
TRP Channels as Cellular Targets of Particulate Matter


Abstract
:1. Introduction
2. Effects of PM on TRP Channels
2.1. Industrial PM
2.1.1. Titanium Nanoparticles
2.1.2. Silica Nanoparticles
2.1.3. Zinc Nanoparticles
2.2. PM Used in Medicine
2.2.1. Bacterial Particles
2.2.2. Carbon Particles
2.2.3. Gold Particles
2.2.4. Lanthanide Particles
2.2.5. Lipid Particles
2.2.6. Iron Particles
2.2.7. Poly(amidoamine) Particles
2.2.8. Poly(lactic-co-glycolic acid) Particles
2.2.9. Semiconductor Particles
2.2.10. Silica Particles
2.3. PM Resulting from Pollution
2.3.1. Cigarette Smoke
2.3.2. Coal Fly Ash
2.3.3. Diesel Exhaust
2.3.4. Ambient Particles
2.3.5. Wood Smoke
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nanoparticle Technology Handbook. Nano Today 2007, 2, 45. [CrossRef]
- Qhobosheane, M.; Santra, S.; Zhang, P.; Tan, W. Biochemically functionalized silica nanoparticles. Analyst 2001, 126, 1274–1278. [Google Scholar] [CrossRef]
- Wang, J.; Asbach, C.; Fissan, H.; Hülser, T.; Kuhlbusch, T.A.J.; Thompson, D.; Pui, D.Y.H. How can nanobiotechnology oversight advance science and industry: Examples from environmental, health, and safety studies of nanoparticles (nano-EHS). J. Nanopart. Res. 2011, 13, 1373–1387. [Google Scholar] [CrossRef]
- Xia, T.; Kovochich, M.; Brant, J.; Hotze, M.; Sempf, J.; Oberley, T.; Sioutas, C.; Yeh, J.I.; Wiesner, M.R.; Nel, A.E. Comparison of the Abilities of Ambient and Manufactured Nanoparticles To Induce Cellular Toxicity According to an Oxidative Stress Paradigm. Nano Lett. 2006, 6, 1794–1807. [Google Scholar] [CrossRef]
- Hamra, G.B.; Guha, N.; Cohen, A.; Laden, F.; Raaschou-Nielsen, O.; Samet, J.M.; Vineis, P.; Forastiere, F.; Saldiva, P.; Yorifuji, T.; et al. Outdoor Particulate Matter Exposure and Lung Cancer: A Systematic Review and Meta-Analysis. Environ. Health Perspect. 2014, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugadoss, S.; Lison, D.; Godderis, L.; Brule, S.V.D.; Mast, J.; Brassinne, F.; Sebaihi, N.; Hoet, P.H. Toxicology of silica nanoparticles: An update. Arch. Toxicol. 2017, 91, 2967–3010. [Google Scholar] [CrossRef] [PubMed]
- Geiser, M.; Rothen-Rutishauser, B.; Kapp, N.; Schürch, S.; Kreyling, W.; Schulz, H.; Semmler, M.; Hof, V.I.; Heyder, J.; Gehr, P. Ultrafine Particles Cross Cellular Membranes by Nonphagocytic Mechanisms in Lungs and in Cultured Cells. Environ. Health Perspect. 2005, 113, 1555–1560. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, V.; Samal, S.S.; Manoharan, N. Safety and Risk Associated with Nanoparticles—A Review. J. Miner. Mater. Charact. Eng. 2010, 9, 455–459. [Google Scholar] [CrossRef]
- Nilius, B.; Flockerzi, V. What Do We Really Know and What Do We Need to Know: Some Controversies, Perspectives, and Surprises. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Nilius, B., Flockerzi, V., Eds.; Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2014; Volume 223, pp. 1239–1280. [Google Scholar] [CrossRef]
- Flockerzi, V.; Nilius, B. TRPs: Truly Remarkable Proteins. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Nilius, B., Flockerzi, V., Eds.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 1–12. [Google Scholar] [CrossRef]
- Owsianik, G.; Talavera, K.; Voets, T.; Nilius, B. Permeation and Selectivity of TRP Channels. Annu. Rev. Physiol. 2006, 68, 685–717. [Google Scholar] [CrossRef]
- Hung, C.-Y.; Tan, C.-H. TRP Channels in Nociception and Pathological Pain. In Advances in Pain Research: Mechanisms and Modulation of Chronic Pain; Springer: Singapore, 2018; Volume 1099, pp. 13–27. [Google Scholar] [CrossRef]
- González-Ramírez, R.; Chen, Y.; Liedtke, W.B.; Morales-Lázaro, S.L. TRP Channels and Pain. In Neurobiology of TRP Channels; CRC Press: Boca Raton, FL, USA, 2017; pp. 125–148. [Google Scholar] [CrossRef]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals 2016, 9, 72. [Google Scholar] [CrossRef]
- Mori, Y.; Voets, T. Sensors and regulatory mechanisms of thermal physiology. Eur. J. Physiol. 2018, 470, 703–704. [Google Scholar] [CrossRef]
