
Myosin Phosphatase Is Implicated in the Control of THP-1 Monocyte to Macrophage Differentiation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
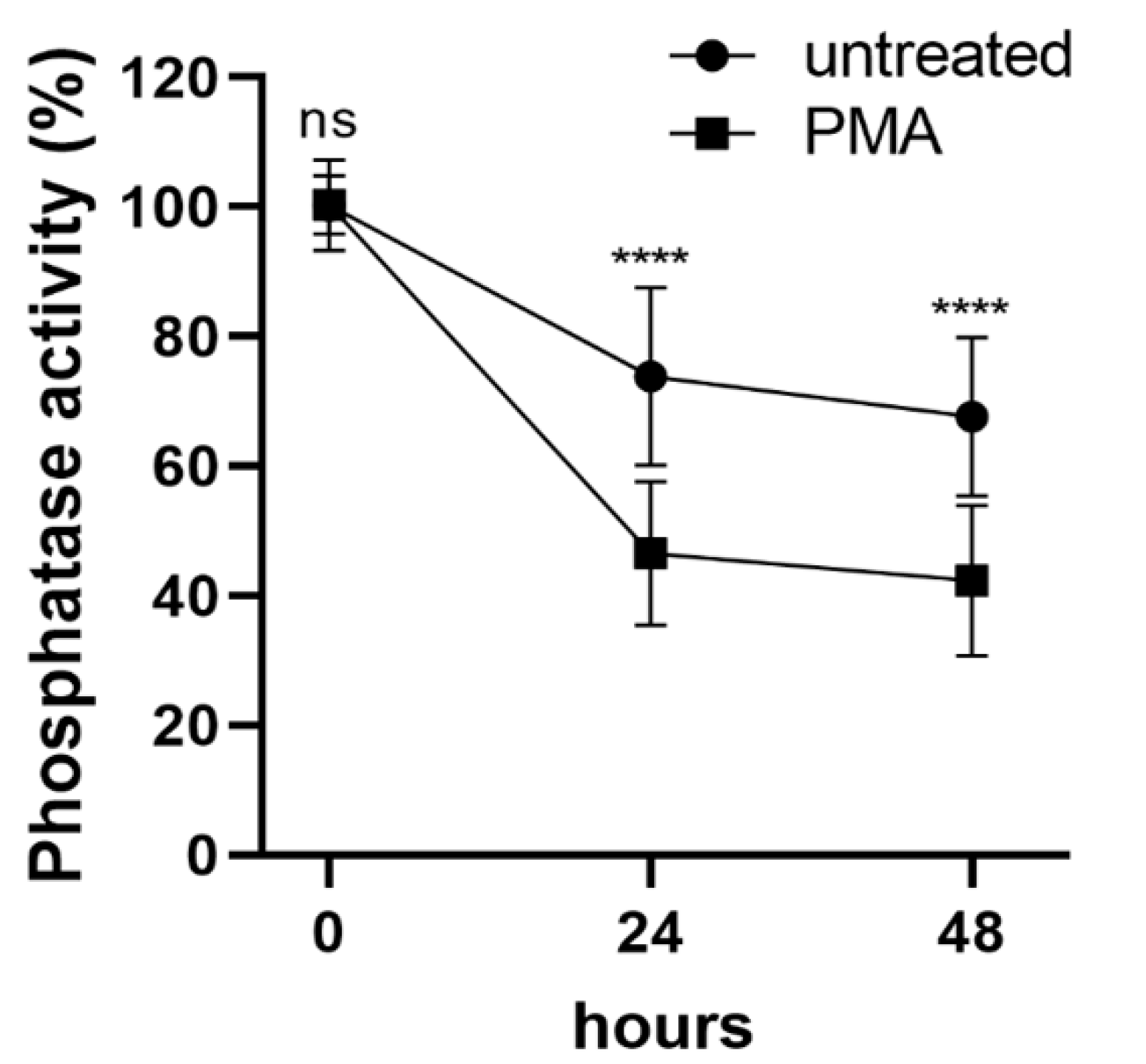
2.1. Differentiation of THP-1 Cells Induced by PMA Is Accompanied by Phosphatase Inhibition
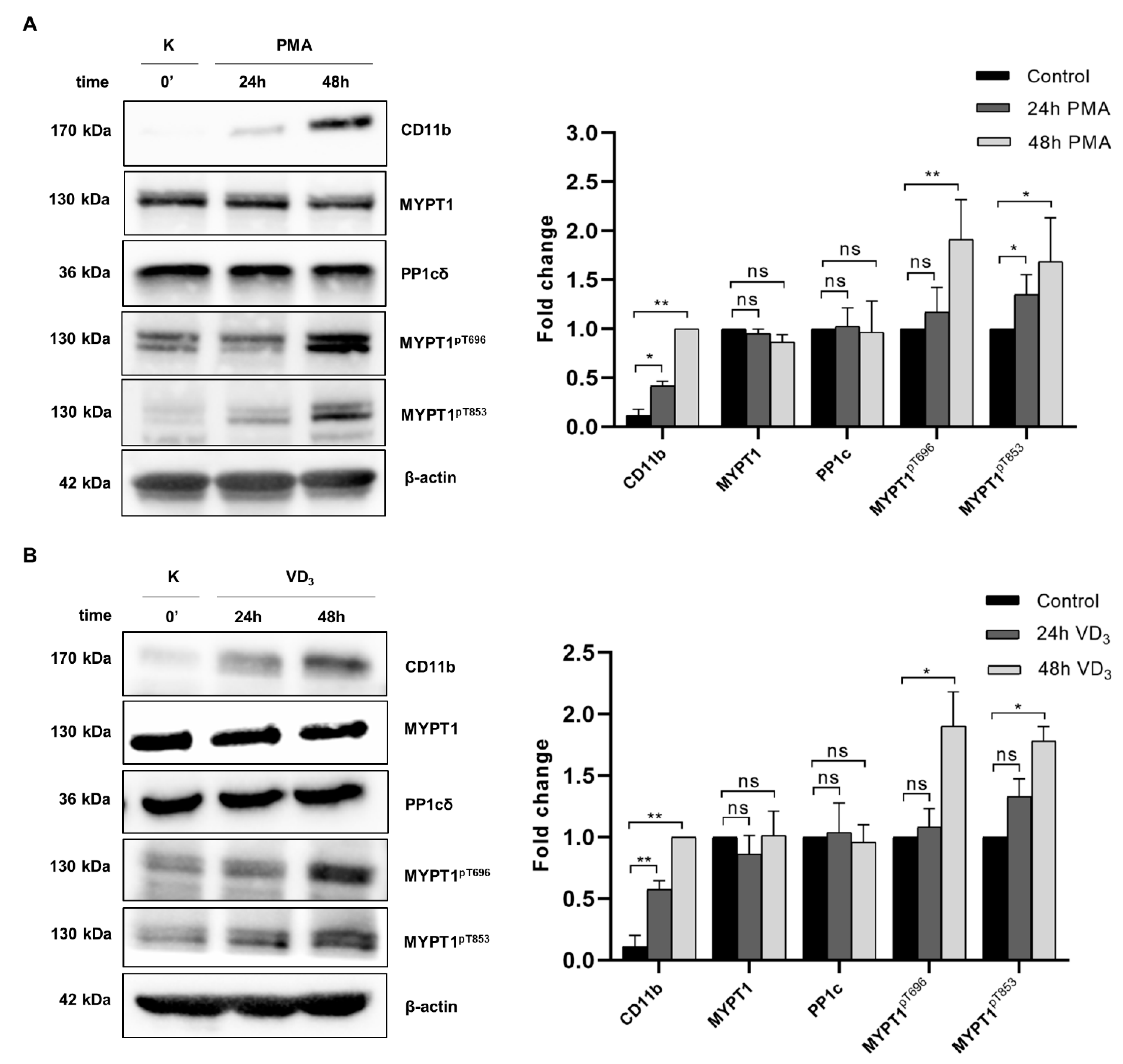
2.2. Myosin Phosphatase Is Inhibited during PMA- and VD3-Induced Differentiation of THP-1 Cells
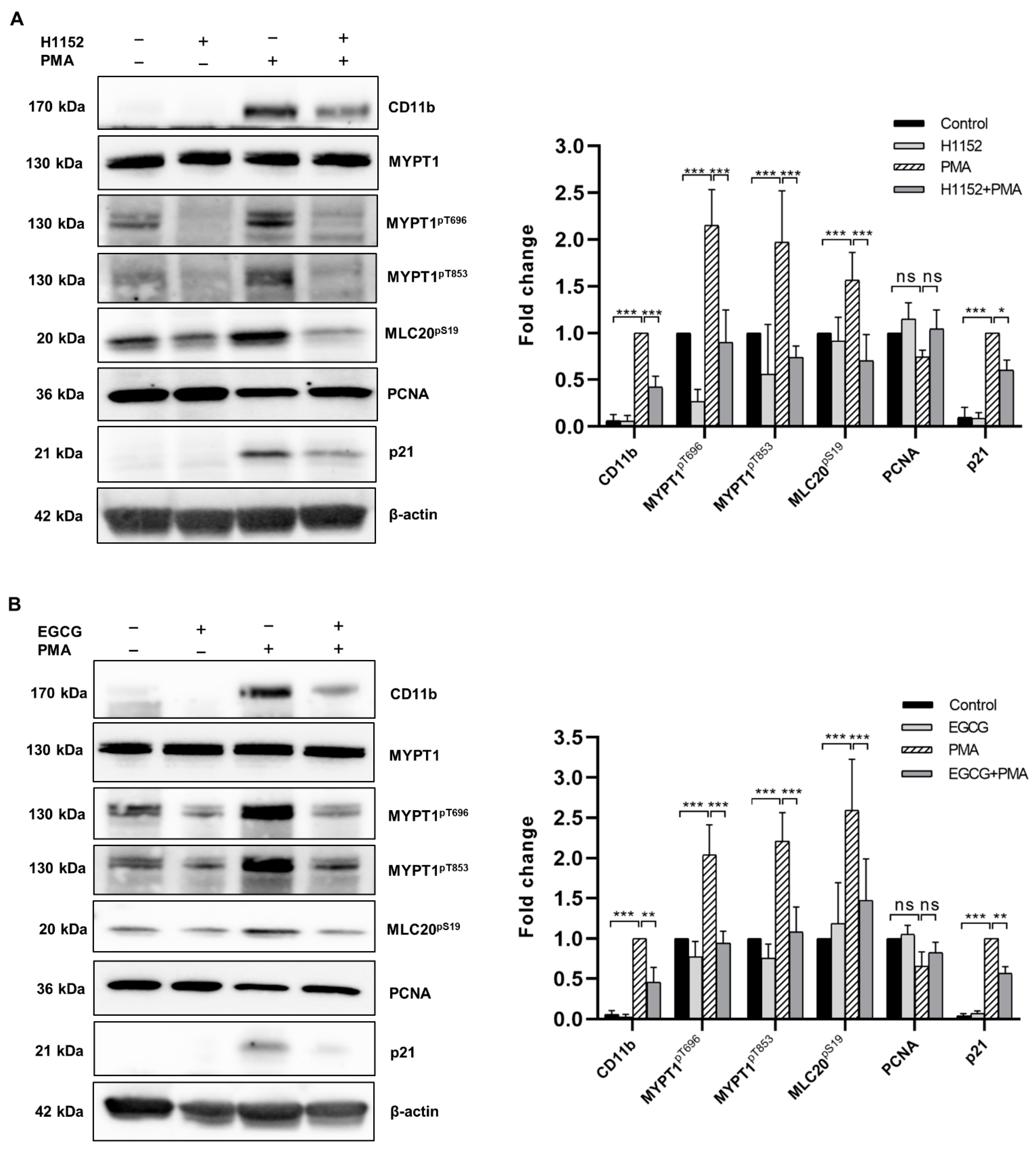
2.3. Both H1152 and EGCG Pretreatment Activates Myosin Phosphatase and Attenuates PMA-Induced Macrophage Differentiation
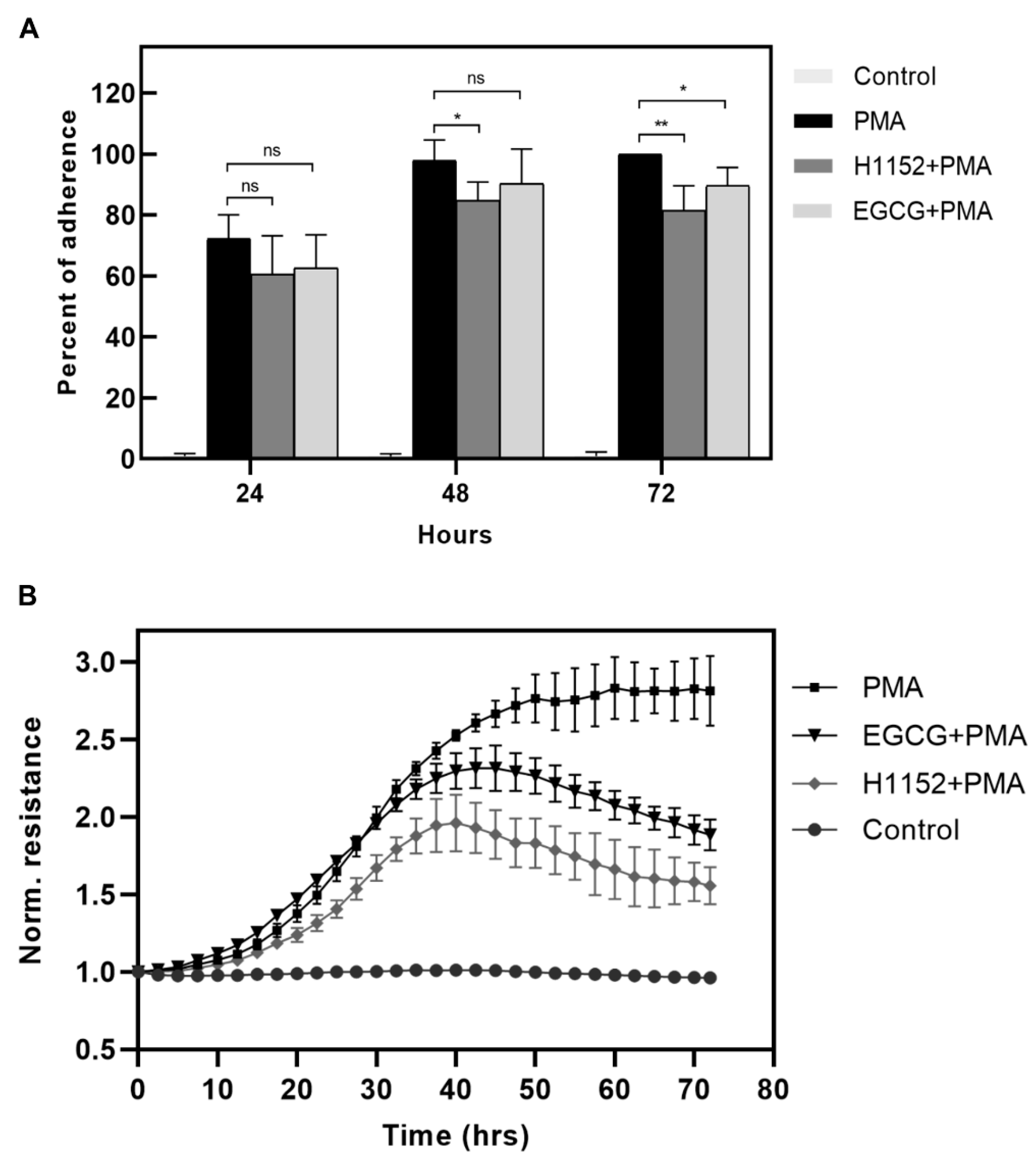
2.4. Activation of Myosin Phosphatase Decreases the Strength of Adherence of Differentiating Macrophages
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture and Treatments
4.3. Phosphatase Activity Assay
4.4. Western Blot Analysis
4.5. Sulforhodamine B Assay
4.6. Electric Cell-Substrate Impedance Sensing (ECIS) Measurement
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coillard, A.; Segura, E. Differentiation of Human Monocytes. Front. Immunol. 2019, 10, 1907. [Google Scholar] [CrossRef]
- Karlmark, K.R.; Tacke, F.; Dunay, I.R. Monocytes in health and disease—Minireview. Eur. J. Microbiol. Immunol. 2012, 2, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, K.; Miller, Y.I.; Hedrick, C.C. Monocyte and macrophage dynamics during atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1506–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, S.; Kobayashi, Y.; Goto, Y.; Okumura, H.; Nakae, S.; Konno, T.; Tada, K. Induction of maturation in cultured human monocytic leukemia cells by a phorbol diester. Cancer Res. 1982, 42, 1530–1536. [Google Scholar] [PubMed]
- Akashi, M.; Osawa, Y.; Koeffler, H.P.; Hachiya, M. p21WAF1 expression by an activator of protein kinase C is regulated mainly at the post-transcriptional level in cells lacking p53: Important role of RNA stabilization. Biochem J. 1999, 337 Pt 3, 607–616. [Google Scholar] [CrossRef]
