3. Discussion
In this study we demonstrated that GPR110 ligands stimulate axon growth in axotomized cortical neurons and retinal explants in culture and after optic nerve injury in adult mice. The axon growth after injury was facilitated through the rapid induction of gpr110 and thus the upregulating of ligand-activated GPR110/cAMP/PKA signaling, even in adult neurons.
Developing neurons can extend their axons after injury, but such regenerative ability is lost with maturity [
15]. A contributing factor to this regeneration failure is the loss of rapid axon growth capacity [
16], which is under developmental control [
17]. GPR110, the target receptor of synaptamide, mediates the neurogenic, neuritogenic, and synaptogenic activity in developing neurons through the activation of cAMP/PKA signaling [
18]. Like numerous genes that undergo dynamic changes in their expression during development [
19],
gpr110 is highly expressed in neural stem cells and prenatal brains, but diminishes after birth [
10], suggesting that the expression of
gpr110 in the brain is also under developmental control. Therefore, the upregulation of GPR110 signaling, a developmental mechanism of stimulated neurite growth, may be a viable strategy for promoting axonal growth after nerve injury in adulthood.
Injury is known to trigger a cascade of interrelating cellular and molecular events and affects the expression of numerous genes for not only degeneration but also reorganization and repair processes [
20,
21,
22]. In the RGC layer of an adult retina,
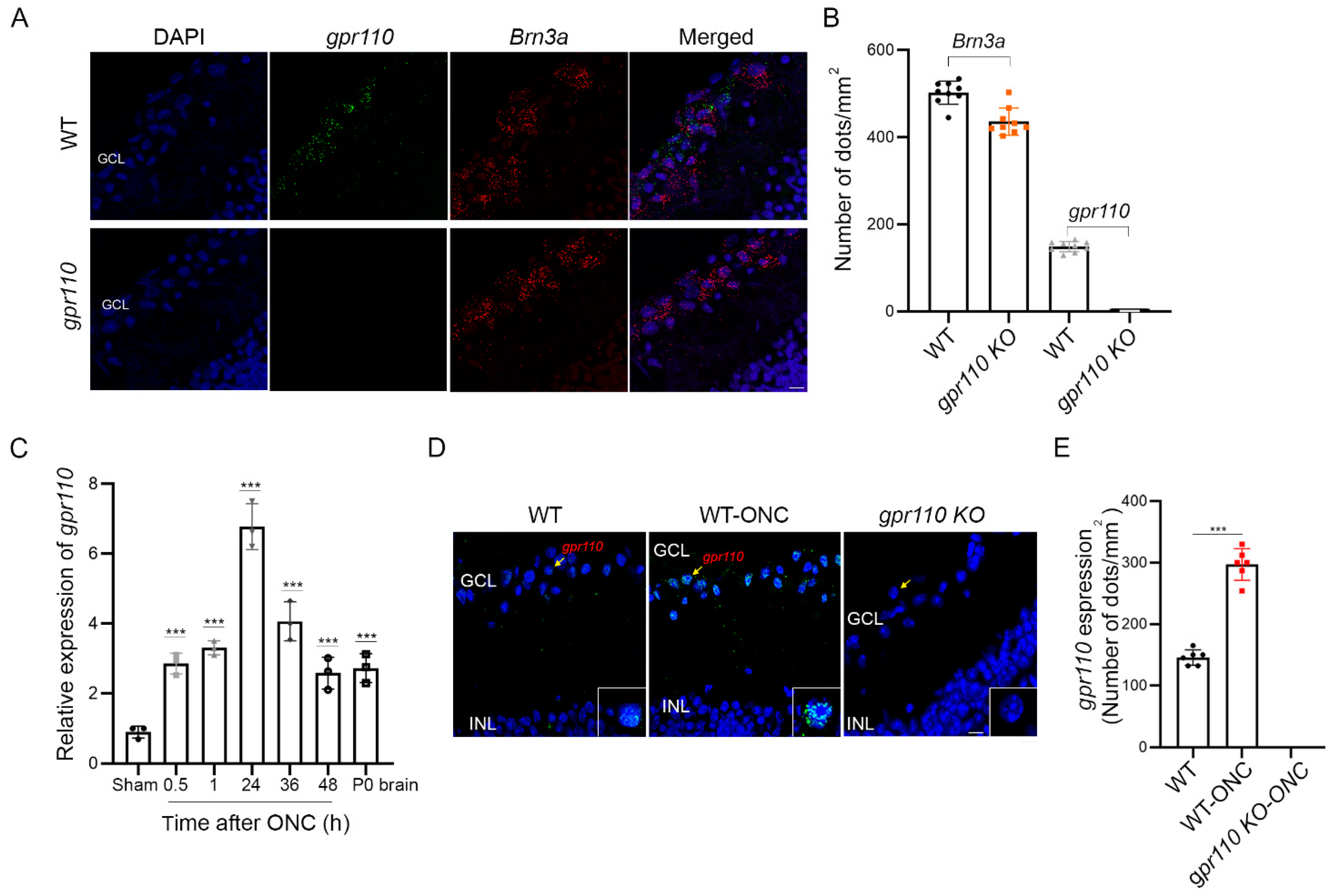
gpr110 expression is low, but is significantly elevated within 30 min after optic nerve injury (
Figure 4C–E). In developing cortical neurons, rapid increase in
gpr110 expression was also apparent after axotomy (
Figure 2G). The rapid injury-response nature of this developmental gene for stimulated neurite growth may be an intrinsic regenerative mechanism for facilitating the repair of injured axons. Indeed, activating GPR110 with its endogenous ligand stimulated axon growth after injury both in vitro (
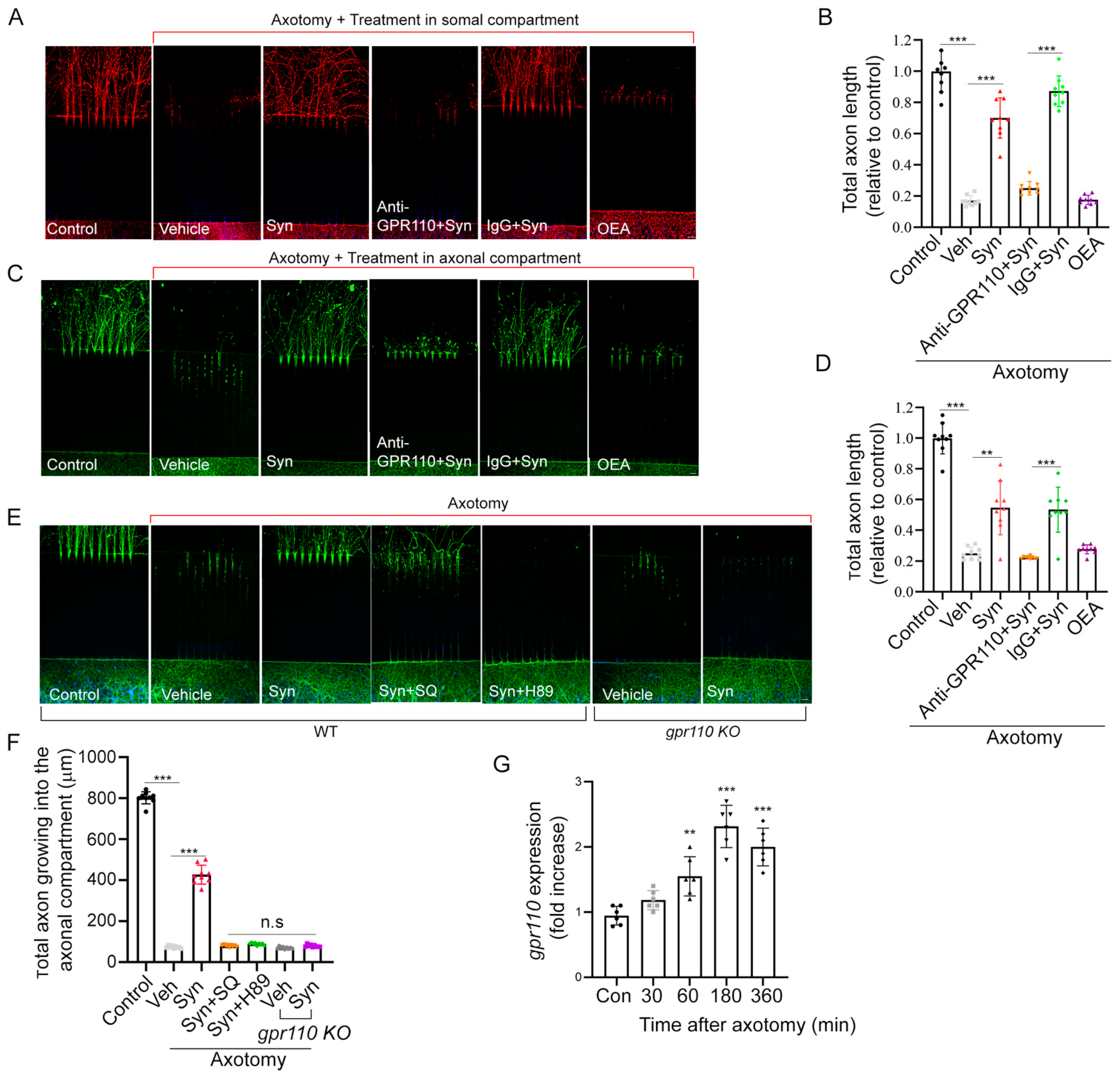
Figure 2) and in vivo, as shown for the axotomized cortical neuron culture (
Figure 2) and crushed optic nerves (
Figure 5).
The neurite extension observed upon synaptamide application to the axonal compartment after axotomy (
