
The Nucleolar Protein Nucleophosmin Is Physiologically Secreted by Endothelial Cells in Response to Stress Exerting Proangiogenic Activity Both In Vitro and In Vivo

 , , , and
, , , and
Abstract
:1. Introduction
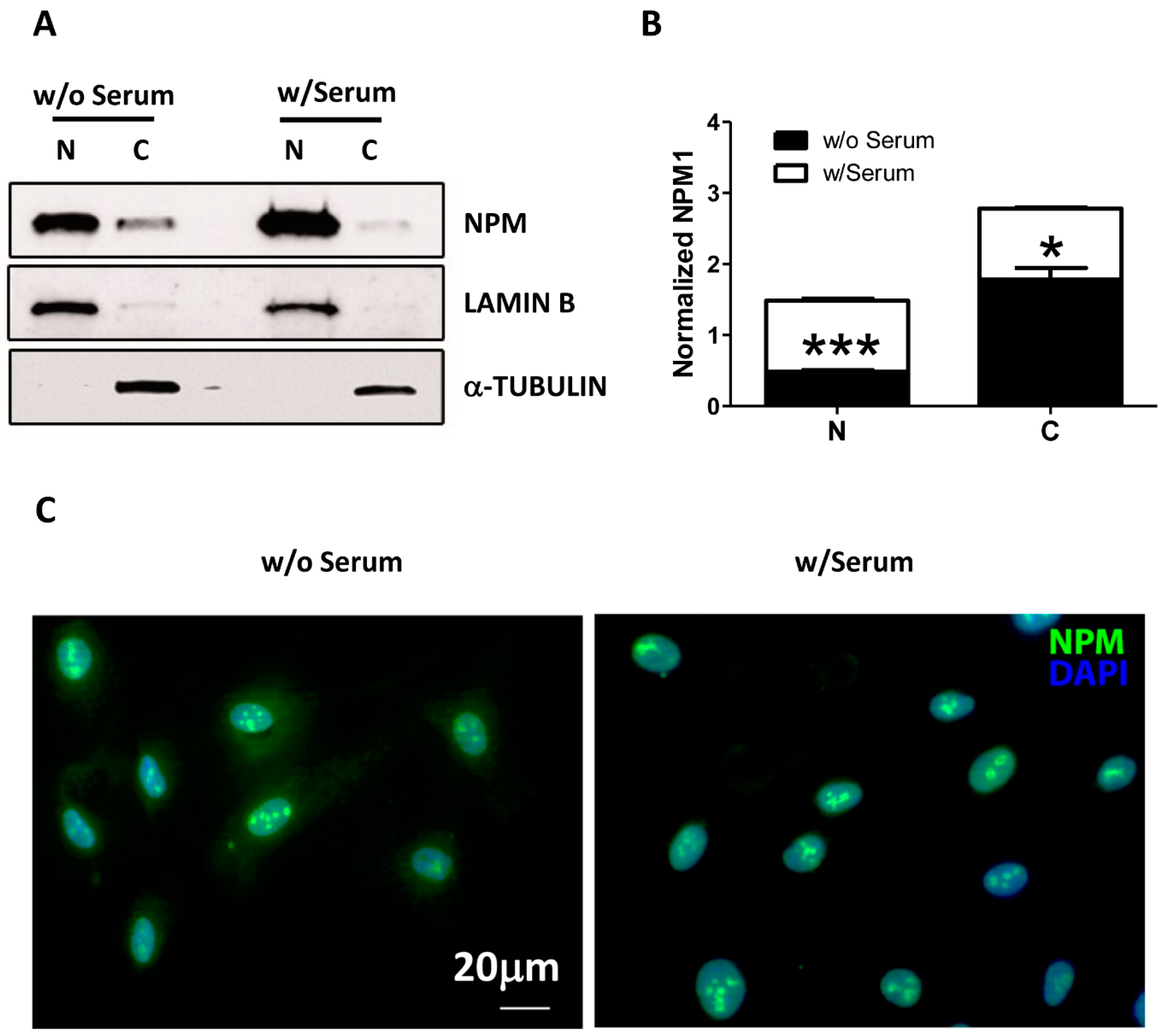
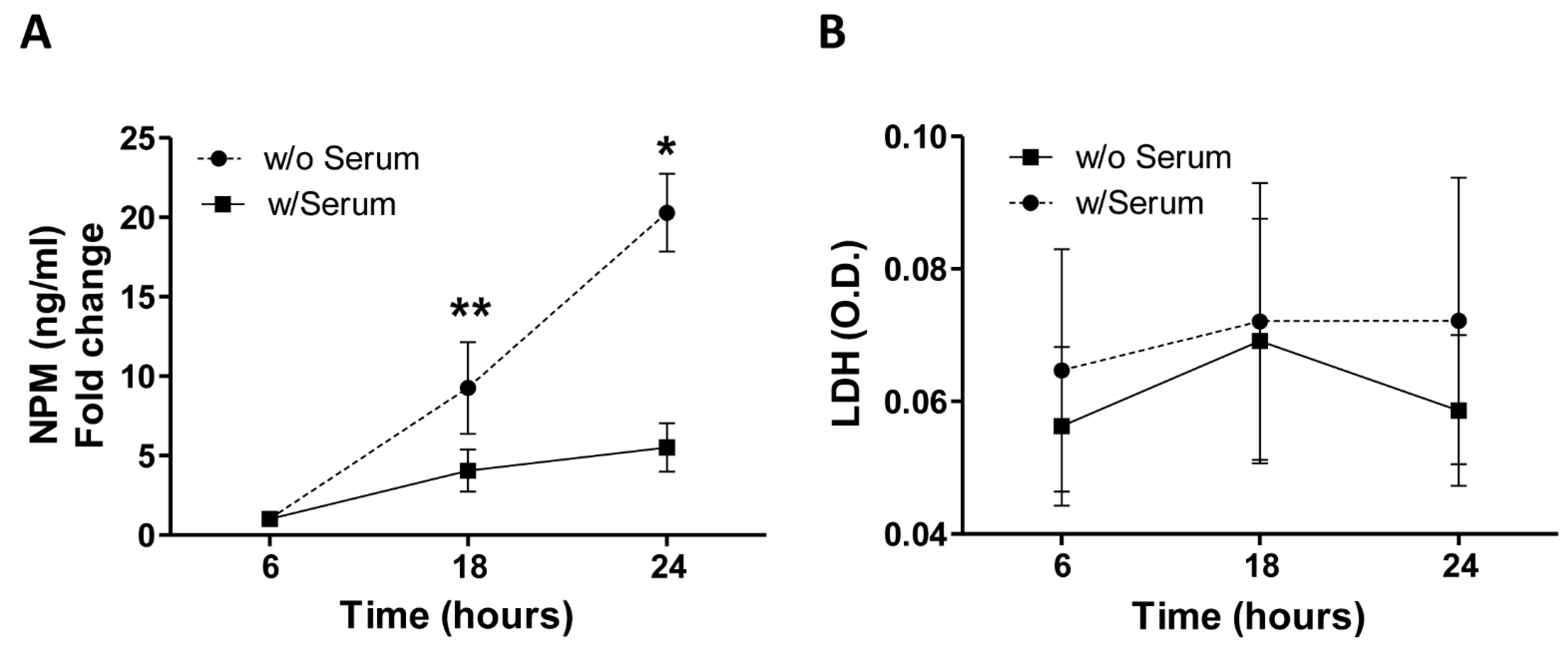
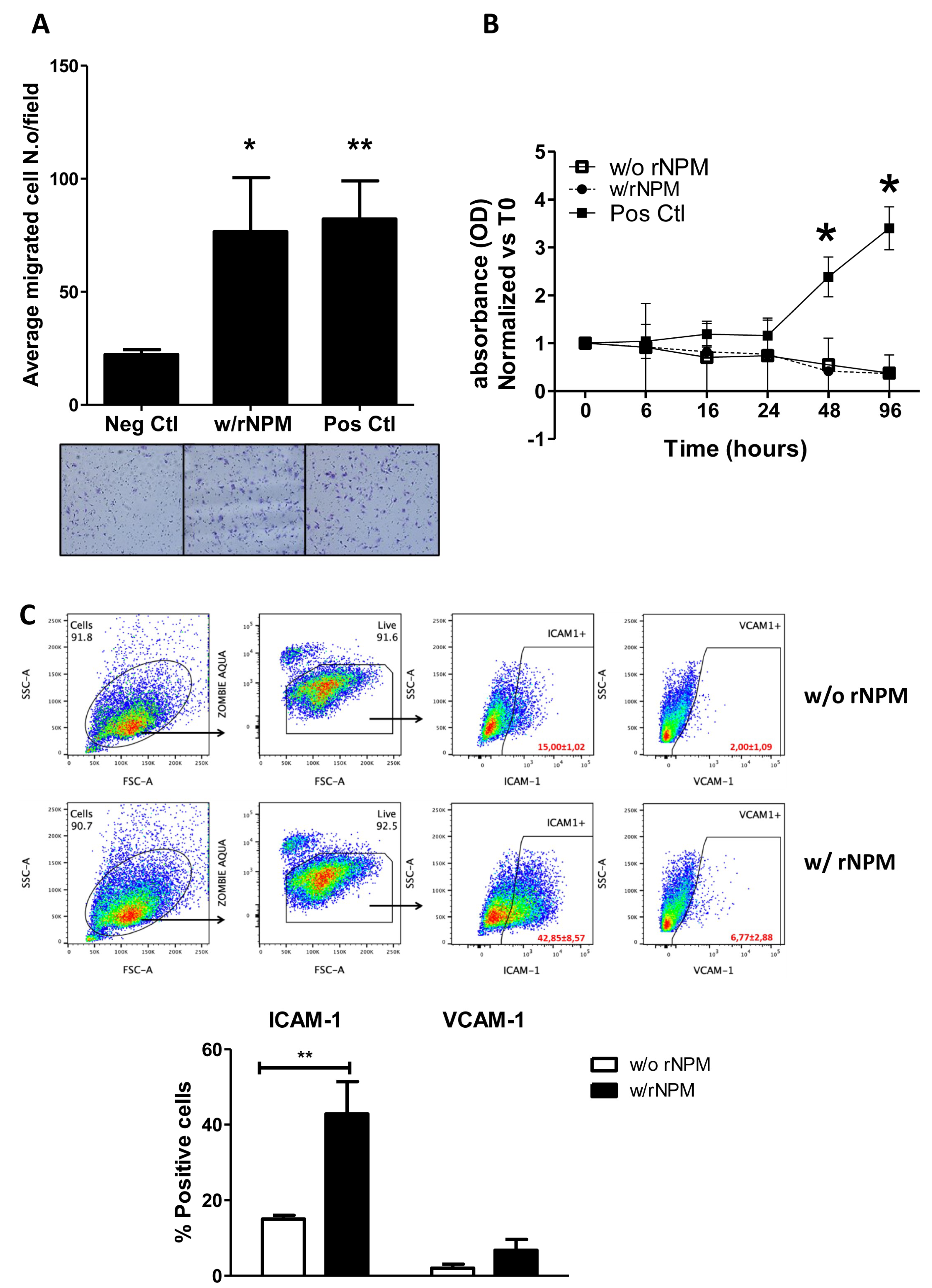
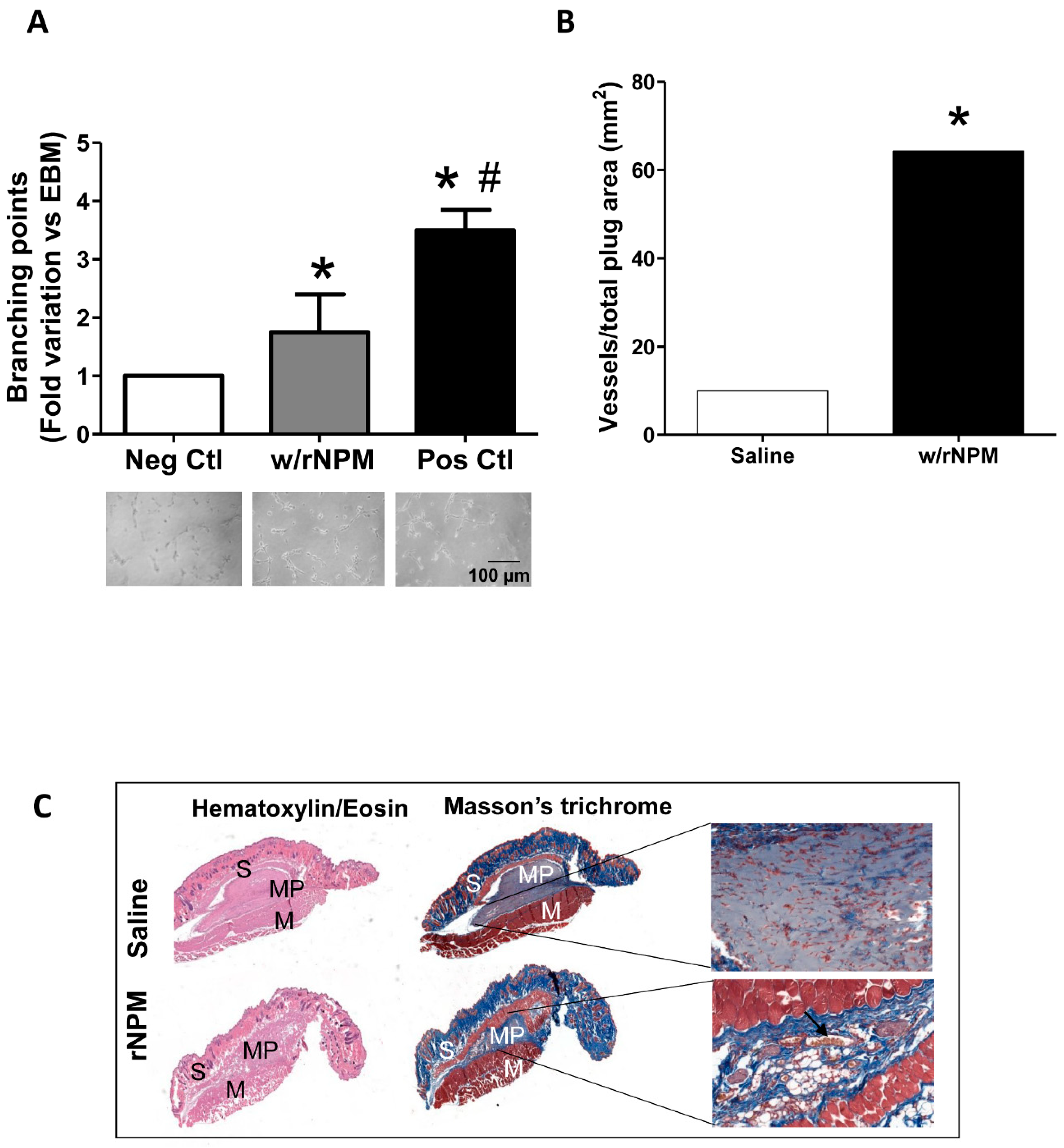
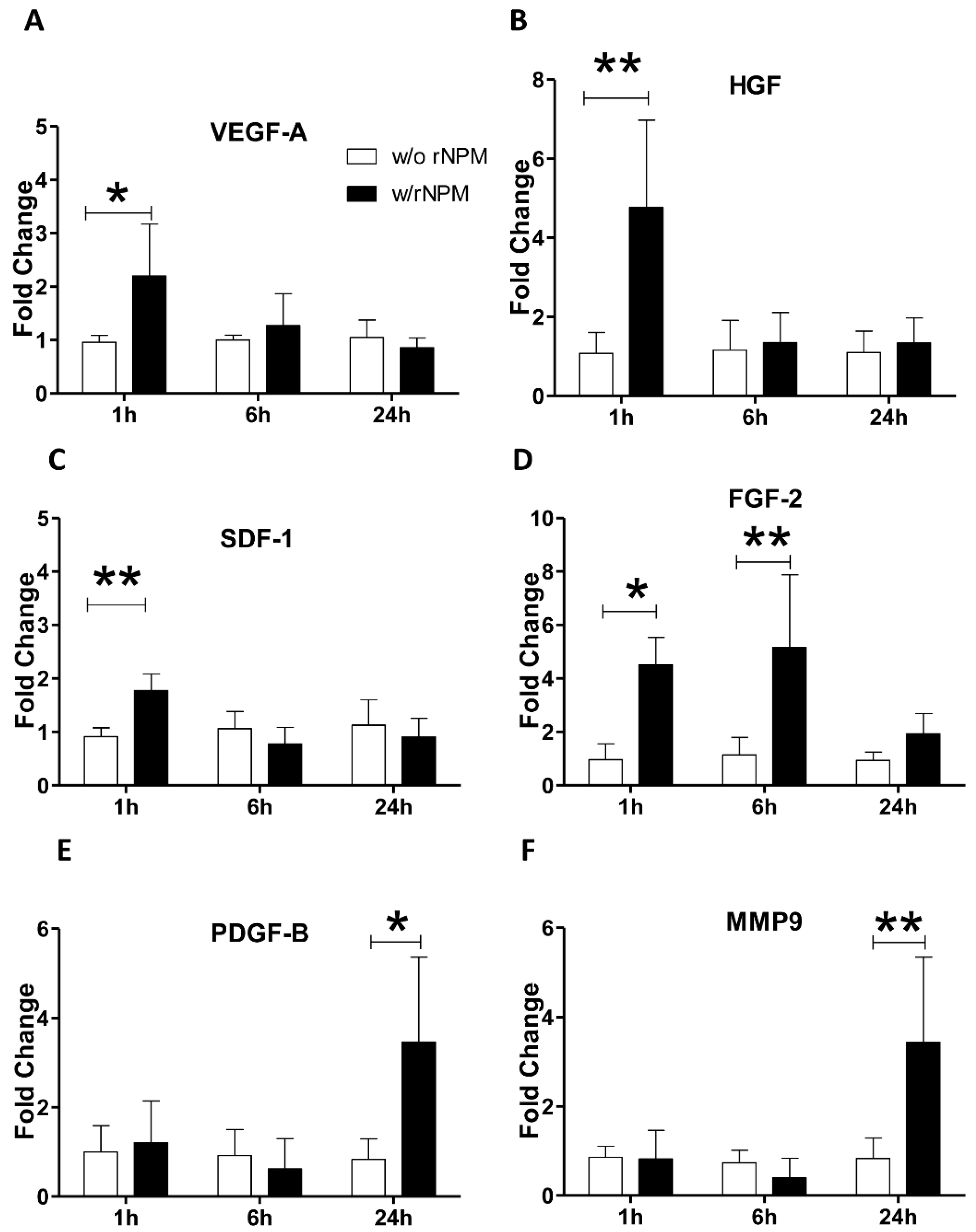
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Stimulation
4.2. Western Blot
4.3. Immunofluorescence
4.4. ELISA and LDH Assay
4.5. Proliferation and Migration
4.6. Angiogenesis Assay
4.7. FACS Analysis
4.8. Real-Time PCR
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a clinical response. J. Clin. Investig. 2012, 122, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.A.; Pober, J.S. Endothelial injury, alarmins, and allograft rejection. Crit. Rev. Immunol. 2008, 28, 229–248. [Google Scholar] [CrossRef]
- Rider, P.; Voronov, E.; Dinarello, C.A.; Apte, R.N.; Cohen, I. Alarmins: Feel the Stress. J. Immunol. 2017, 198, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denk, S.; Perl, M.; Huber-Lang, M. Damage- and pathogen-associated molecular patterns and alarmins: Keys to sepsis? Eur. Surg. Res. 2012, 48, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Box, J.K.; Paquet, N.; Adams, M.N.; Boucher, D.; Bolderson, E.; O’Byrne, K.J.; Richard, D.J. Nucleophosmin: From