- Traore, K.; Trush, M.A.; George, M.; Spannhake, E.W.; Anderson, W.; Asseffa, A. Signal transduction of phorbol 12-myristate 13-acetate (PMA)-induced growth inhibition of human monocytic leukemia THP-1 cells is reactive oxygen dependent. Leuk. Res. 2005, 29, 863–879. [Google Scholar] [CrossRef]
- Schwende, H.; Fitzke, E.; Ambs, P.; Dieter, P. Differences in the state of differentiation of THP-1 cells induced by phorbol ester and 1,25-dihydroxyvitamin D3. J. Leukoc. Biol. 1996, 59, 555–561. [Google Scholar] [CrossRef]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef]
- Hughes, P.J.; Marcinkowska, E.; Gocek, E.; Studzinski, G.P.; Brown, G. Vitamin D3-driven signals for myeloid cell differentiation--implications for differentiation therapy. Leuk. Res. 2010, 34, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Wang, W.; Yu, X.; Yang, L.; Chen, S.; Li, Y. Pathways related to PMA-differentiated THP1 human monocytic leukemia cells revealed by RNA-Seq. Sci. China Life Sci. 2015, 58, 1282–1287. [Google Scholar] [CrossRef] [Green Version]
- Richter, E.; Ventz, K.; Harms, M.; Mostertz, J.; Hochgräfe, F. Induction of Macrophage Function in Human THP-1 Cells Is Associated with Rewiring of MAPK Signaling and Activation of MAP3K7 (TAK1) Protein Kinase. Front. Cell Dev. Biol. 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Ghosh, P.; Prasad, R.; Ghosh, A.; Das, K.; Roy, A.; Mallik, S.; Sinha, D.K.; Sen, P. MAP Kinase driven actomyosin rearrangement is a crucial regulator of monocyte to macrophage differentiation. Cell Signal. 2020, 73, 109691. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Dai, F.; Tang, L.; Le, Y.; Yao, W. Macrophage differentiation induced by PMA is mediated by activation of RhoA/ROCK signaling. J. Toxicol. Sci. 2017, 42, 763–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, A.; Erdődi, F.; Lontay, B. Myosin phosphatase: Unexpected functions of a long-known enzyme. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 2–15. [Google Scholar] [CrossRef]
