
Structural Investigation and Molecular Modeling Studies of Strobilurin-Based Fungicides Active against the Rice Blast Pathogen Pyricularia oryzae

 , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
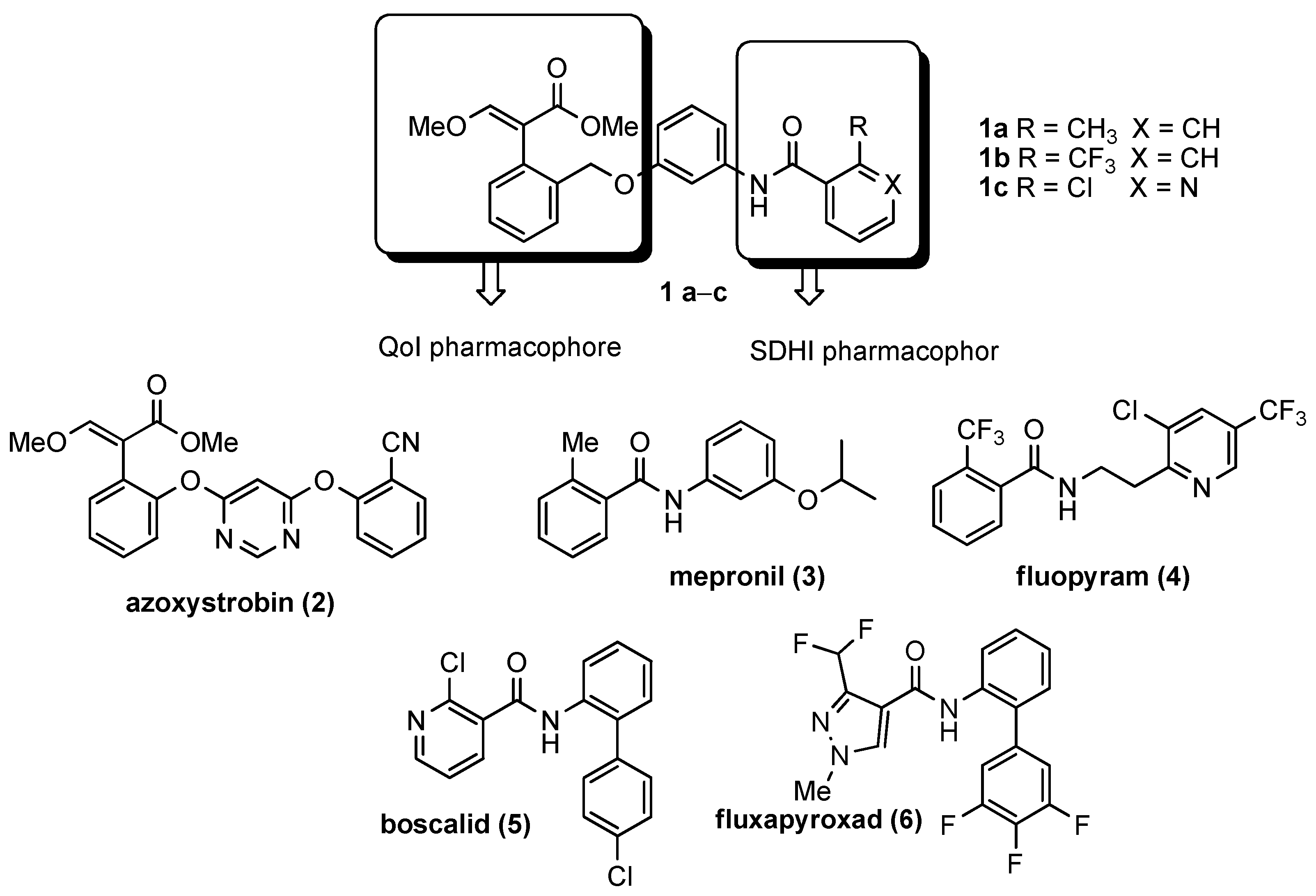
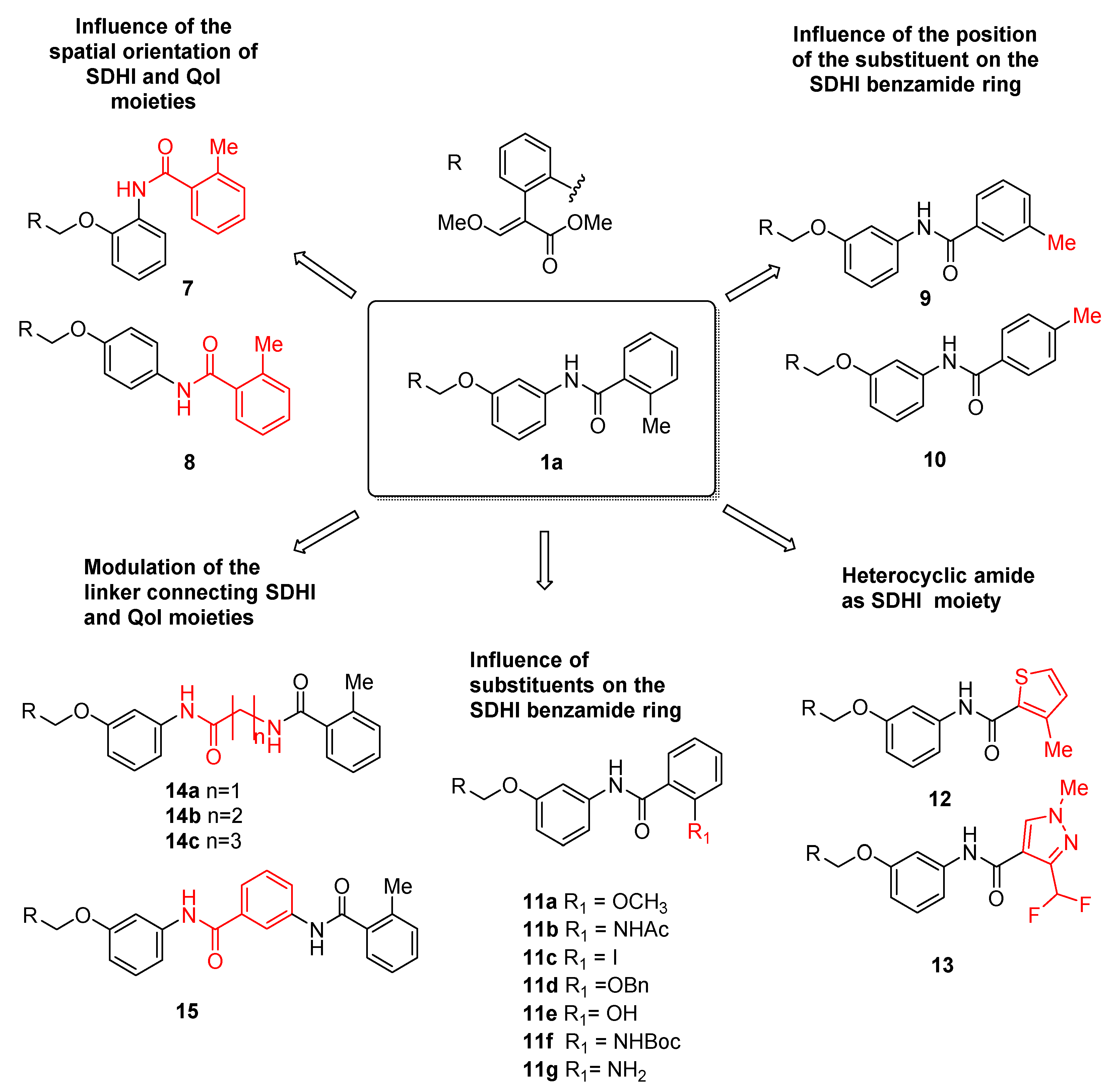
- Spatial orientation of the two pharmacophores
- Role and nature of the linker
- Nature and substitution pattern of the SDHI pharmacophore.
