
Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.)

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
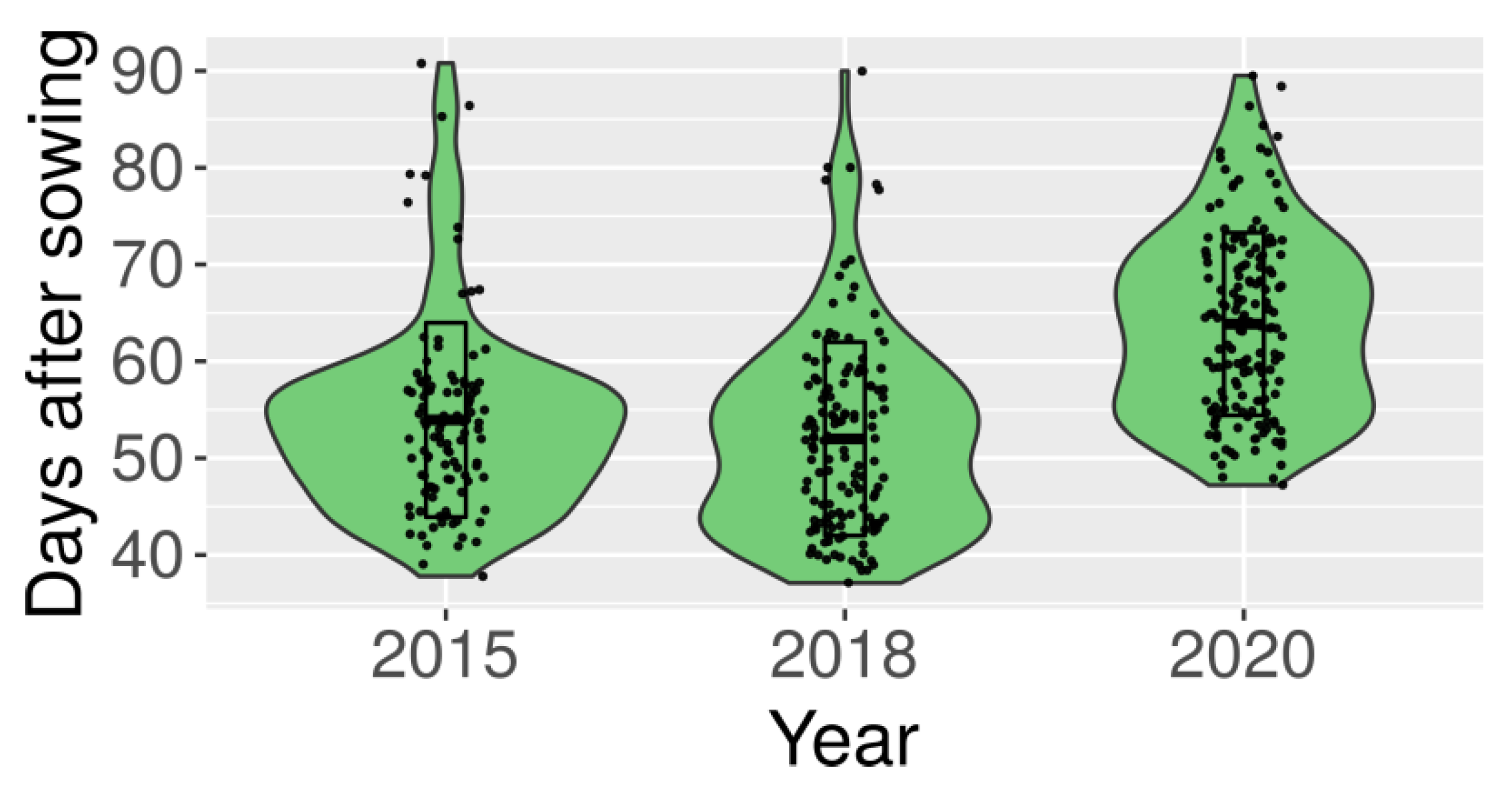
2.1. Early Flowering and Thermoneutrality Is Present in Primitive and Domesticated Germplasm
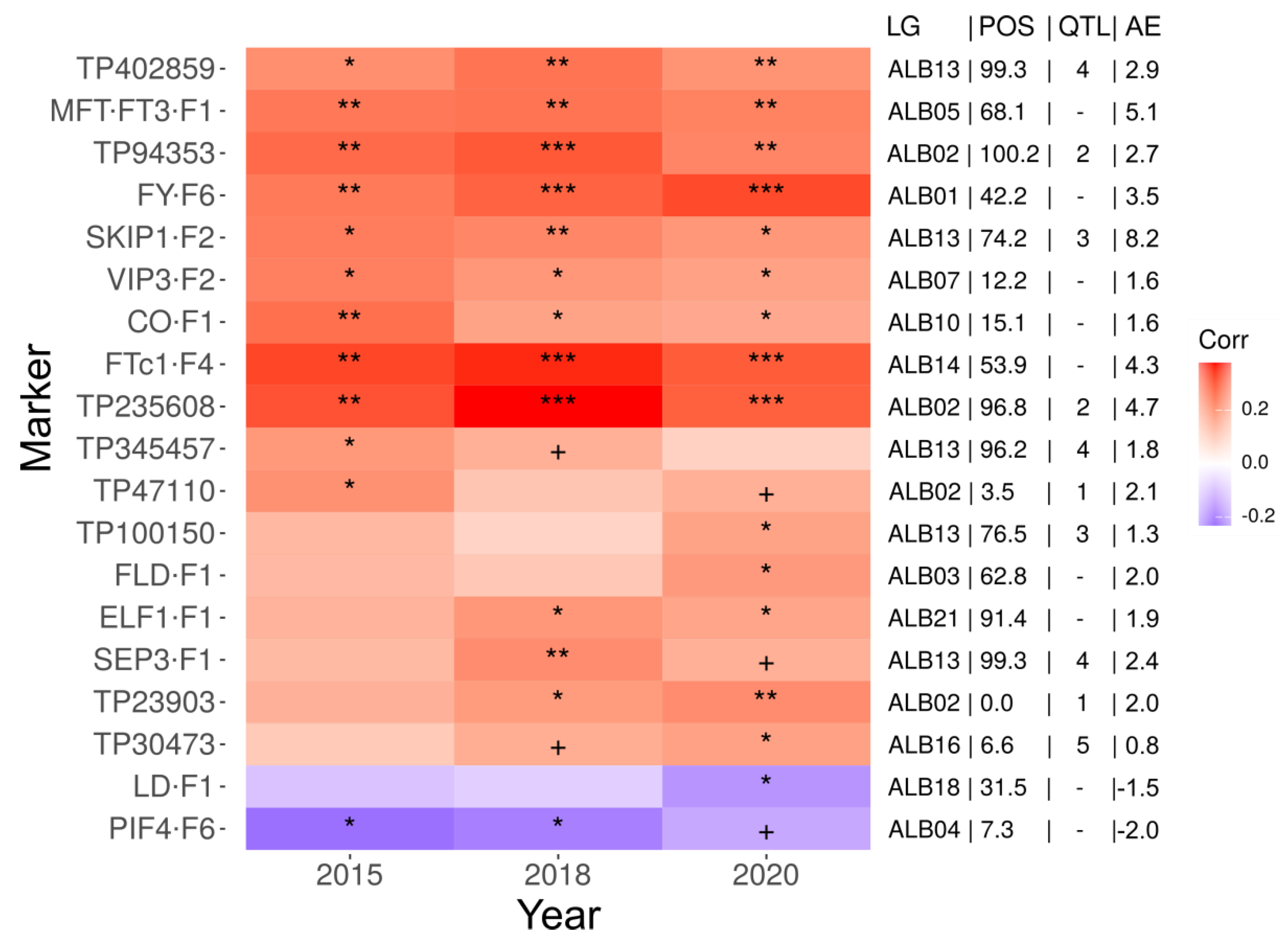
2.2. Flowering Induction in White Lupin Germplasm Collection Is under Polygenic Control
2.3. Several White Lupin Subpopulations Differring in Flowering Time and Allelic Composition Were Identified
3. Discussion
3.1. Contribution of FLOWERING LOCUS T Genes to Early Flowering in White Lupin
3.2. Candidates from Photoperiod, Vernalization, Autonomous and Heat-Responsive Pathways for Flowering Time Control in White Lupin
3.3. Perspectives for Molecular-Assisted Breeding of White Lupin
3.4. Concluding Remarks
4. Materials and Methods
4.1. Plant Material
4.2. Profiling of Time to Flowering and Vernalization Responsiveness in Controlled Environment
4.3. Development of Molecular Markers for Flowering Time QTLs
4.4. Genotyping of White Lupin Germplasm with PCR-Based Markers
4.5. Population Structure Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gladstones, J.S. Lupins as crop plants. Field Crop Abstr. 1970, 23, 26. [Google Scholar]
- Lambers, H.; Clements, J.C.; Nelson, M.N. How a phosphorus-acquisition strategy based on carboxylate exudation powers the success and agronomic potential of lupines (Lupinus, Fabaceae). Am. J. Bot. 2013, 100, 263–288. [Google Scholar] [CrossRef]
- Watt, M.; Evans, J.R. Phosphorus acquisition from soil by white lupin (Lupinus albus L.) and soybean (Glycine max L.), species with contrasting root development. Plant Soil 2003, 248, 271–283. [Google Scholar] [CrossRef]
- Schulze, J.; Temple, G.; Temple, S.J.; Beschow, H.; Vance, C.P. Nitrogen fixation by white lupin under phosphorus deficiency. Ann. Bot. 2006, 98, 731–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papineau, J.; Huyghe, C. Le Lupin Doux Protéagineux; France Agricole: Paris, France, 2004; p. 176. [Google Scholar]
- Annicchiarico, P.; Manunza, P.; Arnoldi, A.; Boschin, G. Quality of Lupinus albus L. (white lupin) seed: Extent of genotypic and environmental effects. J. Agric. Food Chem. 2014, 62, 6539–6545. [Google Scholar] [CrossRef]