- Grassie, M.E.; Moffat, L.D.; Walsh, M.P.; MacDonald, J.A. The myosin phosphatase targeting protein (MYPT) family: A regulated mechanism for achieving substrate specificity of the catalytic subunit of protein phosphatase type 1δ. Arch. Biochem. Biophys. 2011, 510, 147–159. [Google Scholar] [CrossRef]
- Totsukawa, G.; Yamakita, Y.; Yamashiro, S.; Hartshorne, D.J.; Sasaki, Y.; Matsumura, F. Distinct roles of ROCK (Rho-kinase) and MLCK in spatial regulation of MLC phosphorylation for assembly of stress fibers and focal adhesions in 3T3 fibroblasts. J. Cell Biol. 2000, 150, 797–806. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; Iwamatsu, A.; Kaibuchi, K. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef]
- Zagórska, A.; Deak, M.; Campbell, D.G.; Banerjee, S.; Hirano, M.; Aizawa, S.; Prescott, A.R.; Alessi, D.R. New roles for the LKB1-NUAK pathway in controlling myosin phosphatase complexes and cell adhesion. Sci. Signal. 2010, 3, ra25. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Stull, J.T.; Kamm, K.E. Myosin phosphatase targeting subunit 1 affects cell migration by regulating myosin phosphorylation and actin assembly. Exp. Cell Res. 2005, 304, 506–517. [Google Scholar] [CrossRef]
- Eto, M.; Wong, L.; Yazawa, M.; Brautigan, D.L. Inhibition of myosin/moesin phosphatase by expression of the phosphoinhibitor protein CPI-17 alters microfilament organization and retards cell spreading. Cell Motil. Cytoskelet. 2000, 46, 222–234. [Google Scholar] [CrossRef]
- Samson, S.C.; Elliott, A.; Mueller, B.D.; Kim, Y.; Carney, K.R.; Bergman, J.P.; Blenis, J.; Mendoza, M.C. p90 ribosomal S6 kinase (RSK) phosphorylates myosin phosphatase and thereby controls edge dynamics during cell migration. J. Biol. Chem. 2019, 294, 10846–10862. [Google Scholar] [CrossRef] [Green Version]
