
Delivery of Orally Administered Digestible Antibodies Using Nanoparticles

Abstract
:1. Introduction
2. Discussion
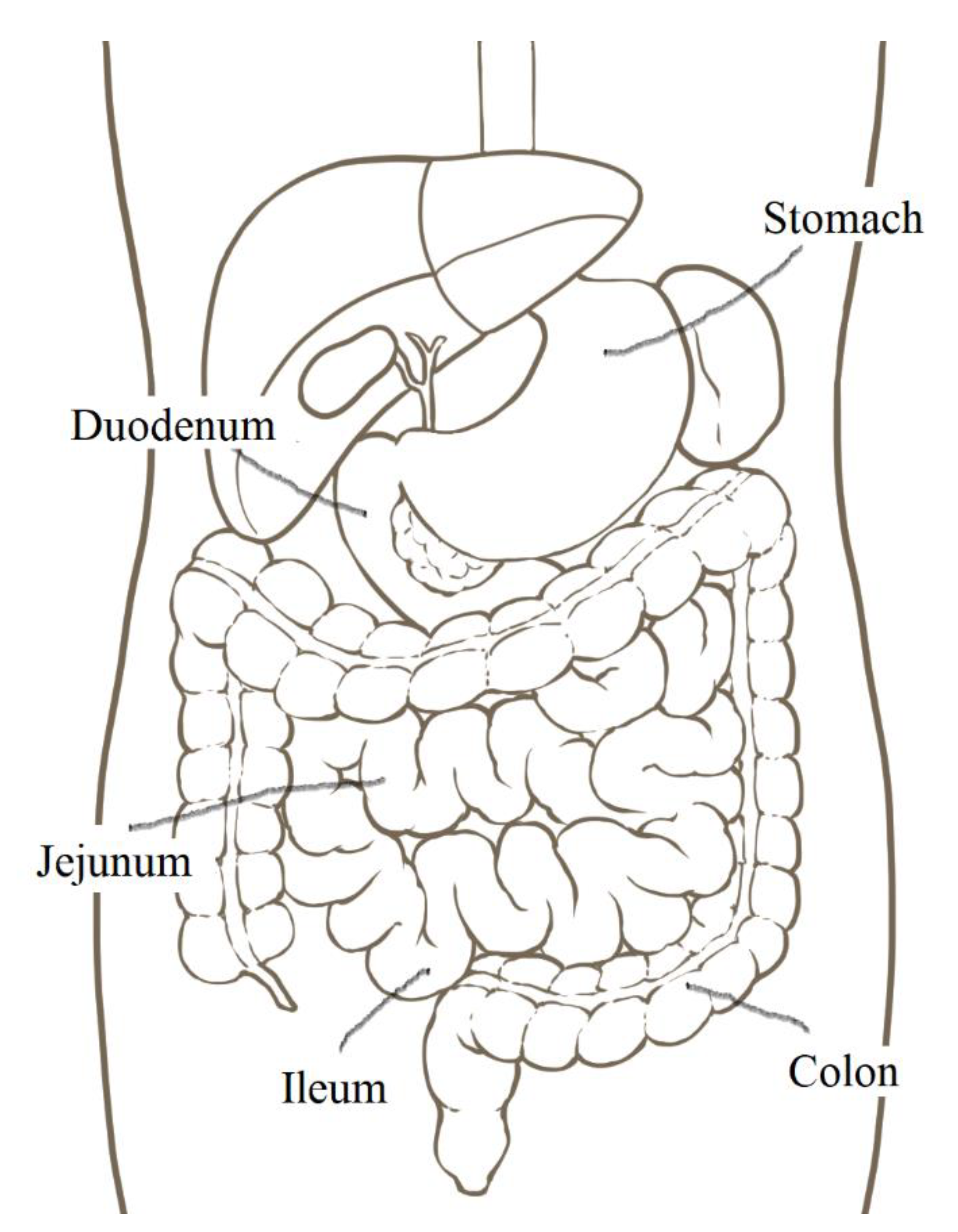
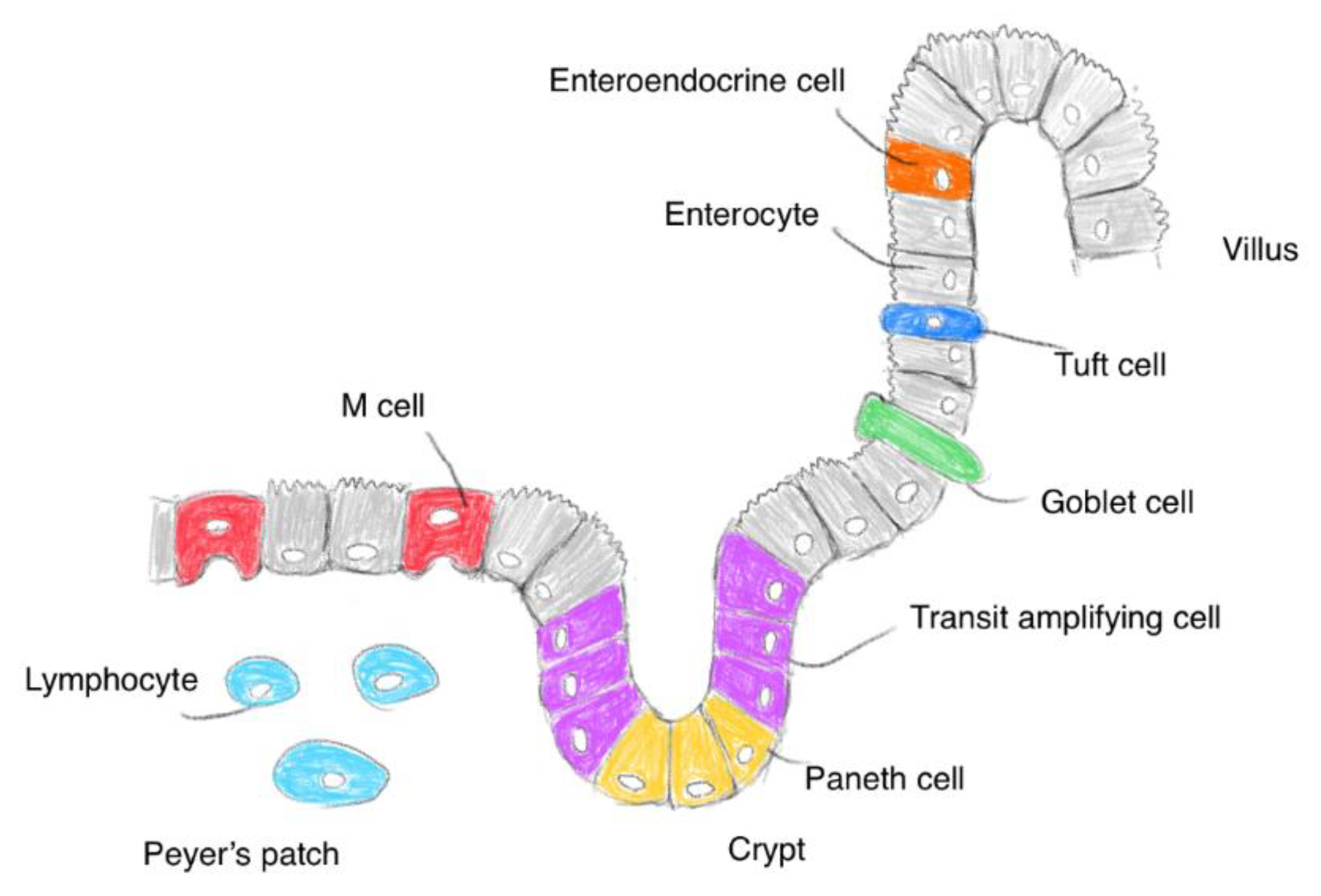
2.1. The Small Intestinal Epithelium
2.2. Viruses Entering the Intestinal Epithelium
2.3. Rise of Intravenous Antibody Drugs on the Market
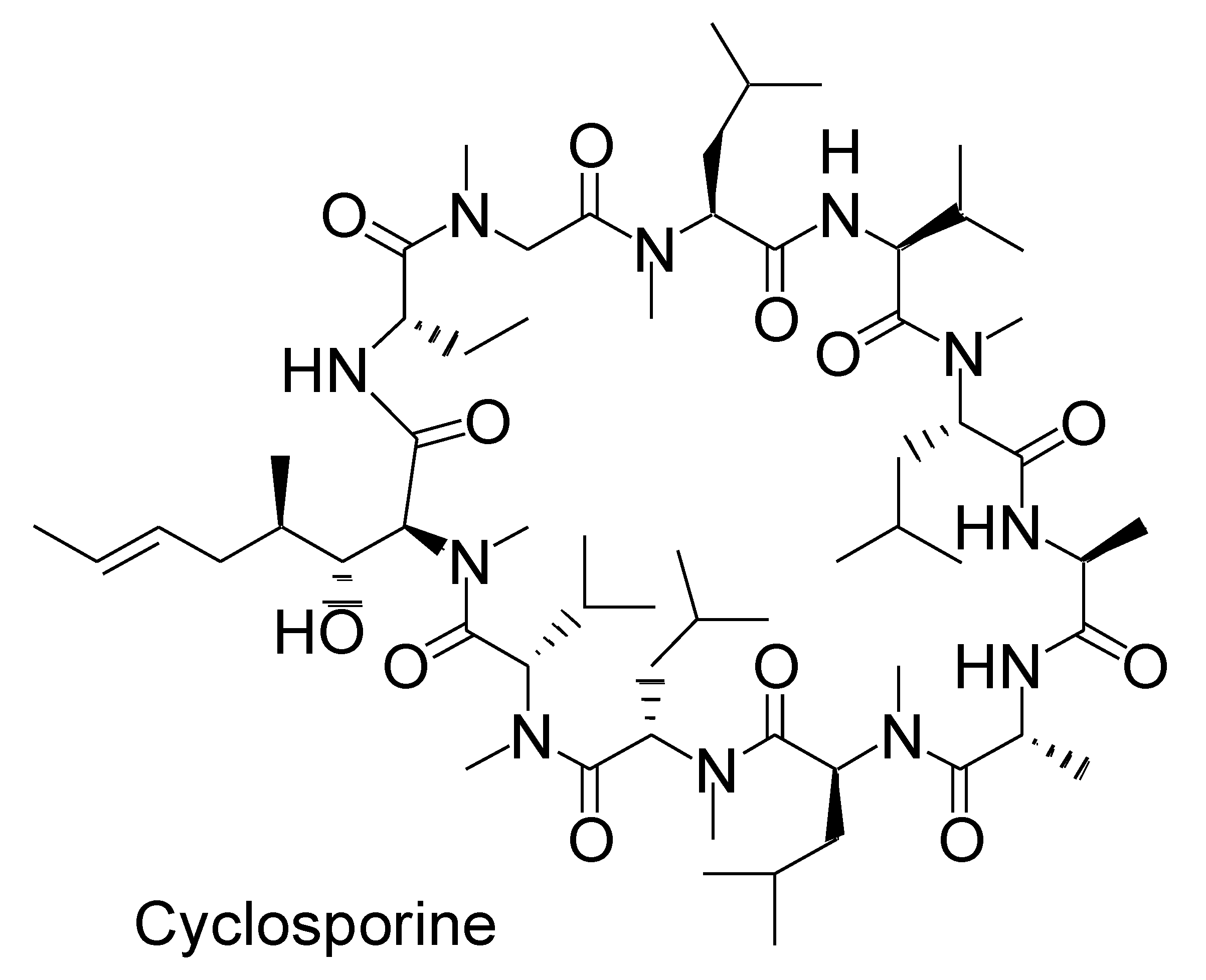
2.4. pH-Dependent Dissolution of Nanoparticles and Subsequent Peptide Cargo Release
2.5. Orally Administered mAbs for Localized Effect in Gut
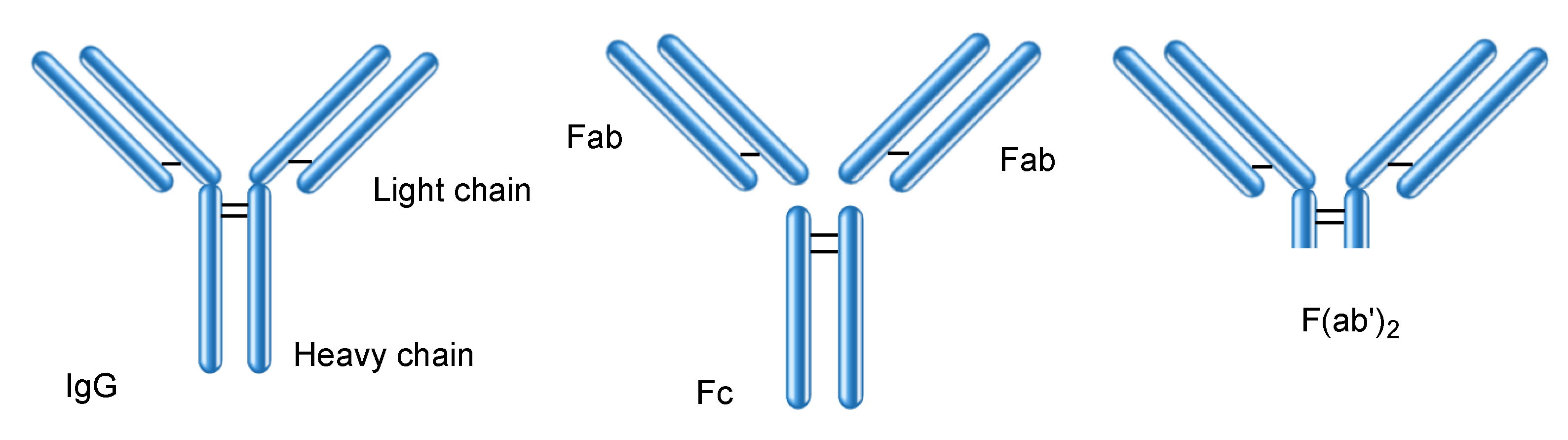
2.6. Proteins Interacting with IgG
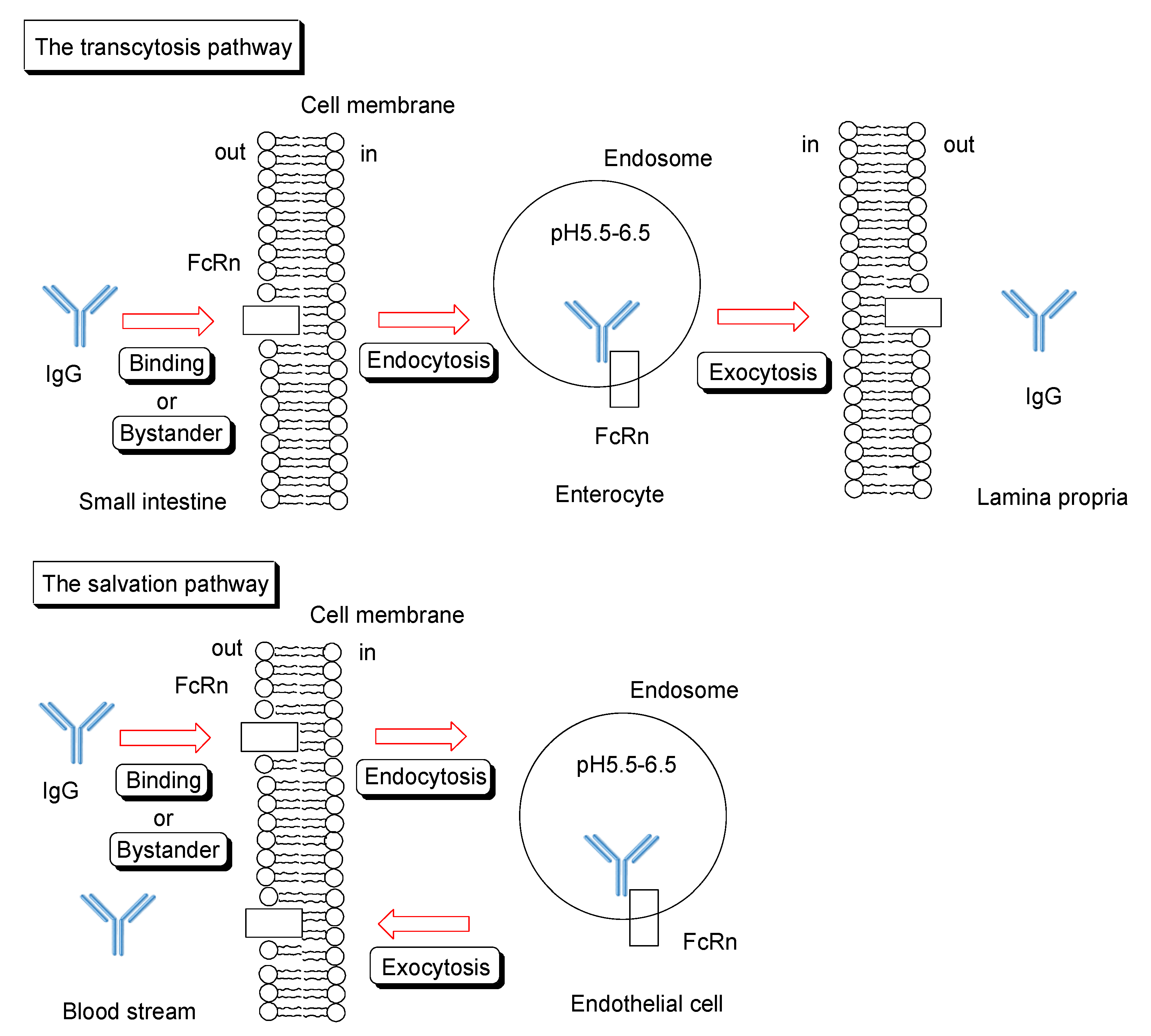
2.6.1. FcRn for Salvation and Transcytosis
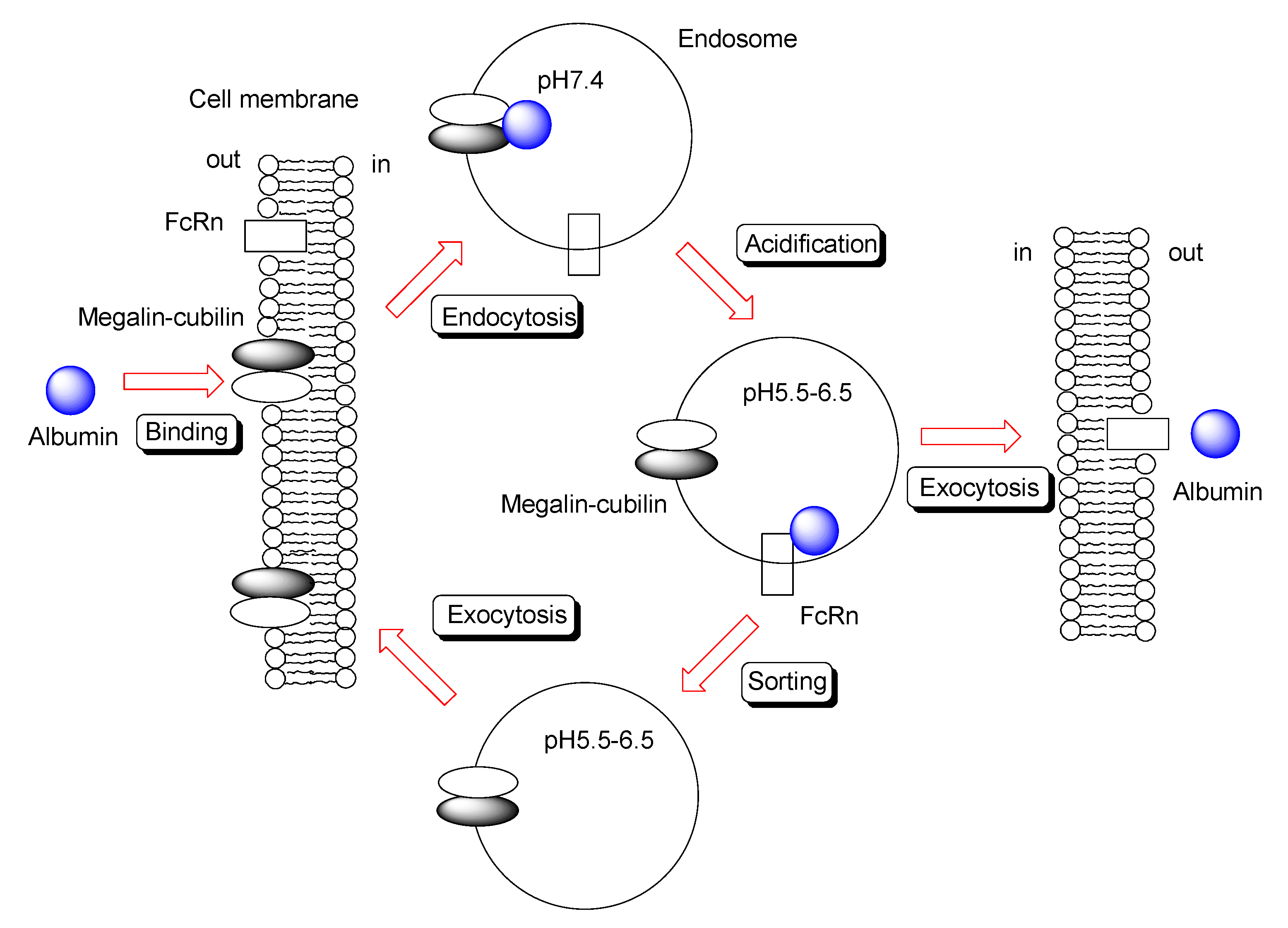
2.6.2. Trafficking of IgG and Albumin by FcRn
2.6.3. IgG Transepithelium through RMT Based on Translational Substance Exchange
2.7. Possible Methods for Applying Orally Administered mAbs for Systemic Treatment across the Epithelium
2.7.1. Oral mAb Delivery Using Nanoparticle through Endocytosis Based on Megalin-Cubilin
2.7.2. Oral mAb Delivery Using Nanoparticles through Endocytosis Based on FcRn
2.7.3. Oral mAb Delivery Using Nanoparticles through Pinocytosis Based on FcRn
2.7.4. Oral mAb Delivery Using Nanoparticles through Transcytosis in M Cells
2.7.5. Bio-Betters and Bio-Superiors
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hua, S. Advances in Oral Drug Delivery for Regional Targeting in the Gastrointestinal Tract—Influence of Physiological, Pathophysiological and Pharmaceutical Factors. Front. Pharmacol. 2020, 11, 524. [Google Scholar] [CrossRef]
- Ibrahim, Y.H.-E.Y.; Regdon, G., Jr.; Hamedelniel, E.I.; Sovány, T. Review of recently used techniques and materials to improve the efficiency of orally administered proteins/peptides. DARU J. Pharm. Sci. 2020, 28, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.-J.; Xu, S.; Wang, H.-M.; Ling, Y.; Dong, J.; Xia, R.-D.; Sun, X.-H. Nanoparticles: Oral Delivery for Protein and Peptide Drugs. AAPS PharmSciTech 2019, 20, 190. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Cui, P.; Chen, B.; Li, S.; Guan, H. Monoclonal antibodies: Formulations of marketed products and recent advances in novel delivery system. Drug Dev. Ind. Pharm. 2017, 43, 519–530. [Google Scholar] [CrossRef]
- Tashima, T. Intriguing possibilities and beneficial aspects of transporter-conscious drug design. Bioorg. Med. Chem. 2015, 23, 4119–4131. [Google Scholar] [CrossRef]
- Tashima, T. Intelligent substance delivery into cells using cell-penetrating peptides. Bioorg. Med. Chem. Lett. 2017, 27, 121–130. [Google Scholar] [CrossRef]