- Vriens, J.; Nilius, B.; Voets, T. Peripheral thermosensation in mammals. Nat. Rev. Neurosci. 2014, 15, 573–589. [Google Scholar] [CrossRef]
- Roper, S.D. TRPs in Taste and Chemesthesis. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2014; Volume 223, pp. 827–871. [Google Scholar] [CrossRef] [Green Version]
- Earley, S.; Brayden, J.E. Transient Receptor Potential Channels in the Vasculature. Physiol. Rev. 2015, 95, 645–690. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Carbajo, L.; Kecskes, M.; Jacobs, G.; Pironet, A.; Syam, N.; Talavera, K.; Vennekens, R. Muscling in on TRP channels in vascular smooth muscle cells and cardiomyocytes. Cell Calcium 2017, 66, 48–61. [Google Scholar] [CrossRef]
- Genova, T.; Gaglioti, D.; Munaron, L. Regulation of Vessel Permeability by TRP Channels. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef]
- Van Gerven, L.; Steelant, B.; Hellings, P.W. Nasal hyperreactivity in rhinitis: A diagnostic and therapeutic challenge. Allergy 2018, 73, 1784–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vennekens, R.; Olausson, J.; Meissner, M.; Bloch, W.; Mathar, I.; Philipp, S.E.; Schmitz, F.; Weissgerber, P.; Nilius, B.; Flockerzi, V.; et al. Increased IgE-dependent mast cell activation and anaphylactic responses in mice lacking the calcium-activated nonselective cation channel TRPM4. Nat. Immunol. 2007, 8, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Alpizar, Y.A.; Boonen, B.; Sanchez, A.; Jung, C.; López-Requena, A.; Naert, R.; Steelant, B.; Luyts, K.; Plata, C.; De Vooght, V.; et al. TRPV4 activation triggers protective responses to bacterial lipopolysaccharides in airway epithelial cells. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Sanchez, A.; Alvarez, J.L.; Demydenko, K.; Jung, C.; Alpizar, Y.A.; Alvarez-Collazo, J.; Cokic, S.M.; Valverde, M.A.; Hoet, P.H.; Talavera, K. Silica nanoparticles inhibit the cation channel TRPV4 in airway epithelial cells. Part. Fibre Toxicol. 2017, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, I.M.; Liedtke, W.; Sanderson, M.J.; Valverde, M.A. TRPV4 channel participates in receptor-operated calcium entry and ciliary beat frequency regulation in mouse airway epithelial cells. Proc. Natl. Acad. Sci. USA 2008, 105, 12611–12616. [Google Scholar] [CrossRef] [Green Version]
- Cantero-Recasens, G.; Butnaru, C.M.; Brouwers, N.; Mitrovic, S.; Valverde, M.A.; Malhotra, V. Sodium channel TRPM4 and sodium/calcium exchangers (NCX) cooperate in the control of Ca2+-induced mucin secretion from goblet cells. J. Biol. Chem. 2019, 294, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zholos, A.V. TRP Channels in Respiratory Pathophysiology: The Role of Oxidative, Chemical Irritant and Temperature Stimuli. Curr. Neuropharmacol. 2015, 13, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Mukaiyama, M.; Usui, T.; Nagumo, Y. Non-electrophilic TRPA1 agonists, menthol, carvacrol and clotrimazole, open epithelial tight junctions via TRPA1 activation. J. Biochem. 2020, 168, 407–415. [Google Scholar] [CrossRef]
- Sokabe, T.; Fukumi-Tominaga, T.; Yonemura, S.; Mizuno, A.; Tominaga, M. The TRPV4 Channel Contributes to Intercellular Junction Formation in Keratinocytes*. J. Biol. Chem. 2010, 285, 18749–18758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokabe, T.; Tominaga, M. The TRPV4 cation channel: A molecule linking skin temperature and barrier function. Commun. Integr. Biol. 2010, 3, 619–621. [Google Scholar] [CrossRef]
- Kida, N.; Sokabe, T.; Kashio, M.; Haruna, K.; Mizuno, Y.; Suga, Y.; Nishikawa, K.; Kanamaru, A.; Hongo, M.; Oba, A.; et al. Importance of transient receptor potential vanilloid 4 (TRPV4) in epidermal barrier function in human skin keratinocytes. Pflugers Arch. Eur. J. Physiol. 2012, 463, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Huang, J.; Yang, Q.; Li, T.; Liu, J.; Qian, Z. Gold nanorods-based thermosensitive hydrogel produces selective long-lasting regional anesthesia triggered by photothermal activation of Transient Receptor Potential Vanilloid Type-1 channels. Colloids Surf. B Biointerfaces 2018, 171, 17–23. [Google Scholar] [CrossRef]
