
Combining Magnetic Resonance Imaging with Systemic Monocyte Evaluation for the Implementation of GBM Management

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
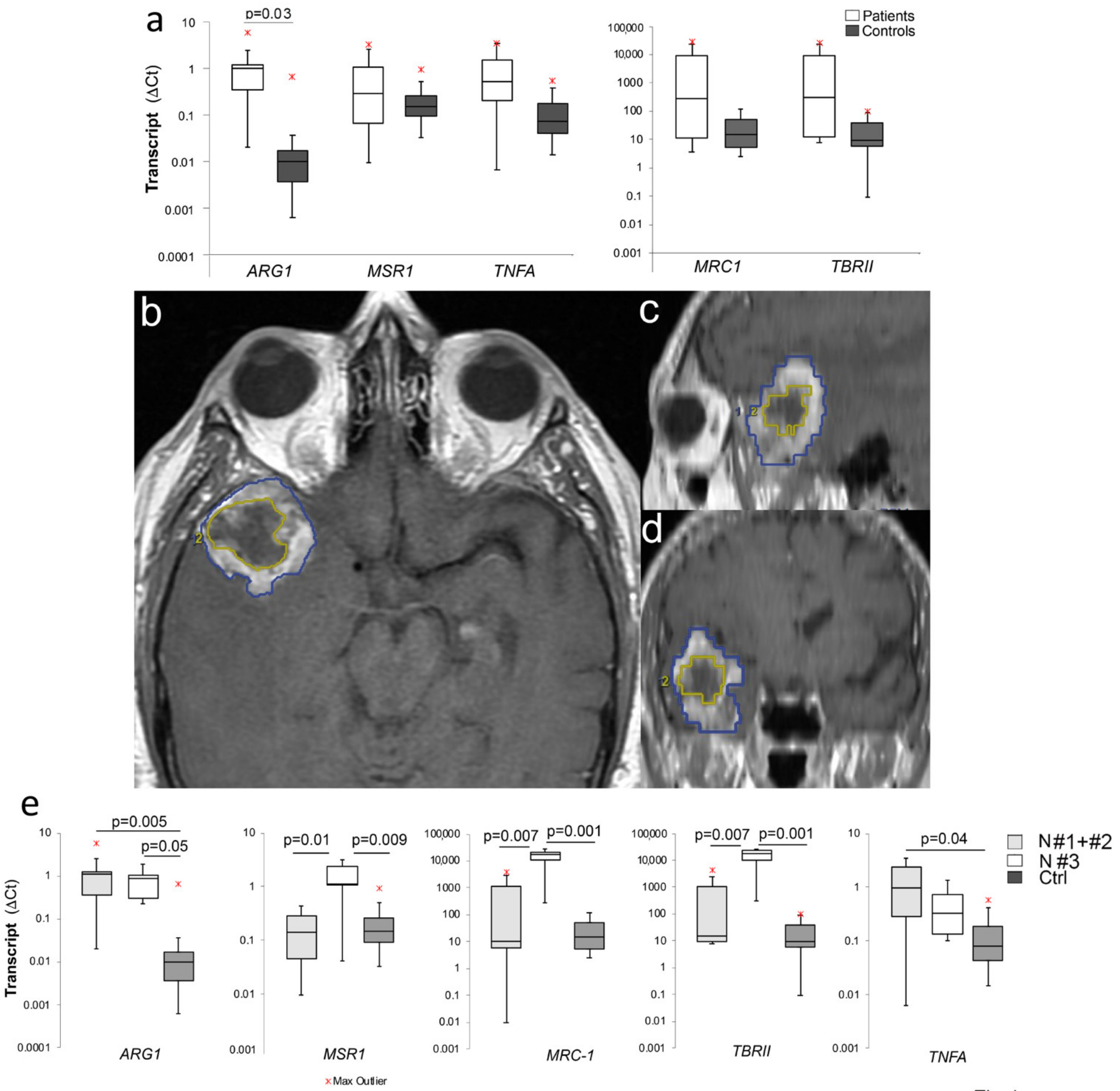
2.1. Increased M2 Gene Expression in PBMCs of GBM Patients
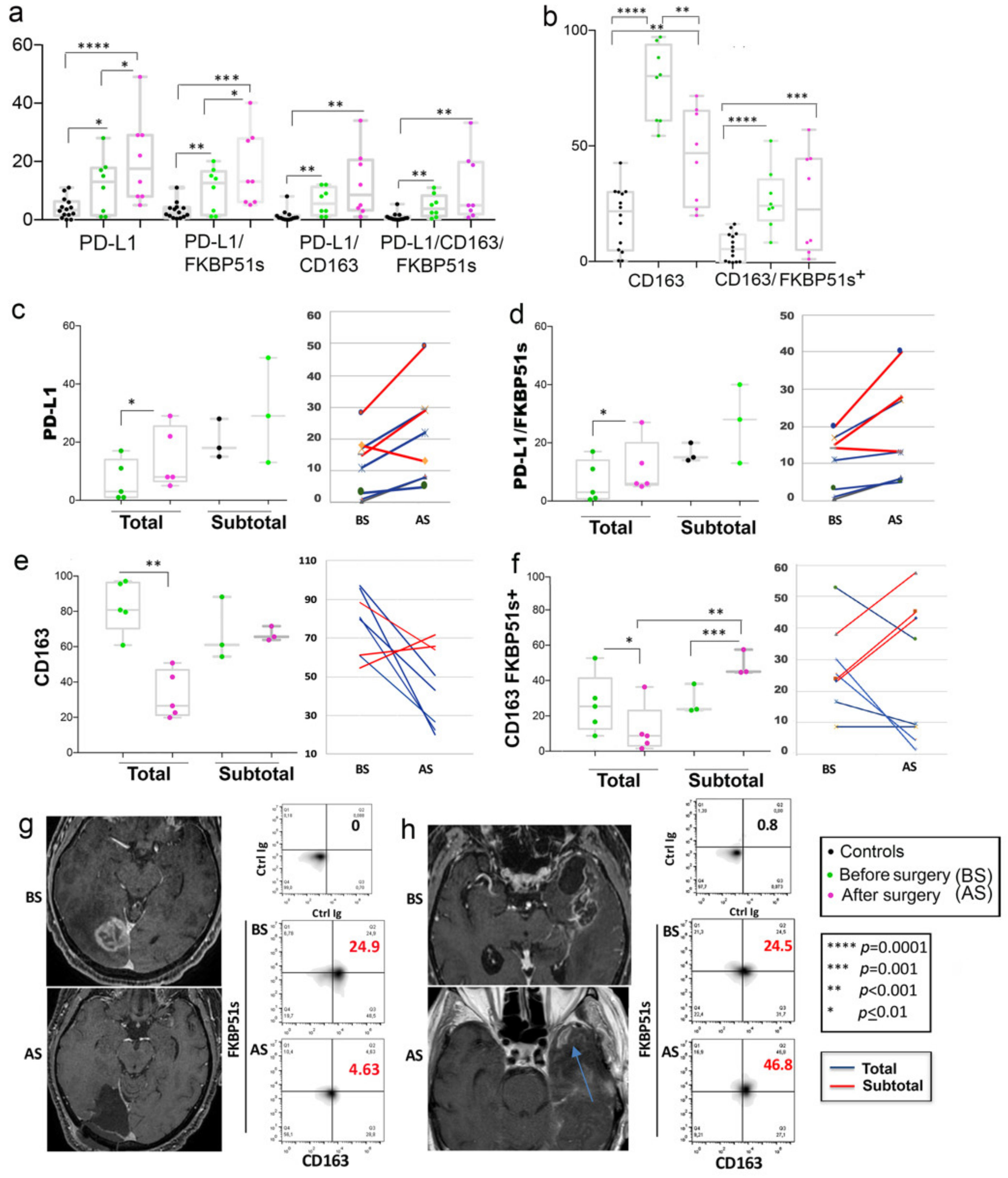
2.2. High Frequency of PD-L1+ and CD163+ Monocytes in Peripheral Blood of GBM Patients
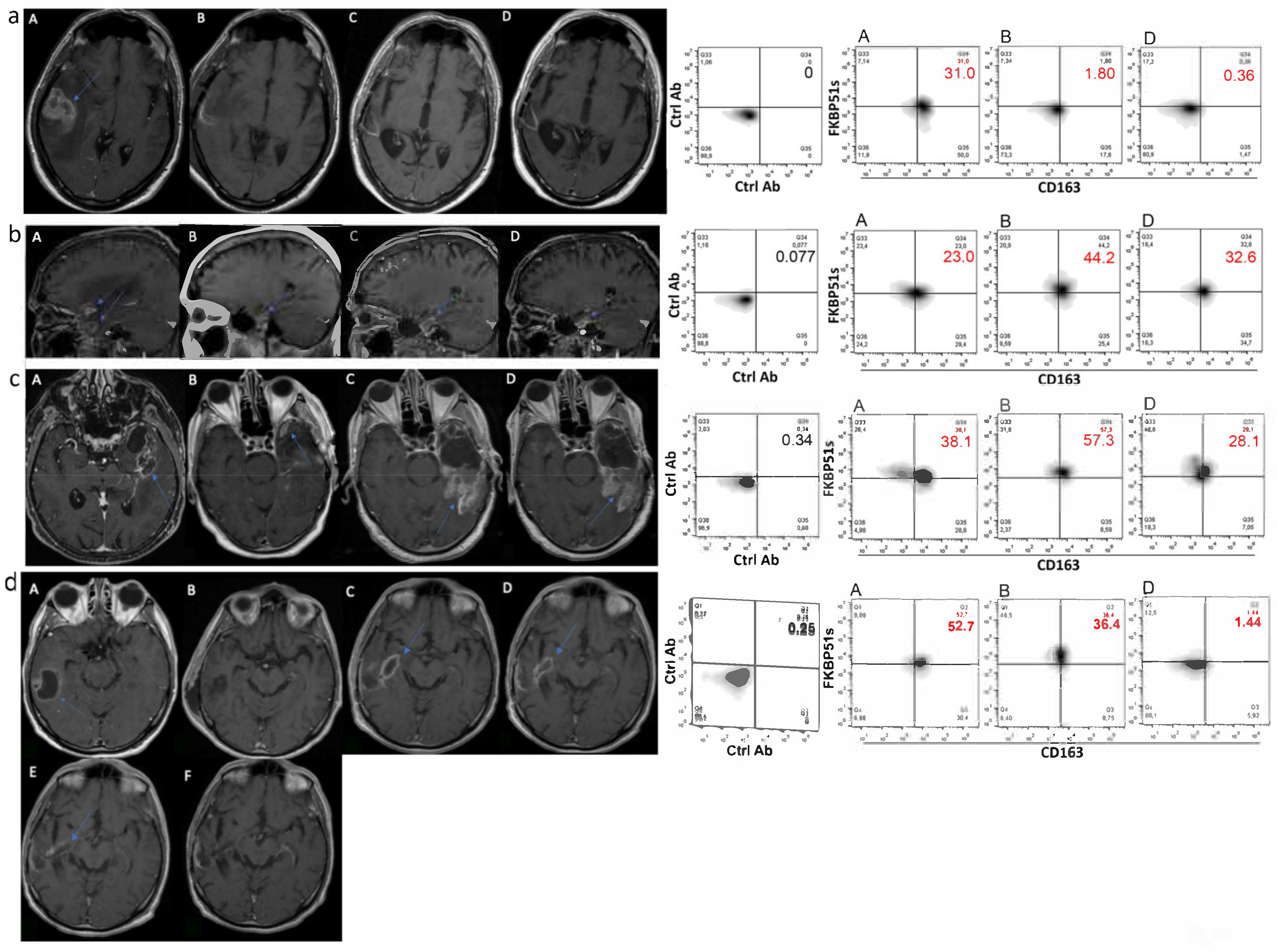
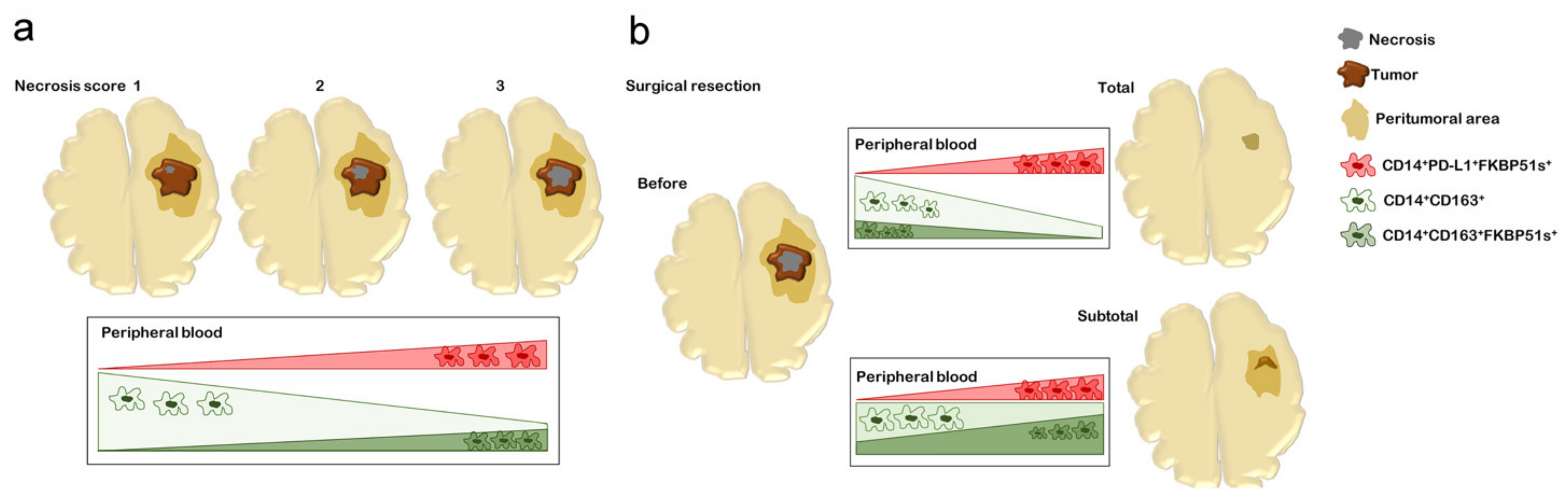
2.3. CD163+FKBP51s+ Monocytes Are Sensitive to Tumor Removal
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Neuropathology
4.3. Imaging and Image Analysis
4.4. Peripheral Blood Mononuclear Cell (PBMC) Isolation
4.5. Flow Cytometry Analysis
4.6. qPCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Farah, P.; Ondracek, A.; Chen, Y.; Wolinsky, Y.; Stroup, N.E.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 2013, 15 (Suppl. S2), ii1–ii56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Ha, E.T.; Antonios, J.P.; Soto, H.; Prins, R.M.; Yang, I.; Kasahara, N.; Liau, L.M.; Kruse, C.A. Chronic inflammation drives glioma growth: Cellular and molecular factors responsible for an immunosuppressive microenvironment. Neuroimmunol. Neuroinflamm. 2014, 1, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Kalpathy-Cramer, J.; Gerstner, E.R.; Emblem, K.E.; Andronesi, O.; Rosen, B. Advanced magnetic resonance imaging of the physical processes in human glioblastoma. Cancer Res. 2014, 74, 4622–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulseh, A.M.; Keller, J.; Klener, J.; Sroubek, J.; Dbalý, V.; Syrůček, M.; Tovaryš, F.; Vymazal, J. Long-term survival of patients suffering from glioblastoma multiforme treated with tumor-treating fields. World J. Surg. Oncol. 