2. Results
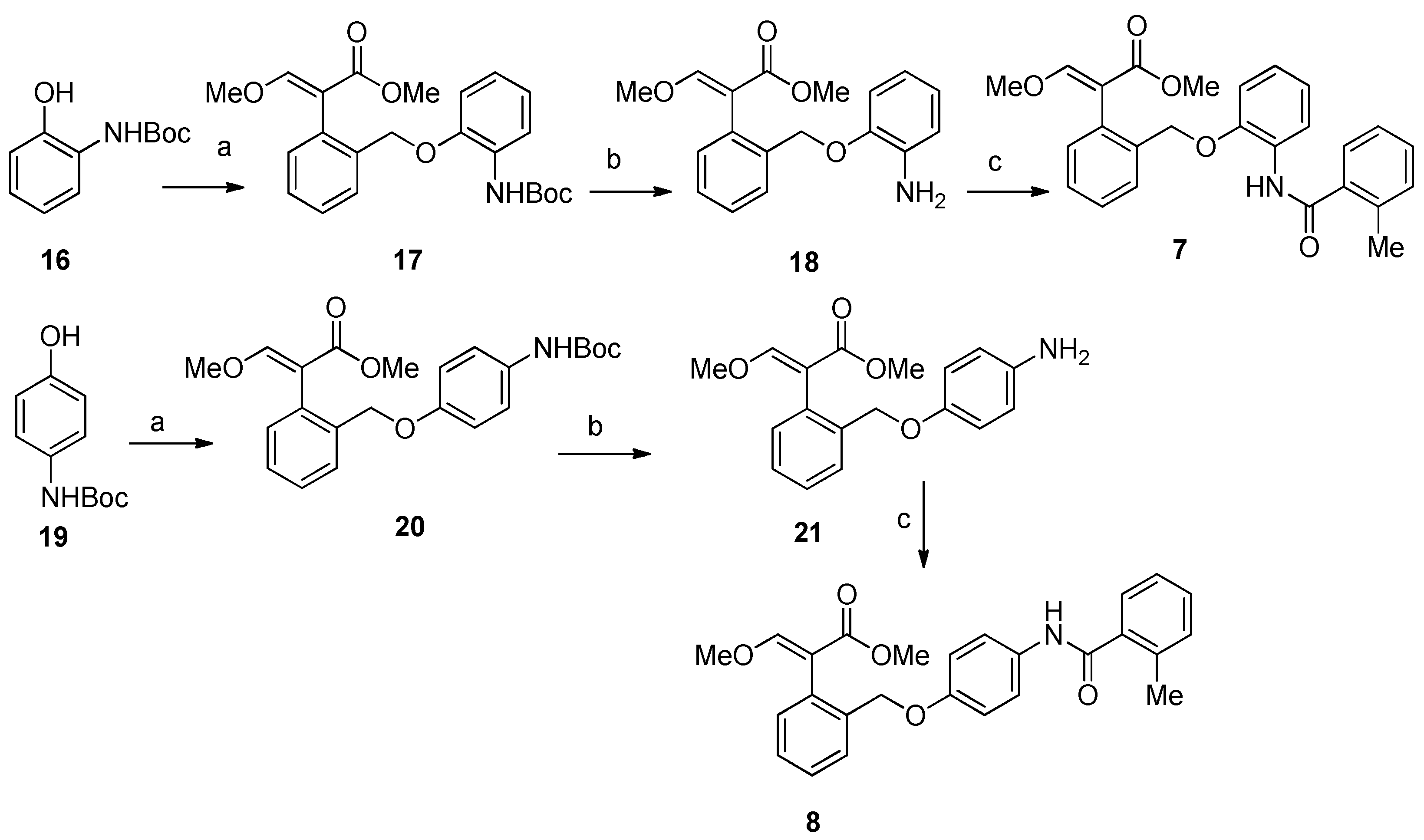
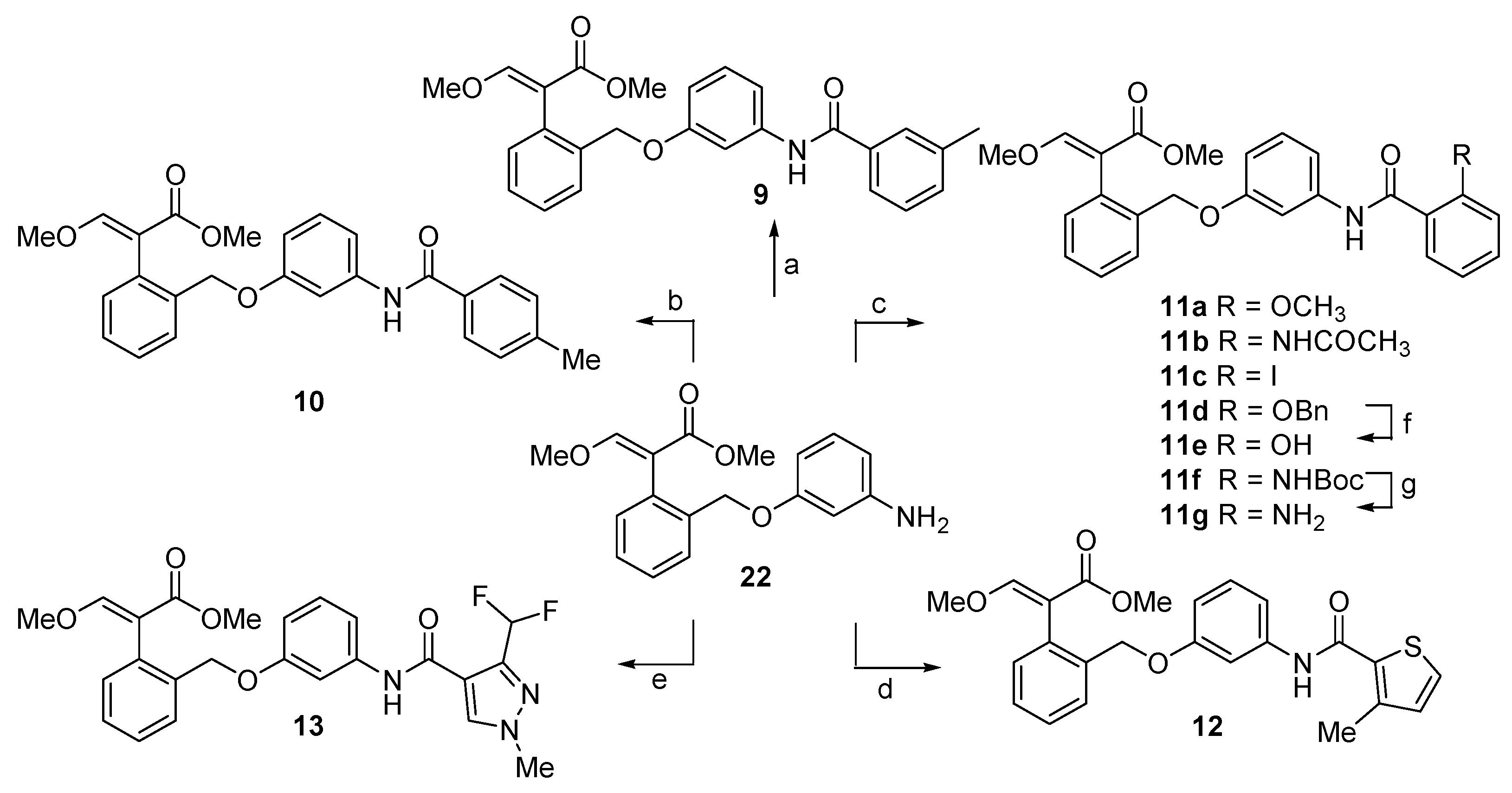
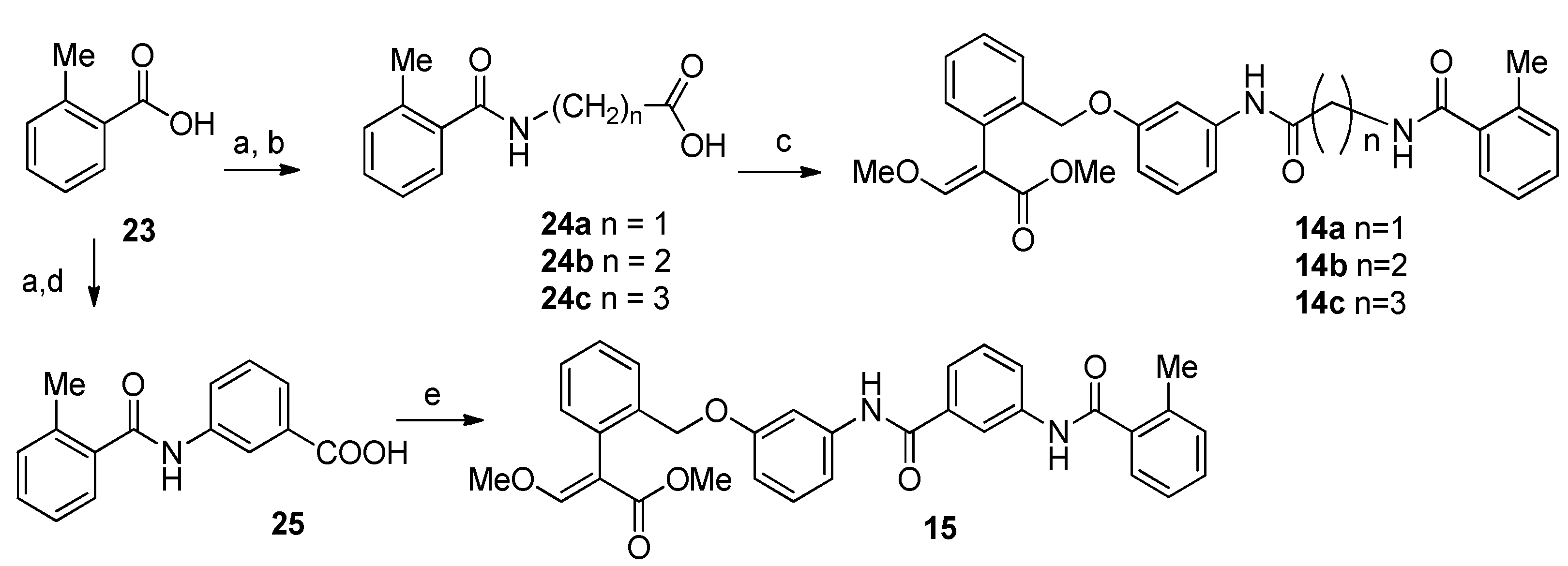
2.1. Chemistry
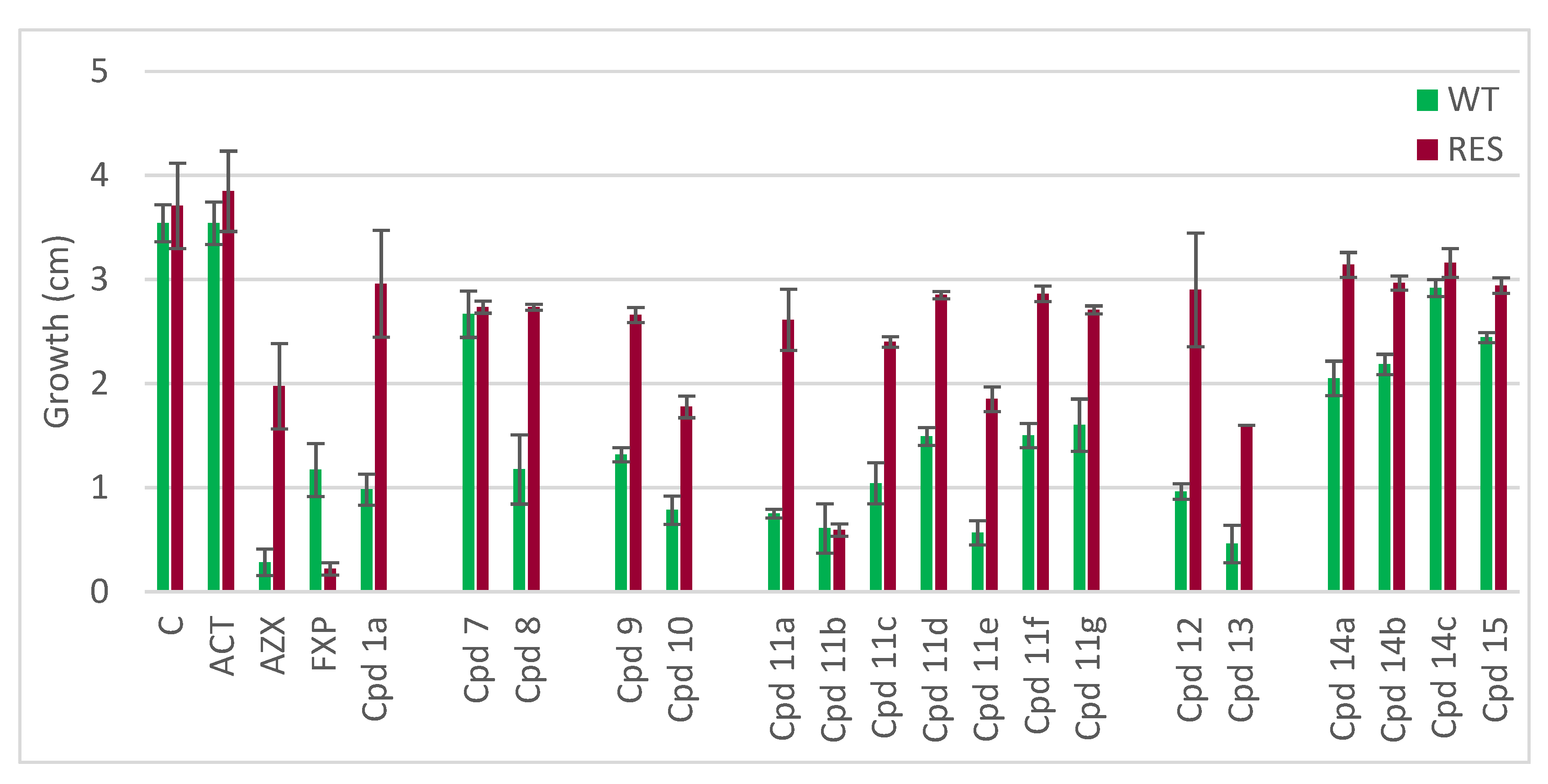
2.2. Biological Activity
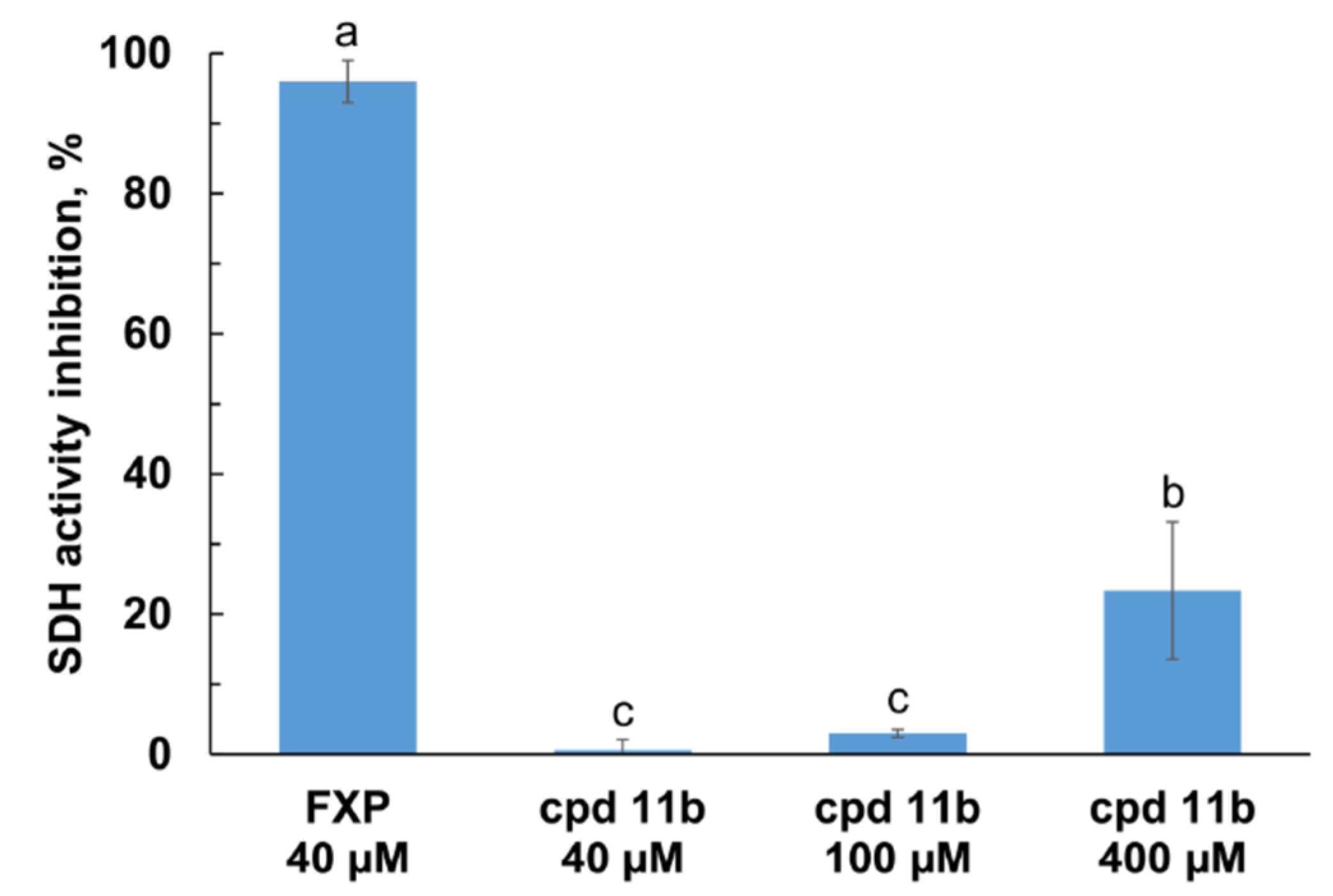
2.3. SDH and Qo Inhibitory Activity of Compound 11b
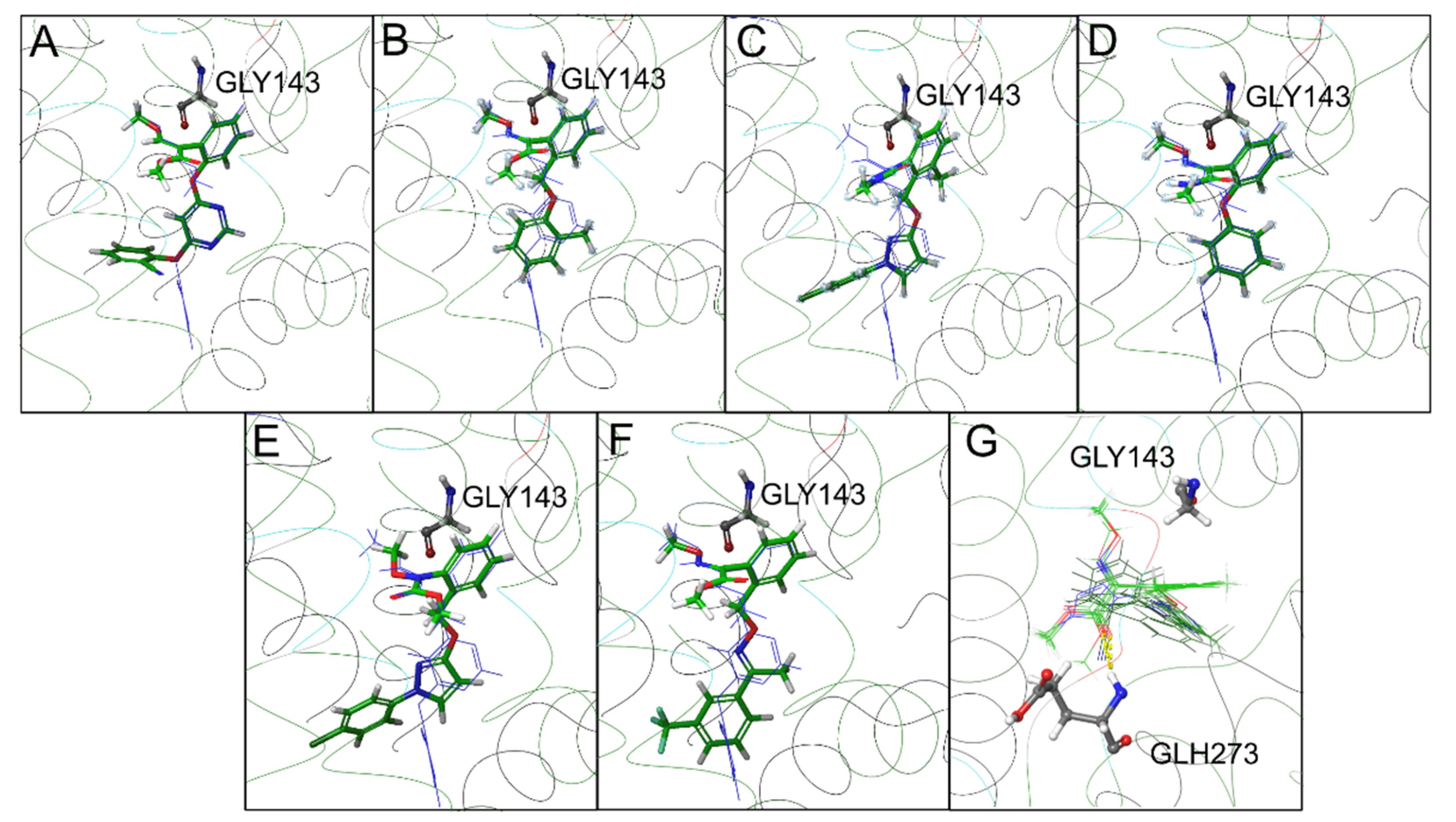
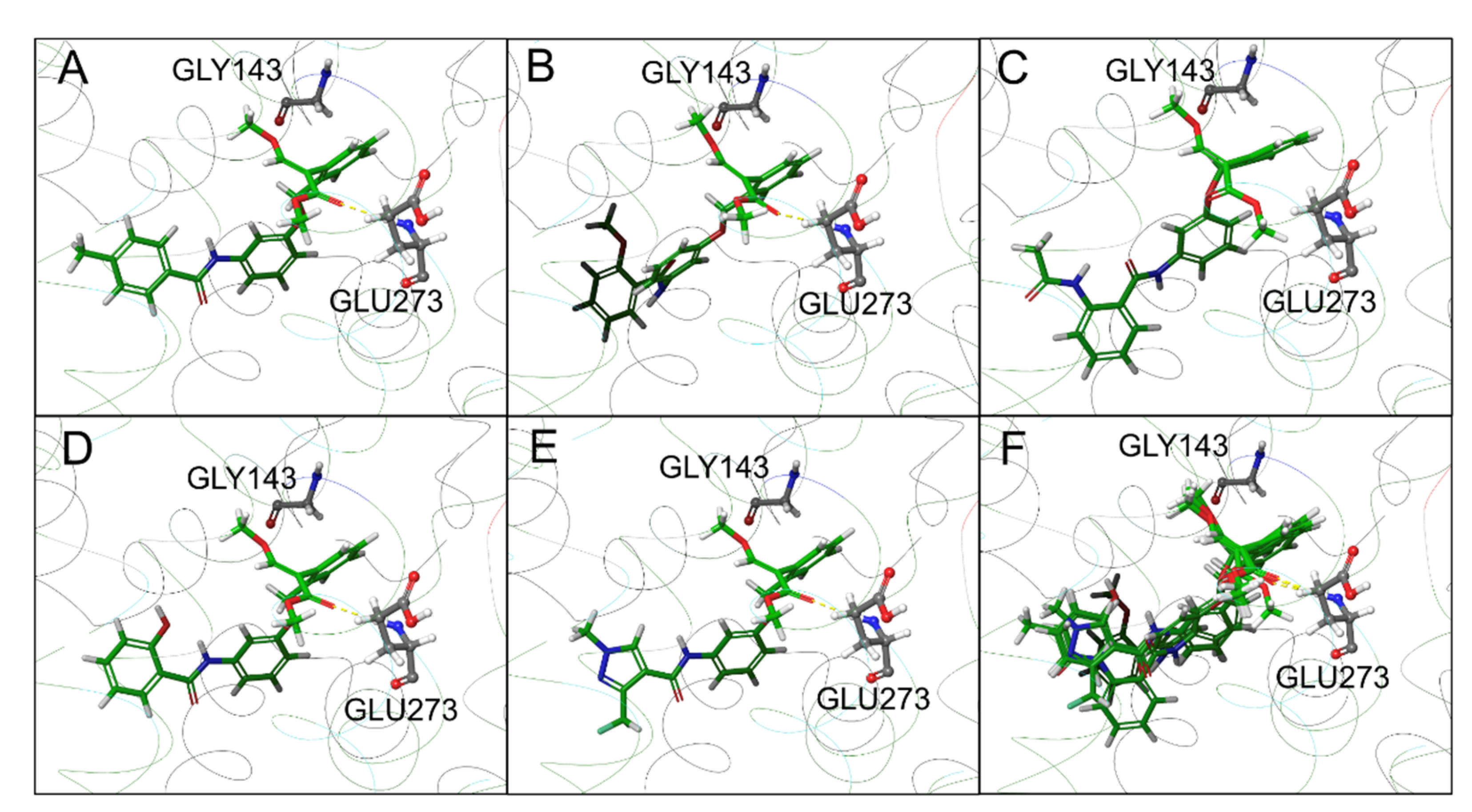
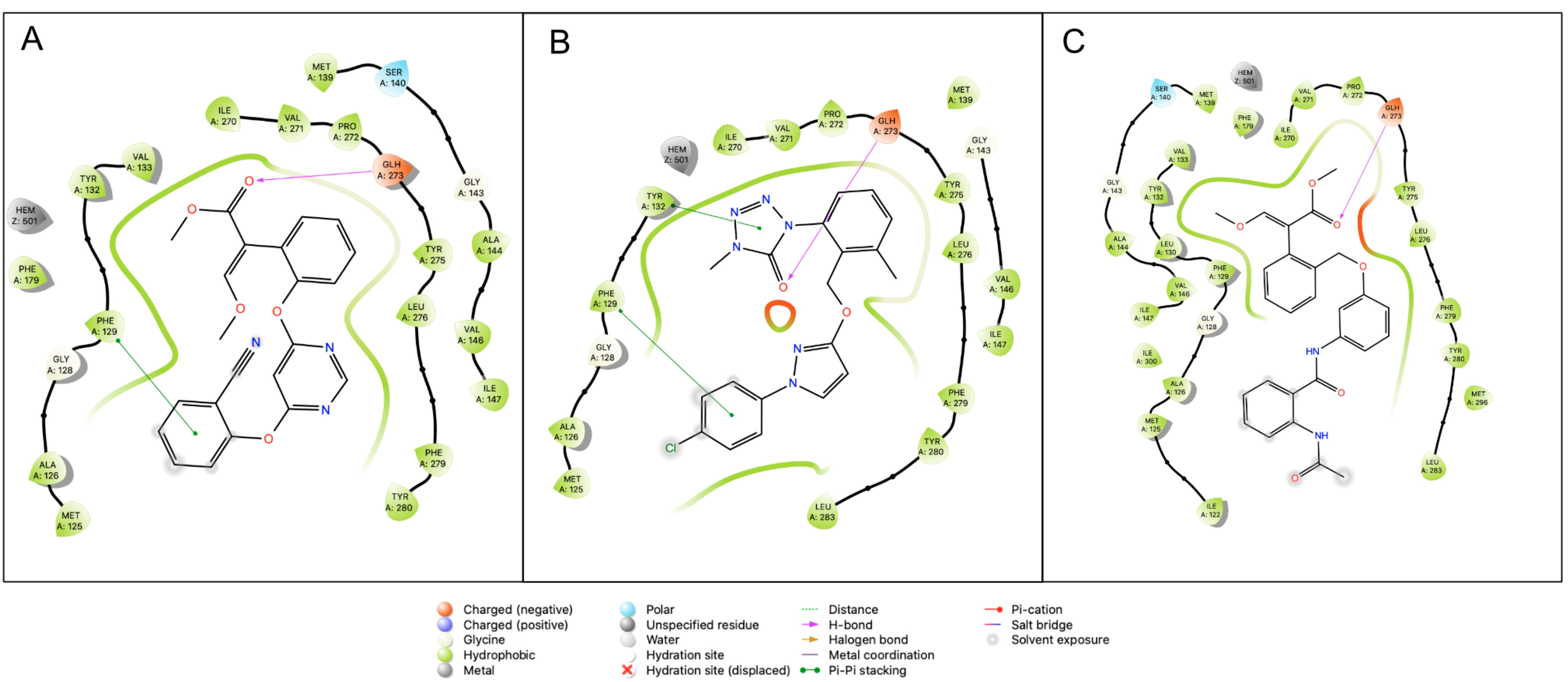
2.4. In Silico Modeling and Docking
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.2. Fungal Strains
4.3. Fungicides
4.4. Inhibition of Mycelium Growth of Pyricularia oryzae by Novel Dual Compounds
4.5. Enzyme Inhibition Assay for the Measurement of SDHI and QoI Action
4.6. Statistical Analysis
4.7. In Silico Modeling
4.7.1. Homology Modeling of Cytochrome bc1 Complex
4.7.2. Ligand Preparation
4.7.3. Molecular Docking and Affinity Calculations