- Virshup, D.M.; Shenolikar, S. From promiscuity to precision: Protein phosphatases get a makeover. Mol. Cell 2009, 33, 537–545. [Google Scholar] [CrossRef]
- Fardilha, M.; Esteves, S.L.; Korrodi-Gregório, L.; da Cruz e Silva, O.A.; da Cruz e Silva, F.F. The physiological relevance of protein phosphatase 1 and its interacting proteins to health and disease. Curr. Med. Chem. 2010, 17, 3996–4017. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, R.; Velmurugan, B.K. Protein phosphatase 2A as therapeutic targets in various disease models. Life Sci. 2018, 210, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.S.; Lin, Y.W.; Huang, C.Y.; Shih, C.M.; Tsai, Y.T.; Tsao, N.W.; Lin, C.S.; Shih, C.C.; Jeng, H.; Lin, F.Y. Thrombomodulin regulates monocye differentiation via PKCδ and ERK1/2 pathway in vitro and in atherosclerotic artery. Sci. Rep. 2016, 6, 38421. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, S.; Huang, Y.; Umeda, D.; Yamada, S.; Yamashita, S.; Kumazoe, M.; Kim, Y.; Murata, M.; Yamada, K.; Tachibana, H. 67-kDa laminin receptor-dependent protein phosphatase 2A (PP2A) activation elicits melanoma-specific antitumor activity overcoming drug resistance. J. Biol. Chem. 2014, 289, 32671–32681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batori, R.; Becsi, B.; Nagy, D.; Konya, Z.; Hegedus, C.; Bordan, Z.; Verin, A.; Lontay, B.; Erdodi, F. Interplay of myosin phosphatase and protein phosphatase-2A in the regulation of endothelial nitric-oxide synthase phosphorylation and nitric oxide production. Sci. Rep. 2017, 7, 44698. [Google Scholar] [CrossRef] [Green Version]