2012, 10, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orringer, D.; Lau, D.; Khatri, S.; Zamora-Berridi, G.J.; Zhang, K.; Wu, C.; Chaudhary, N.; Sagher, O. Extent of resection in patients with glioblastoma: Limiting factors, perception of resectability, and effect on survival. J. Neurosurg. 2012, 117, 851–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Pepa, G.M.; Ius, T.; La Rocca, G.; Gaudino, S.; Isola, M.; Pignotti, F.; Rapisarda, A.; Mazzucchi, E.; Giordano, C.; Dragonetti, V.; et al. 5-Aminolevulinic Acid and Contrast-Enhanced Ultrasound: The Combination of the Two Techniques to Optimize the Extent of Resection in Glioblastoma Surgery. Neurosurgery 2020, 86, E529–E540. [Google Scholar] [CrossRef]
- Thust, S.C.; van den Bent, M.J.; Smits, M. Pseudoprogression of brain tumors. J. Magn. Reson. Imaging 2018, 48, 571–589. [Google Scholar] [CrossRef] [Green Version]
- Hygino da Cruz, L.C.; Rodriguez, I.; Domingues, R.C.; Gasparetto, E.L.; Sorensen, A.G. Pseudoprogression and Pseudoresponse: Imaging Challenges in the Assessment of Posttreatment Glioma. AJNR Am. J. Neuroradiol. 2011, 32, 1978–1985. [Google Scholar] [CrossRef] [Green Version]
- Chae, M.; Peterson, T.E.; Balgeman, A.; Chen, S.; Zhang, L.; Renner, D.N.; Johnson, A.J.; Parney, I.F. Increasing glioma-associated monocytes leads to increased intratumoral and systemic myeloid-derived suppressor cells in a murine model. Neuro Oncol. 2015, 17, 978–991. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Feng, X.; Herting, C.J.; Garcia, V.A.; Nie, K.; Pong, W.W.; Rasmussen, R.; Dwivedi, B.; Seby, S.; Wolf, S.A.; et al. Cellular and Molecular Identity of Tumor-Associated Macrophages in Glioblastoma. Cancer Res. 2017, 77, 2266–2278. [Google Scholar] [CrossRef] [Green Version]