4.7.4. Mutant Generation and Evaluation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FRAC. FRAC Code List©* 2020: Fungal Control Agents Sorted by Cross Resistance Pattern and Mode of Action. Available online: https://cpb-us-w2.wpmucdn.com/u.osu.edu/dist/b/28945/files/2020/02/frac-code-list-2020-final.pdf (accessed on 26 March 2021).
- Lamberth, C. Complex III inhibiting strobilurin esters, amides, and carbamates as broad-spectrum fungicides. In Bioactive Carboxylic Compound Classes: Pharmaceuticals and Agrochemicals; Wiley: Weinheim, Germany, 2016; pp. 371–384. [Google Scholar]
- Walter, H. Fungicidal succinate-dehydrogenase-inhibiting carboxamides. In Bioactive Carboxylic Compound Classes: Pharmaceuticals and Agrochemicals; Wiley: Weinheim, Germany, 2016; pp. 405–425. [Google Scholar]
- Xiong, L.; Shen, Y.-Q.; Jiang, L.-N.; Zhu, X.-L.; Yang, W.-C.; Huang, W.; Yang, G.-F. Succinate Dehydrogenase: An Ideal Target for Fungicide Discovery. In Discovery and Synthesis of Crop Protection Products; ACS Publications: Washington, DC, USA, 2015; pp. 175–194. ISBN 9780841231023. [Google