- Tóth, E.; Erdődi, F.; Kiss, A. Activation of Myosin Phosphatase by Epigallocatechin-Gallate Sensitizes THP-1 Leukemic Cells to Daunorubicin. Anticancer Agents Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, N.; Iwaki, K.; Hashimoto, K.; Mochizuki, S.; Ogasawara, Y.; Sato, M.; Tsujioka, K.; Kajiya, F. Measurements of endothelial cell-to-cell and cell-to-substrate gaps and micromechanical properties of endothelial cells during monocyte adhesion. Proc. Natl. Acad. Sci. USA 2002, 99, 15638–15643. [Google Scholar] [CrossRef] [Green Version]
- Huber, R.; Pietsch, D.; Günther, J.; Welz, B.; Vogt, N.; Brand, K. Regulation of monocyte differentiation by specific signaling modules and associated transcription factor networks. Cell Mol. Life Sci. 2014, 71, 63–92. [Google Scholar] [CrossRef]
- Nishikawa, M.; Omay, S.B.; Nakai, K.; Kihira, H.; Kobayashi, T.; Tamura, S.; Shiku, H. Up-regulation of protein serine/threonine phosphatase type 2C during 1 alpha,25-dihydroxyvitamin D3-induced monocytic differentiation of leukemic HL-60 cells. FEBS Lett. 1995, 375, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Omay, S.B.; Nakai, K.; Kuno, T.; Shiku, H.; Nishikawa, M. 1 alpha,25-dihydroxyvitamin D3-induced upregulation of calcineurin during leukemic HL-60 cell differentiation. Blood 1996, 87, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Omay, S.B.; Ogasawara, H.; Toyoda, H.; Nakai, K.; Shima, H.; Nagao, M.; Mumby, M.C.; Shiku, H.; Nishikawa, M. Translocation of protein phosphatase 1 catalytic subunits during 1,25-dihydroxyvitamin D3-induced monocytic differentiation of HL-60 cells. Cancer Res. 1995, 55, 774–780. [Google Scholar] [PubMed]
- Smith, S.R.; Schaaf, K.; Rajabalee, N.; Wagner, F.; Duverger, A.; Kutsch, O.; Sun, J. The phosphatase PPM1A controls monocyte-to-macrophage differentiation. Sci. Rep. 2018, 8, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartshorne, D.J.; Ito, M.; Erdödi, F. Role of protein phosphatase type 1 in contractile functions: Myosin phosphatase. J. Biol. Chem. 2004, 279, 37211–37214. [Google