- Friebel, E.; Kapolou, K.; Unger, S.; Núñez, N.G.; Utz, S.; Rushing, E.J.; Regli, L.; Weller, M.; Greter, M.; Tugues, S.; et al. Single-Cell Mapping of Human Brain Cancer Reveals Tumor-Specific Instruction of Tissue-Invading Leukocytes. Cell 2020, 181, 1626–1642.e20. [Google Scholar] [CrossRef]
- London, A.; Cohen, M.; Schwartz, M. Microglia and monocyte-derived macrophages: Functionally distinct populations that act in concert in CNS plasticity and repair. Front. Cell. Neurosci. 2013, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hambardzumyan, D. Immune Microenvironment in Glioblastoma Subtypes. Front. Immunol. 2018, 9, 1004. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhang, C.; Maimela, N.R.; Yang, L.; Zhang, Z.; Ping, Y.; Huang, L.; Zhang, Y. Molecular and clinical characterization of CD163 expression via large-scale analysis in glioma. Oncoimmunology 2019, 8, 1601478. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhang, H.; Chen, B.; Liu, X.; Zhang, S.; Zong, Z.; Gao, M. PD-L1-Mediated Immunosuppression in Glioblastoma Is Associated with the Infiltration and M2-Polarization of Tumor-Associated Macrophages. Front. Immunol. 2020, 11, 588552. [Google Scholar] [CrossRef]
- Bloch, O.; Crane, C.A.; Kaur, R.; Safaee, M.; Rutkowski, M.J.; Parsa, A.T. Gliomas promote immunosuppression through induction of B7-H1 expression in tumor-associated macrophages. Clin. Cancer Res. 2013, 19, 3165–3175. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.M.; Choi, J.; Lim, M. Mechanisms of immunotherapy resistance: Lessons from glioblastoma. Nat. Immunol. 2019, 20, 1100–1109. [Google Scholar] [CrossRef]