structure and function to disease development. BMC Mol. Biol. 2016, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siying, C.; Meng, T.; Zheng, X.; Cai, J.; Zhang, W.; You, H.; Xing, J.; Dong, Y. Contribution of nucleophosmin overexpression to multidrug resistance in breast carcinoma. J. Drug Target. 2018, 26, 27–35. [Google Scholar]
- Kim, B.G.; Kwon, H.Y.; Sohn, E.J.; Hwang, S.; Kwon, O.S.; Kim, S.H. Activation of caspases and inhibition of ribosome biogenesis mediate antitumor activity of Chijongdan in A549 non-small lung cancer cells. BMC Complement. Altern. Med. 2014, 14, 420. [Google Scholar] [CrossRef] [Green Version]
- Leotoing, L.; Meunier, L.; Manin, M.; Mauduit, C.; Decaussin, M.; Verrijdt, G.; Claessens, F.; Benahmed, M.; Veyssiere, G.; Morel, L.; et al. Influence of nucleophosmin/B23 on DNA binding and transcriptional activity of the androgen receptor in prostate cancer cell. Oncogene 2008, 27, 2858–2867. [Google Scholar] [CrossRef] [Green Version]
- Holmberg Olausson, K.; Elsir, T.; Moazemi Goudarzi, K.; Nister, M.; Lindstrom, M.S. NPM1 histone chaperone is upregulated in glioblastoma to promote cell survival and maintain nucleolar shape. Sci. Rep. 2015, 5, 16495. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Shen, J.; Xia, L.; Wang, Y. Estrogen mediated expression of nucleophosmin 1 in human endometrial carcinoma clinical stages through estrogen receptor-alpha signaling. Cancer Cell Int. 2014, 14, 540. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.C.; Hasan, M.R.; Rahman, M.; Yu, A.C.; Chan, S.K.; Schaeffer, D.F.; Kennecke, H.F.; Lim, H.J.; Owen, D.; Tai, I.T. Nucleophosmin 1, upregulated in adenomas and cancers of the colon, inhibits p53-mediated cellular senescence. Int. J. Cancer 2013, 133, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Kinumi, T.; Ogawa, Y.; Kimata, J.; Saito, Y.; Yoshida, Y.; Niki, E. Proteomic characterization of oxidative dysfunction in human umbilical vein endothelial cells (HUVEC) induced by exposure to oxidized LDL. Free Radic. Res. 2005, 39, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Csiszar, A.; Wang, M.; Lakatta, E.G.; Ungvari, Z. Inflammation and endothelial dysfunction during aging: Role of NF-kappaB. J. Appl. Physiol. (1985) 2008, 105, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Avitabile, D.; Bailey, B.; Cottage, C.T.; Sundararaman, B.; Joyo, A.; McGregor, M.; Gude, N.; Truffa, S.; Zarrabi, A.; Konstandin, M.; et al. Nucleolar stress is an early response to myocardial damage involving nucleolar proteins nucleostemin and nucleophosmin. Proc. Natl. Acad. Sci. USA 2011, 108, 6145–6150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, C.; Liu, B.; Huang, D.; Chen, R.; Huang, K.; Li, F.; Dong, N. Nucleophosmin contributes to vascular inflammation and endothelial dysfunction in atherosclerosis progression. J. Thorac. Cardiovasc. Surg. 2019. [Google Scholar] [CrossRef] [PubMed]
