
A Comprehensive miRNome Analysis of Macrophages Isolated from db/db Mice and Selected miRNAs Involved in Metabolic Syndrome-Associated Cardiac Remodeling

 , ,
, ,
Abstract
:1. Introduction
2. Results
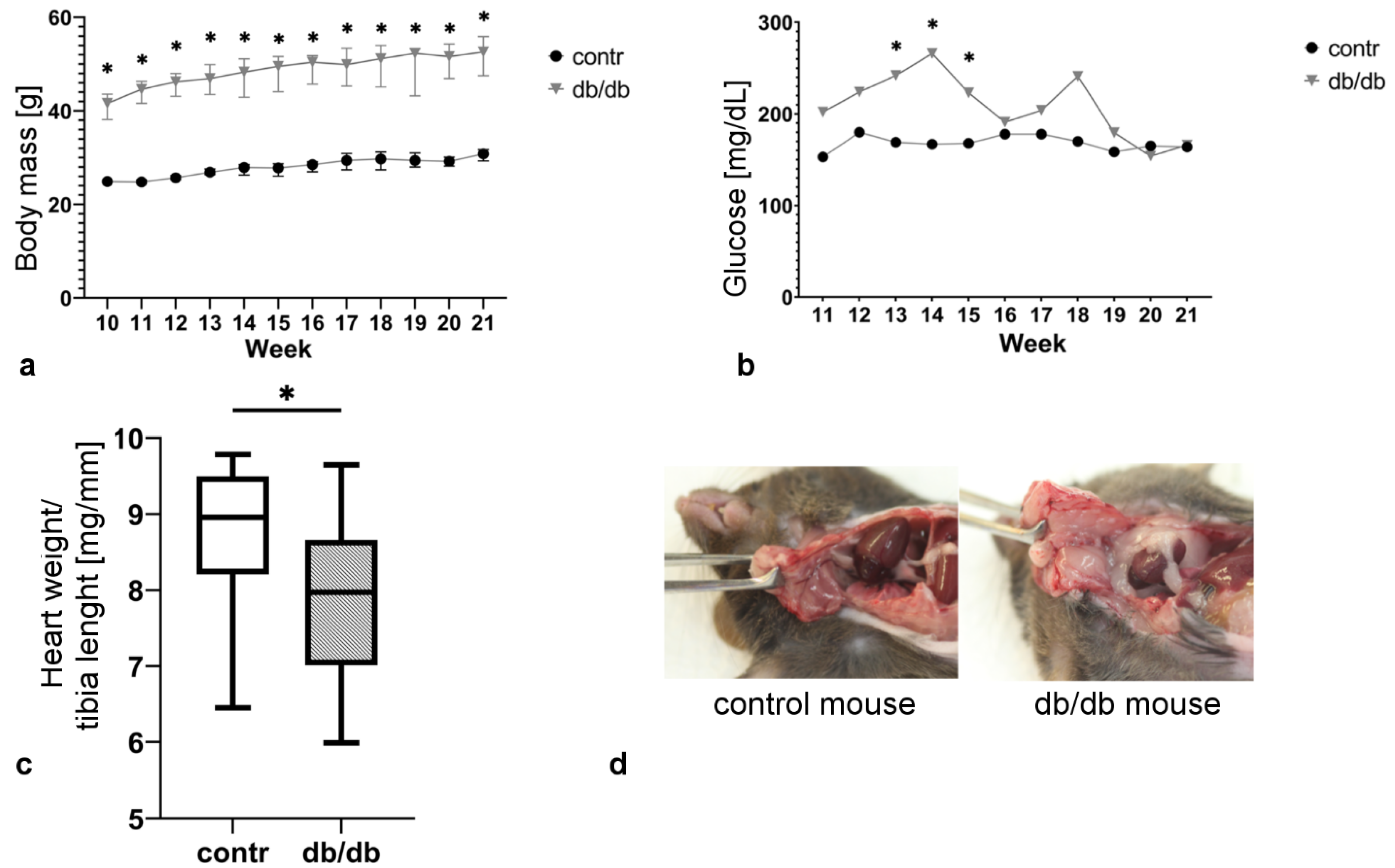
2.1. Db/db Mice Exhibit Obesity and Hyperglycemia
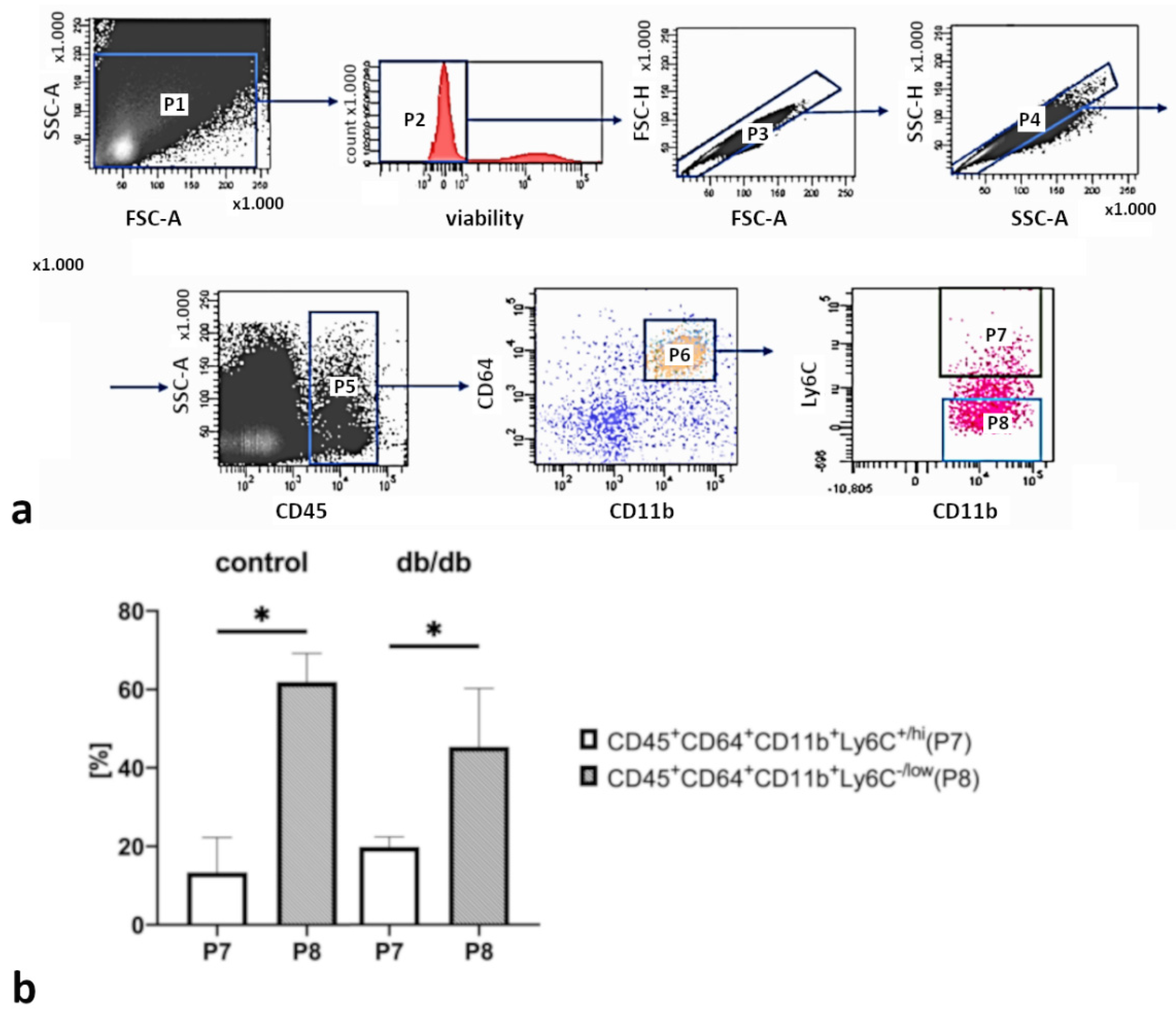
2.2. The Significant Decrease in Macrophage Numbers in db/db vs. Control Mouse Hearts
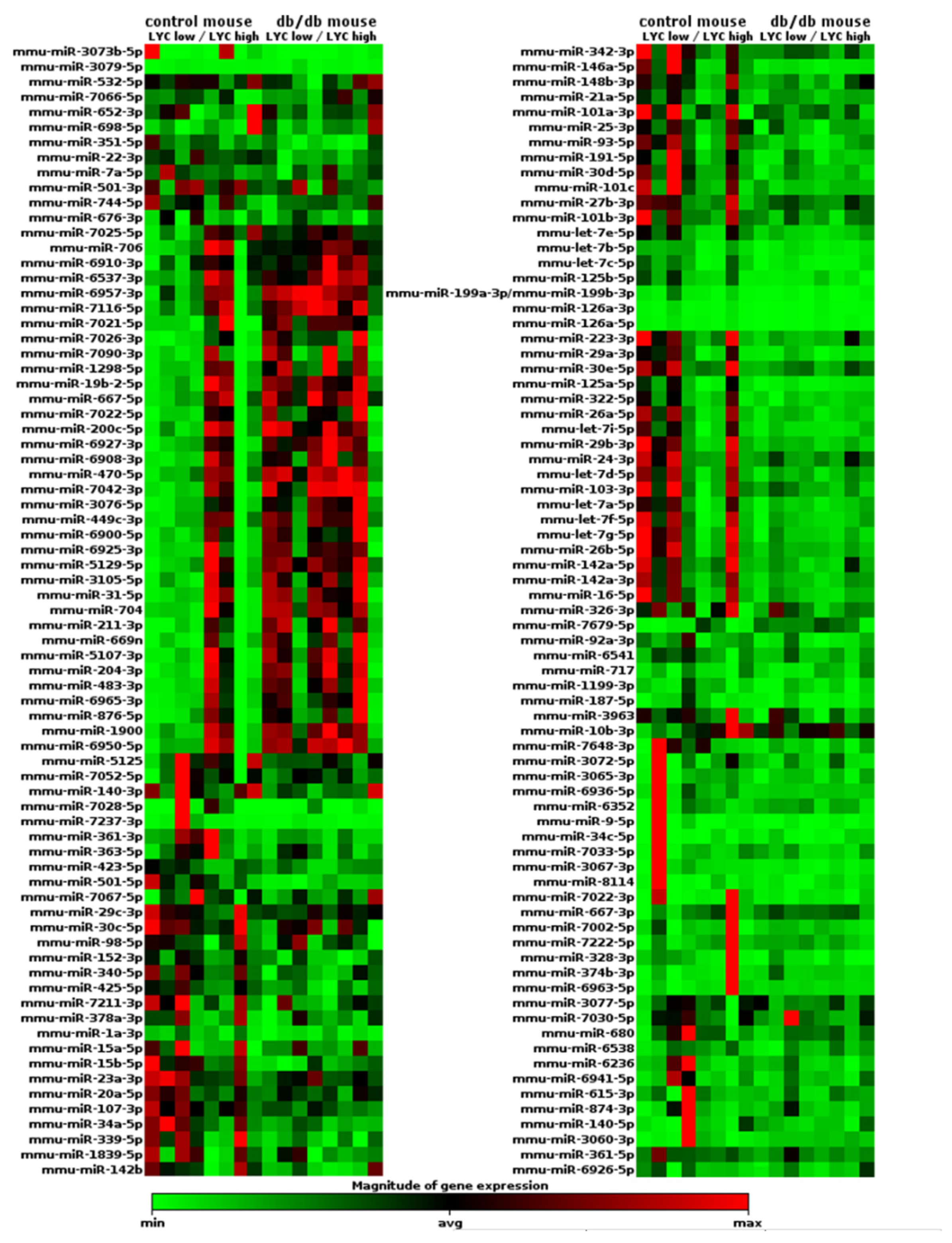
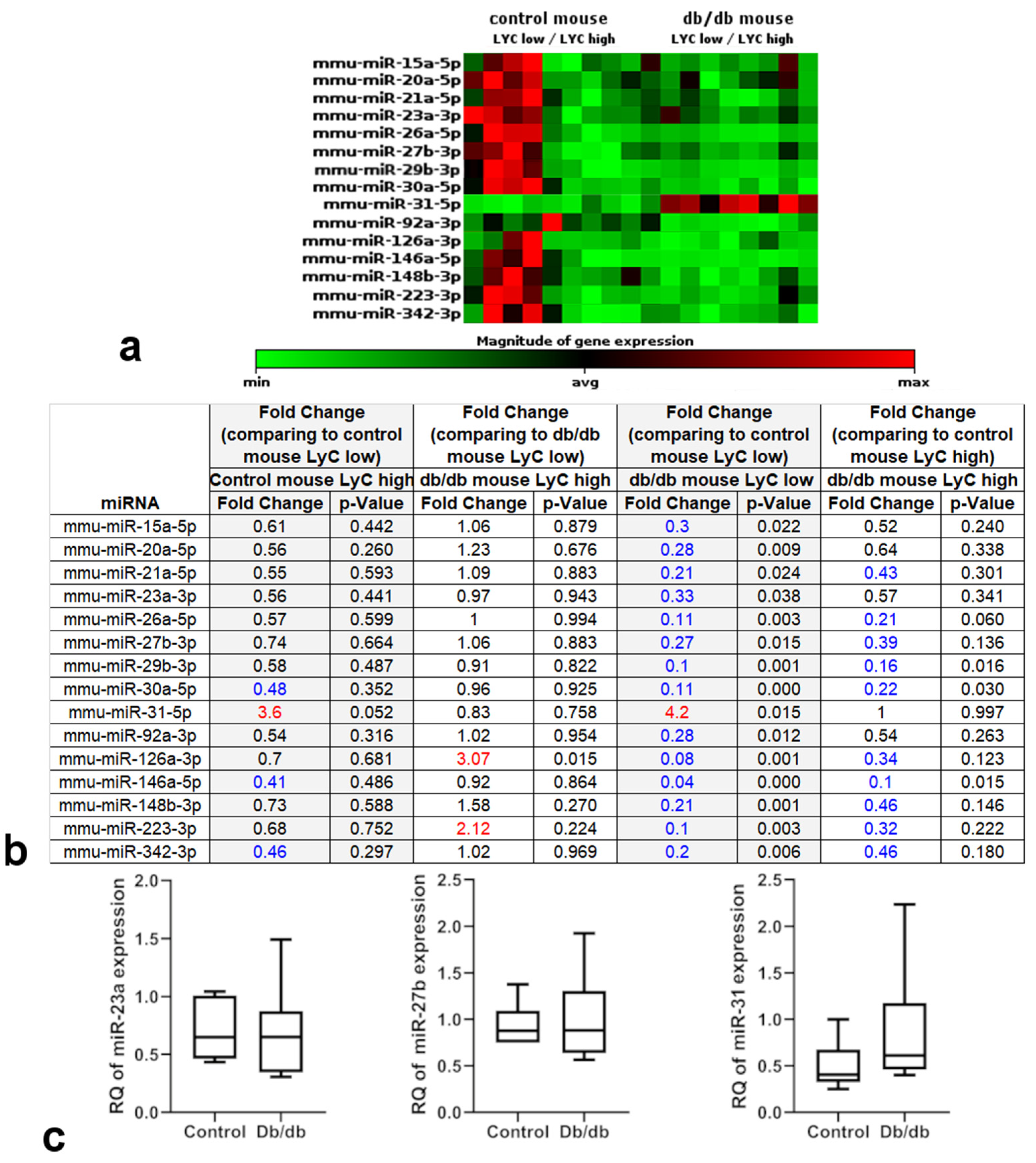
2.3. miRNA Expression Profile Changes in db/db Mouse Cardiac Macrophages but Not in Cardiac Tissue
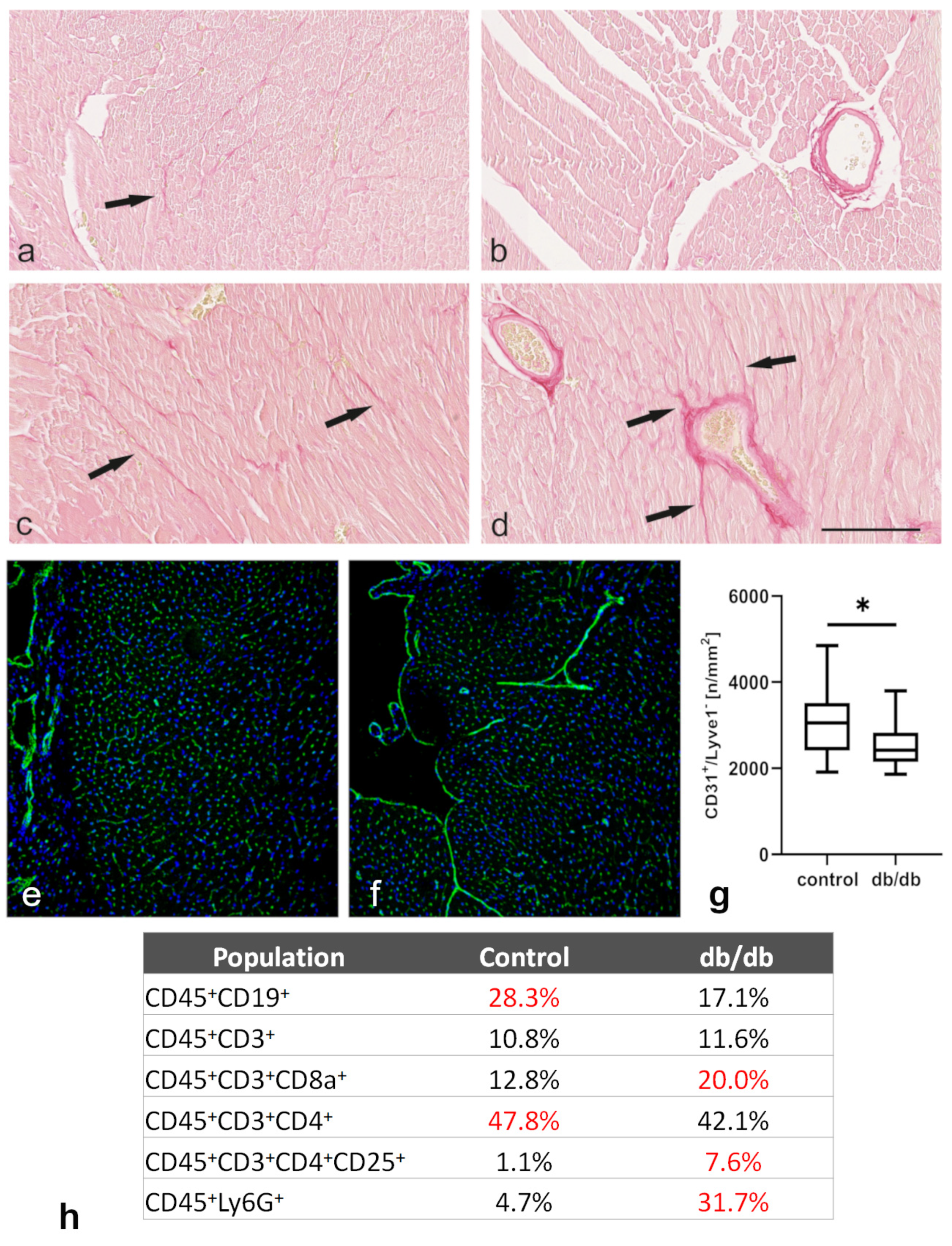
2.4. Morphological Analysis of Tissue Sections and Cell Suspensions Obtained from Control and db/db Mouse Hearts Showed Evidence of Cardiac Fibrosis, Inflammation, and Microvascular Rarefaction
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Assessment of Macrophage and Microvascular Density in a Confocal Microscope
4.3. Cardiac Macrophage Isolation by Flow Cytometry Sorting
4.4. Inflammatory Cell Profile of Cardiac Tissue Cell Suspension
4.5. RNA Isolation, Total miRNA Library Preparation, and miRNA Sequencing
4.6. miRNA-Seq Data Analysis
4.7. RT-PCR Analysis of Cardiac miRNAs
4.8. Picrosirius Red Staining for Collagen Deposits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Function | Target Gene | References |
|---|---|---|---|
| miR-15a-5p | Angiogenesis, fibrosis | Tie-2, fibrosis via VEGF and EMT; and TGF-β1/Smad2 | [61,62,63] |
| miR-20a-5p | fibrosis | ALK-5, TGFβR2, SARA, CD36 | [64,65] |
| miR-21a-5p | fibrosis, inflammation | PPARα, SMAD7, PTEN | [66,67,68] |
| miR-23a-3p | angiogenesis | SEMA6A, Sprouty2 | [69,70] |
| miR-26a-5p | inflammation, angiogenesis | PTEN, VEGF-A, PI3K/AKT | [71,72] |
| miR-27b-3p | fibrosis, inflammation, angiogenesis | ALK5, IL-1, IL-6, TNFα, MCP1, SEMA6A, Sprouty2 | [69,73,74,75,76,77] |
| miR-29b-3p | fibrosis, extracellular matrix deposition, angiogenesis | Wnt, elastin, collagen, fibronectin, IGF-1 | [47,48,78,79] |
| miR-30a-5p | fibrosis, inflammation, angiogenesis | Angiopoietin-2, VCAM-1, CTGF, Beclin1, Dll4, Snail1, Wnt | [80,81,82,83,84,85,86] |
| miR-31-5p | lymphangiogenesis, fibrosis, angiogenesis, inflammation | TNFSF15, PKCε, Prox1, FOXC2, E-selectin | [40,45,87,88] |
| miR-92a-3p | angiogenesis | KLF2, KLF4, TF | [89,90] |
| miR-126a-3p | angiogenesis, inflammation | VCAM-1, Spred-1, PI3KR2, VEGF-A | [91,92,93,94,95] |
| miR-146a-5p | inflammation, fibrosis, angiogenesis | NFκB, TRAF6, IRAK, MYD88, SMAD4, MAPK | [96,97,98] |
| miR-148b-3p | angiogenesis, fibrosis, EMT | FGF-2, DNMT, PTEN, Wnt-β-catenin | [99,100,101] |
| miR-223-3p | inflammation, fibrosis | ICAM-1, RASA1, FBXW7 | [102,103,104] |
| miR-342-3p | angiogenesis | FGF11 | [105] |
| miRNA | Target Gene | Effect on Macrophage and on Macrophage Phenotype Alteration | References |
|---|---|---|---|
| miR-15a-5p | TNIP2 JAK2 | Pro-inflammatory in vitro and in mouse model of sepsis Anti-inflammatory in in vitro model of allergic rhinitis | [49,50] |
| miR-20a-5p | HIF-2α | Anti-angiogenic in tumor associated macrophages (TAMs) | [106] |
| miR-21a-5p | PDCD4 IL-10 | Anti-inflammatory in Brucella infected bone marrow derived macrophages Suppresses M-2 macrophage polarization in primary bone-marrow derived macrophages during particle-induced osteolysis | [107,108] |
| miR-23a-3p | TNFAIP3 | Tumor associated macrophages (TAMs) switching towards M1 phenotype | [109] |
| miR-26a-5p | CTGF | Reduces pro-inflammatory factor TNF-α, IL-6, IL-1β expression of LPS-induced mouse alveolar macrophages | [110] |
| miR-27b-3p | MIP-1β | Anti-inflammatory effect in bone-marrow derived macrophages in vitro | [111] |
| miR-29b-3p | DNMTs (confirmed indirectly) | Inhibits phagocytic function in alveolar macrophages in syngeneic murine model of bone marrow transplantation | [112] |
| miR-30a-5p | SOCS3 | Anti-inflammatory in LPS stimulated RAW 264.7 macrophages | [113] |
| miR-31-5p | Cab39 | Pro-inflammatory in LPS-induced alveolar macrophages in vitro | [114] |
| miR-92a-3p | PTEN KLF4 | Pro-inflammatory in LPS-induced acute lung injury mouse model Pro-inflammatory in monocyte-derived macrophages under atheroprone microenvironment in vitro | [115,116] |
| miR-126a-3p | ATF3, ATP1B1, ATP9A and RAI14 | Reduction of pro-inflammatory cytokine/ chemokine secretion by primary human macrophages and increase in their phagocytic activity | [117] |
| miR-146a-5p | Notch1 | M2 phenotype polarization of microglia following brain stroke in vivo and in vitro | [118] |