- Romano, S.; Simeone, E.; D’Angelillo, A.; D’Arrigo, P.; Russo, M.; Capasso, M.; Lasorsa, V.A.; Zambrano, N.; Ascierto, P.A.; Romano, M.F. FKBP51s signature in peripheral blood mononuclear cells of melanoma patients as a possible predictive factor for immunotherapy. Cancer Immunol. Immunother. 2017, 66, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Troiani, T.; Giunta, E.F.; Tufano, M.; Vigorito, V.; Arrigo, P.; Argenziano, G.; Ciardiello, F.; Romano, M.F.; Romano, S. Alternative macrophage polarisation associated with resistance to anti-PD1 blockade is possibly supported by the splicing of FKBP51 immunophilin in melanoma patients. Br. J. Cancer 2020, 122, 1782–1790. [Google Scholar] [CrossRef]
- Somarelli, J.A.; Lee, S.Y.; Skolnick, J.; Herrera, R.J. Structure-based classification of 45 FK506-binding proteins. Proteins 2008, 72, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, J.; Smith, D.F. Molecular chaperone interactions with steroid receptors: An update. Mol. Endocrinol. 2000, 14, 939–946. [Google Scholar] [CrossRef]
- Romano, S.; Xiao, Y.; Nakaya, M.; D’Angelillo, A.; Chang, M.; Jin, J.; Hausch, F.; Masullo, M.; Feng, X.; Romano, M.F.; et al. FKBP51 employs both scaffold and isomerase functions to promote NF-κB activation in melanoma. Nucleic Acids Res. 2015, 43, 6983–6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baughman, G.; Wiederrecht, G.J.; Campbell, N.F.; Martin, M.M.; Bourgeois, S. FKBP51, a novel T-cell specific immunophilin capable of calcineurin inhibition. Mol. Cell. Biol. 1995, 15, 4395–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, S.; D’Angelillo, A.; Staibano, S.; Simeone, E.; D’Arrigo, P.; Ascierto, P.A.; Scalvenzi, M.; Mascolo, M.; Ilardi, G.; Merolla, F.; et al. Immunomodulatory pathways regulate expression of a spliced FKBP51 isoform in lymphocytes of melanoma patients. Pigment Cell Melanoma Res. 2015, 28, 