
Angiopoietin-Like Growth Factor Involved in Leptin Signaling in the Hypothalamus

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
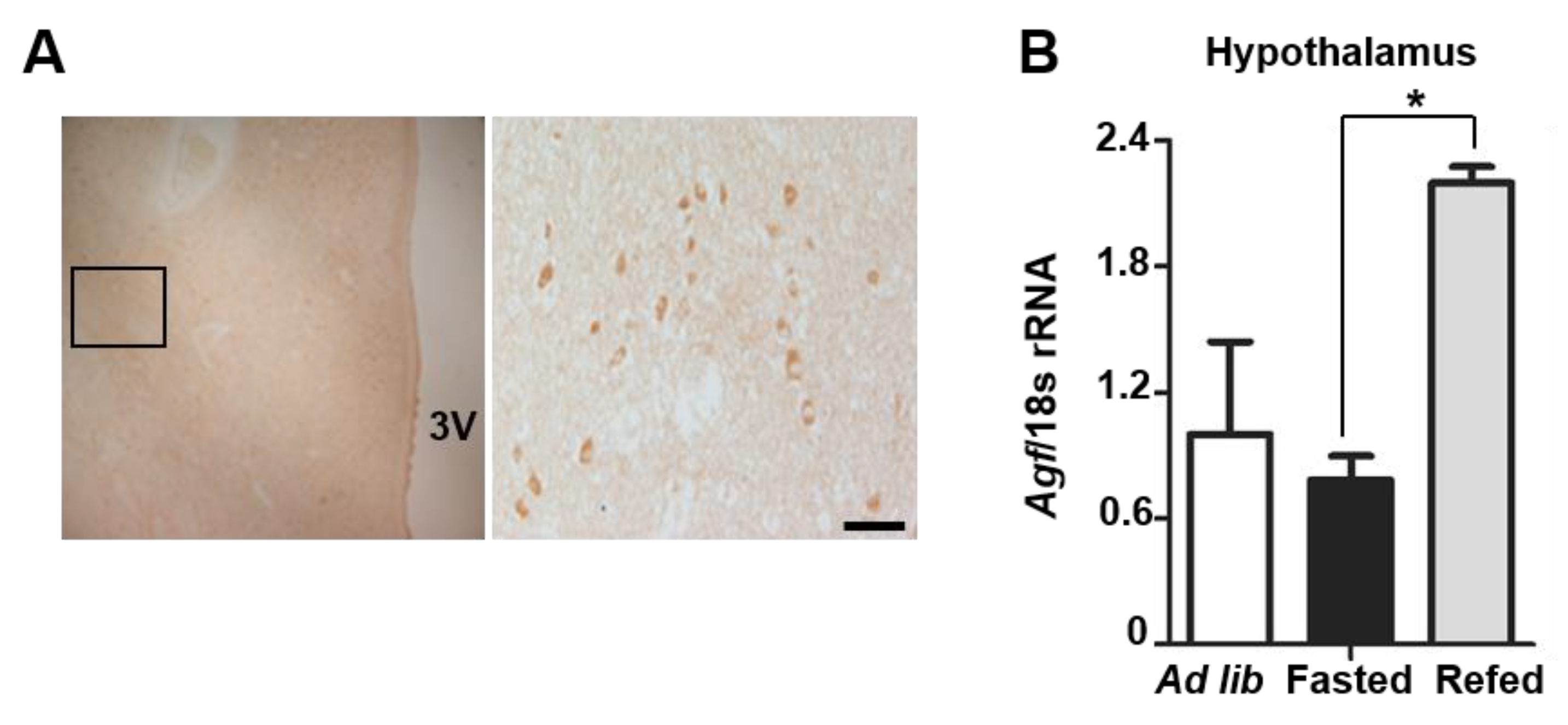
2.1. Angptl6 Is Expressed in the Hypothalamus and Induced after Feeding
2.2. Angptl6 Is Expressed in POMC Neurons of the Hypothalamus
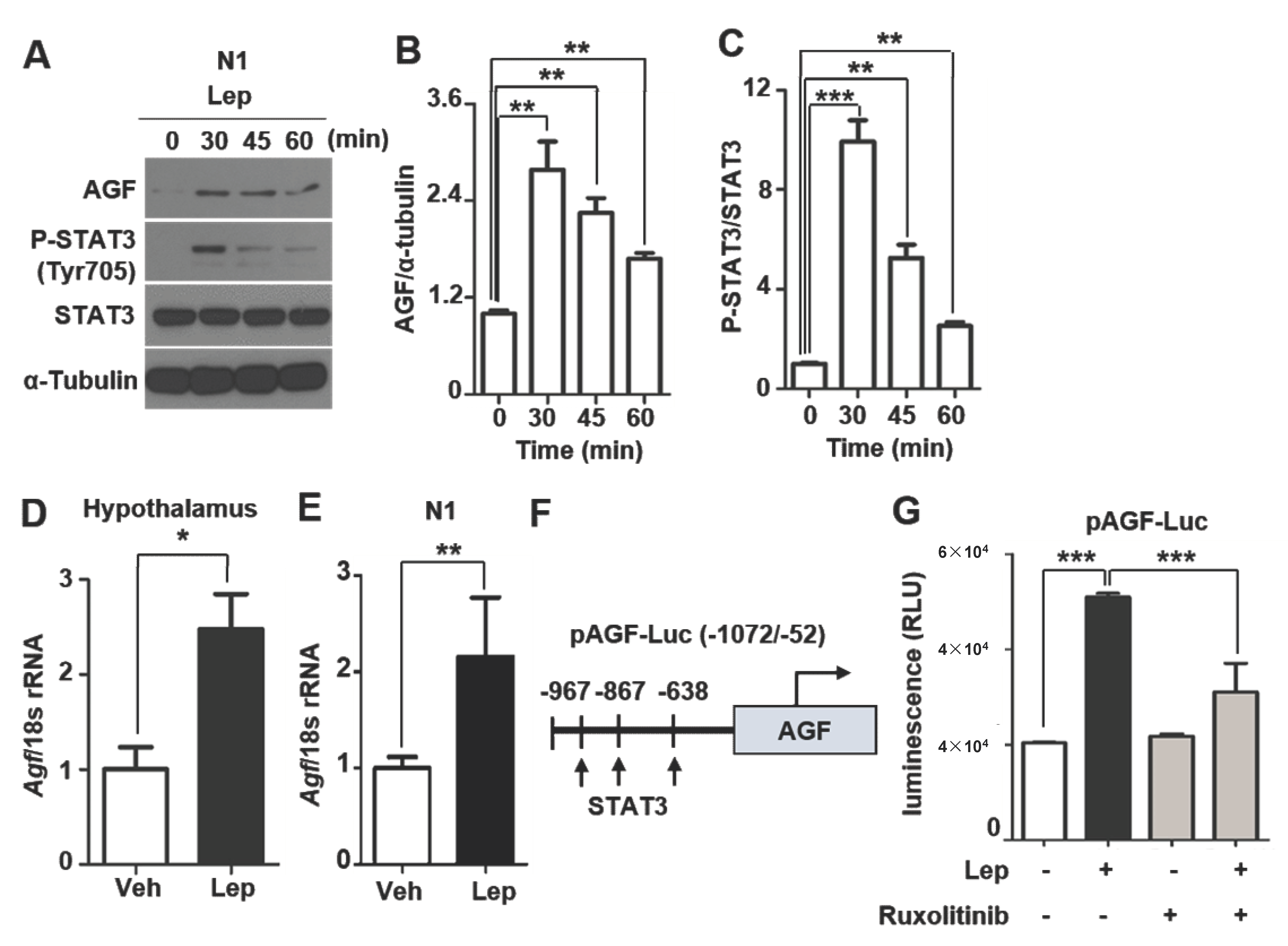
2.3. AGF Expression Is Associated with Leptin-Induced STAT3 Phosphorylation
2.4. AGF Promoter Activity Is Enhanced by Leptin
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Human Brain Tissue
4.4. Immunofluorescence Staining and Immunohistochemistry
4.5. RNA Iolation and Real-Time PCR
4.6. Western Blotting
4.7. Promoter-Luciferase Reporter Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suzuki, K.; Jayasena, C.N.; Bloom, S.R. Obesity and appetite control. Exp. Diabetes Res. 2012, 2012, 824305. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.A.; Bloom, S.R. Hypothalamic neuropeptides and the regulation of appetite. Neuropharmacology 2012, 63, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Barateiro, A.; Mahu, I.; Domingos, A.I. Leptin Resistance and the Neuro-Adipose Connection. Front. Endocrinol. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, Obesity, and Leptin Resistance: Where Are We 25 Years Later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef] [Green Version]
- DePaoli, A.M. 20 years of leptin: Leptin in common obesity and associated disorders of metabolism. J. Endocrinol. 2014, 223, T71–T81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenhill, C. Celastrol identified as a leptin sensitizer and potential novel treatment for obesity. Nat. Rev. Endocrinol. 2015, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Rahmouni, K.; Sigmund, C.D.; Haynes, W.G.; Mark, A.L. Hypothalamic ERK mediates the anorectic and thermogenic sympathetic effects of leptin. Diabetes 2009, 58, 536–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, O.; Kim, K.W.; Kim, M.S. Leptin signalling pathways in hypothalamic neurons. Cell Mol. Life Sci. 2016, 73, 1457–1477. [Google Scholar] [CrossRef] [PubMed]