- Akopian, A.N.; Fanick, E.R.; Brooks, E.G. TRP channels and traffic-related environmental pollution-induced pulmonary disease. Semin. Immunopathol. 2016, 38, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Dubes, V.; Parpaite, T.; Ducret, T.; Quignard, J.-F.; Mornet, S.; Reinhardt, N.; Baudrimont, I.; Dubois, M.; Freund-Michel, V.; Marthan, R.; et al. Calcium signalling induced by in vitro exposure to silicium dioxide nanoparticles in rat pulmonary artery smooth muscle cells. Toxicology 2017, 375, 37–47. [Google Scholar] [CrossRef]
- Mohammadpour, R.; Yazdimamaghani, M.; Reilly, C.A.; Ghandehari, H.; Ghandehari, H. Transient Receptor Potential Ion Channel–Dependent Toxicity of Silica Nanoparticles and Poly(amido amine) Dendrimers. J. Pharmacol. Exp. Ther. 2018, 370, 751–760. [Google Scholar] [CrossRef]
- Gregurec, D.; Senko, A.W.; Chuvilin, A.; Reddy, P.D.; Sankararaman, A.; Rosenfeld, D.; Chiang, P.-H.; Garcia, F.; Tafel, I.; Varnavides, G.; et al. Magnetic Vortex Nanodiscs Enable Remote Magnetomechanical Neural Stimulation. ACS Nano 2020, 14, 8036–8045. [Google Scholar] [CrossRef]
- Jurek, S.C.; Hirano-Kobayashi, M.; Chiang, H.; Kohane, D.S.; Matthews, B.D. Prevention of Ventilator-Induced Lung Edema by Inhalation of Nanoparticles Releasing Ruthenium Red. Am. J. Respir. Cell Mol. Biol. 2014, 50, 1107–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevan, S.; Quallo, T.; Andersson, D.A. TRPV1. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 207–245. [Google Scholar]
- Garcia-Elias, A.; Mrkonjic, S.; Jung, C. The TRPV4 Channel. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 293–319. [Google Scholar]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Zygmunt, P.M.; Hogestatt, E.D. TRPA1. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 583–630. [Google Scholar]
- Viana, F. TRPA1 channels: Molecular sentinels of cellular stress and tissue damage. J. Physiol. 2016, 594, 4151–4169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Li, J.; Jiang, J.; Zhao, Z.; Hui, Z.; Zhang, J.; Zheng, Y.; Ling, D.; Wang, L.; Jiang, L.-H.; et al. A dual role of transient receptor potential melastatin 2 channel in cytotoxicity induced by silica nanoparticles. Sci. Rep. 2015, 5, 18171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Z.; Wang, Z.; Chen, Z.; Jin, H.; Chen, C.; Chai, S.; Lv, H.; Yang, L.; Hu, Y.; Dong, R.; et al. Melatonin Attenuates Chronic Cough Mediated by Oxidative Stress via Transient Receptor Potential Melastatin-2 in Guinea Pigs Exposed to Particulate Matter 2.5. Physiol. Res. 2018, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Gao, Y.; Wang, C.; Tao, R.; Wu, Y.; Zhan, K.; Liao, M.; Lu, N.; Lu, Y.; Wilcox, C.S.; et al. Nitration of TRPM2 as a Molecular Switch Induces Autophagy During Brain Pericyte Injury. Antioxid. Redox Signal. 2017, 27, 1297–1316. [Google Scholar] [CrossRef]
- Liu, Y.; Mikrani, R.; He, Y.; Baig, M.M.F.A.; Abbas, M.; Naveed, M.; Tang, M.; Zhang, Q.; Li, C.; Zhou, X. TRPM8 channels: A review of distribution and clinical role. Eur. J. Pharmacol. 2020, 882, 173312. [Google Scholar] [CrossRef]
- Almaraz, L.; Manenschijn, J.-A.; de la Pena, E.; Viana, F. TRPM8. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 547–579. [Google Scholar]
- Rosenbaum, T.; Benítez-Angeles, M.; Sánchez-Hernández, R.; Morales-Lázaro, S.L.; Hiriart, M.; Morales-Buenrostro, L.E.; Torres-Quiroz, F. TRPV4: A Physio and Pathophysiologically Significant Ion Channel. Int. J. Mol. Sci. 2020, 21, 3837. [Google Scholar] [CrossRef]
- Bonvini, S.J.; Belvisi, M.G. Cough and airway disease: The role of ion channels. Pulm. Pharmacol. Ther. 2017, 47, 21–28. [Google Scholar] [CrossRef]
- Benemei, S.; Patacchini, R.; Trevisani, M.; Geppetti, P. TRP Channels. Curr. Opin. Pharmacol. 2015, 22, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Fariss, M.W.; Gilmour, M.I.; Reilly, C.A.; Liedtke, W.; Ghio, A.J. Emerging Mechanistic Targets in Lung Injury Induced by Combustion-Generated Particles. Toxicol. Sci. 2013, 132, 253–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.P.M.; Cibelli, M.; Urban, L.; Nilius, B.; McGeown, J.G.; Nagy, I. TRPV4: Molecular Conductor of a Diverse Orchestra. Physiol. Rev. 2016, 96, 911–973. [Google Scholar] [CrossRef] [Green Version]