- Tashima, T. Effective cancer therapy based on selective drug delivery into cells across their membrane using receptor-mediated endocytosis. Bioorg. Med. Chem. Lett. 2018, 28, 3015–3024. [Google Scholar] [CrossRef]
- Tashima, T. Smart Strategies for Therapeutic Agent Delivery into Brain across the Blood-Brain Barrier Using Receptor-Mediated Transcytosis. Chem. Pharm. Bull. 2020, 68, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Tashima, T. Shortcut Approaches to Substance Delivery into the Brain Based on Intranasal Administration Using Nanodelivery Strategies for Insulin. Molecules 2020, 25, 5188. [Google Scholar] [CrossRef]
- Reilly, R.M.; Domingo, R.; Sandhu, J. Oral delivery of antibodies. Future pharmacokinetic trends. Clin. Pharmacokinet. 1997, 32, 313–323. [Google Scholar] [CrossRef]
- Nakamura, Y.; Mimuro, H.; Kunisawa, J.; Furusawa, Y.; Takahashi, D.; Fujimura, Y.; Kaisho, T.; Kiyono, H.; Hase, K. Microfold cell-dependent antigen transport alleviates infectious colitis by inducing antigen-specific cellular immunity. Mucosal Immunol. 2020, 13, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Szabóová, R.; Faixová, Z.; Maková, Z.; Piešová, E. The Difference in the Mucus Organization between the Small and Large Intestine and Its Protection of Selected Natural Substances. A Review. Folia Vet. 2018, 62, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; O’Leary, C.E.; Locksley, R.M. Regulation of immune responses by tuft cells. Nat. Rev. Immunol. 2019, 19, 584–593. [Google Scholar] [CrossRef]
- Karandikar, U.C.; Crawford, S.E.; Ajami, N.J.; Murakami, K.; Kou, B.; Ettayebi, K.; Papanicolaou, G.A.; Jongwutiwes, U.; Perales, M.-A.; Shia, J.; et al. Detection of human norovirus in intestinal biopsies from immunocompromised transplant patients. J. Gen. Virol. 2016, 97, 2291–2300. [Google Scholar] [CrossRef]
- Green, K.Y.; Kaufman, S.S.; Nagata, B.M.; Chaimongkol, N.; Kim, D.Y.; Levenson, E.A.; Tin, C.M.; Yardley, A.B.; Johnson, J.A.; Barletta, A.B.F.; et al. Human Norovirus Targets Enteroendocrine Epithelial Cells in the Small Intestine. Nat. Commu. 2020, 11, 2759. [Google Scholar] [CrossRef]
- Murakami, K.; Tenge, V.R.; Karandikar, U.C.; Lin, S.-C.; Ramani, S.; Ettayebi, K.; Crawford, S.E.; Zeng, X.-L.; Neill, F.H.; Ayyar, B.V.; et al. Bile acids and ceramide overcome the entry restriction for GII.3 human norovirus replication in human intestinal enteroids. PNAS 2020, 117, 1700–1710. [Google Scholar] [CrossRef]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef]
- Nielsen, D.S.; Shepherd, N.E.; Xu, W.; Lucke, A.J.; Stoermer, M.J.; Fairlie, D.P. Orally Absorbed Cyclic Peptides. Chem. Rev. 2017, 117, 8094–8128. [Google Scholar] [CrossRef]