Figure 2C,D) indicates the stimulated growth of injured axons in developing neurons. It has been shown that neuronal regenerative capacity can be regulated by cAMP/PKA signaling [
23]. The endogenous cAMP level is much higher in neonatal neurons compared to mature neurons, and this change in cAMP level can effectively switch the glia-derived signal from stimulatory to inhibitory for axon growth [
23]. When damaged CNS axons are first primed with neurotrophins before encountering inhibitory substrates derived from CNS glia around the injury site, axonal regeneration is not inhibited and the regeneration is cAMP- and PKA-dependent [
24]. It is possible that upregulating cAMP/PKA signaling by specifically targeting GPR110 at an early stage after injury allows the injured axons to overcome inhibitory signals and to extend beyond the injury site in the adult CNS.
The synaptamide level in most cases correlates with the tissue level of its precursor, DHA. In the rodent brain, the DHA content is at 14–16% of the total fatty acids, while the synaptamide level is in the range of 6–30 pmol/g tissue [
18]. The human optic nerve contains DHA at about 1.5% of total fatty acids [
25], suggesting that the synaptamide level in the optic nerve would be substantially lower than that observed in the brain. Although the endogenous synaptamide level in the optic nerve is unknwon, it is likely insufficient to properly activate GPR110, despite the receptor upregulation after injury. Exogenously administering GPR110 ligands may have effectively activated GPR110 and downstream cAMP signaling, allowing injured axons to overcome inhibitory signals for regeneration.
It is well-established that ONC produces mild axonal injury without affecting ocular blood flow, and the survival of greater than 40% of RGCs can be achieved, depending on the severity of crush [
26]. It has been also reported that axon degeneration does not require RGC somal loss. It is possible that the activation of GPR110/cAMP signaling stimulates the sprouting of new neurites from the spared soma of traumatized neurons in addition to the regenerative capacity of injured axons. The rapid induction of GPR110 in RGCs after injury may be an intrinsic mechanism for enabling stimulated axon growth through GPR110/cAMP signaling.
