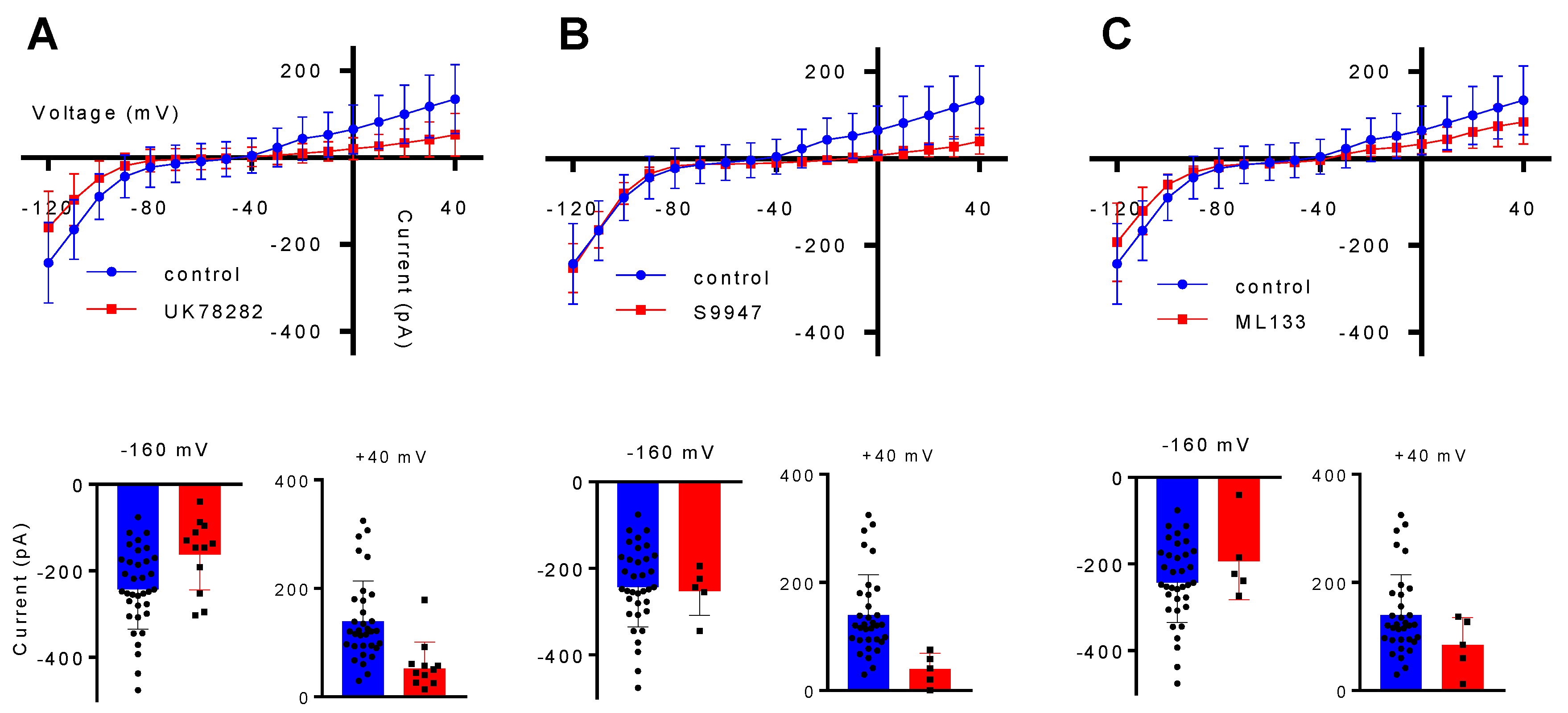
2.1. The Characterization of Potassium Channels Kv1.3, Kv1.5 and Kir 2.1 in the BV2 Microglial Cell Line
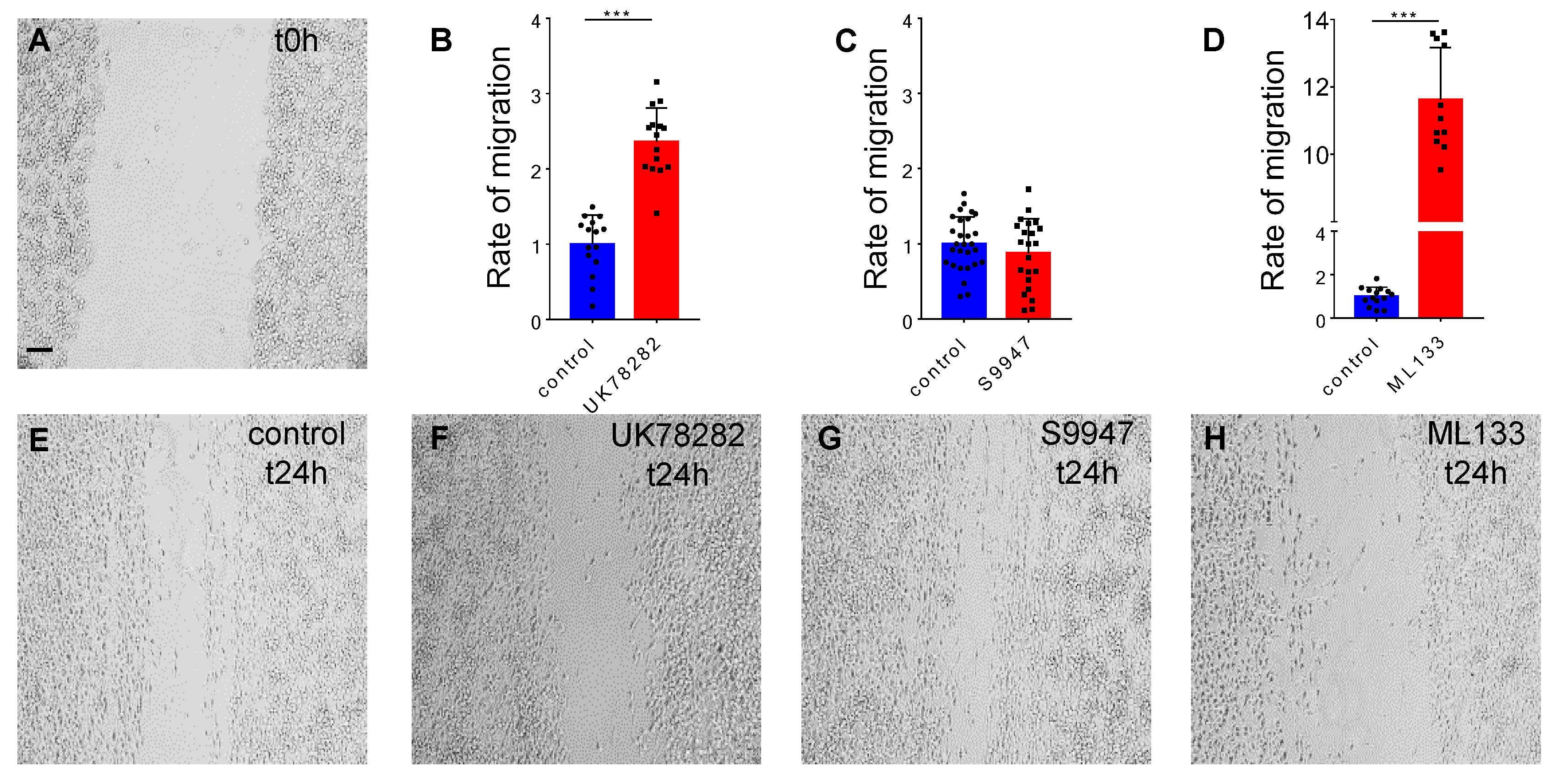
2.2. Potassium Channels Kv1.3 and Kir2.1 But Not Kv.1.5 Are Needed for BV2 Migration
2.3. Microglia Have an Innate Decreased Motility after the Spared Nerve Injury
3. Discussion
4. Materials and Methods
4.1. Animals and Surgery
4.2. Cell cultures
4.2.1. BV2 Cell Line Culture and Transfection
4.2.2. Primary Microglial Culture
4.3. Migration
4.3.1. BV2 Cell Line Migration
4.3.2. Primary Microglial Migration
4.4. Electrophysiology
4.5. Immunocytochemistry
4.6. Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | central nervous system |
| ROS | reactive oxygen species |
| NO | nitric oxide |
| LPS | lipopolysaccharide |
| SC-DH | spinal cord dorsal horn |
| SNI | spared nerve injury |
| I/V | intensity/voltage |
| siRNA | small interfering RNA |
| PBS | phosphate buffer saline |
| PFA | paraformaldehyde |
| BSA | bovine serum albumin |
References
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.-B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Vadakkan, K.I.; Kim, S.S.; Wu, L.-J.; Shang, Y.; Zhuo, M. Selective activation of microglia in spinal cord but not higher cortical regions following nerve injury in adult mouse. Mol. Pain 2008, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Savchenko, V.L. Microglia in ontogeny and brain pathology. Neurophysiology 1998, 30, 107–119. [Google Scholar] [CrossRef]
- Fenrich, K.K.; Weber, P.; Hocine, M.; Zalc, M.; Rougon, G.; Debarbieux, F. Long-term in vivo imaging of normal and pathological mouse spinal cord with sub-cellular resolution using implanted glass windows. J. Physiol. 2012, 16, 3665–3675. [Google Scholar] [CrossRef] [Green Version]
- Catalin, B.; Cupido, A.; Iancau, M.; Albu, C.V.; Kirchhoff, F. Microglia: First responders in the central nervous system. Rom. J. Morphol. Embryol. 2013, 54, 467–472. [Google Scholar]
- Catalin, B.; Stopper, L.; Balseanu, T.-A.; Scheller, A. The in situ morphology of microglia is highly sensitive to the mode of tissue fixation. J. Chem. Neuroanat. 2017, 86, 59–66. [Google Scholar] [CrossRef]
- Hu, X.; Leak, R.K.; Shi, Y.; Suenaga, J.; Gao, Y.; Zheng, P.; Chen, J. Microglial and macrophage polarization—New prospects for brain repair. Nat. Rev. Neurol. 2015, 11, 56–64. [Google Scholar] [CrossRef]