| miR-148b-3p | Nox2 | Anti-inflammatory and lowering ROS production in macrophages in myocardial infarction mouse model | [119] |
| miR-223-3p | Nlrp3 Pknox1 Rasa1 and Nfat5 | Anti-inflammatory in acute and chronic hepatic injury, and in vitro studies M2 phenotype polarization in bone marrow–derived macrophages stimulated with LPS and in adipose tissue of mice on a high-fat diet M2 phenotype polarization in adipose-tissue macrophages isolated from mice on a high-fat diet | [120,121,122] |
| miR-342-3p | Chi3l1 (not confirmed) | Suppresses inflammation and lipid uptake in THP-1 cells | [123] |
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberti, K.G.; Zimmet, P. The metabolic syndrome: Time to reflect. Curr. Diabetes Rep. 2006, 6, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Swarup, S.; Goyal, A.; Grigorova, Y.; Zeltser, R. Metabolic Syndrome. In StatPearls; StatPearls Publishing Copyright © 2021; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020. [Google Scholar]
- Paulus, W.J.; Tschope, C. A novel paradigm for heart failure with preserved ejection fraction: Comorbidities drive myocardial dysfunction and remodeling through coronary microvascular endothelial inflammation. J. Am. Coll. Cardiol. 2013, 62, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, S.J.; Cuijpers, I.; Heymans, S.; Jones, E.A.V. Cellular and Molecular Differences between HFpEF and HFrEF: A Step Ahead in an Improved Pathological Understanding. Cells 2020, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Adameova, A.; Dhalla, N.S. Role of microangiopathy in diabetic cardiomyopathy. Heart Fail. Rev. 2014, 19, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Hulsmans, M.; Sager, H.B.; Roh, J.D.; Valero-Munoz, M.; Houstis, N.E.; Iwamoto, Y.; Sun, Y.; Wilson, R.M.; Wojtkiewicz, G.; Tricot, B.; et al. Cardiac macrophages promote diastolic dysfunction. J. Exp. Med. 2018, 215, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, G.; Schneider, C.; Wong, N.; Bredemeyer, A.; Hulsmans, M.; Nahrendorf, M.; Epelman, S.; Kreisel, D.; Liu, Y.; Itoh, A.; et al. The human heart contains distinct macrophage subsets with divergent origins and functions. Nat. Med. 2018, 24, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Hulin, A.; Anstine, L.J.; Kim, A.J.; Potter, S.J.; DeFalco, T.; Lincoln, J.; Yutzey, K.E. Macrophage Transitions in Heart Valve Development and Myxomatous Valve Disease. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.J.; Xu, N.; Yutzey, K.E. Macrophage lineages in heart valve development and disease. Cardiovasc. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Nahrendorf, M. Myeloid Cells Remodel the Mitral Valve. Circulation 2018, 137, 2494–2496. [Google Scholar] [CrossRef]
- DeBerge, M.; Shah, S.J.; Wilsbacher, L.; Thorp, E.B. Macrophages in Heart Failure with Reduced versus Preserved Ejection Fraction. Trends Mol. Med. 2019, 25, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lu, Y.L.; Zhao, W.T.; Zhong, J.; Lin, X.; Sun, Z.; He, Y.; Chen, M.; Zheng, L.R. Distinct origins and functions of cardiac orthotopic macrophages. Basic Res. Cardiol. 2020, 115, 8. [Google Scholar] [CrossRef]
- Zaman, R.; Hamidzada, H.; Epelman, S. Exploring cardiac macrophage heterogeneity in the healthy and diseased myocardium. Curr. Opin. Immunol. 2020, 68, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sun, X.; Shan, P.F. MicroRNAs and Cardiovascular Disease in Diabetes Mellitus. Biomed. Res. Int. 2017, 2017, 4080364. [Google Scholar] [CrossRef] [Green Version]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: Roles in cardiovascular development and disease. Cardiovasc. Pathol. 2021, 50, 107296. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; Moore, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Riehle, C.; Bauersachs, J. Small animal models of heart failure. Cardiovasc. Res. 2019, 115, 1838–1849. [Google Scholar] [CrossRef] [PubMed]
- Alex, L.; Russo, I.; Holoborodko, V.; Frangogiannis, N.G. Characterization of a mouse model of obesity-related fibrotic cardiomyopathy that recapitulates features of human heart failure with preserved ejection fraction. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H934–H949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puff, R.; Dames, P.; Weise, M.; Goke, B.; Seissler, J.; Parhofer, K.G.; Lechner, A. Reduced proliferation and a high apoptotic frequency of pancreatic beta cells contribute to genetically-determined diabetes susceptibility of db/db BKS mice. Horm. Metab. Res 2011, 43, 306–311. [Google Scholar] [CrossRef]