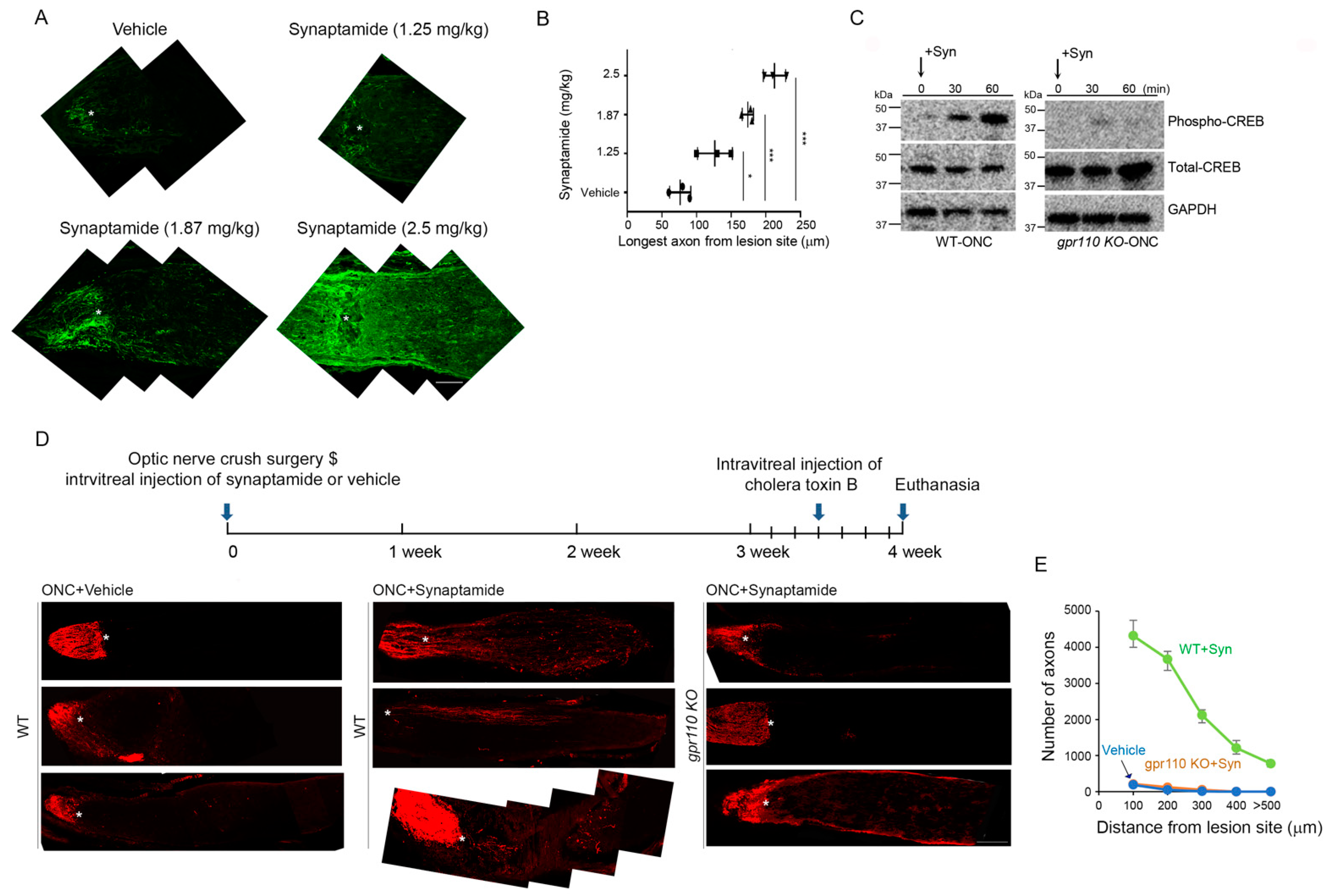
The effectiveness of synaptamide observed with a single treatment after ONC suggests that synaptamide action in an early stage of injury is important in stimulating axon growth and/or preserving axon regeneration capacity. In fact, GPR110-dependent increase in CREB phosphorylation was detected 30 min after synaptamide injection (
Figure 5D). Similarly,
gpr110 was quickly upregulated within 30 min after injury and remained upregulated even after 48 h (
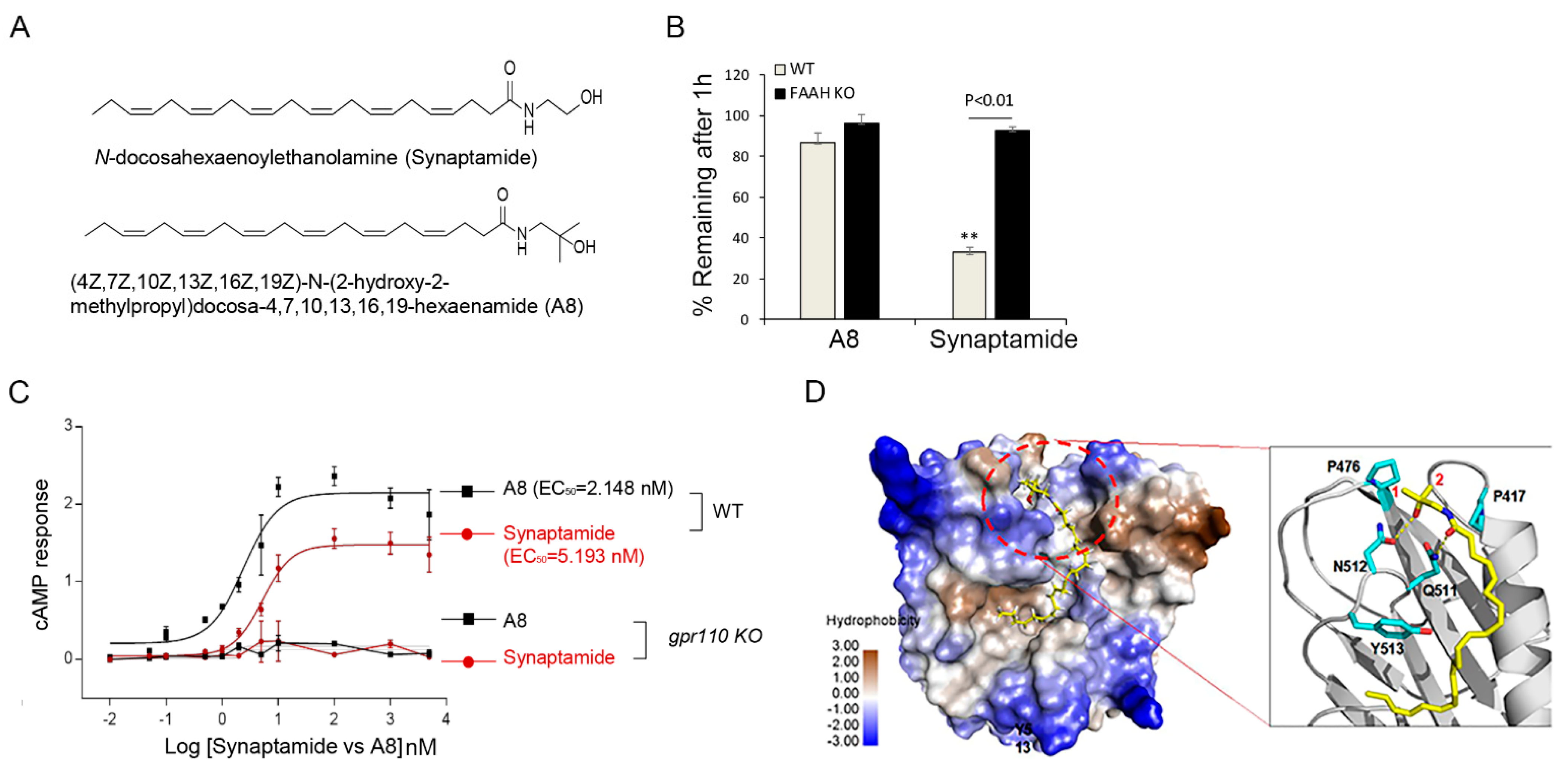
Figure 4C). A8, a FAAH-resistant stable analogue of synaptmaide (
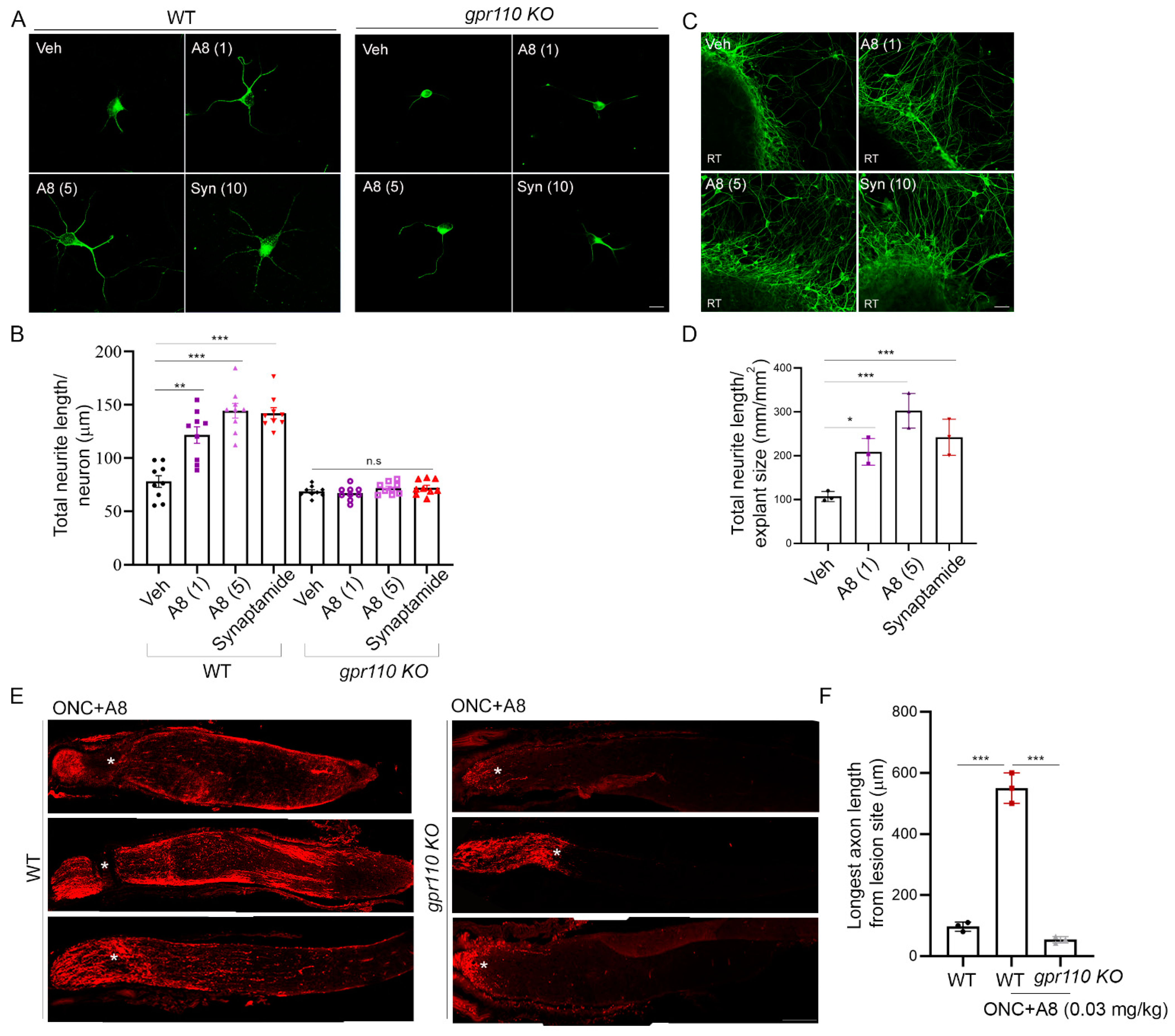
Figure 6A,B) with improved binding affinity, cAMP production, and neurite growth in vitro (
Figure 6C,D,
Figure 7A–D), stimulated optic nerve extension after injury at a dose nearly 100-fold less than synaptamide (
Figure 7E,F). GPR 110 is a member of the adhesion GPCRs that are recognized as emerging targets for drug discovery [
27,
28]. Further investigation is required to verify whether the observed GPR110-dependent stimulated axon growth can result in functional recovery. If so, GPR110 ligands such as A8 may present a new therapeutic opportunity for nerve injury,