- Santiago, A.R.; Bernardino, L.; Agudo-Barriuso, M.; Gonçalves, J. Microglia in Health and Disease: A Double-Edged Sword. Mediat. Inflamm 2017, 2017, 1–2. [Google Scholar] [CrossRef]
- Surugiu, R.; Catalin, B.; Dumbrava, D.; Gresita, A.; Olaru, D.G.; Hermann, D.M.; Popa-Wagner, A. Intracortical Administration of the Complement C3 Receptor Antagonist Trifluoroacetate Modulates Microglia Reaction after Brain Injury. Neural Plast. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettenmann, H.; Hanisch, U.K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef]
- Yaremenko, L.M.; Grabovyi, O.M. Reactions of Microglial Cells in the Sensorimotor Cortex of Rats after Transient Ischemia. Neurophysiology 2017, 49, 107–112. [Google Scholar] [CrossRef]
- Donat, C.K.; Scott, G.; Gentleman, S.M.; Sastre, M. Microglial Activation in Traumatic Brain Injury. Front. Aging Neurosci. 2017, 9, 208. [Google Scholar] [CrossRef] [Green Version]
- Witcher, K.G.; Bray, C.E.; Dziabis, J.E.; McKim, D.B.; Benner, B.N.; Rowe, R.K.; Kokiko-Cochran, O.N.; Popovich, P.G.; Lifshitz, J.; Eiferman, D.S.; et al. Traumatic brain injury-induced neuronal damage in the somatosensory cortex causes formation of rod-shaped microglia that promote astrogliosis and persistent neuroinflammation. Glia 2018, 66, 2719–2736. [Google Scholar] [CrossRef] [PubMed]
- Davalos, D.; Ryu, J.K.; Merlini, M.; Baeten, K.M.; Le Moan, N.; Petersen, M.A.; Deerinck, T.J.; Smirnoff, D.S.; Bedard, C.; Hakozaki, H.; et al. Fibrinogen-induced perivascular microglial clustering is required for the development of axonal damage in neuroinflammation. Nat. Commun. 2012, 3, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orre, M.; Kamphuis, W.; Osborn, L.M.; Jansen, A.H.; Kooijman, L.; Bossers, K.; Hol, E.M. Isolation of glia from Alzheimer’s mice reveals inflammation and dysfunction. Neurobiol. Aging 2014, 35, 2746–2760. [Google Scholar] [CrossRef] [PubMed]
