
Nicotinamide Mononucleotide Potentiates Resistance to Biotrophic Invasion of Fungal Pathogens in Barley

 , ,
, ,  and
and 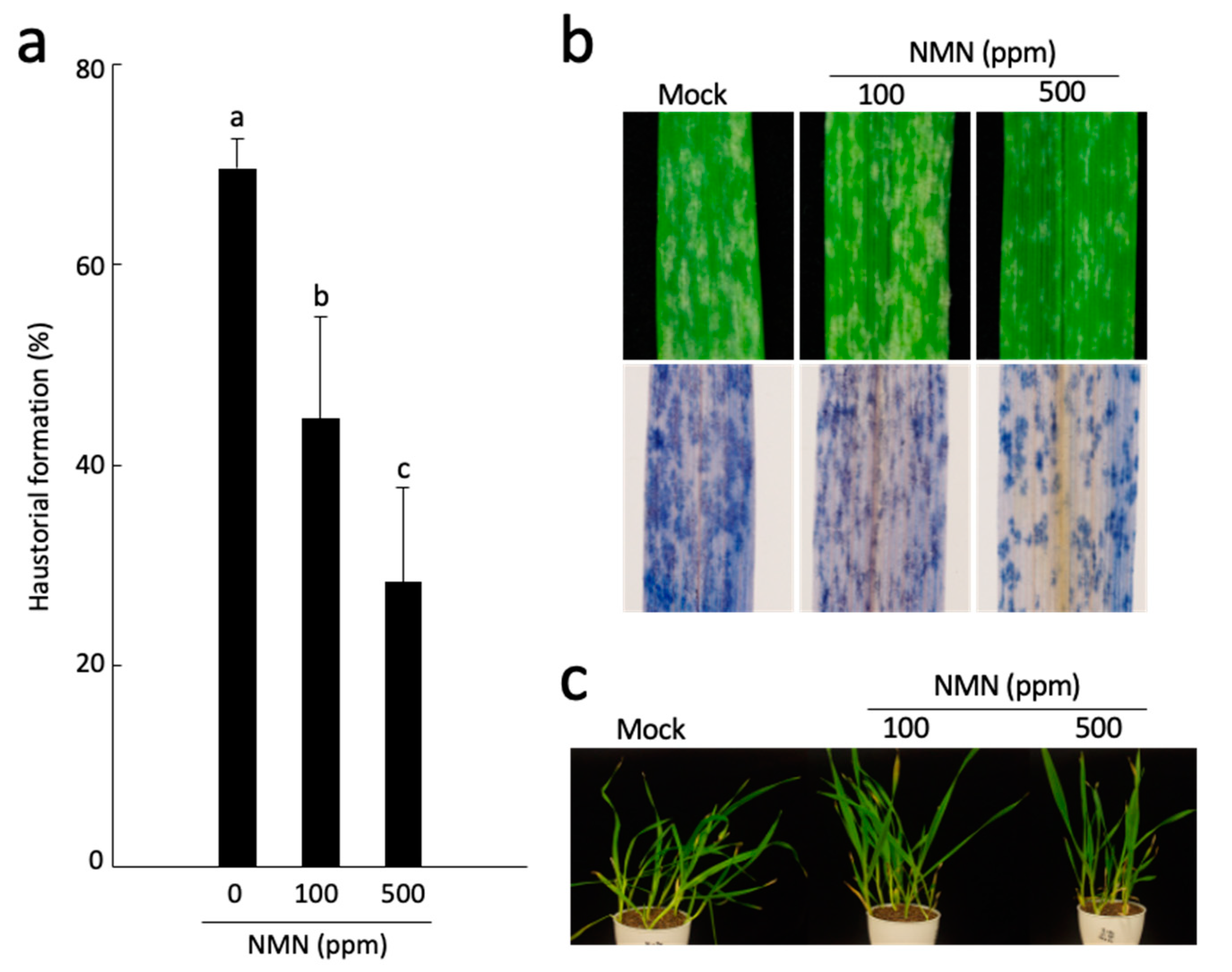{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. NMN Induces Penetration Resistance to Bgh
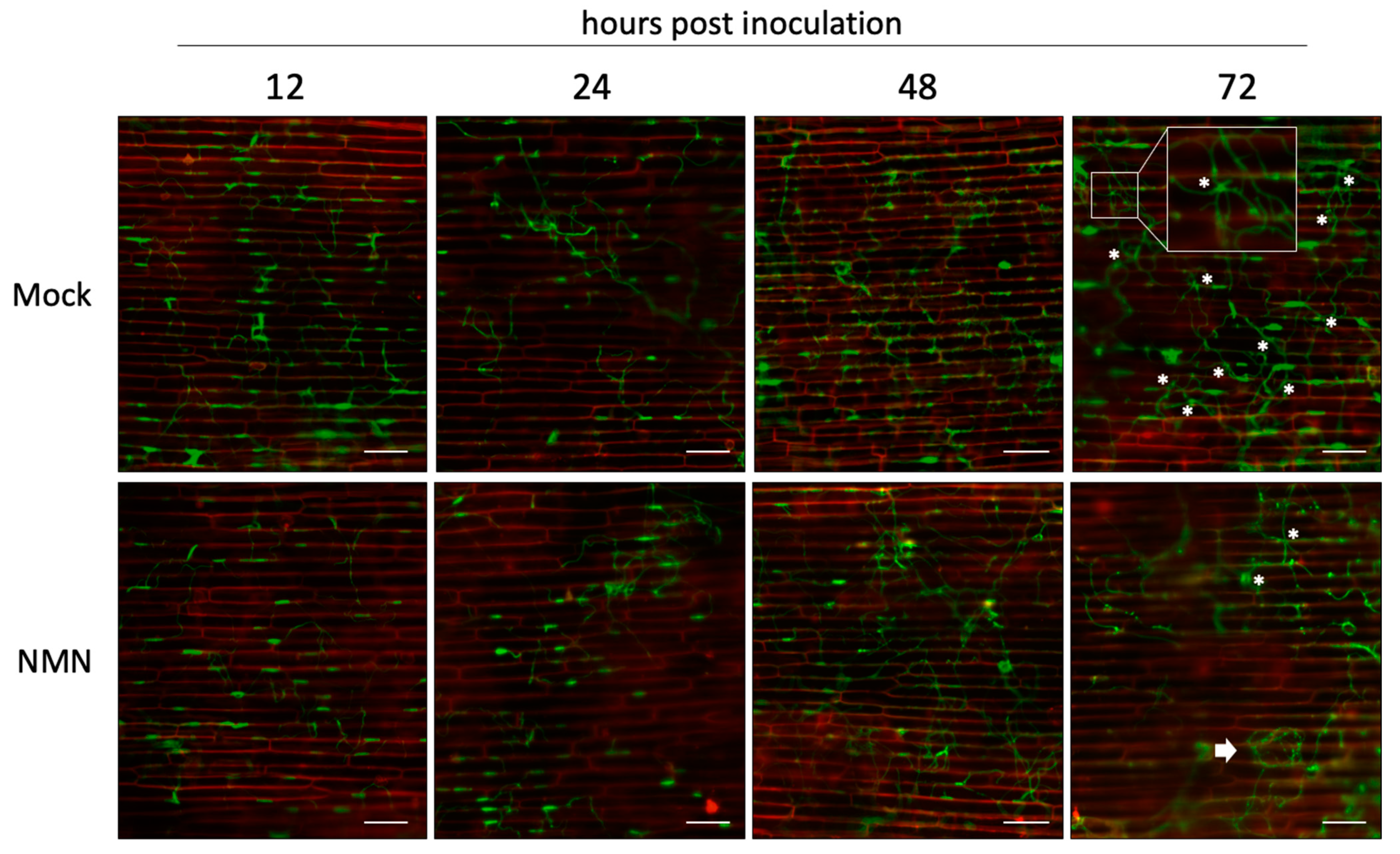
2.2. NMN Interferes with the Biotrophic Process of F. graminearum Infection
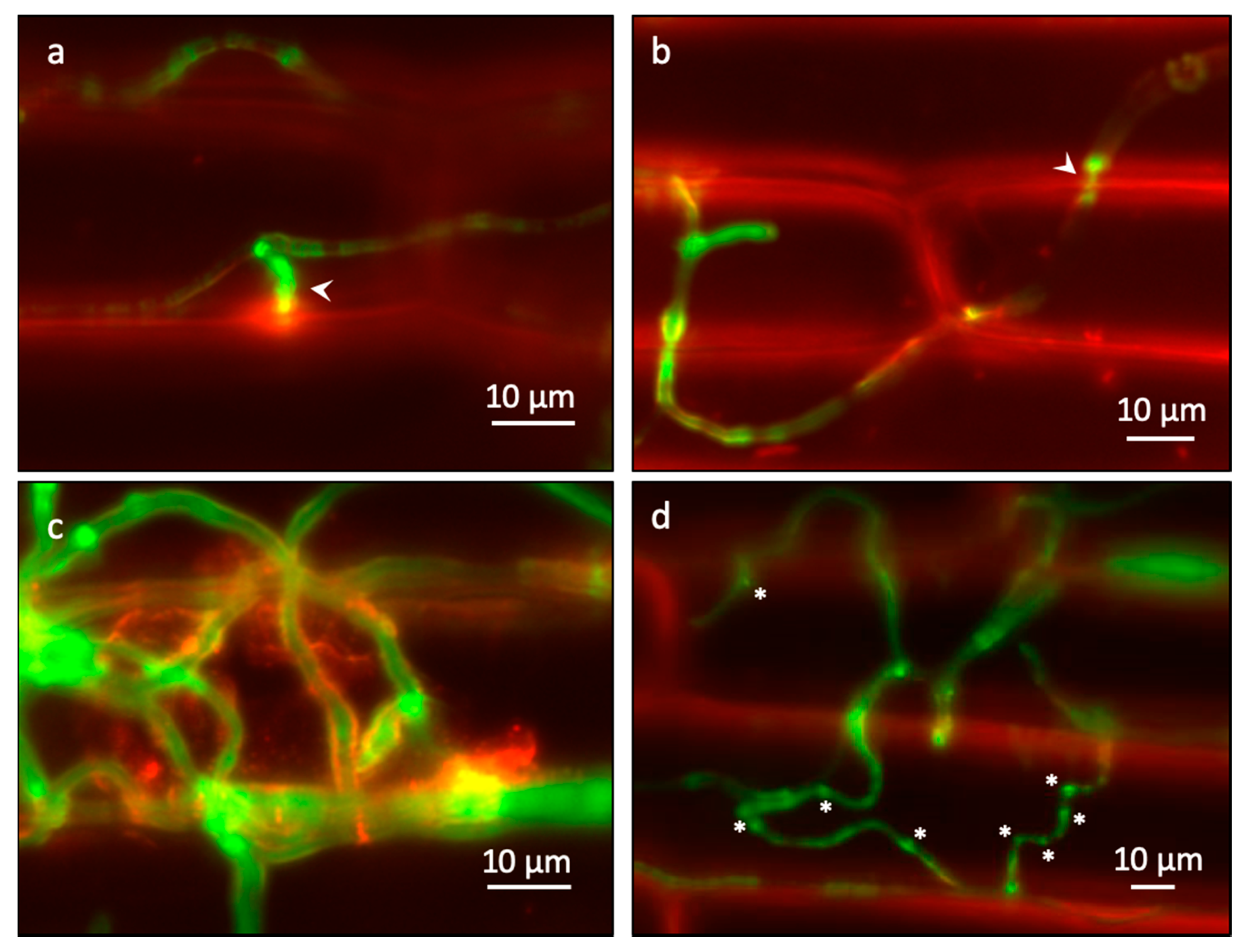
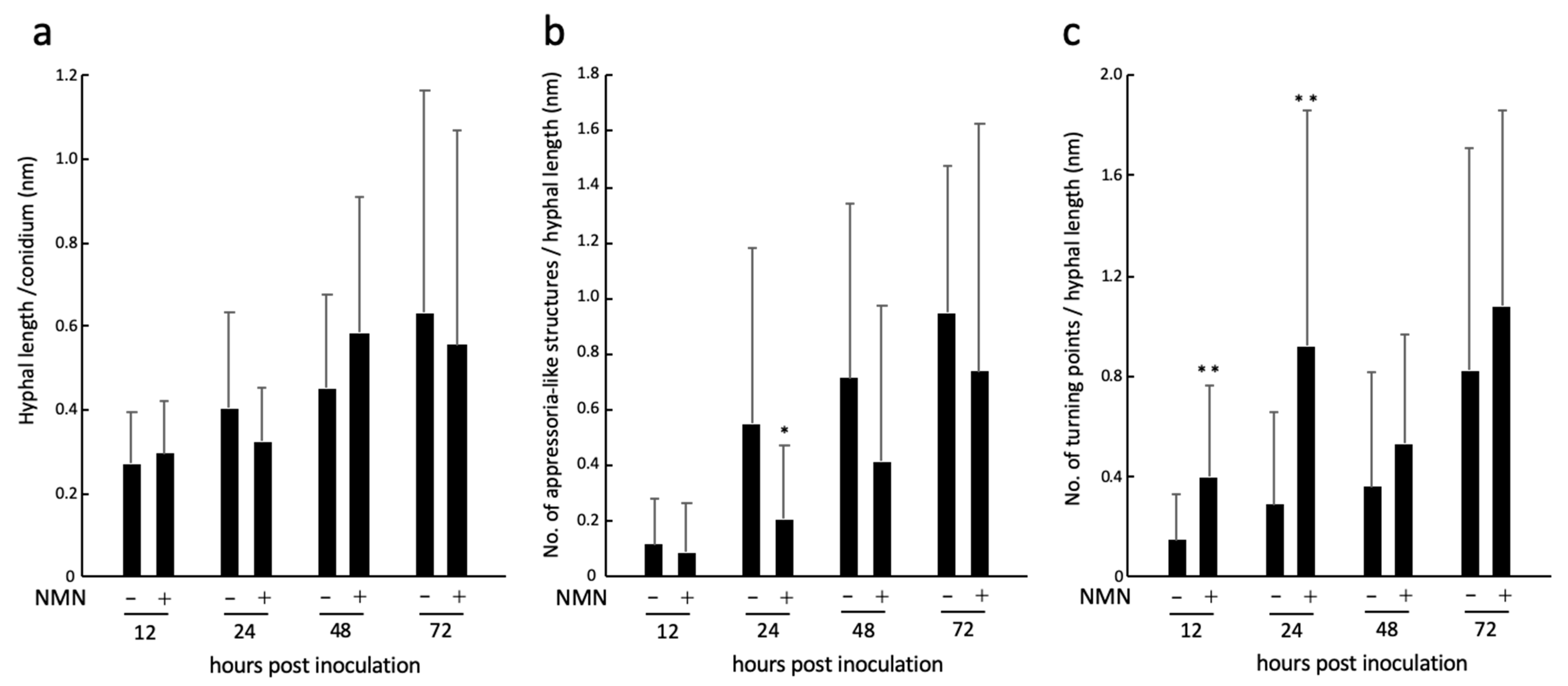
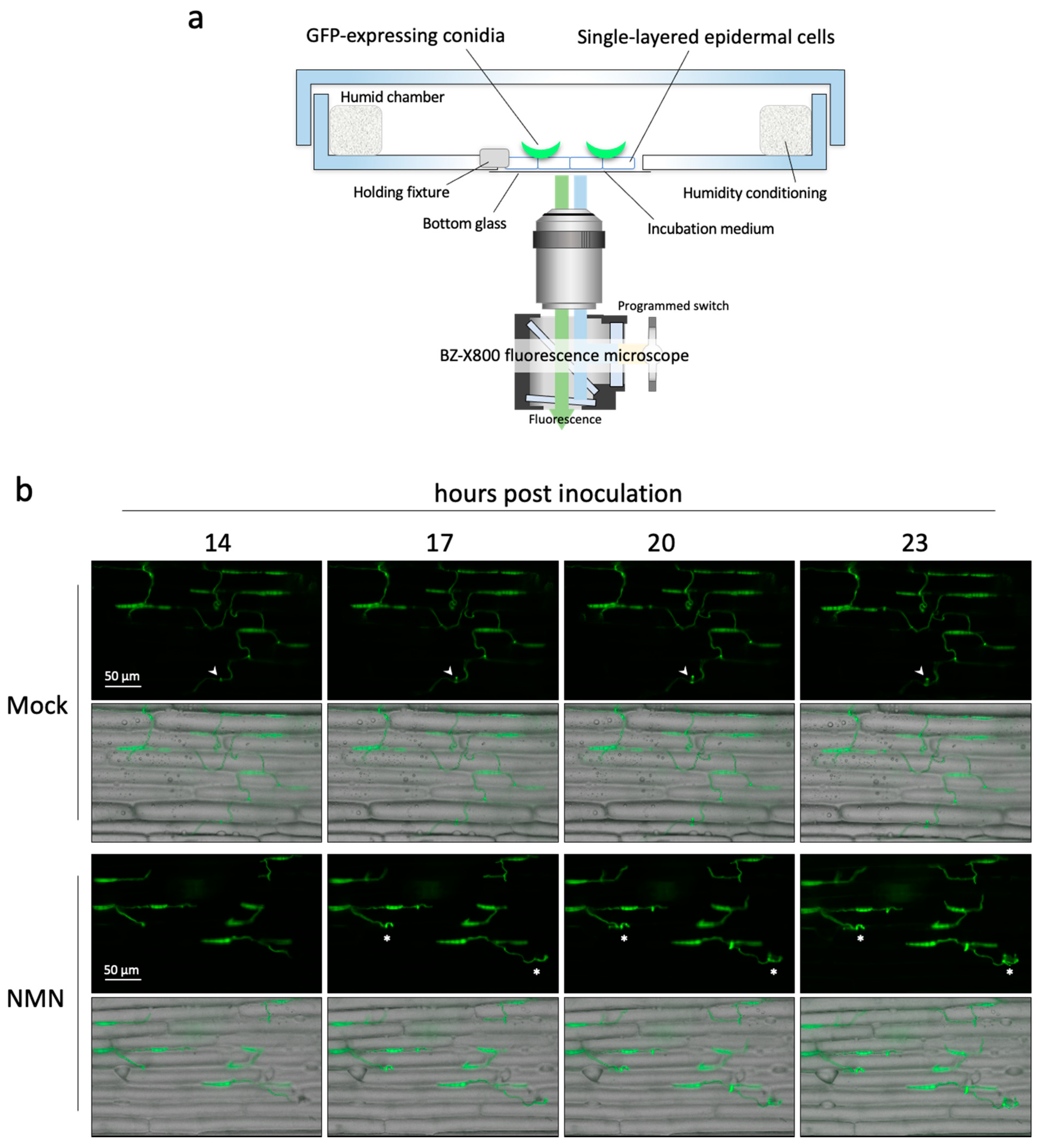
2.3. Real-Time Imaging of Infection Behavior of F. graminearum
3. Materials and Methods
3.1. Plant and Fungal Materials
3.2. Inoculation of Bgh
3.3. Inoculation of F. graminearum
3.4. Real-Time Monitoring of Infection Behavior of F. graminearum
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Desjardins, A.E.; Hohn, T.M. Mycotoxins in plant pathogenesis. Mol. Plant Microbe Interact. 1997, 10, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Liu, X.; Li, B.; Ma, Z. Characterization of Sterol Demethylation Inhibitor-Resistant Isolates of Fusarium asiaticum and F. graminearum Collected from Wheat in China. Phytopathol. 