- Gilardino, A.; Catalano, F.; Ruffinatti, F.A.; Alberto, G.; Nilius, B.; Antoniotti, S.; Martra, G.; Lovisolo, D. Interaction of SiO2 nanoparticles with neuronal cells: Ionic mechanisms involved in the perturbation of calcium homeostasis. Int. J. Biochem. Cell Biol. 2015, 66, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Kan, H.; Wu, Z.; Lin, Y.-C.; Chen, T.-H.; Cumpston, J.L.; Kashon, M.L.; Leonard, S.; E Munson, A.; Castranova, V. The role of nodose ganglia in the regulation of cardiovascular function following pulmonary exposure to ultrafine titanium dioxide. Nanotoxicology 2013, 8, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-G.; Park, M.-K.; Lee, P.-H.; Lee, S.-H.; Hong, J.; Aung, M.M.M.; Moe, K.T.; Han, N.Y.; Jang, A.-S. Effects of nanoparticles on neuroinflammation in a mouse model of asthma. Respir. Physiol. Neurobiol. 2020, 271, 103292. [Google Scholar] [CrossRef]
- Wu, W.; Bromberg, P.A.; Samet, J.M. Zinc ions as effectors of environmental oxidative lung injury. Free Radic. Biol. Med. 2013, 65, 57–69. [Google Scholar] [CrossRef]
- Scheraga, R.G.; Abraham, S.; Niese, K.A.; Southern, B.D.; Grove, L.M.; Hite, R.D.; McDonald, C.; Hamilton, T.A.; Olman, M.A. TRPV4 Mechanosensitive Ion Channel Regulates Lipopolysaccharide-Stimulated Macrophage Phagocytosis. J. Immunol. 2015, 196, 428–436. [Google Scholar] [CrossRef]
- Hajna, Z.; Csekő, K.; Kemény, A.; Kereskai, L.; Kiss, T.; Perkecz, A.; Szitter, I.; Kocsis, B.; Pintér, E.; Helyes, Z. Complex Regulatory Role of the TRPA1 Receptor in Acute and Chronic Airway Inflammation Mouse Models. Int. J. Mol. Sci. 2020, 21, 4109. [Google Scholar] [CrossRef]
- Diogenes, A.; Ferraz, C.; Akopian, A.; Henry, M.; Hargreaves, K. LPS Sensitizes TRPV1 via Activation of TLR4 in Trigeminal Sensory Neurons. J. Dent. Res. 2011, 90, 759–764. [Google Scholar] [CrossRef]
- Boonen, B.; Alpizar, Y.A.; Sanchez, A.; López-Requena, A.; Voets, T.; Talavera, K. Differential effects of lipopolysaccharide on mouse sensory TRP channels. Cell Calcium 2018, 73, 72–81. [Google Scholar] [CrossRef]
- Alpizar, Y.A.; Uvin, P.; Naert, R.; Franken, J.; Pinto, S.; Sanchez, A.; Gevaert, T.; Everaerts, W.; Voets, T.; De Ridder, D.; et al. TRPV4 Mediates Acute Bladder Responses to Bacterial Lipopolysaccharides. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Meseguer, V.M.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.L.; Fajardo, O.; Manenschijn, J.-A.; Fernández-Peña, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5, 3125. [Google Scholar] [CrossRef] [Green Version]
- Soldano, A.; Alpizar, Y.A.; Boonen, B.; Franco, L.; López-Requena, A.; Liu, G.; Mora, N.; Yaksi, E.; Voets, T.; Vennekens, R.; et al. Gustatory-mediated avoidance of bacterial lipopolysaccharides via TRPA1 activation in Drosophila. eLife 2016, 5. [Google Scholar] [CrossRef]
- Startek, J.B.; Talavera, K.; Voets, T.; Alpizar, Y.A. Differential interactions of bacterial lipopolysaccharides with lipid membranes: Implications for TRPA1-mediated chemosensation. Sci. Rep. 2018, 8, 12010. [Google Scholar] [CrossRef] [PubMed]
- Startek, J.B.; Talavera, K. Lipid Raft Destabilization Impairs Mouse TRPA1 Responses to Cold and Bacterial Lipopolysaccharides. Int. J. Mol. Sci. 2020, 21, 3826. [Google Scholar] [CrossRef]
- Boonen, B.; Alpizar, Y.A.; Meseguer, V.M.; Talavera, K. TRP Channels as Sensors of Bacterial Endotoxins. Toxins 2018, 10, 326. [Google Scholar] [CrossRef] [Green Version]
- Startek, J.B.; Boonen, B.; Talavera, K.; Meseguer, V. TRP Channels as Sensors of Chemically-Induced Changes in Cell Membrane Mechanical Properties. Int. J. Mol. Sci. 2019, 20, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryn, D.O.; Melnyk, M.I.; Al Kury, L.T.; Prylutskyy, Y.I.; Ritter, U.; Zholos, A.V. C60 fullerenes disrupt cellular signalling leading to TRPC4 and TRPC6 channels opening by the activation of muscarinic receptors and G-proteins in small intestinal smooth muscles. Cell. Signal. 2018, 43, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, Q.; Kolosov, V.P.; Perelman, J.M.; Zhou, X. Regulation of Particulate Matter-Induced Mucin Secretion by Transient Receptor Potential Vanilloid 1 Receptors. Inflammation 2012, 35, 1851–1859. [Google Scholar] [CrossRef]