- Tischer, J.; Krueger, M.; Mueller, W.; Staszewski, O.; Prinz, M.; Streit, W.J.; Bechmann, I. Inhomogeneous distribution of Iba-1 characterizes microglial pathology in Alzheimer’s disease. Glia 2016, 64, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Suter, M.R.; Wen, Y.R.; Decosterd, I.; Ji, R.R. Do glial cells control pain? Neuron Glia Biol. 2007, 3, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.R.; Tan, P.H.; Cheng, J.K.; Liu, Y.C.; Ji, R.R. Role of microglia in neuropathic pain, postoperative pain, and morphine tolerance. J. Formos. Med. Assoc. 2011, 110, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Baggiolini, M. Chemokines and leukocyte traffic. Nat. Immunol. 2008, 9, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Häusler, K.G.; Prinz, M.; Nolte, C.; Weber, J.R.; Schumann, R.R.; Kettenmann, H.; Hanisch, U.K. Interferon-γ differentially modulates the release of cytokines and chemokines in lipopolysaccharide- and pneumococcal cell wall-stimulated mouse microglia and macrophages. Eur. J. Neurosci. 2002, 16, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.; Musso, N.; Giambirtone, M.; Grasso, M.; Spampinato, S.; Merlo, S.; Drago, F.; Lazzarino, G.; Sortino, M.; et al. Carnosine Prevents Aβ-Induced Oxidative Stress and Inflammation in Microglial Cells: A Key Role of TGF-β1. Cells 2019, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, H.B.; Li, J.; Lv, L.J.; Nie, B.J.; Lu, P.; Xue, F.; Zhang, Z.M. Eupatilin inhibits microglia activation and attenuates brain injury in intracerebral hemorrhage. Exp. Ther. Med. 2018, 16, 4005–4009. [Google Scholar] [CrossRef] [Green Version]
- Petrushenko, Y.A. P2X receptors: Peculiarities of the structure and modulation of the functions. Neurophysiology 2012, 44, 163–173. [Google Scholar] [CrossRef]
- Koizumi, S.; Ohsawa, K.; Inoue, K.; Kohsaka, S. Purinergic receptors in microglia: Functional modal shifts of microglia mediated by P2 and P1 receptors. Glia 2013, 61, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kazuhide, I.; Makoto, T. Microglia and neuropathic pain. Glia 2009, 57, 1469–1479. [Google Scholar] [CrossRef]
- Sperlágh, B.; Illes, P. Purinergic modulation of microglial cell activation. Purinergic Signal. 2007, 3, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, M.-F.; Taupenot, L.; Ulrich, G.; Aunis, D.; Ciesielski-Treska, J. Bacterial endotoxin induces [Ca2+]i transients and changes the organization of actin in microglia. Glia 1994, 11, 336–344. [Google Scholar] [CrossRef]