2009, 99, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, L.; Lawton, K.; Ruess, W.; Masner, P.; Specker, N.; Rella, M.G.; Meier, B.; Dincher, S.; Staub, T.; Uknes, S.; et al. A benzothiadiazole derivative induces systemic acquired resistance in tobacco. Plant J. 1996, 10, 61–70. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.; Chaturvedi, R.; Jeannotte, R.; Sparks, A.A.; Shah, J. Involvement of salicylate and jasmonate signaling pathways in Arabidopsis interaction with Fusarium graminearum. Mol. Plant Microbe Interact. 2010, 7, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Makandar, R.; Nalam, V.J.; Lee, H.; Trick, H.N.; Dong, Y.; Shah, J. Salicylic acid regulates basal resistance to Fusarium Head Blight in wheat. Mol. Plant Microbe Interact. 2012, 25, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Mou, Z. Extracellular pyridine nucleotides induce PR gene expression and disease resistance in Arabidopsis. Plant J. 2009, 57, 302–312. [Google Scholar] [CrossRef]
- Wang, C.; Huang, X.; Li, Q.; Zhang, Y.; Li, J.-L.; Mou, Z. Extracellular pyridine nucleotides trigger plant systemic immunity through a lectin receptor kinase/BAK1 complex. Nat. Commun. 2019, 10, 4810. [Google Scholar] [CrossRef] [Green Version]