- Choi, J.; Kim, K.J.; Koh, E.J.; Lee, B.Y. Gelidium elegans Extract Ameliorates Type 2 Diabetes via Regulation of MAPK and PI3K/Akt Signaling. Nutrients 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.D.; Li, Z.; Wagner, R.; Yue, P.; Tsao, P.; Nestorova, G.; Huang, M.; Hirschberg, D.L.; Yock, P.G.; Quertermous, T.; et al. Transcriptome alteration in the diabetic heart by rosiglitazone: Implications for cardiovascular mortality. PLoS ONE 2008, 3, e2609. [Google Scholar] [CrossRef]
- Wang, S.; Wang, B.; Wang, Y.; Tong, Q.; Liu, Q.; Sun, J.; Zheng, Y.; Cai, L. Zinc Prevents the Development of Diabetic Cardiomyopathy in db/db Mice. Int. J. Mol. Sci. 2017, 18, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, F.C.; Spurgeon, H.A.; Rakusan, K.; Weisfeldt, M.L.; Lakatta, E.G. Use of tibial length to quantify cardiac hypertrophy: Application in the aging rat. Am. J. Physiol. 1982, 243, H941–H947. [Google Scholar] [CrossRef] [PubMed]
- Papinska, A.M.; Soto, M.; Meeks, C.J.; Rodgers, K.E. Long-term administration of angiotensin (1-7) prevents heart and lung dysfunction in a mouse model of type 2 diabetes (db/db) by reducing oxidative stress, inflammation and pathological remodeling. Pharmacol. Res. 2016, 107, 372–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Borght, K.; Lambrecht, B.N. Heart macrophages and dendritic cells in sickness and in health: A tale of a complicated marriage. Cell. Immunol. 2018, 330, 105–113. [Google Scholar] [CrossRef]
- Hilgendorf, I.; Gerhardt, L.M.; Tan, T.C.; Winter, C.; Holderried, T.A.; Chousterman, B.G.; Iwamoto, Y.; Liao, R.; Zirlik, A.; Scherer-Crosbie, M.; et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ. Res. 2014, 114, 1611–1622. [Google Scholar] [CrossRef] [Green Version]
- Epelman, S.; Lavine, K.J.; Beaudin, A.E.; Sojka, D.K.; Carrero, J.A.; Calderon, B.; Brija, T.; Gautier, E.L.; Ivanov, S.; Satpathy, A.T.; et al. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity 2014, 40, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Shen, Y.; Zhang, R.; Sugi, K.; Vasudevan, N.T.; Alaiti, M.A.; Sweet, D.R.; Zhou, L.; Qing, Y.; Gerson, S.L.; et al. Distinct roles of resident and nonresident macrophages in nonischemic cardiomyopathy. Proc. Natl. Acad. Sci. USA 2018, 115, E4661–E4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peet, C.; Ivetic, A.; Bromage, D.I.; Shah, A.M. Cardiac monocytes and macrophages after myocardial infarction. Cardiovasc. Res. 2020, 116, 1101–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.; Tacke, R.; Hedrick, C.C.; Hanna, R.N. Nonclassical patrolling monocyte function in the vasculature. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1306–1316. [Google Scholar] [CrossRef] [Green Version]
- Kimball, A.; Schaller, M.; Joshi, A.; Davis, F.M.; denDekker, A.; Boniakowski, A.; Bermick, J.; Obi, A.; Moore, B.; Henke, P.K.; et al. Ly6C(Hi) Blood Monocyte/Macrophage Drive Chronic Inflammation and Impair Wound Healing in Diabetes Mellitus. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1102–1114. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, C.; Liu, L.; Xi, A.; Chen, B.; Li, Y.; Du, J. Macrophage-Derived mir-155-Containing Exosomes Suppress Fibroblast Proliferation and Promote Fibroblast Inflammation during Cardiac Injury. Mol. Ther. 2017, 25, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol. Cell 2010, 39, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; He, Y.; Ling, S.; Chen, Z.; Yan, G. MiR-15a Functions as a Diagnostic Biomarker for Coronary Artery Disease. Clin. Lab. 2020, 66. [Google Scholar] [CrossRef]
- Sadeghzadeh, S.; Dehghani Ashkezari, M.; Seifati, S.M.; Vahidi Mehrjardi, M.Y.; Dehghan Tezerjani, M.; Sadeghzadeh, S.; Ladan, S.A.B. Circulating miR-15a and miR-222 as Potential Biomarkers of Type 2 Diabetes. Diabetes Metab. Syndr. Obes. 2020, 13, 3461–3469. [Google Scholar] [CrossRef]
- He, Q.F.; Wang, L.X.; Zhong, J.M.; Hu, R.Y.; Fang, L.; Wang, H.; Gong, W.W.; Zhang, J.; Pan, J.; Yu, M. Circulating MicroRNA-21 is Downregulated in Patients with Metabolic Syndrome. Biomed. Environ. Sci. 2016, 29, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, S.; Jia, Q.; Zhang, A.; Li, Y.; Zhu, Y.; Lv, S.; Zhang, J. The microRNA in ventricular remodeling: The miR-30 family. BioSci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.T.; Liu, H.L.; Zhai, B.B.; Zhang, K.; Xu, G.C.; Peng, X.M.; Zhang, Q.Z.; Li, L.Y. Vascular endothelial growth factor suppresses TNFSF15 production in endothelial cells by stimulating miR-31 and miR-20a expression via activation of Akt and Erk signals. FEBS Open Bio 2017, 7, 108–117. [Google Scholar] [CrossRef]