- Hu, Z.; Nizzero, S.; Goel, S.; Hinkle, L.E.; Wu, X.; Li, C. Ferrari M, Shen H Molecular targeting of FATP4 transporter for oral delivery of therapeutic peptide. Sci. Adv. 2020, 6, eaba0145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Yao, W.D.; Rao, Y.F.; Lu, X.Y.; Gao, J.Q. pH-Responsive carriers for oral drug delivery: Challenges and opportunities of current platforms. Drug Deliv. 2017, 24, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Cai, Y.; Xu, Q.; Chen, Z.W. Atomic force bio-analytics of polymerization and aggregation of phycoerythrin-conjugated immunoglobulin G molecules. Mol. Immunol. 2004, 41, 1247–1252. [Google Scholar] [CrossRef] [Green Version]
- Crowe, J.S.; Roberts, K.J.; Carlton, T.M.; Maggiore, L.; Cubitt, M.F.; Clare, S.; Harcourt, K.; Reckless, J.; MacDonald, T.T.; Ray, K.P.; et al. Preclinical Development of a Novel, Orally-Administered Anti-Tumour Necrosis Factor Domain Antibody for the Treatment of Inflammatory Bowel Disease. Sci. Rep. 2018, 8, 4941. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.S.; Hartman, D.; Lemos, B.R.; Erlich, E.C.; Spence, S.; Kennedy, S.; Ptak, T.; Pruitt, R.; Vermeire, S.; Fox, B.S. AVX-470, an Orally Delivered Anti-Tumour Necrosis Factor Antibody for Treatment of Active Ulcerative Colitis: Results of a First-in-Human Trial. J. Crohn’s Colitis 2016, 10, 631–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, E.K.; Canavan, J.B.; Moran, C.J.; McCann, K.; Dunn, W.A.; Farraye, F.A.; Ananthakrishnan, A.N.; Yajnik, V.; Gandhi, R.; Nguyen, D.D.; et al. Immunologic Alterations Associated with Oral Delivery of Anti-CD3 (OKT3) Monoclonal Antibodies in Patients with Moderate-to-Severe Ulcerative Colitis. Crohn’s Colitis 360 2019, 1, otz009. [Google Scholar] [CrossRef] [PubMed]
- Ilan, Y.; Shailubhai, K.; Sanyal, A. Immunotherapy with oral administration of humanized anti-CD3 monoclonal antibody: A novel gut-immune system-based therapy for metaflammation and NASH. Clin. Exp. Immunol. 2018, 193, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Lalazar, G.; Mizrahi, M.; Turgeman, I.; Adar, T.; Ya’acov, A.B.; Shabat, Y.; Nimer, A.; Hemed, N.; Zolotarovya, L.; Lichtenstein, Y.; et al. Oral Administration of OKT3 MAb to Patients with NASH, Promotes Regulatory T-cell Induction, and Alleviates Insulin Resistance: Results of a Phase IIa Blinded Placebo-Controlled Trial. J. Clin. Immunol. 2015, 35, 399–407. [Google Scholar] [CrossRef]