- Ryrie, I.J.; Scott, K.J. Metabolic regulation in diseased leaves II. Changes in nicotinamide nucleotide coenzymes in barley leaves infected with powdery mildew. Plant Physiol. 1968, 43, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Miwa, A.; Sawada, Y.; Tamaoki, D.; Hirai, M.Y.; Kimura, M.; Sato, K.; Nishiuchi, T. Nicotinamide mononucleotide and related metabolites induce disease resistance against fungal phytopathogens in Arabidopsis and barley. Sci. Rep. 2017, 7, 6389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gakière, B.; Hao, J.; de Bont, L.; Pétriacq, P.; Nunes-Nesi, A.; Fernie, A.R. NAD+ biosynthesis and signaling in plants. Crit. Rev. Plant Sci. 2018, 37, 259–307. [Google Scholar] [CrossRef]
- Pritsch, C.; Muehlbauer, G.J.; Bushnell, W.R.; Somers, D.A.; Vance, C.P. Fungal development and induction of defense response genes during early infection of wheat spikes by Fusarium graminearum. Mol. Plant Microbe Interact. 2000, 13, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, S.M.; Bushnell, W.R.; Evans, C.K. Distribution of mycelial colonies and lesions in field-grown barley inoculated with Fusarium graminearum. Phytopathol. 