- Balthasar, N.; Coppari, R.; McMinn, J.; Liu, S.M.; Lee, C.E.; Tang, V.; Kenny, C.D.; McGovern, R.A.; Chua, S.C., Jr.; Elmquist, J.K.; et al. Leptin receptor signaling in POMC neurons is required for normal body weight homeostasis. Neuron 2004, 42, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Wahed, A.; Gautier-Stein, A.; Casteras, S.; Soty, M.; Roussel, D.; Romestaing, C.; Guillou, H.; Tourette, J.A.; Pleche, N.; Zitoun, C.; et al. A link between hepatic glucose production and peripheral energy metabolism via hepatokines. Mol. Metab. 2014, 3, 531–543. [Google Scholar] [CrossRef]
- Oike, Y.; Tabata, M. Angiopoietin-like proteins--potential therapeutic targets for metabolic syndrome and cardiovascular disease. Circ. J. 2009, 73, 2192–2197. [Google Scholar] [CrossRef] [Green Version]
- Oike, Y.; Akao, M.; Yasunaga, K.; Yamauchi, T.; Morisada, T.; Ito, Y.; Urano, T.; Kimura, Y.; Kubota, Y.; Maekawa, H.; et al. Angiopoietin-related growth factor antagonizes obesity and insulin resistance. Nat. Med. 2005, 11, 400–408. [Google Scholar] [CrossRef]
- Kadomatsu, T.; Tabata, M.; Oike, Y. Angiopoietin-like proteins: Emerging targets for treatment of obesity and related metabolic diseases. FEBS J. 2011, 278, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Santulli, G. Angiopoietin-like proteins: A comprehensive look. Front. Endocrinol. 2014, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Shin, M.S.; Youn, B.S.; Kang, G.M.; Gil, S.Y.; Lee, C.H.; Choi, J.H.; Lim, H.S.; Yoo, H.J.; Kim, M.S. Regulation of energy balance by the hypothalamic lipoprotein lipase regulator Angptl3. Diabetes 2015, 64, 1142–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minokoshi, Y.; Alquier, T.; Furukawa, N.; Kim, Y.B.; Lee, A.; Xue, B.; Mu, J.; Foufelle, F.; Ferre, P.; Birnbaum, M.J.; et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature 2004, 428, 569–574. [Google Scholar] [CrossRef]
- Karatsoreos, I.N.; Thaler, J.P.; Borgland, S.L.; Champagne, F.A.; Hurd, Y.L.; Hill, M.N. Food for thought: Hormonal, experiential, and neural influences on feeding and obesity. J. Neurosci. 2013, 33, 17610–17616. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdan, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Varela, L.; Horvath, T.L. Leptin and insulin pathways in POMC and AgRP neurons that modulate energy balance and glucose homeostasis. EMBO Rep. 2012, 13, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Chhabra, K.H.; Thompson, Z.; Jones, G.L.; Kiran, S.; Shangguan, G.; Low, M.J. Hypothalamic POMC deficiency increases circulating adiponectin despite obesity. Mol. Metab. 2020, 35, 100957. [Google Scholar] [CrossRef]