In summary, our data indicate that GPR110 activation by synaptamide or its stable analogue A8 promotes axon growth in an in vitro injury model and in an optic nerve injury model in vivo. Our finding demonstrates a new potential for remedying axon injury by employing a mechanism for developmental neurite outgrowth, namely GPR110-mediated cAMP/PKA signaling. Synaptamide or A8 activates cAMP signaling in adult neurons at the injury site where the specific target receptor GPR110 is induced, and therefore can exert its bioactivity without the considerable side effects associated with systemic intervention of cAMP metabolism. We suggest that activating GPR110 may lead to a valuable strategy for stimulating axon growth after CNS injury in adults.
4. Materials and Methods
4.1. Animals
Timed pregnant female C57BL/6 mice were obtained from the NIH-NCI animal production program or Charles River Laboratories (Germantown, MD, USA), and GPR110 (adhesion G protein-coupled receptor F1: Adgrf1) heterozygous mice of a C57BL/6 background were generated by the Knockout Mouse Project (KOMP) Repository. Animals were housed in an SPF facility and acclimated for a day before their brains were collected for the preparation of primary cortical neurons or immunohistochemistry. gpr110 KO mice and matching WT were generated by heterozygote mating in our animal facility. All experiments were carried out in accordance with the guiding principles for the care and use of animals approved by the National Institute on Alcohol Abuse and Alcoholism (LMS-HK13).