Scholar] [CrossRef] [Green Version]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 2010, 67, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Fukata, Y.; Kimura, K.; Oshiro, N.; Saya, H.; Matsuura, Y.; Kaibuchi, K. Association of the myosin-binding subunit of myosin phosphatase and moesin: Dual regulation of moesin phosphorylation by Rho-associated kinase and myosin phosphatase. J. Cell Biol. 1998, 141, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Fukata, Y.; Matsuoka, Y.; Bennett, V.; Matsuura, Y.; Okawa, K.; Iwamatsu, A.; Kaibuchi, K. Regulation of the association of adducin with actin filaments by Rho-associated kinase (Rho-kinase) and myosin phosphatase. J. Biol. Chem. 1998, 273, 5542–5548. [Google Scholar] [CrossRef] [Green Version]
- Kawano, Y.; Fukata, Y.; Oshiro, N.; Amano, M.; Nakamura, T.; Ito, M.; Matsumura, F.; Inagaki, M.; Kaibuchi, K. Phosphorylation of myosin-binding subunit (MBS) of myosin phosphatase by Rho-kinase in vivo. J. Cell Biol. 1999, 147, 1023–1038. [Google Scholar] [CrossRef] [Green Version]
- Horváth, D.; Sipos, A.; Major, E.; Kónya, Z.; Bátori, R.; Dedinszki, D.; Szöll Si, A.; Tamás, I.; Iván, J.; Kiss, A.; Erd di, F.; Lontay, B. Myosin phosphatase accelerates cutaneous wound healing by regulating migration and differentiation of epidermal keratinocytes via Akt signaling pathway in human and murine skin. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3268–3280. [Google Scholar] [CrossRef]
- Zhou, Q.; Liao, J.K. Rho kinase: An important mediator of atherosclerosis and vascular disease. Curr. Pharm. Des. 2009, 15, 3108–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, P.V.; Liu, D. Green tea catechins and cardiovascular health: An update. Curr. Med. Chem. 2008, 15, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- Eng, Q.Y.; Thanikachalam, P.V.; Ramamurthy, S. Molecular understanding of Epigallocatechin gallate (EGCG) in cardiovascular and metabolic diseases. J. Ethnopharmacol. 