- Nawa, Y.; Kawahara, K.; Tancharoen, S.; Meng, X.; Sameshima, H.; Ito, T.; Masuda, Y.; Imaizumi, H.; Hashiguchi, T.; Maruyama, I. Nucleophosmin may act as an alarmin: Implications for severe sepsis. J. Leukoc. Biol. 2009, 86, 645–653. [Google Scholar] [CrossRef]
- Yang, S.; Xu, L.; Yang, T.; Wang, F. High-mobility group box-1 and its role in angiogenesis. J. Leukoc. Biol. 2014, 95, 563–574. [Google Scholar] [CrossRef]
- Levin, V.A.; Panchabhai, S.C.; Shen, L.; Kornblau, S.M.; Qiu, Y.; Baggerly, K.A. Different changes in protein and phosphoprotein levels result from serum starvation of high-grade glioma and adenocarcinoma cell lines. J. Proteome Res. 2010, 9, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Pirkmajer, S.; Chibalin, A.V. Serum starvation: Caveat emptor. Am. J. Physiol. Cell Physiol. 2011, 301, C272–C279. [Google Scholar] [CrossRef] [Green Version]
- Meder, V.S.; Boeglin, M.; de Murcia, G.; Schreiber, V. PARP-1 and PARP-2 interact with nucleophosmin/B23 and accumulate in transcriptionally active nucleoli. J. Cell Sci. 2005, 118, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, X.; Sejas, D.P.; Bagby, G.C.; Pang, Q. Hypoxia-induced nucleophosmin protects cell death through inhibition of p53. J. Biol. Chem. 2004, 279, 41275–41279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, N.T.; Martin, M.U. Interleukin 33 is a guardian of barriers and a local alarmin. Nat. Immunol. 