- Boschin, G.; D’Agostina, A.; Annicchiarico, P.; Arnoldi, A. The fatty acid composition of the oil from Lupinus albus cv. Luxe as affected by environmental and agricultural factors. Eur. Food Res. Technol. 2007, 225, 769–776. [Google Scholar] [CrossRef]
- Arnoldi, A.; Greco, S. Nutritional and nutraceutical characteristics of lupin protein. Nutrafoods 2011, 10, 23–29. [Google Scholar] [CrossRef]
- Kroc, M.; Rybiński, W.; Wilczura, P.; Kamel, K.A.; Kaczmarek, Z.; Barzyk, P.; Święcicki, W. Quantitative and qualitative analysis of alkaloids composition in the seeds of a white lupin (Lupinus albus L.) collection. Genet. Resour. Crop Evol. 2017, 64, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Annicchiarico, P.; Nazzicari, N.; Ferrari, B. Genetic and genomic resources in white lupin and the application of genomic selection. In The Lupin Genome; Singh, K.B., Kamphuis, L.G., Nelson, M.N., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 139–149. [Google Scholar]
- Annicchiarico, P.; Nazzicari, N.; Ferrari, B.; Harzic, N.; Carroni, A.M.; Romani, M.; Pecetti, L. Genomic prediction of grain yield in contrasting environments for white lupin genetic resources. Mol. Breed. 2019, 39, 142. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Romani, M.; Pecetti, L. White lupin (Lupinus albus) variation for adaptation to severe drought stress. Plant Breed. 2018, 137, 782–789. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Harzic, N.; Carroni, A.M. Adaptation, diversity, and exploitation of global white lupin (Lupinus albus L.) landrace genetic resources. Field Crops Res. 2010, 119, 114–124. [Google Scholar] [CrossRef]
- Gladstones, J.S. The Mediterranean white lupin. J. Dep. Agric. West. Aust. Ser. 4 1976, 17, 70–74. [Google Scholar]
- Adhikari, K.N.; Buirchell, B.J.; Sweetingham, M.W. Length of vernalization period affects flowering time in three lupin species. Plant Breed. 2012, 131, 631–636. [Google Scholar] [CrossRef]
- Weimer, J.L. Anthracnose of lupines. Phytopathology 1943, 33, 249–252. [Google Scholar]
- Talhinhas, P.; Baroncelli, R.; Floch, G.L. Anthracnose of lupins caused by Colletotrichum lupini: A recent disease and a successful worldwide pathogen. J. Plant Pathol. 2016, 98, 5–14. [Google Scholar]