2018, 210, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Wang, H.; Li, Y.F.; Xie, Z.Y.; Ma, Y.; Yan, J.J.; Gao, Y.F.; Wang, Z.M.; Wang, L.S. Inhibition of EMMPRIN and MMP-9 Expression by Epigallocatechin-3-Gallate through 67-kDa Laminin Receptor in PMA-Induced Macrophages. Cell Physiol. Biochem. 2016, 39, 2308–2319. [Google Scholar] [CrossRef]
- Lontay, B.; Serfozo, Z.; Gergely, P.; Ito, M.; Hartshorne, D.J.; Erdodi, F. Localization of myosin phosphatase target subunit 1 in rat brain and in primary cultures of neuronal cells. J. Comp. Neurol. 2004, 478, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Dedinszki, D.; Kiss, A.; Markasz, L.; Marton, A.; Toth, E.; Szekely, L.; Erdodi, F. Inhibition of protein phosphatase-1 and -2A decreases the chemosensitivity of leukemic cells to chemotherapeutic drugs. Cell Signal. 2015, 27, 363–372. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, E.; Erdődi, F.; Kiss, A. Myosin Phosphatase Is Implicated in the Control of THP-1 Monocyte to Macrophage Differentiation. Int. J. Mol. Sci. 2021, 22, 2516. https://doi.org/10.3390/ijms22052516
Tóth E, Erdődi F, Kiss A. Myosin Phosphatase Is Implicated in the Control of THP-1 Monocyte to Macrophage Differentiation. International Journal of Molecular Sciences. 2021; 22(5):2516. https://doi.org/10.3390/ijms22052516
Chicago/Turabian StyleTóth, Emese, Ferenc Erdődi, and Andrea Kiss. 2021. "Myosin Phosphatase Is Implicated in the Control of THP-1 Monocyte to Macrophage Differentiation" International Journal of Molecular Sciences 22, no. 5: 2516. https://doi.org/10.3390/ijms22052516
APA StyleTóth, E., Erdődi, F., & Kiss, A. (2021). Myosin Phosphatase Is Implicated in the Control of THP-1 Monocyte to Macrophage Differentiation. International Journal of Molecular Sciences, 22(5), 2516. https://doi.org/10.3390/ijms22052516