2016, 17, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, F.; Altamura, S.; Frosali, S.; Conti, P. Key Role of DAMP in Inflammation, Cancer, and Tissue Repair. Clin. Ther. 2016, 38, 1017–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limana, F.; Germani, A.; Zacheo, A.; Kajstura, J.; Di Carlo, A.; Borsellino, G.; Leoni, O.; Palumbo, R.; Battistini, L.; Rastaldo, R.; et al. Exogenous high-mobility group box 1 protein induces myocardial regeneration after infarction via enhanced cardiac C-kit+ cell proliferation and differentiation. Circ. Res. 2005, 97, e73–e83. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S. Reappraisal of serum starvation, the restriction point, G0, and G1 phase arrest points. FASEB J. 2003, 17, 333–340. [Google Scholar] [CrossRef]
- Chen, X.L.; Xia, Z.F.; Wei, D.; Liao, H.G.; Ben, D.F.; Wang, G.Q. Expression and regulation of vascular cell adhesion molecule-1 in human umbilical vein endothelial cells induced by sera from severely burned patients. Crit. Care Med. 2004, 32, 77–82. [Google Scholar] [CrossRef]
- Jersmann, H.P.; Hii, C.S.; Ferrante, J.V.; Ferrante, A. Bacterial lipopolysaccharide and tumor necrosis factor alpha synergistically increase expression of human endothelial adhesion molecules through activation of NF-kappaB and p38 mitogen-activated protein kinase signaling pathways. Infect. Immun. 2001, 69, 1273–1279. [Google Scholar] [CrossRef] [Green Version]