- Chao, C.C.; Gekker, G.; Hu, S.; Sheng, W.S.; Shark, K.B.; Bu, D.F.; Archer, S.; Bidlack, J.M.; Peterson, P.K. Kappa Opioid Receptors in Human Microglia Downregulate Human Immunodeficiency Virus 1 Expression. Proc. Natl. Acad. Sci. USA 1996, 93, 8051–8056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heese, K.; Fiebich, B.L.; Bauer, J.; Otten, U. Nerve growth factor (NGF) expression in rat microglia is induced by adenosine A(2a)-receptors. Neurosci. Lett. 1997, 231, 83–86. [Google Scholar] [CrossRef]
- Tanaka, J.; Fujita, H.; Matsuda, S.; Toku, K.; Sakanaka, M.; Maeda, N. Glucocorticoid- and mineralocorticoid receptors in microglial cells: The two receptors mediate differential effects of corticosteroids. Glia 1997, 20, 23–37. [Google Scholar] [CrossRef]
- Boucsein, C.; Kettenmann, H.; Nolte, C. Electrophysiological properties of microglial cells in normal and pathologic rat brain slices. Eur. J. Neurosci. 2000, 12, 2049–2058. [Google Scholar] [CrossRef]
- Ceron, J.; Troncoso, J. Facial nerve injuries cause changes in central nervous system microglial cells. Biomedica 2016, 36, 619–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jou, I.; Pyo, H.; Chung, S.; Jung, S.Y.; Gwag, B.J.; Joe, E.H. Expression of Kv1.5 K+ channels in activated microglia in vivo. Glia 1998, 24, 408–414. [Google Scholar] [CrossRef]
- Kotecha, S.A.; Schlichter, L.C. A Kv1.5 to Kv1.3 switch in endogenous hippocampal microglia and a role in proliferation. J. Neurosci. 1999, 19, 10680–10693. [Google Scholar] [CrossRef] [Green Version]
- Charolidi, N.; Schilling, T.; Eder, C. Microglial Kv1.3 Channels and P2Y12 Receptors Differentially Regulate Cytokine and Chemokine Release from Brain Slices of Young Adult and Aged Mice. PLoS ONE 2015, 10, e0128463. [Google Scholar] [CrossRef] [PubMed]
- Pannasch, U. The potassium channels Kv1 5 and Kv13 modulate distinct functions of microglia. Mol. Cell. Neurosci. 2006, 33, 401–411. [Google Scholar] [CrossRef]
- Rangaraju, S.; Raza, S.A.; Pennati, A.; Deng, Q.; Dammer, E.B.; Duong, D.; Pennington, M.W.; Tansey, M.G.; Lah, J.J.; Betarbet, R.; et al. A systems pharmacology-based approach to identify novel Kv1.3 channel-dependent mechanisms in microglial activation. J. Neuroinflammation 2017, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, P.S.; Kaufhold, J.P.; Blinder, P.; Friedman, B.; Drew, P.J.; Karten, H.J.; Lyden, P.D.; Kleinfeld, D. Correlations of neuronal and microvascular densities in murine cortex revealed by direct counting and colocalization of nuclei and vessels. J. Neurosci. 2009, 29, 14553–14570. [Google Scholar] [CrossRef] [Green Version]