4.2. Primary Cell Culture and Neurite Length Analysis
Primary cortical neurons were prepared from C57BL/6 mouse brains according to the established protocol [
10]. Briefly, cortices were isolated from P0 pups and digested with 100 U of papain for 30 min at 37 °C and mechanically disrupted by pipetting several times in neurobasal medium (Invitrogen, Carlsbad, CA, USA) supplemented with 2% B27 (Invitrogen) and a 1% glutamine/glutamax mixture (1:3) (Invitrogen). The dissociated cortical neurons were seeded in poly-D-lysine-coated 24-well plates (0.5 × 10
4 cells/well) or 8-well glass slide chambers (2 × 10
3 cells/well) for cAMP or neurite outgrowth analysis, respectively.
Neurons were stained for β-III tubulin (1:200, cat# 5568, Cell signaling, Danvers, MA, USA) and images were acquired using a Zeiss LSM 700 confocal laser-scanning microscope. The neurite length was measured using NIH Image J software (National Institutes of Health, Bethesda, MD, USA) as we described earlier [
10].
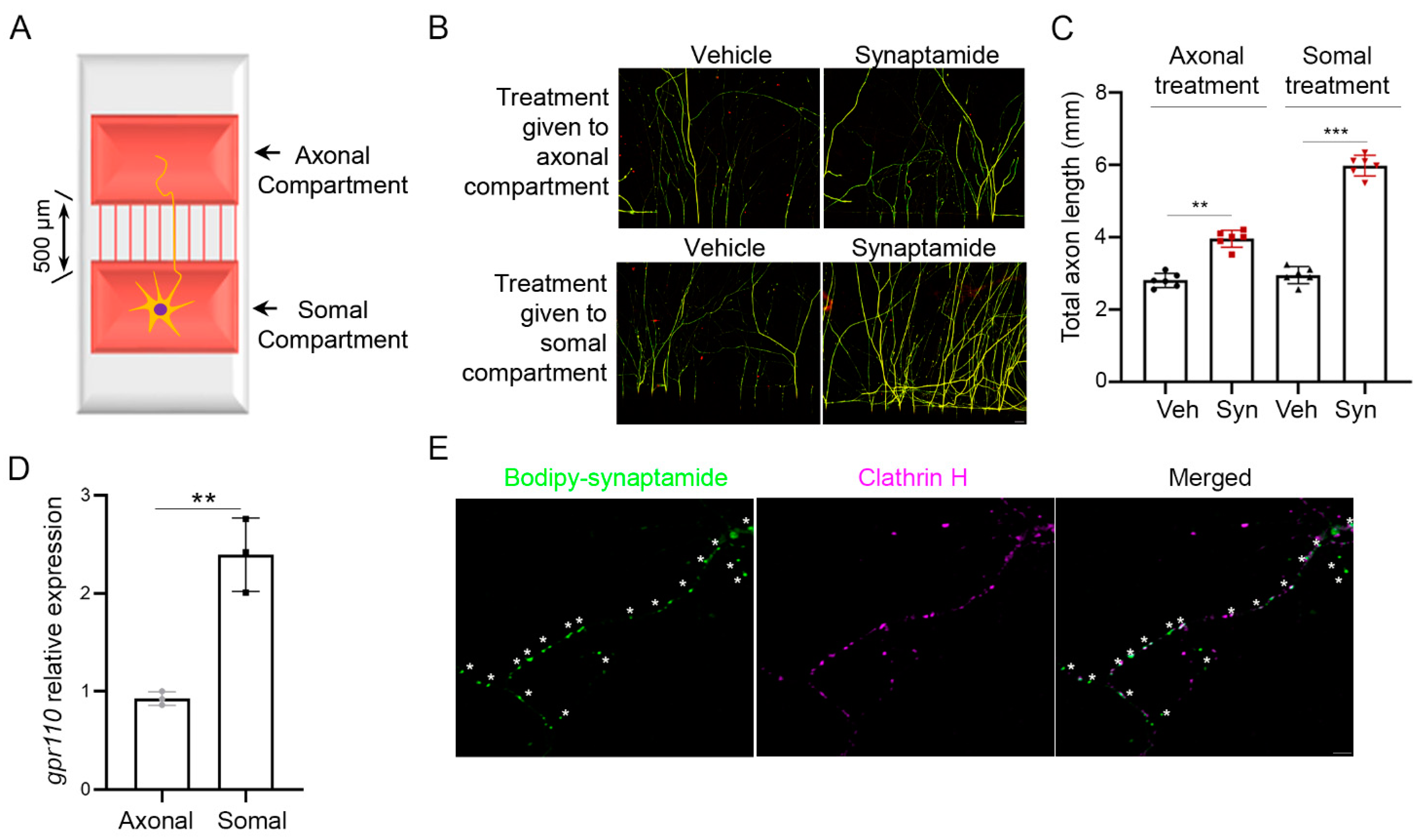
4.3. Cortical Neuron Culture in Microfluidic Compartment Chamber and In Vitro Axotomy
To evaluate the somal and axonal contribution to axon growth in intact or axotomized neurons, we used a microfluidic device with two-compartment chambers separated by 500 μm, as described previously [
29]. The chamber (kindly provided by Dr. In-Hong Yang at the Singapore Institute for Neurotechnology at the National University of Singapore) was uniformly coated with a mixture of polylysine (100 μg/mL) and laminin (10 μg/mL). Primary cortical neurons were loaded onto the cell body side of the device and treated with agents on DIV3 for an additional 4 days. The axonal or somal compartment was added with ligands with or without pretreatment with SQ22536 (10 μM, Sigma-Aldrich, St. Louis, MO, USA), H89 (10 μM, Sigma, CA, USA), or anti-GPR110 (0.4 mg/mL, Abmart, Berkley Heights, NJ, USA) at 60 min prior to the ligand addition. Compounds were introduced into the axonal chamber very slowly and carefully using a micropipette.