- Wada, H.; Satoh, N.; Kitaoka, S.; Ono, K.; Morimoto, T.; Kawamura, T.; Nakano, T.; Fujita, M.; Kita, T.; Shimatsu, A.; et al. Soluble VEGF receptor-2 is increased in sera of subjects with metabolic syndrome in association with insulin resistance. Atherosclerosis 2010, 208, 512–517. [Google Scholar] [CrossRef]
- Wada, H.; Ura, S.; Kitaoka, S.; Satoh-Asahara, N.; Horie, T.; Ono, K.; Takaya, T.; Takanabe-Mori, R.; Akao, M.; Abe, M.; et al. Distinct characteristics of circulating vascular endothelial growth factor-a and C levels in human subjects. PLoS ONE 2011, 6, e29351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastiani, G.; Nigi, L.; Spagnuolo, I.; Morganti, E.; Fondelli, C.; Dotta, F. MicroRNA profiling in sera of patients with type 2 diabetes mellitus reveals an upregulation of miR-31 expression in subjects with microvascular complications. J. Biomed. Sci. Eng. 2013, 6, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.C.; Lilyanna, S.; Wang, P.; Vardy, L.A.; Jiang, X.; Armugam, A.; Jeyaseelan, K.; Richards, A.M. MicroRNA-31 promotes adverse cardiac remodeling and dysfunction in ischemic heart disease. J. Mol. Cell. Cardiol. 2017, 112, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Pedrioli, D.M.; Karpanen, T.; Dabouras, V.; Jurisic, G.; van de Hoek, G.; Shin, J.W.; Marino, D.; Kalin, R.E.; Leidel, S.; Cinelli, P.; et al. miR-31 functions as a negative regulator of lymphatic vascular lineage-specific differentiation in vitro and vascular development in vivo. Mol. Cell. Biol. 2010, 30, 3620–3634. [Google Scholar] [CrossRef] [Green Version]
- Kong, Z.; Wang, Y.; Zhang, Y.; Shan, W.; Wu, J.; Wang, Q. MicroRNA-126 promotes endothelial progenitor cell proliferation and migration ability via the Notch pathway. Cardiovasc. Diagn. Ther. 2020, 10, 490–499. [Google Scholar] [CrossRef]
- Van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [Green Version]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.; Gruter, L.; Werfel, S.; Giosele, S.; Brunner, A.D.; Esfandyari, D.; Papadopoulou, A.S.; De Strooper, B.; et al. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef]
- Lou, Y.; Huang, Z. microRNA-15a-5p participates in sepsis by regulating the inflammatory response of macrophages and targeting TNIP2. Exp. Ther. Med. 2020, 19, 3060–3068. [Google Scholar] [CrossRef]
- Liu, H.W.; Hu, Z.L.; Li, H.; Tan, Q.F.; Tong, J.; Zhang, Y.Q. Knockdown of lncRNA ANRIL suppresses the production of inflammatory cytokines and mucin 5AC in nasal epithelial cells via the miR-15a-5p/JAK2 axis. Mol. Med. Rep. 2021, 23. [Google Scholar] [CrossRef]
- Dangwal, S.; Thum, T. microRNA therapeutics in cardiovascular disease models. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 185–203. [Google Scholar] [CrossRef]
- De Couto, G. Macrophages in cardiac repair: Environmental cues and therapeutic strategies. Exp. Mol. Med. 2019, 51, 159. [Google Scholar] [CrossRef] [Green Version]
- Barwari, T.; Joshi, A.; Mayr, M. MicroRNAs in Cardiovascular Disease. J. Am. Coll. Cardiol. 2016, 68, 2577–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; De Guire, V. How close are miRNAs from clinical practice? A perspective on the diagnostic and therapeutic market. Ejifcc 2019, 30, 114–127. [Google Scholar]
- Femminò, S.; Penna, C.; Margarita, S.; Comità, S.; Brizzi, M.F.; Pagliaro, P. Extracellular vesicles and cardiovascular system: Biomarkers and Cardioprotective Effectors. Vasc. Pharmacol. 2020, 135, 106790. [Google Scholar] [CrossRef]
- Ren, W.; Hou, J.; Yang, C.; Wang, H.; Wu, S.; Wu, Y.; Zhao, X.; Lu, C. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J. Exp. Clin. Cancer Res. 2019, 38, 62. [Google Scholar] [CrossRef]
- He, S.; Chen, D.; Hu, M.; Zhang, L.; Liu, C.; Traini, D.; Grau, G.E.; Zeng, Z.; Lu, J.; Zhou, G.; et al. Bronchial epithelial cell extracellular vesicles ameliorate epithelial-mesenchymal transition in COPD pathogenesis by alleviating M2 macrophage polarization. Nanomedicine 2019, 18, 259–271. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Puchtler, H.; Waldrop, F.S.; Valentine, L.S. Polarization microscopic studies of connective tissue stained with picro-sirius red FBA. Beiträge zur Pathologie 1973, 150, 174–187. [Google Scholar] [CrossRef]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Besnier, M.; Shantikumar, S.; Anwar, M.; Dixit, P.; Chamorro-Jorganes, A.; Sweaad, W.; Sala-Newby, G.; Madeddu, P.; Thomas, A.C.; Howard, L.; et al. miR-15a/-16 Inhibit Angiogenesis by Targeting the Tie2 Coding Sequence: Therapeutic Potential of a miR-15a/16 Decoy System in Limb Ischemia. Mol. Ther. Nucleic Acids 2019, 17, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; He, Q.; Chen, Y.; Yu, D.; Sun, L.; Cheng, G.; Liu, D.; Xiao, J.; Zhao, Z. miR-15a-5p suppresses inflammation and fibrosis of peritoneal mesothelial cells induced by peritoneal dialysis via targeting VEGFA. J. Cell. Physiol. 2019, 234, 9746–9755. [Google Scholar] [CrossRef]