442–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arrigo, P.; Russo, M.; Rea, A.; Tufano, M.; Guadagno, E.; Del Basso De Caro, M.L.; Pacelli, R.; Hausch, F.; Staibano, S.; Ilardi, G.; et al. A regulatory role for the co-chaperone FKBP51s in PD-L1 expression in glioma. Oncotarget 2017, 8, 68291–68304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, S.M.; Lang, F.F.; Aggarwal, B.B.; Fuller, G.N.; Wildrick, D.M.; Sawaya, R. Necrosis and glioblastoma: A friend or a foe? A review and a hypothesis. Neurosurgery 2002, 51, 2–12. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: New molecules and patterns of gene expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [Green Version]
- Pierallini, A.; Bonamini, M.; Pantano, P.; Palmeggiani, F.; Raguso, M.; Osti, M.F.; Anaveri, G.; Bozzao, L. Radiological assessment of necrosis in glioblastoma: Variability and prognostic value. Neuroradiology 1998, 40, 150–153. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Y.; Xu, K.; Wang, Z.; Fan, X.; Zhang, C.; Li, S.; Qiu, X.; Jiang, T. Relationship between necrotic patterns in glioblastoma and patient survival: Fractal dimension and lacunarity analyses using magnetic resonance imaging. Sci. Rep. 2017, 7, 8302. [Google Scholar] [CrossRef]
- Li, W.; Graeber, M.B. The molecular profile of microglia under the influence of glioma. Neuro Oncol. 2012, 14, 958–978. [Google Scholar] [CrossRef] [Green Version]
- Huettner, C.; Czub, S.; Kerkau, S.; Roggendorf, W.; Tonn, J.C. Interleukin 10 is expressed in human gliomas in vivo and increases glioma cell proliferation and motility in vitro. Anticancer Res. 1997, 17, 3217–3224. [Google Scholar]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [Green Version]