- Wood, R.A.; Kim, J.S.; Lindblad, R.; Nadeau, K.; Henning, A.K.; Dawson, P.; Plaut, M.; Sampson, H.A. A randomized, double-blind, placebo-controlled study of omalizumab combined with oral immunotherapy for the treatment of cow’s milk allergy. J. Allergy Clin. Immunol. 2016, 137, 1103–1110. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Gadir, A.; Schneider, L.; Casini, A.; Charbonnier, L.-M.; Little, S.V.; Harrington, T.; Umetsu, D.T.; Rachid, R.; Chatila, T.A. Oral immunotherapy with omalizumab reverses the Th2 cell-like programme of regulatory T cells and restores their function. Clin. Exp. Allergy 2018, 48, 825–836. [Google Scholar] [CrossRef]
- Handen, B.L.; Melmed, R.D.; Hansen, R.L.; Aman, M.G.; Burnham, D.L.; Bruss, J.B.; McDougle, C.J. A double-blind, placebo-controlled trial of oral human immunoglobulin for gastrointestinal dysfunction in children with autistic disorder. J. Autism Dev. Disord. 2009, 39, 796–805. [Google Scholar] [CrossRef] [Green Version]
- Martyushev-Poklad, A.; Bruhwyler, J.; Heijmans, S.; Thiry, M. Efficacy of a Novel Antibody TLR3 Modulator in the Self-Treatment of Common Cold: The ESTUAR Trial. Adv. Infect. Dis. 2015, 5, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Savarino, S.J.; McKenzie, R.; Tribble, D.R.; Porter, C.K.; O’Dowd, A.; Sincock, S.A.; Poole, S.T.; DeNearing, B.; Woods, C.M.; Kim, H.; et al. Hyperimmune Bovine Colostral Anti-CS17 Antibodies Protect Against Enterotoxigenic Escherichia coli Diarrhea in a Randomized, Doubled-Blind, Placebo-Controlled Human Infection Model. J. Infect. Dis. 2019, 220, 505–513. [Google Scholar] [CrossRef]
- Savarino, S.J.; McKenzie, R.; Tribble, D.R.; Porter, C.K.; O’Dowd, A.; Cantrell, J.A.; Sincock, S.A.; Poole, S.T.; DeNearing, B.; Woods, C.M.; et al. Prophylactic Efficacy of Hyperimmune Bovine Colostral Antiadhesin Antibodies Against Enterotoxigenic Escherichia coli Diarrhea: A Randomized, Double-Blind, Placebo-Controlled, Phase 1 Trial. J. Infect. Dis. 2017, 216, 7–13. [Google Scholar] [CrossRef]
- Utay, N.S.; Somasunderam, A.; Hinkle, J.E.; Petschow, B.W.; Detzel, C.J.; Somsouk, M.; Fichtenbaum, C.J.; Weaver, E.M.; Shaw, A.L.; Asmuth, D.M. Serum Bovine Immunoglobulins Improve Inflammation and Gut Barrier Function in Persons with HIV and Enteropathy on Suppressive ART. Pathog. Immun. 2019, 4, 124–146. [Google Scholar] [CrossRef] [PubMed]
- Thiry, M.; Martyushev-Poklad, A. Pharmaceutical Compositions of Antibodies for Treatment of Diseases Caused by Influenza Virus. US Patent 9,409,991 B1, 9 August 2016. [Google Scholar]
- Shailubhai, K. Anti-CD3 Formulations. U.S. Patent 2018/0,177,880 A1, 28 June 2018. [Google Scholar]
- Pyzik, M.; Sand, K.M.K.; Hubbard, J.J.; Andersen, J.T.; Sandlie, I.; Blumberg, R.S. The Neonatal Fc Receptor (FcRn): A Misnomer? Front. Immunol. 2019, 10, 1540. [Google Scholar] [CrossRef]
- Patel, D.D.; Bussel, J.B. Neonatal Fc receptor in human immunity: Function and role in therapeutic intervention. J. Allergy Clin. Immunol. 2020, 146, 467–478. [Google Scholar] [CrossRef]