- He, Q.; Wen, L.; Wang, L.; Zhang, Y.; Yu, W.; Zhang, F.; Zhang, W.; Xiao, J.; Wen, X.; Zhao, Z. miR-15a-5p suppresses peritoneal fibrosis induced by peritoneal dialysis via targeting VEGF in rats. Ren. Fail. 2020, 42, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.C.; Moonen, J.R.; Brinker, M.G.; Krenning, G. FGF2 inhibits endothelial-mesenchymal transition through microRNA-20a-mediated repression of canonical TGF-beta signaling. J. Cell Sci. 2016, 129, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, Y.; Yang, L.Y.; Zhao, D. MicroRNA-20a-5p Ameliorates Non-alcoholic Fatty Liver Disease via Inhibiting the Expression of CD36. Front. Cell Dev. Biol. 2020, 8, 596329. [Google Scholar] [CrossRef]
- Roy, S.; Khanna, S.; Hussain, S.R.; Biswas, S.; Azad, A.; Rink, C.; Gnyawali, S.; Shilo, S.; Nuovo, G.J.; Sen, C.K. MicroRNA expression in response to murine myocardial infarction: miR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovasc. Res. 2009, 82, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, K.C.; Wu, W.; Subramaniam, S.; Shyy, J.Y.; Chiu, J.J.; Li, J.Y.; Chien, S. MicroRNA-21 targets peroxisome proliferators-activated receptor-alpha in an autoregulatory loop to modulate flow-induced endothelial inflammation. Proc. Natl. Acad. Sci. USA 2011, 108, 10355–10360. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chen, H.; Ge, D.; Xu, Y.; Xu, H.; Yang, Y.; Gu, M.; Zhou, Y.; Zhu, J.; Ge, T.; et al. Mir-21 Promotes Cardiac Fibrosis After Myocardial Infarction Via Targeting Smad7. Cell. Physiol. Biochem. 2017, 42, 2207–2219. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gallagher, R.; Ufret-Vincenty, R.; Li, X.; Olson, E.N.; Wang, S. Regulation of angiogenesis and choroidal neovascularization by members of microRNA-23~27~24 clusters. Proc. Natl. Acad. Sci. USA 2011, 108, 8287–8292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghiman, T.; Barghchi, B.; Esmaeili, S.A.; Shabestari, M.M.; Tabaee, S.S.; Momtazi-Borojeni, A.A. Therapeutic angiogenesis with exosomal microRNAs: An effectual approach for the treatment of myocardial ischemia. Heart Fail. Rev. 2021, 26, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Zhong, Q.Q.; Long, T.Y.; Pan, W.; Qian, Z.X. Downregulated miRNA-26a-5p induces the apoptosis of endothelial cells in coronary heart disease by inhibiting PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4940–4947. [Google Scholar] [CrossRef]
- Chai, Z.T.; Kong, J.; Zhu, X.D.; Zhang, Y.Y.; Lu, L.; Zhou, J.M.; Wang, L.R.; Zhang, K.Z.; Zhang, Q.B.; Ao, J.Y.; et al. MicroRNA-26a inhibits angiogenesis by down-regulating VEGFA through the PIK3C2alpha/Akt/HIF-1alpha pathway in hepatocellular carcinoma. PLoS ONE 2013, 8, e77957. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cai, H.; Li, H.; Gao, Z.; Song, K. Atrial overexpression of microRNA-27b attenuates angiotensin II-induced atrial fibrosis and fibrillation by targeting ALK5. Hum. Cell 2018, 31, 251–260. [Google Scholar] [CrossRef]
- Urbich, C.; Kaluza, D.; Fromel, T.; Knau, A.; Bennewitz, K.; Boon, R.A.; Bonauer, A.; Doebele, C.; Boeckel, J.N.; Hergenreider, E.; et al. MicroRNA-27a/b controls endothelial cell repulsion and angiogenesis by targeting semaphorin 6A. Blood 2012, 119, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.D.; Shen, Y.; Wei, X.; Zhang, F.Q.; Liu, Y.Y.; Ma, L. Inhibitory effect of microRNA-27b on interleukin 17 (IL-17)-induced monocyte chemoattractant protein-1 (MCP1) expression. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Lei, S.; Chen, G.; Deng, L.; He, J. Upregulation of miR-27b Facilitates Apoptosis of TNF-alpha-Stimulated Fibroblast-Like Synoviocytes. Yonsei Med. J. 2019, 60, 585–591. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, L.; Zhu, G.; Pei, Z.; Zhang, W. Downregulated microRNA-27b attenuates lipopolysaccharide-induced acute lung injury via activation of NF-E2-related factor 2 and inhibition of nuclear factor kappaB signaling pathway. J. Cell. Physiol. 2019, 234, 6023–6032. [Google Scholar] [CrossRef]