For in vitro axotomy, mouse primary cortical neurons were cultured for at least 7 days in the microfluidic chamber to allow the axons to grow sufficiently into the axonal compartment. Subsequently, axotomy was carried out by carefully applying suction with fine tip glass pipette positioned at the entrance of the axonal channel. Suction was maintained for approximately 3 s, or until all visible liquid in the axonal compartment was removed. The axonal compartment was then washed with media by placing the tip at the channel entrance and vigorously pipetting up and down to remove all detached axons and debris. Following this step, fresh medium was added to the compartment for washing and the microfluidic chamber was inspected under the microscope to ensure axotomy. Axotomized and non-axotomized control cultures were treated with agents and incubated for an additional 7 days for immunocytochemistry.
4.4. Quantitation of Axon Growth In Vitro
Images were acquired using a Zeiss LSM 700 confocal microscope (Zeiss, White Plains, NY, USA) and analyzed using Image J-neurite tracker software 1.34. Neurons in the microfluidic chamber were fixed by 4% paraformaldehyde and incubated with anti-β-III tubulin (1:200, Cell signaling) overnight followed by Alexa 488- or 555-labeled secondary antibody. Nuclei were counter-stained by DAPI (1:10,000, Thermo Fisher Scientific Inc., Waltham, MA, USA). Axons were recognized by β-III tubulin-positive fibers and quantified with NIH Image J-neurite tracker software 1.34 to show total axon length in a field. To rule out possible dendrite contamination in the microgrooves, we selected the cross-line between the ends of microgrooves and the axonal compartment as the reference line through the entire axonal compartment. For each experiment, at least three randomly selected axon fields were evaluated to represent each individual sample for quantification in a manner blinded to each culture condition.
4.5. Retinal Explant Cultures and Axon Length Measurements
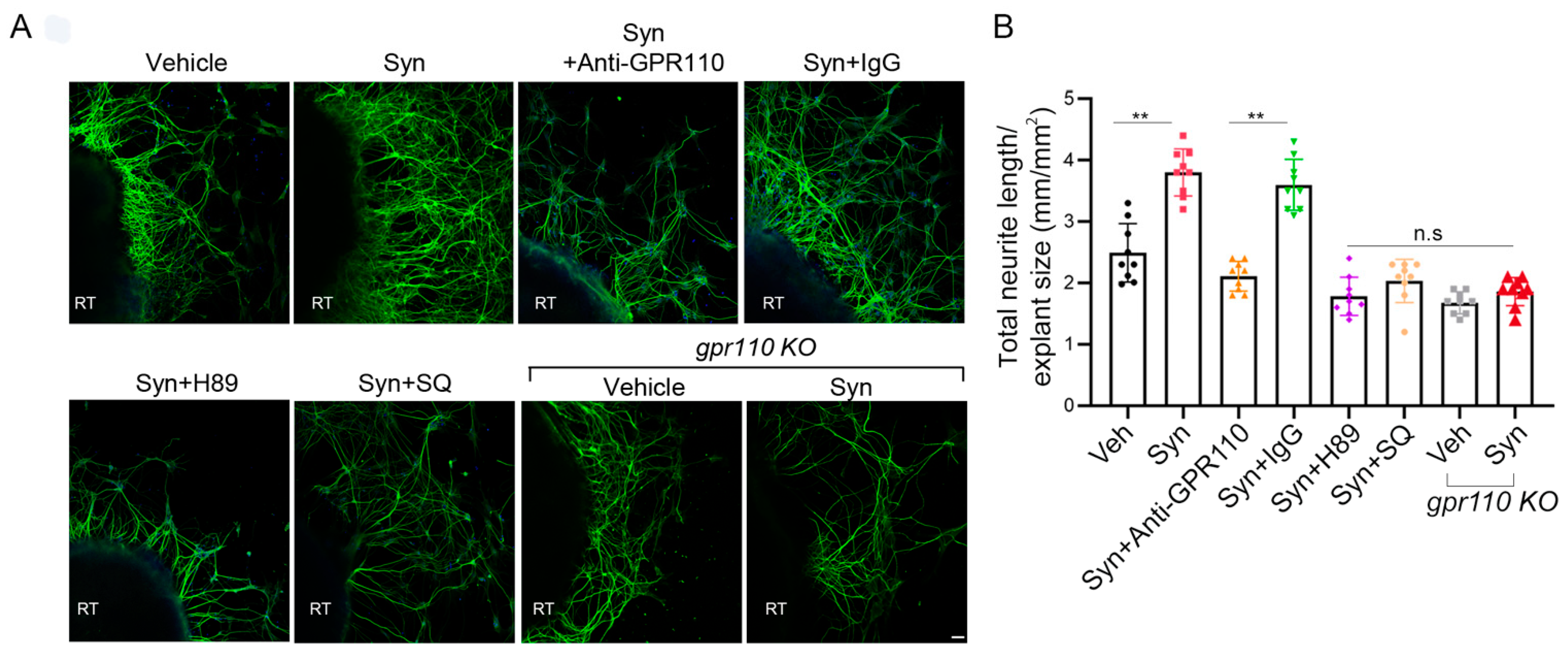
Retinal explant cultures were performed according to the procedure previously described [
30]. P0 mouse retina was dissected, and 3 explants were taken from each animal along the dorsal–ventral axis in the center of the nasotemporal axis. The dissected explants of retina tissues were placed onto poly-D-lysine/laminin coated glass cover slips that were treated with agents including vehicle (DMSO), synaptamide, or A8 on DIV 3. After 7 days, explants were fixed with 4% paraformaldehyde (PFA) and subsequently immunostained with antibody against β-III tubulin. Axon outgrowth was quantified using NIH Image J neurite tracker software 1.34 and the total outgrowth for each explant was normalized to the explant size. The final data are presented as the axon growth relative to control.
4.6. Synaptamide and A8 Application
Synaptamide and A8 were added according to our protocol as previously described [
10]. Primary cortical neuron or retina explant tissues were treated with synaptamide or A8 dissolved in DMSO in the presence of 40 μM vitamin E and 0.01% BSA (Sigma). As a control treatment, DMSO in medium containing BSA and vitamin E was used. In some cases, OEA in DMSO was also used as a lipid control. Anti-GPR110 (0.4 mg/mL, Santa Cruz, CA, USA), SQ22536 (Sigma), or H89 (Sigma) were added to the culture medium 60 min prior to the addition of synaptamide.