Scholar]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Musso, L.; Fabbrini, A.; Dallavalle, S. Natural compound-derived cytochrome bc1 complex inhibitors as antifungal agents. Molecules 2020, 25, 4582. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, D.; Salat, M.; Frey, R.; Mosbach, A.; Luksch, T.; Balmer, D.; Hansen, R.; Widdison, S.; Logan, G.; Dietrich, R.A.; et al. A dispensable paralog of succinate dehydrogenase subunit C mediates standing resistance towards a subclass of SDHI fungicides in Zymoseptoria tritici. PLoS Pathog. 2019, 15, e1007780. [Google Scholar] [CrossRef] [PubMed]
- FRAC. FRAC List of Plant Pathogenic Organisms Resistant to Disease Control Agents. Available online: https://www.frac.info/docs/default-source/working-groups/sdhi-fungicides/group/list-of-resistant-plant-pathogens_2012-edition.pdf?sfvrsn=ef18469a_2 (accessed on 26 March 2021).
- Torriani, S.F.F.; Frey, R.; Buitrago, C.; Wullschleger, J.; Waldner, M.; Kuehn, R.; Scalliet, G.; Sierotzki, H. Succinate-dehydrogenase inhibitor (SDHI) resistance evolution in plant pathogens. In Modern Fungicides and Antifungal Compounds; Deising, H.B., Fraaije, B., Mehl, A., Oerke, E.C., Sierotzki, H., Stammler, G., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2016; Volume VIII, pp. 89–94. ISBN 978-3-941261-15-0. [Google Scholar]
- Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. The QoI fungicides, the rise and fall of a successful class of agricultural fungicides. In Fungicides; Carisse, O., Ed.; IntechOpen: London, UK, 2010; pp. 203–220. ISBN 978-953-307-266-1. [Google Scholar]
- FRAC. FRAC Reccomendations for QoI Fungicides. Available online: https://www.frac.info/frac-teams/working-groups/qol-fungicides/recommendations-for-qoi (accessed on 16 March 2021).
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Rice blast: A disease with implications for global food security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Nalley, L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.; Thoma, G. Economic and environmental impact of rice blast pathogen (Magnaporthe oryzae) alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, K.K.; Raimondi, J.V.; Marschalek, R.; de Andrade, A.; Wickert, E. Magnaporthe oryzae genetic diversity and its outcomes on the search for durable resistance. In The Molecular Basis of Plant Genetic Diversity; IntechOpen: London, UK, 2012; pp. 331–356. [Google Scholar]
- Ishii, H. Rice pathogens in Japan. In Fungicide Resistance in Plant Pathogens; Springer: Tokyo, Japan, 2015; pp. 341–354. [Google Scholar]
- Kunova, A.; Cortesi, P. Tricyclazole and azoxystrobin in rice blast management: A review of their activity and pathogen responses. In Fungicides: Classification, Role in Disease Management and Toxicity Effects; Wheeler, M.N., Johnston, B.R., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2013; pp. 39–66. ISBN 978-1-62948-045-9. [Google Scholar]
- Fitogest® Image Line Fitogest®. Available online: https://fitogest.imagelinenetwork.com/it/ (accessed on 6 November 2020).