- Dylewski, J.; Dobrinskikh, E.; Lewis, L.; Tonsawan, P.; Miyazaki, M.; Jat, P.S.; Blaine, J. Differential trafficking of albumin and IgG facilitated by the neonatal Fc receptor in podocytes in vitro and in vivo. PLoS ONE 2019, 14, e0209732. [Google Scholar] [CrossRef] [Green Version]
- Vidarsson, G.; Stemerding, A.M.; Stapleton, N.M.; Spliethoff, S.E.; Janssen, H.; Rebers, F.E.; Haas, M. d.; Winkel, J.G. v.d. FcRn: An IgG receptor on phagocytes with a novel role in phagocytosis. Blood 2006, 108, 3573–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdiche, Y.N.; Yeung, Y.A.; Chaparro-Riggers, J.; Barman, I.; Strop, P.; Chin, S.M.; Pham, A.; Bolton, G.; McDonough, D.; Lindquist, K.; et al. The neonatal Fc receptor (FcRn) binds independently to both sites of the IgG homodimer with identical affinity. MAbs 2015, 7, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrok, M.J.; Wu, Y.; Beyaz, N.; Yu, X.-Q.; Oganesyan, V.; Dall’Acqua, W.F.; Tsui, P. pH-dependent binding engineering reveals an FcRn affinity threshold that governs IgG recycling. J. Biol. Chem. 2015, 290, 4282–4290. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.G.W.; Hvam, M.L.; Antunes, F.; Cameron, J.; Viuff, D.; Andersen, B.; Kristensen, N.N.; Howard, K.A. Direct demonstration of a neonatal Fc receptor (FcRn)-driven endosomal sorting pathway for cellular recycling of albumin. J. Biol. Chem. 2017, 292, 13312–13322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyzik, M.; Rath, T.; Lencer, W.I.; Baker, K.; Blumberg, R.S. FcRn: The Architect Behind the Immune and Nonimmune Functions of IgG and Albumin. J. Immunol. 2015, 194, 4595–4603. [Google Scholar] [CrossRef] [Green Version]
- Birn, H.; Fyfe, J.C.; Jacobsen, C.; Mounier, F.; Verroust, P.J.; Orskov, H.; Willnow, T.E.; Moestrup, S.K.; Christensen, E.I. Cubilin is an albumin binding protein important for renal tubular albumin reabsorption. J. Clin. Investig. 2000, 105, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Carretero, M.D.; Palomo, M.; García-Miranda, P.; Sánchez-Aguayo, I.; Peral, M.J.; Calonge, M.L.; Ilundain, A.A. Dab2, megalin, cubilin and amnionless receptor complex might mediate intestinal endocytosis in the suckling rat. J. Cell. Biochem. 2014, 115, 510–522. [Google Scholar] [CrossRef]
- Nagai, J.; Sato, K.; Yumoto, R.; Takano, M. Megalin/cubilin-mediated uptake of FITC-labeled IgG by OK kidney epithelial cells. Drug Metab. Pharmacokinet. 2011, 26, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Christensen, E.I.; Birn, H. Megalin and cubilin: Multifunctional endocytic receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 258–267. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 23 January 2021).