- Gheorghe, R.; Deftu, A.; Filippi, A.; Grosu, A.; Bica-Popi, M.; Chiritoiu, M.; Chiritoiu, G.; Munteanu, C.; Silvestro, L.; Ristoiu, V.; et al. Silencing the Cytoskeleton Protein Iba1 (Ionized Calcium Binding Adapter Protein 1 ) Interferes with BV2 Microglia Functioning. Cell. Mol. Neurobiol. 2020, 40, 1011–1027. [Google Scholar] [CrossRef]
- Fomina, A.F.; Nguyen, H.M.; Wulff, H. Kv1.3 inhibition attenuates neuroinflammation through disruption of microglial calcium signaling ABSTRACT. Channels 2021, 15, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Januszyk, M.; Rennert, R.C.; Sorkin, M.; Maan, Z.N.; Wong, L.K.; Whittam, A.J.; Whitmore, A.; Duscher, D.; Gurtner, G.C. Evaluating the Effect of Cell Culture on Gene Expression in Primary Tissue Samples Using Microfluidic-Based Single Cell Transcriptional Analysis. Microarrays 2015, 4, 540–550. [Google Scholar] [CrossRef]
- Henn, A.; Lund, S.; Hedtjärn, M.; Schrattenholz, A.; Pörzgen, P.; Leist, M. The suitability of BV2 cells as alternative model system for primary microglia cultures or for animal experiments examining brain inflammation. ALTEX 2009, 26, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a unique TGF-beta-dependent molecular and functional signature in microglia. Nat. Neurosci. 2014, 17, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shi, Y.; Wang, G.; Li, Y.; Cheng, L.; Wang, Y. Memantine selectively prevented the induction of dynamic allodynia by blocking Kir2.1 channel and inhibiting the activation of microglia in spinal dorsal horn of mice in spared nerve injury model. Mol. Pain 2019, 15, 174480691983894. [Google Scholar] [CrossRef] [Green Version]
- Bozic, I.; Tesovic, K.; Laketa, D.; Adzic, M.; Jakovljevic, M.; Bjelobaba, I.; Savic, D.; Nedeljkovic, N.; Pekovic, S.; Lavrnja, I. Voltage Gated Potassium Channel Kv1.3 Is Upregulated on Activated Astrocytes in Experimental Autoimmune Encephalomyelitis. Neurochem. Res. 2018, 43, 1020–1034. [Google Scholar] [CrossRef] [Green Version]