- Gielen, P.R.; Schulte, B.M.; Kers-Rebel, E.D.; Verrijp, K.; Bossman, S.A.; Ter Laan, M.; Wesseling, P.; Adema, G.J. Elevated levels of polymorphonuclear myeloid-derived suppressor cells in patients with glioblastoma highly express S100A8/9 and arginase and suppress T cell function. Neuro Oncol. 2016, 18, 1253–1264. [Google Scholar] [CrossRef]
- Gabrusiewicz, K.; Li, X.; Wei, J.; Hashimoto, Y.; Marisetty, A.L.; Ott, M.; Wang, F.; Hawke, D.; Yu, J.; Healy, L.M.; et al. Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. Oncoimmunology 2018, 7, e1412909. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Minutti, C.M.; Knipper, J.A.; Allen, J.E.; Zaiss, D.M. Tissue-specific contribution of macrophages to wound healing. Semin. Cell Dev. Biol. 2017, 61, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronning, P.A.; Helseth, E.; Meling, T.R.; Johannesen, T.B. A population-based study on the effect of temozolomide in the treatment of glioblastoma multiforme. Neuro Oncol. 2012, 14, 1178–1184. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.; Ogaki, H.; Wiestler, O.D.; Cavenee, W.K.; Ellison, D.W.; Figarella-Branger, D.; Perry, A.; Reifenberger, G.; von Deimling, A. WHO Classification of Tumours of the Central Nervous System, Revised 4th, ed.; Louis, D.N., Ohgaki, H., Wiestler, O.D., Cavenee, W.K., Eds.; IARC Press: Lyon, France, 2016; Volume 1, pp. 10–76. [Google Scholar]
- Park, M.J.; Park, M.J.; Kim, H.S.; Jahng, G.H.; Ryu, C.W.; Park, S.M.; Kim, S.Y. Semiquantitative assessment of intratumoral susceptibility signals using non-contrast-enhanced high-field high-resolution susceptibility-weighted imaging in patients with gliomas: Comparison with MR perfusion imaging. Am. J. Neuroradiol. 2009, 7, 1402–1408. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Reddy, M.V.; Wilson, B.K.J.; Blair, D.A.; Taha, A.; Frampton, C.M.; Eiholzer, R.A.; Gan, P.Y.C.; Ziad, F.; Thotathil, Z.; et al. MR Imaging Characteristics Associate with Tumor-Associated Macrophages in Glioblastoma and Provide an Improved Signature for Survival Prognostication. AJNR Am. J. Neuroradiol. 2018, 39, 252–259. [Google Scholar] [CrossRef]
- Grimaldi, E.; Carandente, P.; Scopacasa, F.; Romano, M.F.; Pellegrino, M.; Bisogni, R.; De Caterina, M. Evaluation of the monocyte counting by two automated haematology analysers compared with flow cytometry. Clin. Lab. Haematol. 2005, 27, 91–97. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
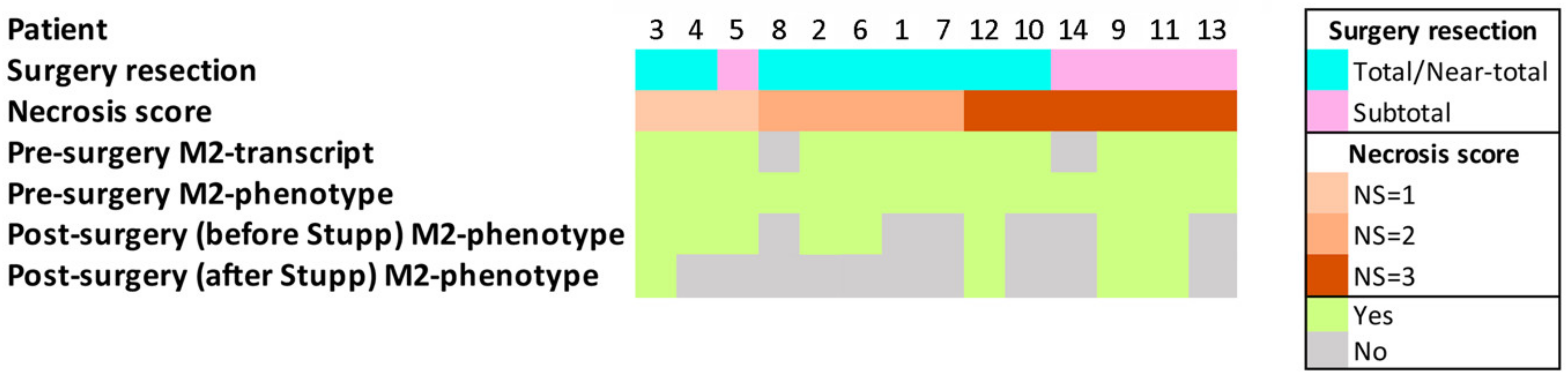
| Patient | Gender | Age | Localization | Pathology | SR | NS | EE | CCI | MS | MH | ES | ITSS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | M | 61 | occipital | GBM IDH wt | Total | 2 | NO | NO | NO | NO | 1 | 3 |
| 2 | F | 45 | frontal | GBM IDH mutant | Total | 2 | NO | NO | NO | NO | 0 | 3 |
| 3 | M | 75 | temporo-occipital | GBM IDH wt | Near-total | 1 | YES | YES | YES | YES | 0 | 3 |
| 4 | F | 59 | frontal | GBM IDH wt | Subtotal | 1 | YES | YES | YES | NO | 1 | 3 |
| 5 | M | 55 | temporal | GBM IDH wt | Total | 1 | NO | NO | YES | NO | 1 | 3 |
| 6 | M | 41 | temporal | GBM IDH wt | Near-total | 2 | YES | NO | YES | NO | 0 | 3 |
| 7 | M | 63 | temporal | GBM IDH wt | Total | 2 | NO | NO | NO | NO | 0 | 2 |
| 8 | F | 60 | parieto-occipital | GBM IDH wt | Near-total | 2 | NO | YES | YES | NO | 0 | 3 |
| 9 | M | 78 | temporal | GBM IDH wt | Subtotal | 3 | NO | NO | NO | NO | 0 | 2 |
| 10 | M | 66 | temporo-parietal | GBM IDH wt | Near-total | 3 | NO | NO | NO | NO | 1 | 2 |
| 11 | M | 52 | temporal | GBM IDH wt | Subtotal | 3 | NO | NO | NO | NO | 1 | 3 |
| 12 | M | 55 | temporal | GBM IDH wt | Total | 3 | NO | NO | NO | NO | 0 | 3 |