- Harrison, P.T.; Davis, W.; Norman, J.C.; Hockaday, A.R.; Allen, J.M. Binding of monomeric immunoglobulin G triggers FcγRI-mediated endocytosis. J. Biol. Chem. 1994, 269, 24396–24402. [Google Scholar] [CrossRef]
- Kiyoshi, M.; Caaveiro, J.M.M.; Kawai, T.; Tashiro, S.; Ide, T.; Asaoka, Y.; Hatayama, K.; Tsumoto, K. Structural basis for binding of human IgG1 to its high-affinity human receptor FcγRI. Nat. Commun. 2015, 6, 6866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbero, G.J.; Runge, G.; Fischer, D.; Crawford, M.N.; Torres, F.E.; György, P. Investigations on the bacterial flora, pH, and sugar content in the intestinal tract of infants. J. Pediatr. 1952, 40, 152–163. [Google Scholar] [CrossRef]
- Klassen, R.B.; Allen, P.L.; Batuman, V.; Crenshaw, K.; Hammond, T.G. Light chains are a ligand for megalin. J. Appl. Physiol. 2005, 98, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Gerbal-Chaloin, S.; Gondeau, C.; Aldrian-Herrada, G.; Heitz, F.; Gauthier-Rouvière, C.; Divita, G. First step of the cell-penetrating peptide mechanism involves Rac1 GTPase-dependent actin-network remodeling. Biol. Cell 2007, 99, 223–238. [Google Scholar] [CrossRef]
- Fujii, M.; Kawai, K.; Egami, Y.; Araki, N. Dissecting the roles of Rac1 activation and deactivation in macropinocytosis using microscopic photo-manipulation. Sci. Rep. 2013, 3, 2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, E.S.; Ober, R.J. Chapter 4: Multitasking by exploitation of intracellular transport functions the many faces of FcRn. Adv. Immunol. 2009, 103, 77–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohl, W.R. Current progress in innovative engineered antibodies. Protein Cell 2018, 9, 86–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Administrated Drug | Formulation/Co-Administrated Drug | Disease | Sponsor | Phase | Study Start Date | Study Completion Date | ClinicalTrials.gov Identifier (accessed on 23 January 2021) | Status | References |
|---|---|---|---|---|---|---|---|---|---|---|
| (i) | Oral V565 | Crohn’s disease | VHsquared Ltd. | Phase 2 | December 2016 | December 2018 | NCT02976129 | Unknown | - | |
| (ii) | Oral V565 | Capsule | Ulcerative colitis | VHsquared Ltd. | Phase 1 | October 2017 | October 2017 | NCT03705117 | Completed | - |
| (iii) | Oral AVX-470 | Enteric-coated capsule | Ulcerative colitis | Avaxia Biologics, Incorporated | Phase 1 | February 2013 | December 2013 | NCT01759056 | Completed | [23] |
| (iv) | Oral muromonab-CD3 | Omeprazole | Ulcerative colitis | Brigham and Women’s Hospital | Phase 1 Phase 2 | April 2011 | May 2013 | NCT01287195 | Completed | [24] |