- Maurer, B.; Stanczyk, J.; Jungel, A.; Akhmetshina, A.; Trenkmann, M.; Brock, M.; Kowal-Bielecka, O.; Gay, R.E.; Michel, B.A.; Distler, J.H.; et al. MicroRNA-29, a key regulator of collagen expression in systemic sclerosis. Arthritis Rheumatol. 2010, 62, 1733–1743. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, R.; Yue, Q.; Peng, H. MicroRNA-29 regulates myocardial microvascular endothelial cells proliferation and migration in association with IGF1 in type 2 diabetes. Biochem. Biophys. Res. Commun. 2017, 487, 15–21. [Google Scholar] [CrossRef]
- Duisters, R.F.; Tijsen, A.J.; Schroen, B.; Leenders, J.J.; Lentink, V.; van der Made, I.; Herias, V.; van Leeuwen, R.E.; Schellings, M.W.; Barenbrug, P.; et al. miR-133 and miR-30 regulate connective tissue growth factor: Implications for a role of microRNAs in myocardial matrix remodeling. Circ. Res. 2009, 104, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Demolli, S.; Doebele, C.; Doddaballapur, A.; Lang, V.; Fisslthaler, B.; Chavakis, E.; Vinciguerra, M.; Sciacca, S.; Henschler, R.; Hecker, M.; et al. MicroRNA-30 mediates anti-inflammatory effects of shear stress and KLF2 via repression of angiopoietin 2. J. Mol. Cell. Cardiol. 2015, 88, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Zhong, Y.; Cheng, C.; Liu, B.; Wang, L.; Li, A.; Xiong, L.; Liu, S. MiR-30-regulated autophagy mediates angiotensin II-induced myocardial hypertrophy. PLoS ONE 2013, 8, e53950. [Google Scholar] [CrossRef]
- Bridge, G.; Monteiro, R.; Henderson, S.; Emuss, V.; Lagos, D.; Georgopoulou, D.; Patient, R.; Boshoff, C. The microRNA-30 family targets DLL4 to modulate endothelial cell behavior during angiogenesis. Blood 2012, 120, 5063–5072. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.T.; Li, X.X.; Cheng, Q.J.; Wang, Y.H.; Wang, J.H.; Liu, C.L. MiR-30a regulates the atrial fibrillation-induced myocardial fibrosis by targeting snail 1. Int. J. Clin. Exp. Pathol. 2015, 8, 15527–15536. [Google Scholar] [PubMed]
- Maciejak, A.; Kostarska-Srokosz, E.; Gierlak, W.; Dluzniewski, M.; Kuch, M.; Marchel, M.; Opolski, G.; Kiliszek, M.; Matlak, K.; Dobrzycki, S.; et al. Circulating miR-30a-5p as a prognostic biomarker of left ventricular dysfunction after acute myocardial infarction. Sci. Rep. 2018, 8, 9883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sygitowicz, G.; Maciejak-Jastrzębska, A.; Sitkiewicz, D. MicroRNAs in the development of left ventricular remodeling and postmyocardial infarction heart failure. Pol. Arch. Intern. Med. 2020, 130, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Suarez, Y.; Wang, C.; Manes, T.D.; Pober, J.S. Cutting edge: TNF-induced microRNAs regulate TNF-induced expression of E-selectin and intercellular adhesion molecule-1 on human endothelial cells: Feedback control of inflammation. J. Immunol. 2010, 184, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Men, M.; Yang, W.; Zheng, H.; Xue, S. MiR-31 Downregulation Protects Against Cardiac Ischemia/Reperfusion Injury by Targeting Protein Kinase C Epsilon (PKCepsilon) Directly. Cell. Physiol. Biochem. 2015, 36, 179–190. [Google Scholar] [CrossRef]
- Fang, Y.; Davies, P.F. Site-specific microRNA-92a regulation of Kruppel-like factors 4 and 2 in atherosusceptible endothelium. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yuan, L.; Su, L.; Lian, Z.; Liu, C.; Zhang, F.; Cui, Y.; Wu, M.; Chen, H. Decreased miR-92a-3p expression potentially mediates the pro-angiogenic effects of oxidative stress-activated endothelial cell-derived exosomes by targeting tissue factor. Int. J. Mol. Med. 2020, 46, 1886–1898. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Hoelscher, M.; Cattelan, A.; Schmitz, T.; Proebsting, S.; Wenzel, D.; Vosen, S.; Franklin, B.S.; Fleischmann, B.K.; et al. Endothelial microparticle-mediated transfer of MicroRNA-126 promotes vascular endothelial cell repair via SPRED1 and is abrogated in glucose-damaged endothelial microparticles. Circulation 2013, 128, 2026–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Schober, A.; Weber, C. Pathogenic arterial remodeling: The good and bad of microRNAs. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1050–H1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, H.Y.; Han, Z.J.; Tian, P.; Sun, W.J.; Xue, D.X.; Bi, Y.; Yang, Z.H.; Liu, P. VEGFA Expression Is Inhibited by Arsenic Trioxide in HUVECs through the Upregulation of Ets-2 and miRNA-126. PLoS ONE 2015, 10, e0135795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Lo, W.Y.; Wang, S.J.; Wang, H.J. Non-canonical Interaction Between O-Linked N-Acetylglucosamine Transferase and miR-146a-5p Aggravates High Glucose-Induced Endothelial Inflammation. Front. Physiol. 2020, 11, 1091. [Google Scholar] [CrossRef]