- Frencel, I.M. Report on first detection of anthracnose (Colletotrichum gloeosporioides) on lupins in Poland. Plant Dis. 1998, 82, 350. [Google Scholar] [CrossRef]
- Gondran, J.; Bateman, G.L.; Milford, G.F.J.; Bayer, J.; Beerepoot, L.; Boller, B.; Caligari, P.D.S.; Carrasco-Lopez, J.M.; Crowley, J.G.; da Rocha, J.J.P.; et al. Anthracnose of white lupin (Lupinus albus): European prospects for a future sustainable crop. In Towards the 21st Century. Proceedings of the 8th International Lupin Conference; Hill, G.D., Ed.; International Lupin Association: Asilomar, CA, USA, 1996; pp. 512–518. [Google Scholar]
- Thomas, G.J.; Sweetingham, M.W. Cultivar and environment influence the development of lupin anthracnose caused by Colletotrichum lupini. Australas. Plant Path. 2004, 33, 571–577. [Google Scholar] [CrossRef]
- Sweetingham, M.W.; Frencel, I.; Buirchell, B.J.; Barzyk, P.; Lewartowska, E.; Yang, H.A.; Ponomaryova, L.; Yakasheva, A.; Adhikari, K.N.; Francis, C.M.; et al. Collaborative anthracnose resistance screening in Poland, Russia and Australia. México, where old and new world lupins meet. In Proceedings of the 11th International Lupin Conference, Guadalajara, Mexico, 4–9 May 2005; International Lupin Association: Canterbury, New Zealand, 2006. [Google Scholar]
- Adhikari, K.N.; Buirchell, B.J.; Thomas, G.J.; Sweetingham, M.W.; Yang, H. Identification of anthracnose resistance in Lupinus albus L. and its transfer from landraces to modern cultivars. Crop Pasture Sci. 2009, 60, 472–479. [Google Scholar] [CrossRef]
- Atnaf, M.; Yao, N.; Martina, K.; Dagne, K.; Wegary, D.; Tesfaye, K. Molecular genetic diversity and population structure of Ethiopian white lupin landraces: Implications for breeding and conservation. PLoS ONE 2017, 12, e0188696. [Google Scholar] [CrossRef] [Green Version]
- Phan, H.T.T.; Ellwood, S.R.; Adhikari, K.; Nelson, M.N.; Oliver, R.P. The first genetic and comparative map of white lupin (Lupinus albus L.): Identification of QTLs for anthracnose resistance and flowering time, and a locus for alkaloid content. DNA Res. 2007, 14, 59–70. [Google Scholar] [CrossRef]