- Lee, D.U.; Park, J.-Y.; Kwon, S.; Park, J.Y.; Kim, Y.H.; Khang, D.; Hong, J.H.; Kim, Y.H. Apoptotic lysosomal proton sponge effect in tumor tissue by cationic gold nanorods. Nanoscale 2019, 11, 19980–19993. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, Y.; Liu, L.; Xu, Y.; Liu, X.; Lin, J.; Zhou, W.; Wei, P.; Jin, P.; Wen, L.-P. Inhibition of lanthanide nanocrystal-induced inflammasome activation in macrophages by a surface coating peptide through abrogation of ROS production and TRPM2-mediated Ca2+ influx. Biomaterials 2016, 108, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Puglia, C.; Santonocito, D.; Bonaccorso, A.; Musumeci, T.; Ruozi, B.; Pignatello, R.; Carbone, C.; Parenti, C.; Chiechio, S. Lipid Nanoparticle Inclusion Prevents Capsaicin-Induced TRPV1 Defunctionalization. Pharmaceutics 2020, 12, 339. [Google Scholar] [CrossRef]
- Sharma, G.; Chopra, K.; Puri, S.; Bishnoi, M.; Rishi, P.; Kaur, I.P. Topical delivery of TRPsiRNA-loaded solid lipid nanoparticles confer reduced pain sensation via TRPV1 silencing, in rats. J. Drug Target. 2017, 26, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Grolez, G.P.; Hammadi, M.; Barras, A.; Gordienko, D.; Slomianny, C.; Völkel, P.; Angrand, P.O.; Pinault, M.; Guimaraes, C.; Potier-Cartereau, M.; et al. Encapsulation of a TRPM8 Agonist, WS12, in Lipid Nanocapsules Potentiates PC3 Prostate Cancer Cell Migration Inhibition through Channel Activation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Chechetka, S.A.; Doi, M.; Pichon, B.P.; Bégin-Colin, S.; Miyako, E. Photothermal and mechanical stimulation of cells via dualfunctional nanohybrids. Nanotechnology 2016, 27, 475102. [Google Scholar] [CrossRef]
- Huang, H.; Delikanli, S.; Zeng, H.; Ferkey, D.M.; Pralle, A. Remote control of ion channels and neurons through magnetic-field heating of nanoparticles. Nat. Nanotechnol. 2010, 5, 602–606. [Google Scholar] [CrossRef]
- Rosenfeld, D.; Senko, A.W.; Moon, J.; Yick, I.; Varnavides, G.; Gregureć, D.; Koehler, F.; Chiang, P.-H.; Christiansen, M.G.; Maeng, L.Y.; et al. Health and Medicine Transgene-free remote magnetothermal regulation of adrenal hormones. Sci. Adv. 2020, 6, eaaz3734. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; McBrayer, M.K.; Wolfe, D.M.; Haslett, L.J.; Kumar, A.; Sato, Y.; Lie, P.P.; Mohan, P.; Coffey, E.E.; Kompella, U.; et al. Presenilin 1 Maintains Lysosomal Ca2+ Homeostasis via TRPML1 by Regulating vATPase-Mediated Lysosome Acidification. Cell Rep. 2015, 12, 1430–1444. [Google Scholar] [CrossRef] [Green Version]
- Jian, T.; Chen, J.; Ding, X.; Lv, H.; Li, J.; Wu, Y.; Ren, B.; Tong, B.; Zuo, Y.; Su, K.; et al. Flavonoids isolated from loquat (Eriobotrya japonica) leaves inhibit oxidative stress and inflammation induced by cigarette smoke in COPD mice: The role of TRPV1 signaling pathways. Food Funct. 2020, 11, 3516–3526. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.; Liu, X.; Sun, J. Dual-Functional Dendritic Mesoporous Bioactive Glass Nanospheres for Calcium Influx-Mediated Specific Tumor Suppression and Controlled Drug Delivery in Vivo. ACS Appl. Mater. Interfaces 2018, 10, 23548–23559. [Google Scholar] [CrossRef]
- Nie, Y.; Huang, C.; Zhong, S.; Wortley, M.A.; Luo, Y.; Luo, W.; Xie, Y.; Lai, K.; Zhong, N. Cigarette smoke extract (CSE) induces transient receptor potential ankyrin 1(TRPA1) expression via activation of HIF1αin A549 cells. Free Radic. Biol. Med. 2016, 99, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Andrè, E.; Campi, B.; Materazzi, S.; Trevisani, M.; Amadesi, S.; Massi, D.; Creminon, C.; Vaksman, N.; Nassini, R.; Civelli, M.; et al. Cigarette smoke–induced neurogenic inflammation is mediated by α,β-unsaturated aldehydes and the TRPA1 receptor in rodents. J. Clin. Investig. 