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef]
- Auerbach, R.; Lewis, R.; Shinners, B.; Kubai, L.; Akhtar, N. Angiogenesis assays: A critical overview. Clin. Chem. 2003, 49, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Stieger, S.M.; Bloch, S.H.; Foreman, O.; Wisner, E.R.; Ferrara, K.W.; Dayton, P.A. Ultrasound assessment of angiogenesis in a matrigel model in rats. Ultrasound Med. Biol. 2006, 32, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Farina, A.R.; Mackay, A.R. Gelatinase B/MMP-9 in Tumour Pathogenesis and Progression. Cancers 2014, 6, 240–296. [Google Scholar] [CrossRef]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef]
- Taylor, S.M.; Nevis, K.R.; Park, H.L.; Rogers, G.C.; Rogers, S.L.; Cook, J.G.; Bautch, V.L. Angiogenic factor signaling regulates centrosome duplication in endothelial cells of developing blood vessels. Blood 2010, 116, 3108–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawagishi, H.; Nakamura, H.; Maruyama, M.; Mizutani, S.; Sugimoto, K.; Takagi, M.; Sugimoto, M. ARF suppresses tumor angiogenesis through translational control of VEGFA mRNA. Cancer Res. 2010, 70, 4749–4758. [Google Scholar] [CrossRef] [Green Version]