| (v) | Oral foralumab | Omeprazole | NASH, NAFLD, Type 2 diabetes mellitus | Tiziana Life Sciences, PLC | Phase 2 | December 2017 | June 2019 | NCT03291249 | Withdrawn | [25] |
| (vi) | Oral muromonab-CD3 | NASH | Hadassah Medical Organization | Phase 2 | September 2010 | April 2011 | NCT01205087 | Completed | [26] | |
| (vii) | Oral anti-CD3 mAb | Omeprazole | Chronic hepatitis C | Inspira Medical AB | Phase 2 | November 2011 | October 2013 | NCT01459419 | Unknown | - |
| (viii) | Oral omalizumab | Milk allergy | Hugh A Sampson, MD | Phase 2 | August 2010 | October 2015 | NCT01157117 | Completed | [27] | |
| (ix) | Oral omalizumab | Peanut Allergy | Lynda Schneider | Phase 1Phase 2 | February 2011 | September 2013 | NCT01290913 | Completed | [28] | |
| (x) | Subcutaneous omalizumab | Peanut Allergy, Multi-food Allergy | National Institute of Allergy and Infectious Diseases | Phase 3 | July 2019 | December 2023 | NCT03881696 | Recruiting | - | |
| (xi) | Oral AGY | Capsule | Celiac disease | Igy Inc. | Phase 1 | May 2014 | August 2015 | NCT01765647 | Completed | - |
| (xii) | Oral AGY | Capsule | Celiac disease | Igy Inc. | Phase 2 | October 2019 | December 2022 | NCT03707730 | Recruiting | - |
| (xiii) | Oral Oralgam | Autistic Disorder, Gastrointestinal Diseases | PediaMed Pharmaceuticals | Phase 2 | April 2005 | June 2006 | NCT00110708 | Unknown | [29] | |
| (xiv) | Oral TAO1 | Tablet | Common Cold | Theranor s.p.r.l | Phase 1Phase 2 | September 2012 | August 2013 | NCT01651715 | Completed | [30] |
| (xv) | Oral anti-influenza antibody | Tablet | Influenza | Hadassah Medical Organization | Phase 1 | January 2010 | January 2011 | NCT01026350 | Unknown | - |
| (xvi) | Oral anti-CsbD bovine IgG, Oral anti-CS17 bovine IgG | Diarrhea | Johns Hopkins Bloomberg School of Public Health | Phase 2 | January 2007 | October 2007 | NCT00524004 | Completed | [31] | |
| (xvii) | Oral anti-CFA/I bovine IgG, Oral anti-CfaE bovine IgG | Diarrhea | Johns Hopkins Bloomberg School of Public Health | Phase 1 | March 2006 | October 2006 | NCT00435526 | Completed | [32] | |
| (xviii) | Oral SBI | Mucositis | MercyOne Des Moines Medical Center | Phase 2 | January 2020 | December 2021 | NCT04239261 | Recruiting | - | |
| (xix) | Oral SBI | HIV-associated enteropathy | Entera Health, Inc | Not applicable | April 2013 | September 2014 | NCT01828593 | Completed | [33] | |
| (xx) | Oral SBI | Female reproductive cancer | Mayo Clinic | Phase 2 | October 2013 | October 2021 | NCT01867606 | Active, not recruiting | - | |
| (xxi) | Oral SBI | Advanced COPD with cachexia | Medical University of South Carolina | Not Applicable | December 2013 | April 2016 | NCT02067377 | Completed | - | |