2006, 96, 567–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanjiru, W.M.; Zhensheng, K.; Buchenauer, H. Importance of cell wall degrading enzymes produced by Fusarium graminearum during infection of wheat heads. Eur. J. Plant Pathol. 2002, 108, 803–810. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Dehne, H.-W.; Steiner, U. Maize leaf trichomes represent an entry point of infection for Fusarium species. Fungal Biol. 2016, 120, 895–903. [Google Scholar] [CrossRef]
- Imboden, L.; Afton, D.; Trail, F. Surface interactions of Fusarium graminearum on barley. Mol. Plant Pathol. 2018, 19, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Zhao, X.; Fang, W.; Wu, H.; Abubakar, Y.S.; Lu, G.-D.; Wang, Z.; Zheng, W. Spatiotemporal nature of Fusarium graminearum-wheat coleoptile interactions. Phytopathol. Res. 2019, 1, 26. [Google Scholar] [CrossRef] [Green Version]
- Sugai, K.; Inoue, H.; Inoue, C.; Sato, M.; Wakazaki, M.; Kobayashi, K.; Nishiguchi, M.; Toyooka, K.; Yamaoka, N.; Yaeno, T. High humidity causes abnormalities in the process of appressorial formation of Blumeria graminis f. sp. hordei. Pathogens 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Jansen, C.; von Wettstein, D.; Schafer, W.; Kogel, K.H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Proc. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [Green Version]
- Talbot, N.J. Appressoria. Curr. Biol. 2019, 29, R144–R146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryce-Jones, E.; Carver, T.; Gurr, S.J. The role of cellulase enzymes and mechanical force in host penetration by Erysiphe graminis f. sp. hordei. Physiol. Mol. Plant Pathol. 1999, 55, 175–182. [Google Scholar] [CrossRef]
- Hao, Q.; Wang, W.; Han, X.; Wu, J.; Lyu, B.; Chen, F.; Caplan, A.; Li, C.; Wu, J.; Wang, W.; et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol. Plant Pathol. 2018, 19, 1995–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamoto, Y.; Toda, H.; Inoue, H.; Kobayashi, K.; Yamaoka, N.; Araki, T.; Yaeno, T. Fast and inexpensive phenotyping and genotyping methods for evaluation of barley mutant population. Plants 2020, 9, 1153. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, N.; Yoshida, S.; Motoyama, E.; Takeuchi, Y.; Takada, Y.; Fukunaga, N. Resistance induction in barley coleoptile cells by intracellular pH decline. Plant Cell Physiol. 2000, 41, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Wahara, M.; Inoue, C.; Kohguchi, T.; Sugai, K.; Kobayashi, K.; Nishiguchi, M.; Yamaoka, N.; Yaeno, T. Improved method for in situ biolistic transformation to analyze barley-powdery mildew interactions. J. Gen. Plant Pathol. 2017, 83, 140–146. [Google Scholar] [CrossRef]
- Nakajima, Y.; Maeda, K.; Ohsato, S.; Kanamaru, K.; Kobayashi, T.; Kimura, M. Nuclear localization and relative stability of the zinc finger domain of TRI6 trichothecene regulator. JSM Mycotoxins 2016, 66, 13–15. [Google Scholar] [CrossRef]
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Nakajima, Y.; Tanahashi, Y.; Kitou, Y.; Miwa, A.; Kanamaru, K.; Kobayashi, T.; Nishiuchi, T.; Kimura, M. L-Threonine and its analogue added to autoclaved solid medium suppress trichothecene production by Fusarium graminearum. Arch. Microbiol. 2017, 199, 945–952. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, K.; Nakajima, Y.; Inoue, H.; Kobayashi, K.; Nishiuchi, T.; Kimura, M.; Yaeno, T. Nicotinamide Mononucleotide Potentiates Resistance to Biotrophic Invasion of Fungal Pathogens in Barley. Int. J. Mol. Sci. 2021, 22, 2696. https://doi.org/10.3390/ijms22052696
Ueda K, Nakajima Y, Inoue H, Kobayashi K, Nishiuchi T, Kimura M, Yaeno T. Nicotinamide Mononucleotide Potentiates Resistance to Biotrophic Invasion of Fungal Pathogens in Barley. International Journal of Molecular Sciences. 2021; 22(5):2696. https://doi.org/10.3390/ijms22052696
Chicago/Turabian StyleUeda, Kana, Yuichi Nakajima, Hiroshi Inoue, Kappei Kobayashi, Takumi Nishiuchi, Makoto Kimura, and Takashi Yaeno. 2021. "Nicotinamide Mononucleotide Potentiates Resistance to Biotrophic Invasion of Fungal Pathogens in Barley" International Journal of Molecular Sciences 22, no. 5: 2696. https://doi.org/10.3390/ijms22052696
APA StyleUeda, K., Nakajima, Y., Inoue, H., Kobayashi, K., Nishiuchi, T., Kimura, M., & Yaeno, T. (2021). Nicotinamide Mononucleotide Potentiates Resistance to Biotrophic Invasion of Fungal Pathogens in Barley. International Journal of Molecular Sciences, 22(5), 2696. https://doi.org/10.3390/ijms22052696