- Onikubo, T.; Nicklay, J.J.; Xing, L.; Warren, C.; Anson, B.; Wang, W.L.; Burgos, E.S.; Ruff, S.E.; Shabanowitz, J.; Cheng, R.H.; et al. Developmentally Regulated Post-translational Modification of Nucleoplasmin Controls Histone Sequestration and Deposition. Cell Rep. 2015, 10, 1735–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s double life: Intracellular regulator and extracellular signal. Biochim. Biophys. Acta 2009, 1793, 1008–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, N.Q.K. Fertility-GRU: Identifying Fertility-Related Proteins by Incorporating Deep-Gated Recurrent Units and Original Position-Specific Scoring Matrix Profiles. J. Proteome Res. 2019, 18, 3503–3511. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence |
|---|---|
| 18S | Fwd: CATGGCCTCAGTTCCGAAAA Rev: CGAGCCGCCTGGATACC |
| VEGF-A | Fwd: CTACCTCCACCATGCCAAGT Rev: CACACAGGATGGCTTGAAGA |
| SDF-1 | Fwd: GCTGGTCCTCGTGCTGAC Rev: GCATGGGCATCTGTAGCTC |
| HGF | Fwd: GGACGCAGCTACAAGGGAAC Rev: CCTTCTTCCCCTCGAGGATT |
| FGF-2 | Fwd: GAGACACCCATCCGTGAACC Rev: GGCAGCGTGGTGATGCTC |
| PDGF-B | Fwd: CTCGTCCGTCTGTCTCGATG Rev: GGAAGAAGATGGCGATGGAG |
| MMP9 | Fwd: GAACCAATCTCACCGACAGG Rev: GCCACCCGAGTGTAACCATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Carlo, A.; Beji, S.; Palmerio, S.; Picozza, M.; D’Agostino, M.; Petrozza, V.; Melchionna, R.; Germani, A.; Magenta, A.; De Falco, E.; et al. The Nucleolar Protein Nucleophosmin Is Physiologically Secreted by Endothelial Cells in Response to Stress Exerting Proangiogenic Activity Both In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 3672. https://doi.org/10.3390/ijms22073672