| (xxii) | Oral SBI | Diarrhea-predominant irritable bowel syndrome | Mayo Clinic | Not Applicable | June 2014 | November 2016 | NCT02163213 | Completed | - | |
| (xxiii) | Oral SBI | Irritable bowel syndrome | Louisiana State University Health Sciences Center in New Orleans | Not Applicable | March 2017 | March 2017 | NCT02609529 | Completed | - | |
| (xxiv) | Oral antibodies in colostrum | Clostridium difficile-associated diarrhea | Hadassah Medical Organization | Phase 2Phase 3 | September 2011 | November 2013 | NCT00747071 | Withdrawn | - | |
| (xxv) | Oral mAbs | Enteric nanoparticles | Systemic treatment | Under analysis in Tashima lab | - |
| # | Protein (Gene Name) | Protein Expression | Level | RNA Expression | NX Values |
|---|---|---|---|---|---|
| (i) | FcRn (FCGRT) | Cerebellum | Medium | Granulocytes | 164.3 |
| Cerebral cortex | Low | Small intestine | 125.2 | ||
| Hippocampus | Low | Colon | 104.2 | ||
| Caudate | Low | Monocytes | 100.2 | ||
| Lung | Low | Liver | 87.4 | ||
| Testis | Low | Total PBMC | 72.1 | ||
| Heart muscle | Low | Dendritic cells | 70.7 | ||
| (ii) | Megalin/LRP2 (LRP2) | Kidney | High | Kidney | 61.7 |
| Parathyroid gland | Medium | Parathyroid gland | 61.4 | ||
| Testis | Medium | Placenta | 10.5 | ||
| Placenta | Low | Small intestine | 0.4 | ||
| Small intestine | Not detected | ||||
| (iii) | Cubilin (CUBN) | Kidney | High | Kidney | 79.7 |
| Small intestine | Low | Small intestine | 28.9 | ||
| Epididymis | 11.5 | ||||
| (iv) | FcγRI (FCGR1A, CD64A) | not shown | Monocytes | 35.6 | |
| Epididymis | 33.2 | ||||
| Granulocytes | 25.5 | ||||
| Small intestine | 0.1 | ||||
| (v) | FcγRI (FCGR1B, CD64B) | not shown | Epididymis | 36.9 | |
| Granulocytes | 21.8 | ||||
| Monocytes | 19.2 | ||||
| Small intestine | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tashima, T. Delivery of Orally Administered Digestible Antibodies Using Nanoparticles. Int. J. Mol. Sci. 2021, 22, 3349. https://doi.org/10.3390/ijms22073349
Tashima T. Delivery of Orally Administered Digestible Antibodies Using Nanoparticles. International Journal of Molecular Sciences. 2021; 22(7):3349. https://doi.org/10.3390/ijms22073349
Chicago/Turabian StyleTashima, Toshihiko. 2021. "Delivery of Orally Administered Digestible Antibodies Using Nanoparticles" International Journal of Molecular Sciences 22, no. 7: 3349. https://doi.org/10.3390/ijms22073349
APA StyleTashima, T. (2021). Delivery of Orally Administered Digestible Antibodies Using Nanoparticles. International Journal of Molecular Sciences, 22(7), 3349. https://doi.org/10.3390/ijms22073349