- Zhang, Q.; Cai, R.; Tang, G.; Zhang, W.; Pang, W. MiR-146a-5p targeting SMAD4 and TRAF6 inhibits adipogenensis through TGF-β and AKT/mTORC1 signal pathways in porcine intramuscular preadipocytes. J. Anim. Sci. Biotechnol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, Q.; Du, Z.; Huang, M.; Zhang, M.; Tan, H. MiR-148b-3p inhibits renal carcinoma cell growth and pro-angiogenic phenotype of endothelial cell potentially by modulating FGF2. Biomed. Pharmacother. 2018, 107, 359–367. [Google Scholar] [CrossRef]
- Tian, L.; Wu, D.; Dasgupta, A.; Chen, K.H.; Mewburn, J.; Potus, F.; Lima, P.D.A.; Hong, Z.; Zhao, Y.Y.; Hindmarch, C.C.T.; et al. Epigenetic Metabolic Reprogramming of Right Ventricular Fibroblasts in Pulmonary Arterial Hypertension: A Pyruvate Dehydrogenase Kinase-Dependent Shift in Mitochondrial Metabolism Promotes Right Ventricular Fibrosis. Circ. Res. 2020, 126, 1723–1745. [Google Scholar] [CrossRef] [PubMed]
- Shan, G.; Zhou, X.; Gu, J.; Zhou, D.; Cheng, W.; Wu, H.; Wang, Y.; Tang, T.; Wang, X. Downregulated exosomal microRNA-148b-3p in cancer associated fibroblasts enhance chemosensitivity of bladder cancer cells by downregulating the Wnt/β-catenin pathway and upregulating PTEN. Cell. Oncol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xu, Y.; Deng, Y.; Li, H. MicroRNA-223 Regulates Cardiac Fibrosis After Myocardial Infarction by Targeting RASA1. Cell. Physiol. Biochem. 2018, 46, 1439–1454. [Google Scholar] [CrossRef]
- Shao, J.; Fan, G.; Yin, X.; Gu, Y.; Wang, X.; Xin, Y.; Yao, Y. A novel transthyretin/STAT4/miR-223-3p/FBXW7 signaling pathway affects neovascularization in diabetic retinopathy. Mol. Cell. Endocrinol. 2019, 498, 110541. [Google Scholar] [CrossRef]
- Cheng, S.; Cui, Y.; Fan, L.; Mu, X.; Hua, Y. T2DM inhibition of endothelial miR-342-3p facilitates angiogenic dysfunction via repression of FGF11 signaling. Biochem. Biophys. Res. Commun. 2018, 503, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhao, L.; Zhu, L.Y.; He, M.; Zheng, L.; Wu, Y. MicroRNA-17, 20a regulates the proangiogenic function of tumor-associated macrophages via targeting hypoxia-inducible factor 2alpha. PLoS ONE 2013, 8, e77890. [Google Scholar] [CrossRef]
- Corsetti, P.P.; de Almeida, L.A.; Goncalves, A.N.A.; Gomes, M.T.R.; Guimaraes, E.S.; Marques, J.T.; Oliveira, S.C. miR-181a-5p Regulates TNF-alpha and miR-21a-5p Influences Gualynate-Binding Protein 5 and IL-10 Expression in Macrophages Affecting Host Control of Brucella abortus Infection. Front. Immunol. 2018, 9, 1331. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ge, J.; Li, W.; Zhou, W.; Xu, L. LncRNA KCNQ1OT1 ameliorates particle-induced osteolysis through inducing macrophage polarization by inhibiting miR-21a-5p. Biol. Chem. 2018, 399, 375–386. [Google Scholar] [CrossRef]
- Ma, S.; Liu, M.; Xu, Z.; Li, Y.; Guo, H.; Ge, Y.; Liu, Y.; Zheng, D.; Shi, J. A double feedback loop mediated by microRNA-23a/27a/24-2 regulates M1 versus M2 macrophage polarization and thus regulates cancer progression. Oncotarget 2016, 7, 13502–13519. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Han, T.; Li, R.; Fu, L.; Yue, L. miR-26a-5p mediates TLR signaling pathway by targeting CTGF in LPS-induced alveolar macrophage. BioSci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Li, W.; Chang, N.; Tian, L.; Yang, J.; Ji, X.; Xie, J.; Yang, L.; Li, L. miR-27b-3p, miR-181a-1-3p, and miR-326-5p are involved in the inhibition of macrophage activation in chronic liver injury. J. Mol. Med. 2017, 95, 1091–1105. [Google Scholar] [CrossRef]
- Domingo-Gonzalez, R.; Wilke, C.A.; Huang, S.K.; Laouar, Y.; Brown, J.P.; Freeman, C.M.; Curtis, J.L.; Yanik, G.A.; Moore, B.B. Transforming growth factor-beta induces microRNA-29b to promote murine alveolar macrophage dysfunction after bone marrow transplantation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L86–L95. [Google Scholar] [CrossRef] [Green Version]
- Shangxun, Z.; Junjie, L.; Wei, Z.; Yutong, W.; Wenyuan, J.; Shanshou, L.; Yanjun, W.; Qianmei, W.; Zhusheng, F.; Chaoping, Y.; et al. ADAR1 Alleviates Inflammation in a Murine Sepsis Model via the ADAR1-miR-30a-SOCS3 Axis. Mediat. Inflamm. 2020, 2020, 9607535. [Google Scholar] [CrossRef]