2008, 118, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Kichko, T.I.; Kobal, G.; Reeh, P.W. Cigarette smoke has sensory effects through nicotinic and TRPA1 but not TRPV1 receptors on the isolated mouse trachea and larynx. Am. J. Physiol. Cell. Mol. Physiol. 2015, 309, L812–L820. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.-H.; Liu, M.-H.; Ko, H.-K.; Perng, D.-W.; Lee, T.-S.; Kou, Y.R. Lung Epithelial TRPA1 Transduces the Extracellular ROS into Transcriptional Regulation of Lung Inflammation Induced by Cigarette Smoke: The Role of Influxed Ca2+. Mediat. Inflamm. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, Y.; Xu, M.; Zhang, H.; Chen, Y.; Chung, K.F.; Adcock, I.M.; Li, F. Roles of TRPA1 and TRPV1 in cigarette smoke -induced airway epithelial cell injury model. Free Radic. Biol. Med. 2019, 134, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, D.N.; Liu, B.; Ha, M.A.; Jordt, S.; Morris, J.B. Menthol attenuates respiratory irritation responses to multiple cigarette smoke irritants. FASEB J. 2011, 25, 4434–4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, W.-H.; Hsu, C.-C.; Chiang, L.-L.; Lin, Y.-J.P.; Lin, Y.S.; Su, C.-L. Role of TRPV1 and P2X receptors in the activation of lung vagal C-fiber afferents by inhaled cigarette smoke in rats. Mol. Med. Rep. 2013, 7, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Hsu, C.-C.; Bien, M.-Y.; Hsu, H.-C.; Weng, H.-T.; Kou, Y.R. Activations of TRPA1 and P2X receptors are important in ROS-mediated stimulation of capsaicin-sensitive lung vagal afferents by cigarette smoke in rats. J. Appl. Physiol. 2010, 108, 1293–1303. [Google Scholar] [CrossRef]
- Baxter, M.; Eltom, S.; Dekkak, B.; Yew-Booth, L.; Dubuis, E.D.; Maher, S.A.; Belvisi, M.G.; Birrell, M.A. Role of transient receptor potential and pannexin channels in cigarette smoke-triggered ATP release in the lung. Thorax 2014, 69, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Yang, C.; Huang, L.; Chen, M.; Shi, J.; Ouyang, L.; Tang, T.; Zhang, W.; Li, Y.; Liang, R.; et al. Upregulation of TRPM7 augments cell proliferation and interleukin-8 release in airway smooth muscle cells of rats exposed to cigarette smoke. Mol. Med. Rep. 2016, 13, 4995–5004. [Google Scholar] [CrossRef] [Green Version]
- Deering-Rice, C.E.; Stockmann, C.; Romero, E.G.; Lu, Z.; Shapiro, D.; Stone, B.L.; Fassl, B.; Nkoy, F.; Uchida, D.A.; Ward, R.M.; et al. Characterization of Transient Receptor Potential Vanilloid-1 (TRPV1) Variant Activation by Coal Fly Ash Particles and Associations with Altered Transient Receptor Potential Ankyrin-1 (TRPA1) Expression and Asthma. J. Biol. Chem. 2016, 291, 24866–24879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimilli, S.; Makena, P.; Prasad, G. Combustible Cigarette and Smokeless Tobacco Product Preparations Differentially Regulate Intracellular Calcium Mobilization in HL60 Cells. Inflammation 2019, 42, 1641–1651. [Google Scholar] [CrossRef] [Green Version]
- Kiss, F.; Pohóczky, K.; Szállási, A.; Helyes, Z. Transient Receptor Potential (TRP) Channels in Head-and-Neck Squamous Cell Carcinomas: Diagnostic, Prognostic, and Therapeutic Potentials. Int. J. Mol. Sci. 2020, 21, 6374. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.G.; Romero, E.G.; Lu, Z.; Marcus, S.K.; Peterson, H.C.; Veranth, J.M.; Deering-Rice, C.E.; Reilly, C.A. Activation of Human Transient Receptor Potential Melastatin-8 (TRPM8) by Calcium-Rich Particulate Materials and Effects on Human Lung Cells. Mol. Pharmacol. 2017, 92, 653–664. [Google Scholar] [CrossRef]
- Deering-Rice, C.E.; Romero, E.G.; Shapiro, D.; Hughen, R.W.; Light, A.R.; Yost, G.S.; Veranth, J.M.; Reilly, C.A. Electrophilic Components of Diesel Exhaust Particles (DEP) Activate Transient Receptor Potential Ankyrin-1 (TRPA1): A Probable Mechanism of Acute Pulmonary Toxicity for DEP. Chem. Res. Toxicol. 2011, 24, 950–959. [Google Scholar] [CrossRef]