4.7. Optic Nerve Crush (ONC)
Mice were anesthetized by injection of ketamine (100 mg/kg, i.p.) and xylazine (10 mg/kg, i.p.). Under a binocular operating scope, a small incision was made with spring scissors (cat. #RS-5619; Roboz, Gaithersburg, MD, USA) in the conjunctiva, beginning inferior to the globe and around the eye temporally. Caution was taken, as making this cut too deep can result in cutting into the underlying musculature (inferior oblique, inferior rectus muscles inferiorly, or the lateral rectus temporally) or the supplying vasculature. With micro-forceps (Dumont #5/45 forceps, cat. #RS-5005; Roboz), the edge of the conjunctiva next to the globe was grasped and retracted, rotating the globe nasally. This exposed the posterior aspect of the globe, allowing for the visualization of the optic nerve. The exposed optic nerve was grasped approximately 1–3 mm from the globe with Dumont #N7 cross-action forceps (cat. #RS-5027; Roboz) for 3 s to apply pressure on the nerve by the self-clamping action. The Dumont cross-action forceps were chosen because its spring action applied a constant and consistent force to the optic nerve. During the 5 s clamping, we were able to observe mydriasis. After 5 s the pressure on the optic nerve was released and the forceps removed, allowing the eye to rotate back into place.
4.8. Intravitreal Administration of Synaptamide or A8
Synaptamide (2.5 mg/kg), A8 (0.03 mg/kg), or vehicle (DMSO) was intravitreally injected immediately after ONC while the mice were still under anesthesia. The experimenter was blinded to the identity of the compounds, including the vehicle. Before the injection of the compounds, PBS was applied to the cornea for cleaning purposes. A pulled glass micropipette attached to a 10 µL Hamilton syringe was used to deliver 2 µL of a solution into the vitreous chamber of the eye, posterior to the limbus. Care was taken to prevent damage to the lens. The pipette was held in place for 3 s after injection and slowly withdrawn from the eye to prevent reflux. Injections were performed under a surgical microscope to visualize pipette entry into the vitreous chamber and to confirm the delivery of the injected solution.
4.9. Anterograde Labeling
On the third day prior to euthanasia, the animals were injected with CTB–Alexa Fluor 555-conjugated cholera toxin subunit B (CTB, Life Technologies, Carlsbad, CA, USA) as an anterograde tracer to visualize axons in the optic nerve originating from RGCs.
4.10. Evaluation of Axon Growth after ONC
The micrographic images of the longitudinal optic nerve sections were obtained using a Zeiss LSM700 confocal microscope under 40Xagnification with automatic stitching and analyzed using Image J in a blinded fashion. Three longitudinal Z-stack sections were analyzed per each optic nerve tissue section and each experimental group included 4–6 mice. In some cases, the optic nerve micrographs were obtained after manually stitching the image sections using Photoshop. To analyze the extent of the axon growth after optic nerve injury, the number of the CTB-labeled regenerating axons (N) that passed through the measuring point at a specific distance (d) from the lesion site was estimated using the following formula as described earlier [
31]: σd =
πr2 × [average axon numbers per millimeter]/
t, where
r is half the cross-sectional width of the nerve at the counting site (d), the average number of axons per millimeter nerve width was from three nerve sections, and
t is the section thickness (12 μm). The number of the regenerating axons extended from the lesion site was determined at a 100 μm interval. The longest axon length was determined by analyzing the micrographs of two longitudinal optic nerve sections per animal, taken under 40X magnification using a Zeiss LSM700 confocal microscope (Zeiss).
4.11. RNA In Situ Hybridization
Perfused tissue sections (25 μm thickness) were processed for in situ RNA detection using the RNAscope Detection Kit (Fluorescent) (Newark, CA, USA) according to the manufacturer’s instructions (Advanced Cell Diagnostics, Hayward, CA, USA). Briefly, frozen tissue samples were placed on slides and incubated with Pretreat 1 buffer for 10 min at room temperature. Slides were boiled in Pretreat 2 buffer for 15 min, followed by incubation with Pretreat 3 buffer for 30 min at 40 °C. Slides were then incubated with the relevant probes for 2 h at 40 °C, followed by successive incubation with Amp 1 to 6 reagents. Staining was visualized with fluorescent multiplex reagents (Cat# 320851, RNA scope). The in situ probes used were gpr110 (NM_133776.2, cat# 300031), DapB (EF191515, cat # 310043), and Brn3 (NM_011143.4 cat# 414671).
4.12. Bodipy–Synaptamide Binding to GPR110
Bodipy–synaptamide puncta staining was performed as previously described [
10]. Briefly, cortical neurons were plated on 4-well glass slide chambers and treated with 100 nM bodipy–synaptamide (green) for 30 min and fixed with 4% paraformaldehyde for 10 min at room temperature. After washing, the cells were incubated with anti-clathrin H antibody (1:200, red), followed by Alexa-555-conjugated secondary antibody for 1 h at room temperature. Images were acquired on a Zeiss LSM 700 confocal laser-scanning microscope (Zeiss).
4.13. Immunostaining of Optic Nerve
At 4 weeks after ONC, tissues were collected after perfusion for immunostaining. Frozen sections (25 μm thickness) were prepared using a cryostat microtome (Leica) and fixed in 4% PBS-buffered polyformaldehyde solution, and permeabilized using 0.3% Triton-X 100 and 3% goat serum in PBS for immunostaining for GAP-43, a regenerating axon marker. Tissue samples were incubated with anti-GAP43 antibody (1:500, Cell Signaling) diluted in PBS containing 3% goat serum at 4 °C overnight. After washing 3 times with PBS, samples were incubated with Alexa Fluor 488 goat anti-rabbit (1:200) secondary antibody (Molecular Probes, Thermo Fisher Scientific Inc.) for 1 h at room temperature. Samples were mounted with fluoro mounting medium (Millipore, Carlsbad, CA, USA) and images were taken with a Zeiss LSM700 confocal microscope (Oberkochen, Germany).
4.14. Quantitative Polymerase Chain Reaction
One microgram of total RNA was reverse transcribed in the presence of 0.5 μg of Oligo(dT)15 primer (ABI, Foster, CA, USA) for 1 h at 37 °C using a reverse transcriptase kit (Cat# Qiagen, Germantown, USA) according to the manufacturer’s instructions. One microliter of first-strand cDNA was then subjected to PCR performed on ABI9700 Cycler (Foster, CA, USA) with the following conditions: 95 °C for 3 min (1 cycle), 95 °C for 30 s, 62 °C for 1 min, 72 °C for 30 s (30 cycles), and 72 °C for 10 min. RT-PCR products of the genes of interest in the wild-type and gpr110 KO mice were grouped and analyzed simultaneously. All reactions were repeated twice with different RT reactions. Relative expression levels were determined by normalization to the glyceraldehyde 3-phosphate dehydrogenase (gapdh) mRNA level using 2−ΔΔCt with logarithmic transformation.