Di Carlo A, Beji S, Palmerio S, Picozza M, D’Agostino M, Petrozza V, Melchionna R, Germani A, Magenta A, De Falco E, et al. The Nucleolar Protein Nucleophosmin Is Physiologically Secreted by Endothelial Cells in Response to Stress Exerting Proangiogenic Activity Both In Vitro and In Vivo. International Journal of Molecular Sciences. 2021; 22(7):3672. https://doi.org/10.3390/ijms22073672
Chicago/Turabian StyleDi Carlo, Anna, Sara Beji, Silvia Palmerio, Mario Picozza, Marco D’Agostino, Vincenzo Petrozza, Roberta Melchionna, Antonia Germani, Alessandra Magenta, Elena De Falco, and et al. 2021. "The Nucleolar Protein Nucleophosmin Is Physiologically Secreted by Endothelial Cells in Response to Stress Exerting Proangiogenic Activity Both In Vitro and In Vivo" International Journal of Molecular Sciences 22, no. 7: 3672. https://doi.org/10.3390/ijms22073672
APA StyleDi Carlo, A., Beji, S., Palmerio, S., Picozza, M., D’Agostino, M., Petrozza, V., Melchionna, R., Germani, A., Magenta, A., De Falco, E., & Avitabile, D. (2021). The Nucleolar Protein Nucleophosmin Is Physiologically Secreted by Endothelial Cells in Response to Stress Exerting Proangiogenic Activity Both In Vitro and In Vivo. International Journal of Molecular Sciences, 22(7), 3672. https://doi.org/10.3390/ijms22073672