- Deering-Rice, C.E.; Shapiro, D.; Romero, E.G.; Stockmann, C.; Bevans, T.S.; Phan, Q.M.; Stone, B.L.; Fassl, B.; Nkoy, F.; Uchida, D.A.; et al. Activation of Transient Receptor Potential Ankyrin-1 by Insoluble Particulate Material and Association with Asthma. Am. J. Respir. Cell Mol. Biol. 2015, 53, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Deering-Rice, C.E.; Johansen, M.E.; Roberts, J.K.; Thomas, K.C.; Romero, E.G.; Lee, J.; Yost, G.S.; Veranth, J.M.; Reilly, C.A. Transient Receptor Potential Vanilloid-1 (TRPV1) Is a Mediator of Lung Toxicity for Coal Fly Ash Particulate Material. Mol. Pharmacol. 2011, 81, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Deering-Rice, C.E.; Memon, T.; Lu, Z.; Romero, E.G.; Cox, J.; Taylor-Clark, T.; Veranth, J.M.; Reilly, C.A. Differential Activation of TRPA1 by Diesel Exhaust Particles: Relationships between Chemical Composition, Potency, and Lung Toxicity. Chem. Res. Toxicol. 2019, 32, 1040–1050. [Google Scholar] [CrossRef]
- Stevens, J.S.; Padilla, S.; DeMarini, D.M.; Hunter, D.L.; Martin, W.K.; Thompson, L.C.; Gilmour, M.I.; Hazari, M.S.; Farraj, A.K. Zebrafish Locomotor Responses Reveal Irritant Effects of Fine Particulate Matter Extracts and a Role for TRPA1. Toxicol. Sci. 2017, 161, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.K.; Birrell, M.A.; Adcock, J.J.; Wortley, M.A.; Dubuis, E.D.; Chen, S.; McGilvery, C.M.; Hu, S.; Shaffer, M.S.; Bonvini, S.J.; et al. Mechanistic link between diesel exhaust particles and respiratory reflexes. J. Allergy Clin. Immunol. 2018, 141, 1074–1084.e9. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.C.; Walsh, L.; Martin, B.L.; McGee, J.; Wood, C.; Kovalcik, K.; Pancras, J.P.; Haykal-Coates, N.; Ledbetter, A.D.; Davies, D.; et al. Ambient Particulate Matter and Acrolein Co-Exposure Increases Myocardial Dyssynchrony in Mice via TRPA1. Toxicol. Sci. 2018, 167, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kanju, P.; Patterson, M.; Chew, W.-L.; Cho, S.-H.; Gilmour, I.; Oliver, T.; Yasuda, R.; Ghio, A.; Simon, S.A.; et al. TRPV4-Mediated Calcium Influx into Human Bronchial Epithelia upon Exposure to Diesel Exhaust Particles. Environ. Health Perspect. 2011, 119, 784–793. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.U.; Ji, M.J.; Kang, J.Y.; Kyung, S.Y.; Hong, J.H. Dust particles-induced intracellular Ca2+ signaling and reactive oxygen species in lung fibroblast cell line MRC5. Korean J. Physiol. Pharmacol. 2017, 21, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, L.; Moreno-Vinasco, L.; Lang, G.D.; Siegler, J.H.; Mathew, B.; Usatyuk, P.V.; Samet, J.M.; Geyh, A.S.; Breysse, P.N.; et al. Particulate matter air pollution disrupts endothelial cell barrier via calpain-mediated tight junction protein degradation. Part. Fibre Toxicol. 2012, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, K.; Park, S.-H.; Han, B.S.; Oh, S.W.; Lee, S.E.; Yoo, J.A.; Park, S.J.; Kim, J.; Kim, J.W.; Cho, J.Y.; et al. Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1. Int. J. Mol. Sci. 2018, 19, 2660. [Google Scholar] [CrossRef] [Green Version]
- Ghelfi, E.; Rhoden, C.R.; Wellenius, G.A.; Lawrence, J.; Gonzalez-Flecha, B. Cardiac Oxidative Stress and Electrophysiological Changes in Rats Exposed to Concentrated Ambient Particles are Mediated by TRP-Dependent Pulmonary Reflexes. Toxicol. Sci. 2008, 102, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Yue, J.; Chen, Z.; Chai, S.; Cao, X.; Zhan, J.; Ji, Z.; Zhang, H.; Dong, R.; Lai, K. Effect of transient receptor potential vanilloid-1 on cough hypersensitivity induced by particulate matter 2.5. Life Sci. 2016, 151, 157–166. [Google Scholar] [CrossRef]
- Liu, H.; Fan, X.; Wang, N.; Zhang, Y.; Yu, J. Exacerbating effects of PM2.5 in OVA-sensitized and challenged mice and the expression of TRPA1 and TRPV1 proteins in lungs. J. Asthma 2017, 54, 807–817. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, Y.; Wang, M.; Zhang, H.; Chen, Y.; Adcock, I.M.; Chung, K.F.; Mo, J.; Zhang, Y.; Li, F. TRPV1 and TRPA1 in Lung Inflammation and Airway Hyperresponsiveness Induced by Fine Particulate Matter (PM2.5). Oxid. Med. Cell. Longev. 2019, 2019, 7450151. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, D.; Deering-Rice, C.E.; Romero, E.G.; Hughen, R.W.; Light, A.R.; Veranth, J.M.; Reilly, C.A. Activation of Transient Receptor Potential Ankyrin-1 (TRPA1) in Lung Cells by Wood Smoke Particulate Material. Chem. Res. Toxicol. 