- Zuccolo, M.; Kunova, A.; Musso, L.; Forlani, F.; Pinto, A.; Vistoli, G.; Gervasoni, S.; Cortesi, P.; Dallavalle, S. Dual-active antifungal agents containing strobilurin and SDHI-based pharmacophores. Sci. Rep. 2019, 9, 11377. [Google Scholar] [CrossRef] [PubMed]
- Suemoto, H.; Matsuzaki, Y.; Iwahashi, F. Metyltetraprole, a novel putative complex III inhibitor, targets known QoI-resistant strains of Zymoseptoria tritici and Pyrenophora teres. Pest Manag. Sci. 2019, 75, 1181–1189. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, Y.; Yoshimoto, Y.; Arimori, S.; Kiguchi, S.; Harada, T.; Iwahashi, F. Discovery of metyltetraprole: Identification of tetrazolinone pharmacophore to overcome QoI resistance. Bioorg. Med. Chem. 2020, 28, 115211. [Google Scholar] [CrossRef]
- Esser, L.; Quinn, B.; Li, Y.-F.; Zhang, M.; Elberry, M.; Yu, L.; Yu, C.-A.; Xia, D. Crystallographic studies of quinol oxidation site inhibitors: A modified classification of inhibitors for the cytochrome bc 1 complex. J. Mol. Biol. 2004, 341, 281–302. [Google Scholar] [CrossRef]
- Palsdottir, H.; Lojero, C.G.; Trumpower, B.L.; Hunte, C. Structure of the yeast cytochrome bc1 complex with a hydroxyquinone anion Qo site inhibitor bound. J. Biol. Chem. 2003, 278, 31303–31311. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.M.; Jung, C.K.; Choi, E.Y.; Gao, C.; Kim, S.W.; Lee, S.H.; Kwon, O.P. Highly conductive polyaniline copolymers with dual-functional hydrophilic dioxyethylene side chains. Polymer 2011, 52, 4451–4455. [Google Scholar] [CrossRef]
- Huang, R.; Li, Z.; Ren, P.; Chen, W.; Kuang, Y.; Chen, J.; Zhan, Y.; Chen, H.; Jiang, B. N- Phenyl- N- aceto-vinylsulfonamides as efficient and chemoselective handles for N-terminal modification of peptides and proteins. Eur. J. Org. Chem. 2018, 2018, 829–836. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Q.; Zhao, W.; Xu, S.; Zhang, W.; Chen, J. Palladium-catalyzed ortho-arylation of benzoic acid derivatives via C-H bond activation using an aminoacetic acid bidentate directing group. Tetrahedron Lett. 2015, 56, 851–855. [Google Scholar] [CrossRef]
- Crestey, F.; Frederiksen, K.; Jensen, H.S.; Dekermendjian, K.; Larsen, P.H.; Bastlund, J.F.; Lu, D.; Liu, H.; Yang, C.R.; Grunnet, M.; et al. Identification and electrophysiological evaluation of 2-methylbenzamide derivatives as Nav1.1 modulators. ACS Chem. Neurosci. 2015, 6, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Baramov, T.; Schmid, B.; Ryu, H.; Jeong, J.; Keijzer, K.; von Eckardstein, L.; Baik, M.; Süssmuth, R.D. How many O-donor groups in enterobactin does it take to bind a metal cation? Chem. Eur. J. 2019, 25, 6955–6962. [Google Scholar] [CrossRef]
- Vilaivan, T. A rate enhancement of tert-butoxycarbonylation of aromatic amines with Boc2O in alcoholic solvents. Tetrahedron Lett. 2006, 47, 6739–6742. [Google Scholar] [CrossRef]
- Kunova, A.; Pizzatti, C.; Cortesi, P. Impact of tricyclazole and azoxystrobin on growth, sporulation and secondary infection of the rice blast fungus, Magnaporthe oryzae. Pest Manag. Sci. 2013, 69, 278–284. [Google Scholar] [CrossRef]
- Kunova, A.; Pizzatti, C.; Bonaldi, M.; Cortesi, P. Sensitivity of nonexposed and exposed populations of Magnaporthe oryzae from rice to tricyclazole and azoxystrobin. Plant Dis. 2014, 98, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.-H.; Ma, L.; Dai, Z.-C.; Xiao, Y.; Zhang, Y.-Y.; Li, D.-D.; Wang, J.-X.; Zhu, H.-L. Synthesis and antifungal activity of nicotinamide derivatives as succinate dehydrogenase inhibitors. J. Agric. Food Chem. 2014, 62, 4063–4071. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, F.; Li, H.; Yang, W.; Chen, Q.; Yang, G. Design, synthesis, and bioevaluation of novel strobilurin derivatives. Chin. J. Chem. 2012, 30, 1999–2008. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team, The R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Faria, J.C.; Jelihovschi, E.G.; Allaman, I.B. Conventional Tukey Test; UESC: Ilheus, Brasil, 2020. [Google Scholar]
- Schrödinger Release 2020-3; BioLuminate, Schrödinger, LLC: New York, NY, USA, 2020.
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A force field providing broad coverage of drug-like small molecules and proteins. J. Chem. Theory Comput. 2016, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2020-3; Schrödinger, LLC: New York, NY, USA, 2020.