The primers used in q RT-PCR analysis were: mouse Gpr110: forward, 5′-CCAAGAGAAGCCAAACCTCC-3′; reverse, 5′-TTCGATAAGCCAGCAGGATG-3′, and mouse Gapdh: forward, 5′-ACCACAGTCCATGCCATCAC-3′: reverse, 5′-CACCACCCTGTTGCTGTAGCC-3′.
4.15. cAMP Assay
cAMP analysis was performed as described previously [
10]. Primary cortical cells were cultured and treated on DIV3 with synaptamide or A8 for 10 min to determine cAMP levels using a cyclic AMP XP assay kit (Cell Signaling Technology, Danvers, MA, USA).
4.16. Western Blot Analysis
WT or gpr110 KO mice were perfused intracardially with phosphate-buffered saline (PBS) to clear the blood before the brains were removed. Retina and optic nerve tissues were dissected out and homogenized in ice-cold solubilization buffer (25 mM Tris pH 7.2, 150 mM NaCl, 1 mM CaCl2, 1 mM MgCl2) containing 0.5% NP-40 (Thermo Fisher Scientific, Waltham, MA, USA) and protease inhibitors (Sigma). The protein concentrations of the lysates were determined by micro BCA protein assay (Thermo Fisher Scientific). Samples for SDS-PAGE were prepared at 1 μg protein/μL concentration using 4X SDS-PAGE buffer (Thermo Fisher Scientific, Waltham, MA, USA) and 20 μg of protein was loaded onto each well. Proteins were separated by SDS-PAGE on 4–15% polyacrylamide gels (Thermo Fisher Scientific) and transferred onto a PVDF membrane (Thermo Fisher Scientific, Waltham, MA, USA). After treating with blocking buffer containing 0.01% Tween-20, 10% BSA (Thermo Fisher Scientific) for 1 h at room temperature, blots were incubated with primary antibody diluted in blocking buffer (anti-CREB, anti-phospho-CREB, rabbit anti-GAPDH 1:1000, Cell Signaling Technology) overnight at 4 °C followed by HPR-conjugated secondary antibodies (1:5000, Cell Signaling Technology) for 1 h at room temperature. The detection was performed using the KODAK Imaging system.
4.17. Preparation of A8 (4Z,7Z,10Z,13Z,16Z,19Z)-N-(2-Hydroxy-2-methylpropyl)docosa-4,7,10,13,16,19-hexaenamide (NCGC0024843)
100 mg (0.304 mmol) of (4Z,7Z,10Z,13Z,16Z,19Z)-docosa-4,7,10,13,16,19-hexaenoic acid (Enzo) and 32.6 mg (0.365 mmol) of 1-amino-2-methylpropan-2-ol (Matrix Scientific) were taken in a nitrogen filled dry 25 mL pear shaped flask charged with a magnetic stirrer. 2 mL dichloromethane (Sigma Aldrich) was added, followed by 127 mg (0.335 mmol) hexafluorophosphate azabenzotriazole tetramethyl uronium (HATU, Combi-Blocks) and 0.085 mL (0.609 mmol) triethylamine (Sigma Aldrich). The reaction was stirred for 10 min at room temperature and then diluted with more dichloromethane (~20 mL) washed with saturated aqueous ammonium chloride (~20 mL) and then saturated aqueous sodium bicarbonate. The dichloromethane layer was separated, dried with magnesium sulfate, filtered, and concentrated in vacuo. The residue was purified via flash silica gel chromatography (0 to 75% ethyl acetate in dichloromethane); fractions could be visualized by thin layer chromatography followed by staining with para-anisaldehyde stain or iodine. Fractions containing the product were combined, dried via rotary evaporation, and then under high vacuum to obtain 105 mg (0.263 mmol, 86% yield) of A8 which was protected from light and stored under nitrogen at −80 °C.
1H spectra were recorded on a Varian Inova 400 MHz spectrometer (Palo Alto, CA, USA). Chemical shifts are reported in ppm with the solvent resonance as the internal standard (MeOH-d4 3.31 ppm for Me). Data are reported as follows: chemical shift, multiplicity (s = singlet, d = doublet, t = triplet, q = quartet, m = multiplet), coupling constants, and number of protons, apparent multiplicity (if observed). 1H NMR (400 MHz, methanol-d4) δ 5.45–5.24 (m, 12H), 3.18 (s, 2H), 2.91–2.80 (m, 10H), 2.45–2.33 (m, 2H), 2.29 (ddd, J = 7.9, 6.9, 1.1 Hz, 2H, app triplet), 2.09 (dqdd, J = 7.5, 6.7, 1.4, 0.7 Hz, 2H, app pentet), 1.16 (s, 6H), 0.97 (t, J = 7.5 Hz, 3H). Analytical purity analysis and retention times (RT) reported here were performed on an Agilent LC/MS (Agilent Technologies, Santa Clara, CA, USA). A Phenomenex Luna C18 column (3 µ, 3 × 75 mm) was used at a temperature of 50 °C. The solvent gradients are mentioned for each compound and consist of a percentage of acetonitrile (containing 0.025% trifluoroacetic acid) in water (containing 0.05% trifluoroacetic acid). A 4.5 min run time at a flow rate of 1 mL/min was used. A8 was at >95% purity based on the above methods and had RT = 3.9 min. Mass determination was performed using an Agilent 6130 mass spectrometer with electrospray ionization (ESI) in the positive ion mode. Calculated mass for C26H42NO2+ (M+H)+ was 400.3, found 400.3.
4.18. Docking Studies
The 3D structure of GPR110 was generated using the I-TASSER program as described previously [
10,
11]. Docking of synaptamide and A8 to the GAIN domain of GPR110 was performed using the MOE program [
14]. The induced fit protocol was used for ligand docking, and the binding affinity was evaluated using the GBVI/WSA score [
14].
4.19. Statistical Analysis
Data were analyzed using GraphPad Prism 7 software (San Diego, CA, USA) and all data are presented as mean ± s.e.m and are representative of at least two independent experiments. Statistical significance was determined by an unpaired Student’s t test or one-way ANOVA. * p < 0.05, ** p < 0.01 and *** p < 0.001.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