- Jiang, W.L.; Zhao, K.C.; Yuan, W.; Zhou, F.; Song, H.Y.; Liu, G.L.; Huang, J.; Zou, J.J.; Zhao, B.; Xie, S.P. MicroRNA-31-5p Exacerbates Lipopolysaccharide-Induced Acute Lung Injury via Inactivating Cab39/AMPKalpha Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 8822361. [Google Scholar] [CrossRef]
- Fu, L.; Zhu, P.; Qi, S.; Li, C.; Zhao, K. MicroRNA-92a antagonism attenuates lipopolysaccharide (LPS)-induced pulmonary inflammation and injury in mice through suppressing the PTEN/AKT/NF-kappaB signaling pathway. Biomed. Pharmacother. 2018, 107, 703–711. [Google Scholar] [CrossRef]
- Chang, Y.J.; Li, Y.S.; Wu, C.C.; Wang, K.C.; Huang, T.C.; Chen, Z.; Chien, S. Extracellular MicroRNA-92a Mediates Endothelial Cell-Macrophage Communication. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 2492–2504. [Google Scholar] [CrossRef]
- Laffont, B.; Corduan, A.; Rousseau, M.; Duchez, A.C.; Lee, C.H.; Boilard, E.; Provost, P. Platelet microparticles reprogram macrophage gene expression and function. Thromb. Haemost. 2016, 115, 311–323. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, M.; Zhang, X.; Kuai, Y.; Mei, Y.; Tan, Q.; Zhong, K.; Sun, X.; Tan, W. Isosteviol Sodium Protects against Ischemic Stroke by Modulating Microglia/Macrophage Polarization via Disruption of GAS5/miR-146a-5p sponge. Sci. Rep. 2019, 9, 12221. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Brown, M.E.; Zhang, H.; Martinez, M.; Zhao, Z.; Bhutani, S.; Yin, S.; Trac, D.; Xi, J.J.; Davis, M.E. High-throughput screening identifies microRNAs that target Nox2 and improve function after acute myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1002–H1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, G.; Meng, C.; Guo, X.; Cheruku, P.S.; Shi, L.; Xu, H.; Li, H.; Wang, G.; Evans, A.R.; Safe, S.; et al. A novel regulator of macrophage activation: miR-223 in obesity-associated adipose tissue inflammation. Circulation 2012, 125, 2892–2903. [Google Scholar] [CrossRef] [Green Version]
- Ying, W.; Tseng, A.; Chang, R.C.; Morin, A.; Brehm, T.; Triff, K.; Nair, V.; Zhuang, G.; Song, H.; Kanameni, S.; et al. MicroRNA-223 is a crucial mediator of PPARgamma-regulated alternative macrophage activation. J. Clin. Investig. 2015, 125, 4149–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez Calvente, C.; Del Pilar, H.; Tameda, M.; Johnson, C.D.; Feldstein, A.E. MicroRNA 223 3p Negatively Regulates the NLRP3 Inflammasome in Acute and Chronic Liver Injury. Mol. Ther. 2020, 28, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xia, J.W.; Ke, Z.P.; Zhang, B.H. Blockade of NEAT1 represses inflammation response and lipid uptake via modulating miR-342-3p in human macrophages THP-1 cells. J. Cell. Physiol. 2019, 234, 5319–5326. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niderla-Bielińska, J.; Ścieżyńska, A.; Moskalik, A.; Jankowska-Steifer, E.; Bartkowiak, K.; Bartkowiak, M.; Kiernozek, E.; Podgórska, A.; Ciszek, B.; Majchrzak, B.; et al. A Comprehensive miRNome Analysis of Macrophages Isolated from db/db Mice and Selected miRNAs Involved in Metabolic Syndrome-Associated Cardiac Remodeling. Int. J. Mol. Sci. 2021, 22, 2197. https://doi.org/10.3390/ijms22042197
Niderla-Bielińska J, Ścieżyńska A, Moskalik A, Jankowska-Steifer E, Bartkowiak K, Bartkowiak M, Kiernozek E, Podgórska A, Ciszek B, Majchrzak B, et al. A Comprehensive miRNome Analysis of Macrophages Isolated from db/db Mice and Selected miRNAs Involved in Metabolic Syndrome-Associated Cardiac Remodeling. International Journal of Molecular Sciences. 2021; 22(4):2197. https://doi.org/10.3390/ijms22042197
Chicago/Turabian StyleNiderla-Bielińska, Justyna, Aneta Ścieżyńska, Aneta Moskalik, Ewa Jankowska-Steifer, Krzysztof Bartkowiak, Mateusz Bartkowiak, Ewelina Kiernozek, Anna Podgórska, Bogdan Ciszek, Barbara Majchrzak, and et al. 2021. "A Comprehensive miRNome Analysis of Macrophages Isolated from db/db Mice and Selected miRNAs Involved in Metabolic Syndrome-Associated Cardiac Remodeling" International Journal of Molecular Sciences 22, no. 4: 2197. https://doi.org/10.3390/ijms22042197
APA StyleNiderla-Bielińska, J., Ścieżyńska, A., Moskalik, A., Jankowska-Steifer, E., Bartkowiak, K., Bartkowiak, M., Kiernozek, E., Podgórska, A., Ciszek, B., Majchrzak, B., & Ratajska, A. (2021). A Comprehensive miRNome Analysis of Macrophages Isolated from db/db Mice and Selected miRNAs Involved in Metabolic Syndrome-Associated Cardiac Remodeling. International Journal of Molecular Sciences, 22(4), 2197. https://doi.org/10.3390/ijms22042197