- Bozic, I.; Savic, D.; Milosevic, A.; Janjic, M.; Laketa, D.; Tesovic, K.; Bjelobaba, I.; Jakovljevic, M.; Nedeljkovic, N.; Pekovic, S.; et al. The Potassium Channel Kv1.5 Expression Alters During Experimental Autoimmune Encephalomyelitis. Neurochem. Res. 2019, 44, 2733–2745. [Google Scholar] [CrossRef]
- Wang, H.R.; Wu, M.; Yu, H.; Long, S.; Stevens, A.; Engers, D.W.; Sackin, H.; Daniels, J.S.; Dawson, E.S.; Hopkins, C.R.; et al. Selective inhibition of the K ir2 family of inward rectifier potassium channels by a small molecule probe: The discovery, SAR, and pharmacological characterization of ML133. ACS Chem. Biol. 2011, 6, 845–856. [Google Scholar] [CrossRef] [Green Version]
- Hanson, D.C.; Nguyen, A.; Mather, R.J.; Rauer, H.; Koch, K.; Burgess, L.E.; Rizzi, J.P.; Donovan, C.B.; Bruns, M.J.; Canniff, P.C.; et al. UK-78,282, a novel piperidine compound that potently blocks the Kv1.3 voltage-gated potassium channel and inhibits human T cell activation. Br. J. Pharmacol. 1999, 126, 1707–1716. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, A.; Gutcher, I.; Kopp, K.; Brendel, J.; Bosch, R.F.; Busch, A.E.; Gögelein, H. Characterization of a novel Kv1.5 channel blocker in Xenopus oocytes, CHO cells, human and rat cardiomyocytes. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 472–478. [Google Scholar] [CrossRef]
- Haynes, S.E.; Hollopeter, G.; Yang, G.; Kurpius, D.; Dailey, M.E.; Gan, W.B.; Julius, D. The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 2006, 9, 1512–1519. [Google Scholar] [CrossRef]
- Jung, S.; Aliberti, J.; Graemmel, P.; Sunshine, M.J.; Kreutzberg, G.W.; Sher, A.; Littman, D.R. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol. Cell. Biol. 2000, 20, 4106–4114. [Google Scholar] [CrossRef] [Green Version]
- Pertin, M.; Gosselin, R.-D.; Decosterd, I. The Spared Nerve Injury Model of Neuropathic Pain. Methods Mol. Biol. 2012, 851, 205–212. [Google Scholar] [PubMed]
- Gattlen, C.; Deftu, A.F.; Tonello, R.; Ling, Y.; Berta, T.; Ristoiu, V.; Suter, M.R. The inhibition of Kir2.1 potassium channels depolarizes spinal microglial cells, reduces their proliferation, and attenuates neuropathic pain. Glia 2020, 68, 2119–2135. [Google Scholar] [CrossRef]