| 13 | M | 66 | parieto-occipital | GBM IDH wt | Subtotal | 3 | NO | YES | NO | NO | 0 | 3 |
| 14 | M | 52 | frontal | GBM IDH wt | Subtotal | 3 | NO | YES | NO | NO | 0 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, C.; Sabatino, G.; Romano, S.; Della Pepa, G.M.; Tufano, M.; D’Alessandris, Q.G.; Cottonaro, S.; Gessi, M.; Balducci, M.; Romano, M.F.; et al. Combining Magnetic Resonance Imaging with Systemic Monocyte Evaluation for the Implementation of GBM Management. Int. J. Mol. Sci. 2021, 22, 3797. https://doi.org/10.3390/ijms22073797
Giordano C, Sabatino G, Romano S, Della Pepa GM, Tufano M, D’Alessandris QG, Cottonaro S, Gessi M, Balducci M, Romano MF, et al. Combining Magnetic Resonance Imaging with Systemic Monocyte Evaluation for the Implementation of GBM Management. International Journal of Molecular Sciences. 2021; 22(7):3797. https://doi.org/10.3390/ijms22073797
Chicago/Turabian StyleGiordano, Carolina, Giovanni Sabatino, Simona Romano, Giuseppe Maria Della Pepa, Martina Tufano, Quintino Giorgio D’Alessandris, Simone Cottonaro, Marco Gessi, Mario Balducci, Maria Fiammetta Romano, and et al. 2021. "Combining Magnetic Resonance Imaging with Systemic Monocyte Evaluation for the Implementation of GBM Management" International Journal of Molecular Sciences 22, no. 7: 3797. https://doi.org/10.3390/ijms22073797
APA StyleGiordano, C., Sabatino, G., Romano, S., Della Pepa, G. M., Tufano, M., D’Alessandris, Q. G., Cottonaro, S., Gessi, M., Balducci, M., Romano, M. F., Olivi, A., Gaudino, S., & Colosimo, C. (2021). Combining Magnetic Resonance Imaging with Systemic Monocyte Evaluation for the Implementation of GBM Management. International Journal of Molecular Sciences, 22(7), 3797. https://doi.org/10.3390/ijms22073797