- Air, E.L.; Benoit, S.C.; Clegg, D.J.; Seeley, R.J.; Woods, S.C. Insulin and leptin combine additively to reduce food intake and body weight in rats. Endocrinology 2002, 143, 2449–2452. [Google Scholar] [CrossRef]
- Baskin, D.G.; Figlewicz Lattemann, D.; Seeley, R.J.; Woods, S.C.; Porte, D., Jr.; Schwartz, M.W. Insulin and leptin: Dual adiposity signals to the brain for the regulation of food intake and body weight. Brain Res. 1999, 848, 114–123. [Google Scholar] [CrossRef]
- Shimizu, H.; Inoue, K.; Mori, M. The leptin-dependent and -independent melanocortin signaling system: Regulation of feeding and energy expenditure. J. Endocrinol. 2007, 193, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, C.; Pocai, A.; Muse, E.D.; Etgen, A.M.; Myers, M.G., Jr.; Rossetti, L. Critical role of STAT3 in leptin’s metabolic actions. Cell Metab. 2006, 4, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Namkung, J.; Chang, J.S.; Kim, S.J.; Park, K.S.; Kong, I.D. Leptin regulates the expression of angiopoietin-like 6. Biochem. Biophys. Res. Commun. 2018, 502, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.; Claret, M. Hypothalamic ER stress: A bridge between leptin resistance and obesity. FEBS Lett. 2015, 589, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Sainz, N.; Barrenetxe, J.; Moreno-Aliaga, M.J.; Martinez, J.A. Leptin resistance and diet-induced obesity: Central and peripheral actions of leptin. Metabolism 2015, 64, 35–46. [Google Scholar] [CrossRef]
- Scarpace, P.J.; Matheny, M.; Tumer, N. Hypothalamic leptin resistance is associated with impaired leptin signal transduction in aged obese rats. Neuroscience 2001, 104, 1111–1117. [Google Scholar] [CrossRef]
- Darambazar, G.; Nakata, M.; Okada, T.; Wang, L.; Li, E.; Shinozaki, A.; Motoshima, M.; Mori, M.; Yada, T. Paraventricular NUCB2/nesfatin-1 is directly targeted by leptin and mediates its anorexigenic effect. Biochem. Biophys. Res. Commun. 2015, 456, 913–918. [Google Scholar] [CrossRef]