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A new approach for rapid, accurate docking and scoring. 2. Enrichment factors in database screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Abel, R.; Zhu, K.; Cao, Y.; Zhao, S.; Friesner, R.A. The VSGB 2.0 model: A next generation energy model for high resolution protein structure modeling. Proteins Struct. Funct. Bioinform. 2011, 79, 2794–2812. [Google Scholar] [CrossRef] [Green Version]
- Eberini, I.; Rocco, A.G.; Mantegazza, M.; Gianazza, E.; Baroni, A.; Vilardo, M.C.; Donghi, D.; Galliano, M.; Beringhelli, T. Computational and experimental approaches assess the interactions between bovine β-lactoglobulin and synthetic compounds of pharmacological interest. J. Mol. Graph. Model. 2008, 26, 1004–1013. [Google Scholar] [CrossRef]
- Beard, H.; Cholleti, A.; Pearlman, D.; Sherman, W.; Loving, K.A. Applying physics-based scoring to calculate free energies of binding for single amino acid mutations in protein-protein complexes. PLoS ONE 2013, 8, e82849. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | RES | |||
|---|---|---|---|---|
| Compound | % Inhibition a | Tukey HSD b | % Inhibition | Tukey HSD |
| Control | 0 | a | 0 | a |
| ACT | 0 | a | −4 | a |
| AZX | 92 | m | 47 | c |
| FXP | 67 | f h i | 94 | d |
| 1a | 72 | h i j | 20 | b |
| 7 | 25 | b c | 26 | b |
| 8 | 67 | f g h i | 26 | b |
| 9 | 63 | e f g h | 28 | b |
| 10 | 78 | j k l | 52 | c |
| 11a | 79 | i j k l | 30 | b |
| 11b | 83 | k l | 84 | d |
| 11c | 71 | h i j | 35 | b |
| 11d | 58 | e f g | 23 | b |
| 11e | 84 | k l m | 50 | c |
| 11f | 58 | e f | 23 | b |
| 11g | 55 | e | 27 | b |
| 12 | 73 | h i j k | 22 | b |
| 13 | 87 | l m | 57 | c |
| 14a | 42 | d | 15 | b |
| 14b | 38 | d | 20 | b |
| 14c | 18 | b | 15 | b |
| 15 | 31 | c d | 21 | b |
| Ligand | Glide XP a [kcal/mol] | MM-GBSA b [kcal/mol] | Number of Poses c | Range d [kcal/mol] | Orientation of the Generated Poses e |
|---|---|---|---|---|---|
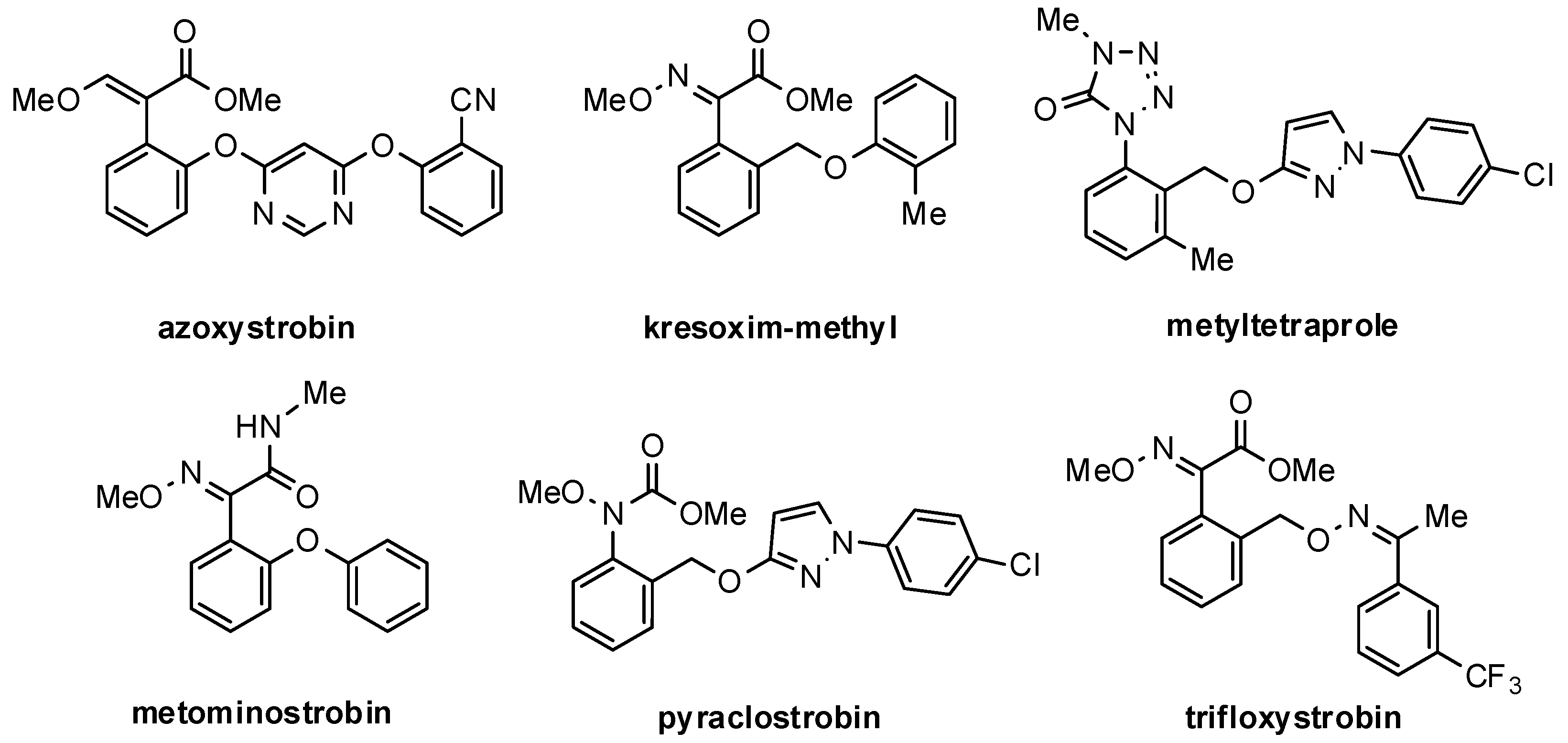
| azoxystrobin | −10.6 | −76.4 | 5 | (−10.6; −9.4) | 5/5 |
| trifloxystrobin | −9.8 | −65.6 | 2 | (−9.8; −9.7) | 2/2 |
| kresoxym-methyl | −10.4 | −68.9 | 1 | (−10.4) | 1/1 |
| metominostrobin | −9.9 | −79.3 | 1 | (−9.9) | 1/1 |
| pyraclostrobin | −8.4 | −61.7 | 5 | (−10.0; −5.7) | 3/5 |
| metyltetraprole | −10.2 | −62.9 | 2 | (−10.2; −9.8) | 2/2 |
| Ligand | Glide XP a [kcal/mol] | MM-GBSA b [kcal/mol] | Number of Poses c | Range d [kcal/mol] | Orientation of the Generated Poses e |
|---|---|---|---|---|---|
| 10 | −11.7 | −83.9 | 3 | (−11.7; −9.1) | 3/3 |
| 11a | −9.1 | −60.3 | 5 | (−9.1; −7.4) | 1/5 |
| 11b | −9.4 | −59.6 | 5 | (−9.4; −7.4) | 3/5 |
| 11e | −12.6 | −83.3 | 5 | (−12.6; −11.6) | 5/5 |
| 13 | −12.2 | −78.7 | 2 | (−12.2; −10.8) | 2/2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunova, A.; Palazzolo, L.; Forlani, F.; Catinella, G.; Musso, L.; Cortesi, P.; Eberini, I.; Pinto, A.; Dallavalle, S. Structural Investigation and Molecular Modeling Studies of Strobilurin-Based Fungicides Active against the Rice Blast Pathogen Pyricularia oryzae. Int. J. Mol. Sci. 2021, 22, 3731. https://doi.org/10.3390/ijms22073731
Kunova A, Palazzolo L, Forlani F, Catinella G, Musso L, Cortesi P, Eberini I, Pinto A, Dallavalle S. Structural Investigation and Molecular Modeling Studies of Strobilurin-Based Fungicides Active against the Rice Blast Pathogen Pyricularia oryzae. International Journal of Molecular Sciences. 2021; 22(7):3731. https://doi.org/10.3390/ijms22073731
Chicago/Turabian StyleKunova, Andrea, Luca Palazzolo, Fabio Forlani, Giorgia Catinella, Loana Musso, Paolo Cortesi, Ivano Eberini, Andrea Pinto, and Sabrina Dallavalle. 2021. "Structural Investigation and Molecular Modeling Studies of Strobilurin-Based Fungicides Active against the Rice Blast Pathogen Pyricularia oryzae" International Journal of Molecular Sciences 22, no. 7: 3731. https://doi.org/10.3390/ijms22073731
APA StyleKunova, A., Palazzolo, L., Forlani, F., Catinella, G., Musso, L., Cortesi, P., Eberini, I., Pinto, A., & Dallavalle, S. (2021). Structural Investigation and Molecular Modeling Studies of Strobilurin-Based Fungicides Active against the Rice Blast Pathogen Pyricularia oryzae. International Journal of Molecular Sciences, 22(7), 3731. https://doi.org/10.3390/ijms22073731