2013, 26, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Deering-Rice, C.E.; Nguyen, N.; Lu, Z.; Cox, J.E.; Shapiro, D.; Romero, E.G.; Mitchell, V.K.; Burrell, K.L.; Veranth, J.M.; Reilly, C.A. Activation of TRPV3 by Wood Smoke Particles and Roles in Pneumotoxicity. Chem. Res. Toxicol. 2018, 31, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.D.; Memon, T.A.; Burrell, K.L.; Almestica-Roberts, M.; Rapp, E.; Sun, L.; Scott, A.F.; Deering-Rice, C.; Rower, J.E.; Reilly, C.A. Transient Receptor Potential Ankyrin-1 (TRPA1) and Vanilloid-3 (TRPV3) Differentially Regulate Endoplasmic Reticulum Stress and Cytotoxicity in Human Lung Epithelial Cells Following Pneumotoxic Wood Smoke Particle Exposure. Mol. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Memon, T.A.; Nguyen, N.D.; Burrell, K.L.; Scott, A.F.; Almestica-Roberts, M.; Rapp, E.; Deering-Rice, C.E.; Reilly, C.A. Wood Smoke Particles Stimulate MUC5AC Overproduction by Human Bronchial Epithelial Cells Through TRPA1 and EGFR Signaling. Toxicol. Sci. 2020, 174, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Amarouch, M.-Y.; El Hilaly, J. Inherited Cardiac Arrhythmia Syndromes: Focus on Molecular Mechanisms Underlying TRPM4 Channelopathies. Cardiovasc. Ther. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Naert, R.; Talavera, A.; Startek, J.B. TRPA1 gene variants hurting our feelings. Pflugers Arch. Eur. J. Physiol. 2020, 472, 953–960. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. The puzzle of TRPV4 channelopathies. EMBO Rep. 2013, 14, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. Transient receptor potential channelopathies. Pflugers Arch. Eur. J. Physiol. 2010, 460, 437–450. [Google Scholar] [CrossRef]






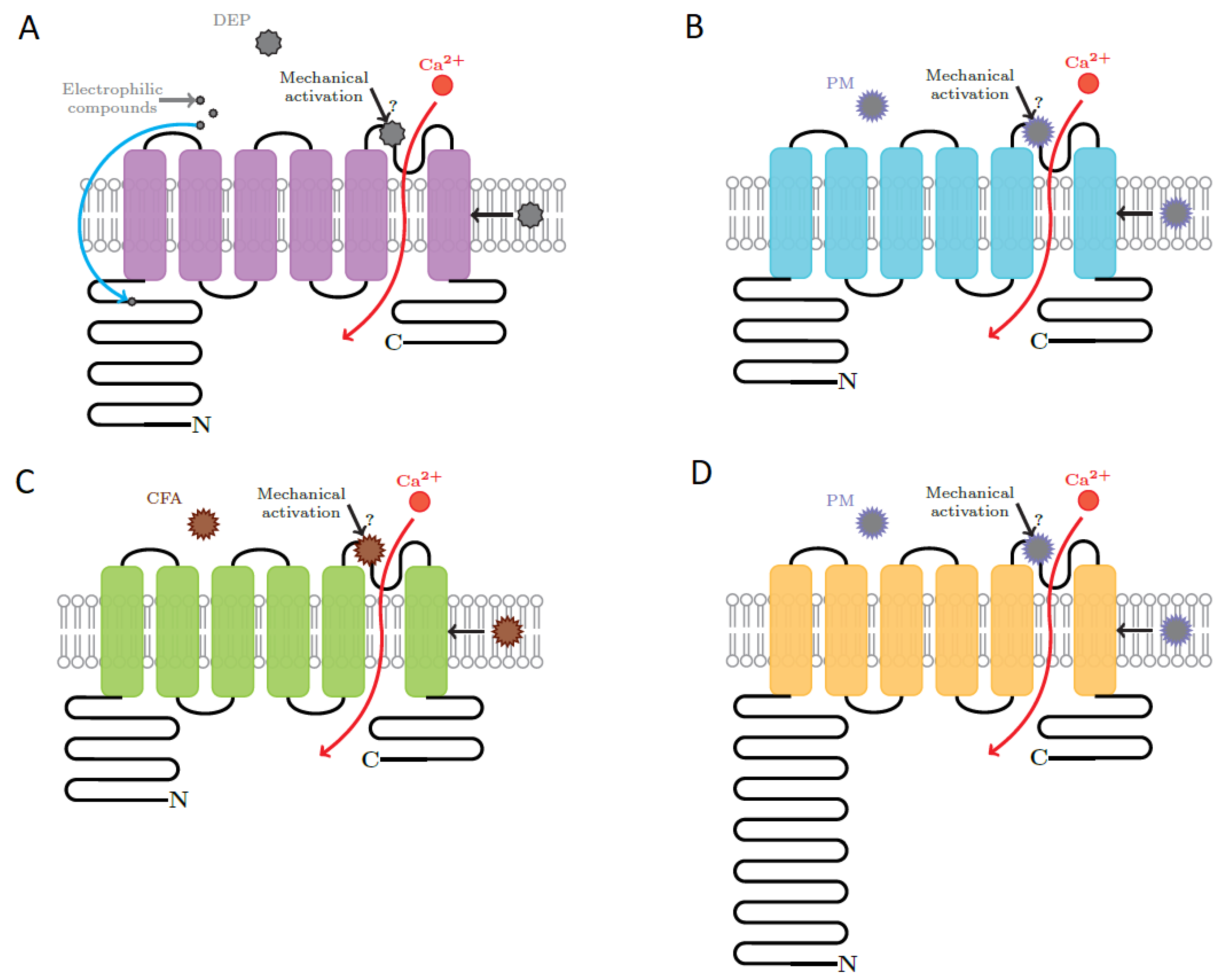{kind=link}
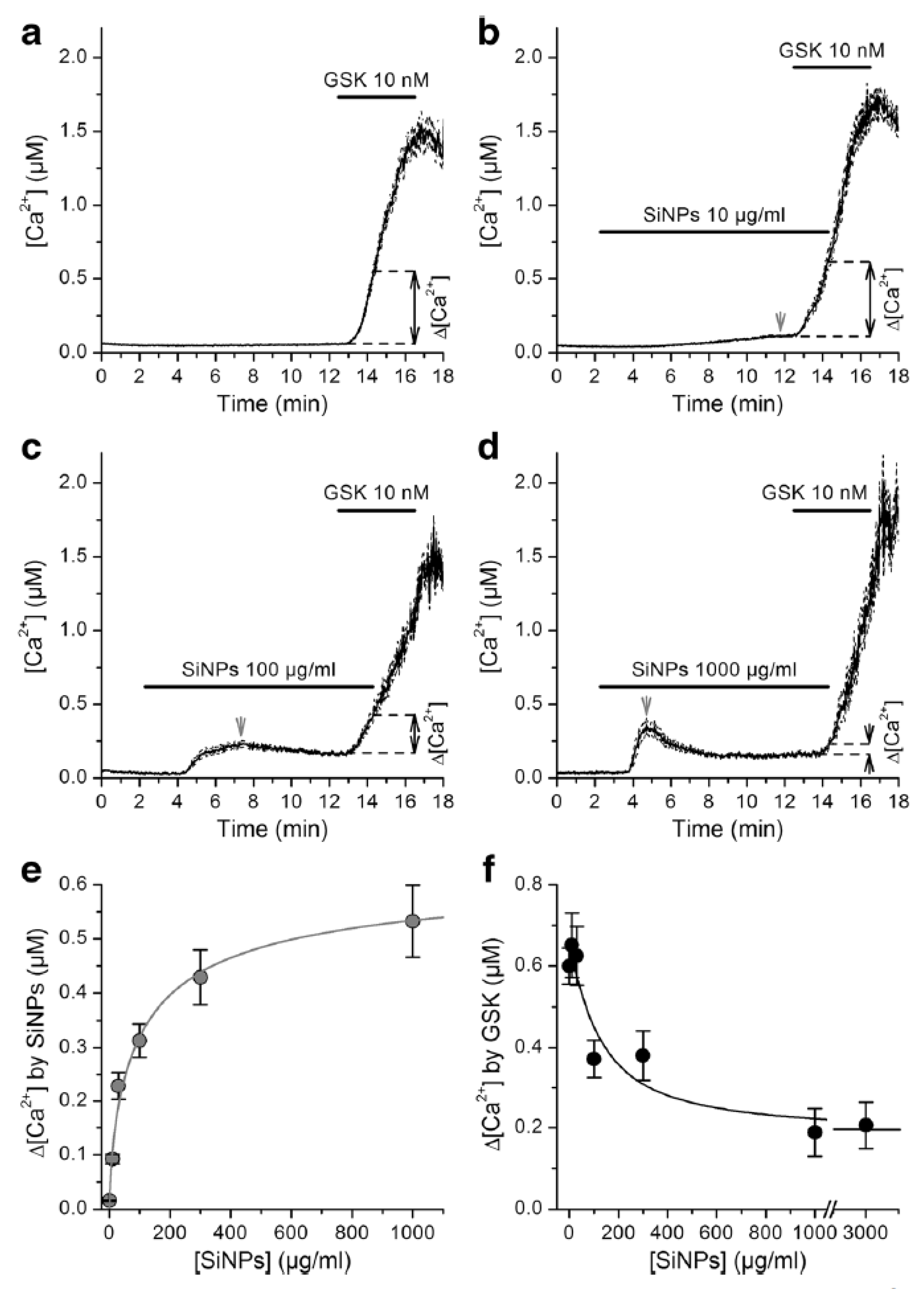{kind=link}
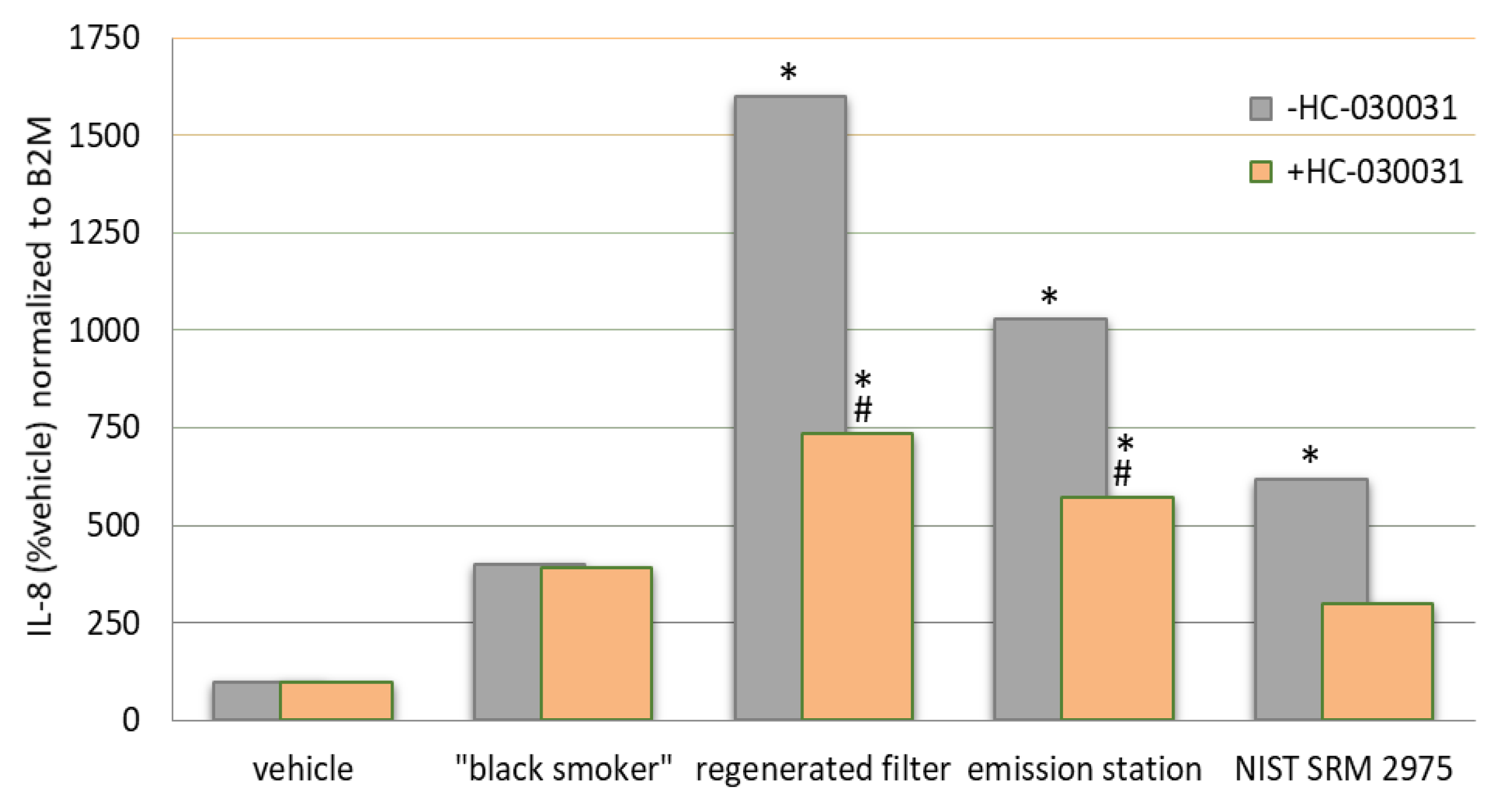{kind=link}
{kind=link}
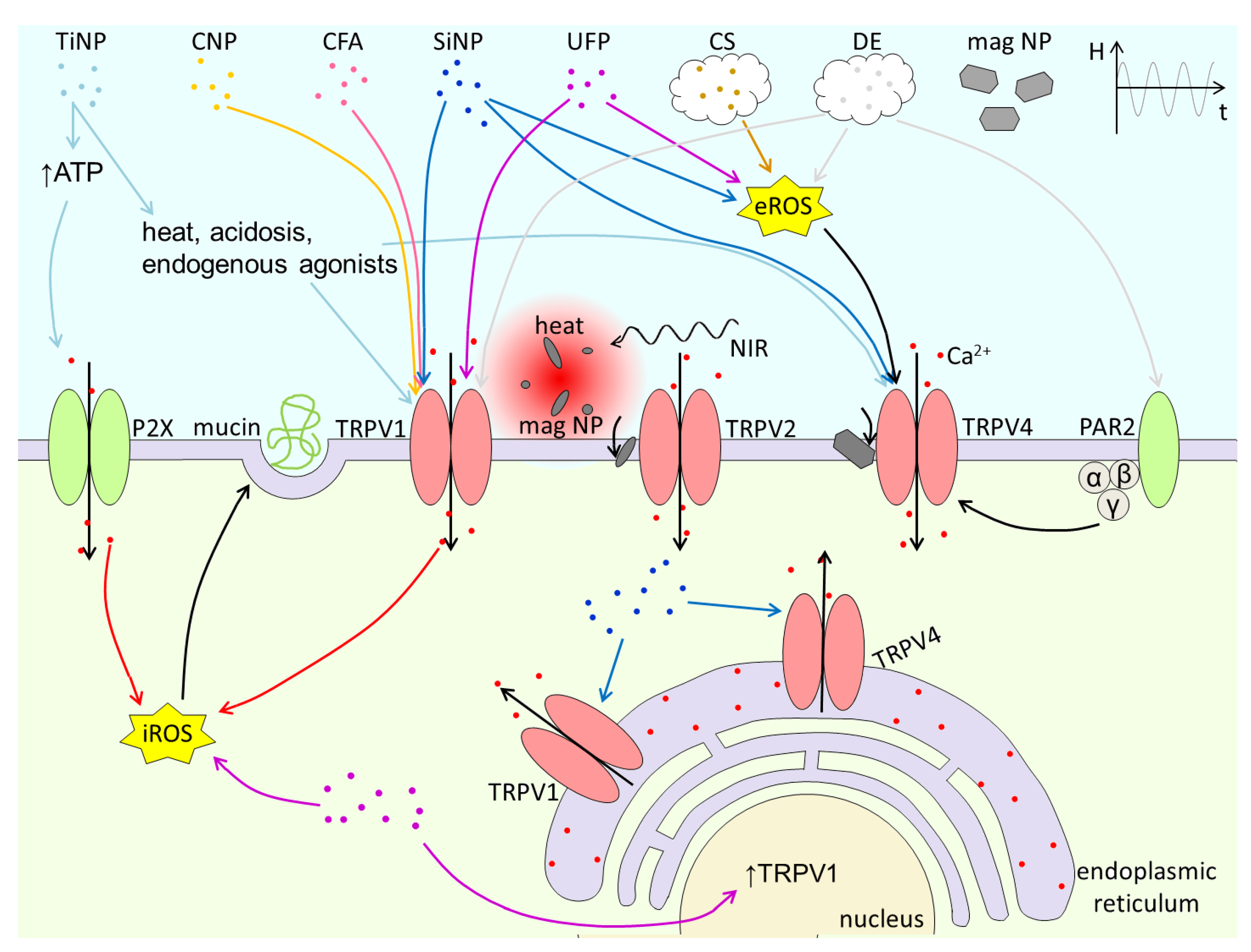{kind=link}
| Type of PM | Target | Effects | Observations |
|---|---|---|---|
| Ambient | General effect |
| |
| TRPA1 |
|
| |
| exposure | |||
| |||
| |||
| TRPM2 |
| ||
| |||
| |||
| |||
| TRPV1 |
| ||
| |||
| |||
| |||
| |||
| |||
| Bacterial | TRPV4 |
| |
| Carbon | TRPC4/TRPC6 |
| |
| TRPV1 |
| ||
| Cigarette smoke | General effect |
| |
| |||
| TRPA1 |
| ||
| |||
| |||
| |||
| |||
| |||
| |||
| TRPM2 |
| ||
| TRPM7 |
| ||
| TRPV1 |
|
| |
| TRPV4 |
|
| |
| |||
| Coal fly ash | General effect |
| |
| |||
| |||
| TRPA1 |
| ||
| TRPM8 |
| ||
| TRPV1 |
|
| |
| Diesel exhaust | General effect |
| |
| TRPA1 |
|
| |
| TRPM8 |
| ||
| TRPV1 |
| ||
| TRPV4 |
| ||
| Gold | TRPM2 |
| |
| TRPV1 |
| ||
| Iron | TRPV1 |
| |
| TRPV2 |
| ||
| TRPV4 |
| ||
| Lanthanide | TRPM2 |
| |
| Lipid | TRPM8 |
| |
| TRPV1 |
| ||
| PAMAM | TRPA1 |
| |
| TRPM2 |
| ||
| TRPM8 |
| ||
| TRPV4 |
| ||
| PLGA | TRPML1 |
| |
| TRPV4 |
| ||
| Semiconductor | TRPV1 |
| |
| Silica | TRPA1 |
| |
| TRPC6 |
| ||
| TRPM2 |
| ||
| |||
| TRPM4 |
| ||
| TRPM8 |
|
| |
| TRPV1 |
|
| |
| TRPV4 |
| ||
| Titanium | General effect |
| |
| TRPV1 |
| ||
| TRPV4 |
| ||
| Wood smoke | General effect |
| |
| TRPA1 |
|
| |
| TRPV3 |
|
| |
| Zinc | General effect |
| |
| TRPA1 |
| ||
| TRPM2 |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milici, A.; Talavera, K. TRP Channels as Cellular Targets of Particulate Matter. Int. J. Mol. Sci. 2021, 22, 2783. https://doi.org/10.3390/ijms22052783
Milici A, Talavera K. TRP Channels as Cellular Targets of Particulate Matter. International Journal of Molecular Sciences. 2021; 22(5):2783. https://doi.org/10.3390/ijms22052783
Chicago/Turabian StyleMilici, Alina, and Karel Talavera. 2021. "TRP Channels as Cellular Targets of Particulate Matter" International Journal of Molecular Sciences 22, no. 5: 2783. https://doi.org/10.3390/ijms22052783
APA StyleMilici, A., & Talavera, K. (2021). TRP Channels as Cellular Targets of Particulate Matter. International Journal of Molecular Sciences, 22(5), 2783. https://doi.org/10.3390/ijms22052783