- Perello, M.; Stuart, R.C.; Nillni, E.A. The role of intracerebroventricular administration of leptin in the stimulation of prothyrotropin releasing hormone neurons in the hypothalamic paraventricular nucleus. Endocrinology 2006, 147, 3296–3306. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Cha, D.; Kim, D.W.; Hoang, T.V.; Blackshaw, S. Tanycyte-Independent Control of Hypothalamic Leptin Signaling. Front. Neurosci. 2019, 13, 240. [Google Scholar] [CrossRef] [PubMed]
- Dodd, G.T.; Decherf, S.; Loh, K.; Simonds, S.E.; Wiede, F.; Balland, E.; Merry, T.L.; Munzberg, H.; Zhang, Z.Y.; Kahn, B.B.; et al. Leptin and insulin act on POMC neurons to promote the browning of white fat. Cell 2015, 160, 88–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shintani, T.; Higashi, S.; Suzuki, R.; Takeuchi, Y.; Ikaga, R.; Yamazaki, T.; Kobayashi, K.; Noda, M. PTPRJ Inhibits Leptin Signaling, and Induction of PTPRJ in the Hypothalamus Is a Cause of the Development of Leptin Resistance. Sci. Rep. 2017, 7, 11627. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, Y.; Heo, J.Y.; Lee, M.J.; Zhu, J.; Seo, C.; Go, D.H.; Yoon, S.K.; Yukari, D.; Oike, Y.; Sohn, J.-W.; et al. Angiopoietin-Like Growth Factor Involved in Leptin Signaling in the Hypothalamus. Int. J. Mol. Sci. 2021, 22, 3443. https://doi.org/10.3390/ijms22073443
Jang Y, Heo JY, Lee MJ, Zhu J, Seo C, Go DH, Yoon SK, Yukari D, Oike Y, Sohn J-W, et al. Angiopoietin-Like Growth Factor Involved in Leptin Signaling in the Hypothalamus. International Journal of Molecular Sciences. 2021; 22(7):3443. https://doi.org/10.3390/ijms22073443
Chicago/Turabian StyleJang, Yunseon, Jun Young Heo, Min Joung Lee, Jiebo Zhu, Changjun Seo, Da Hyun Go, Sung Kyung Yoon, Date Yukari, Yuichi Oike, Jong-Woo Sohn, and et al. 2021. "Angiopoietin-Like Growth Factor Involved in Leptin Signaling in the Hypothalamus" International Journal of Molecular Sciences 22, no. 7: 3443. https://doi.org/10.3390/ijms22073443
APA StyleJang, Y., Heo, J. Y., Lee, M. J., Zhu, J., Seo, C., Go, D. H., Yoon, S. K., Yukari, D., Oike, Y., Sohn, J. -W., Shong, M., & Kweon, G. R. (2021). Angiopoietin-Like Growth Factor Involved in Leptin Signaling in the Hypothalamus. International Journal of Molecular Sciences, 22(7), 3443. https://doi.org/10.3390/ijms22073443