- Deftu, A.F.; Ristoiu, V.; Suter, M.R. Intrathecal Administration of CXCL1 Enhances Potassium Currents in Microglial Cells. Pharmacology 2018, 101, 262–268. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strutz-Seebohm, N.; Gutcher, I.; Decher, N.; Steinmeyer, K.; Lang, F.; Seebohm, G. Comparison of potent Kv1.5 potassium channel inhibitors reveals the molecular basis for blocking kinetics and binding mode. Cell. Physiol. Biochem. 2007, 20, 791–800. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham | 1 ± 0.4923, n = 70 | |||
|---|---|---|---|---|
| SNI | 0.113 ± 0.13, n = 61, p < 0.001 | |||
| K1.3 (UK78282) | Kv1.5 (S9947) | Kir2.1 (ML133) | ||
| Sham | Control | 1 ± 0.4188, n = 21 | 1 ± 0.6214, n = 28 | 1 ± 0.3736, n = 21 |
| Inhibitor | 0.425 ± 0.294, n = 21, p < 0.001 | 0.271 ± 0.2633, n = 28, p < 0.001 | 0.075 ± 0.06508, n = 21, p < 0.001 | |
| SNI | Control | 1 ± 1.141, n = 21 | 1 ± 1.147, n = 21 | 1 ± 0.9143, n = 21 |
| Inhibitor | 0.170 ± 0.2331, n = 21, p < 0.001 | 1.065 ± 1.415, n = 20, p > 0.05 | 0.447 ± 0.5559, n = 21, p < 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anton, R.; Ghenghea, M.; Ristoiu, V.; Gattlen, C.; Suter, M.-R.; Cojocaru, P.A.; Popa-Wagner, A.; Catalin, B.; Deftu, A.-F. Potassium Channels Kv1.3 and Kir2.1 But Not Kv1.5 Contribute to BV2 Cell Line and Primary Microglial Migration. Int. J. Mol. Sci. 2021, 22, 2081. https://doi.org/10.3390/ijms22042081
Anton R, Ghenghea M, Ristoiu V, Gattlen C, Suter M-R, Cojocaru PA, Popa-Wagner A, Catalin B, Deftu A-F. Potassium Channels Kv1.3 and Kir2.1 But Not Kv1.5 Contribute to BV2 Cell Line and Primary Microglial Migration. International Journal of Molecular Sciences. 2021; 22(4):2081. https://doi.org/10.3390/ijms22042081
Chicago/Turabian StyleAnton, Ruxandra, Mihail Ghenghea, Violeta Ristoiu, Christophe Gattlen, Marc-Rene Suter, Petre Alexandru Cojocaru, Aurel Popa-Wagner, Bogdan Catalin, and Alexandru-Florian Deftu. 2021. "Potassium Channels Kv1.3 and Kir2.1 But Not Kv1.5 Contribute to BV2 Cell Line and Primary Microglial Migration" International Journal of Molecular Sciences 22, no. 4: 2081. https://doi.org/10.3390/ijms22042081
APA StyleAnton, R., Ghenghea, M., Ristoiu, V., Gattlen, C., Suter, M. -R., Cojocaru, P. A., Popa-Wagner, A., Catalin, B., & Deftu, A. -F. (2021). Potassium Channels Kv1.3 and Kir2.1 But Not Kv1.5 Contribute to BV2 Cell Line and Primary Microglial Migration. International Journal of Molecular Sciences, 22(4), 2081. https://doi.org/10.3390/ijms22042081