- Vipin, C.A.; Luckett, D.J.; Harper, J.D.I.; Ash, G.J.; Kilian, A.; Ellwood, S.R.; Phan, H.T.T.; Raman, H. Construction of integrated linkage map of a recombinant inbred line population of white lupin (Lupinus albus L.). Breed. Sci. 2013, 63, 292–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Książkiewicz, M.; Nazzicari, N.; Yang, H.; Nelson, M.N.; Renshaw, D.; Rychel, S.; Ferrari, B.; Carelli, M.; Tomaszewska, M.; Stawiński, S.; et al. A high-density consensus linkage map of white lupin highlights synteny with narrow-leafed lupin and provides markers tagging key agronomic traits. Sci. Rep. 2017, 7, 15335. [Google Scholar] [CrossRef]
- Hufnagel, B.; Marques, A.; Soriano, A.; Marquès, L.; Divol, F.; Doumas, P.; Sallet, E.; Mancinotti, D.; Carrere, S.; Marande, W.; et al. High-quality genome sequence of white lupin provides insight into soil exploration and seed quality. Nat. Commun. 2020, 11, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Zhang, Q.; Yuan, W.; Xu, F.; Muhammad Aslam, M.; Miao, R.; Li, Y.; Wang, Q.; Li, X.; Zhang, X.; et al. The genome evolution and low-phosphorus adaptation in white lupin. Nat. Commun. 2020, 11, 1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychel-Bielska, S.; Nazzicari, N.; Plewiński, P.; Bielski, W.; Annicchiarico, P.; Książkiewicz, M. Development of PCR-based markers and whole-genome selection model for anthracnose resistance in white lupin (Lupinus albus L.). J. Appl. Genet. 2020, 61, 531–545. [Google Scholar] [CrossRef]
- Rychel, S.; Książkiewicz, M.; Tomaszewska, M.; Bielski, W.; Wolko, B. FLOWERING LOCUS T, GIGANTEA, SEPALLATA and FRIGIDA homologs are candidate genes involved in white lupin (Lupinus albus L.) early flowering. Mol. Breed. 2019, 39, 43. [Google Scholar] [CrossRef] [Green Version]
- Rychel, S.; Książkiewicz, M. Development of gene-based molecular markers tagging low alkaloid pauper locus in white lupin (Lupinus albus L.). J. Appl. Genet. 2019, 60, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, K.N.; Thomas, G.; Diepeveen, D.; Trethowan, R. Overcoming the barriers of combining early flowering and anthracnose resistance in white lupin (Lupinus albus L.) for the Northern Agricultural Region of Western Australia. Crop Pasture Sci. 2013, 64, 914–921. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Yang, S.S.; Miller, S.S.; Bucciarelli, B.; Liu, J.; Rydeen, A.; Bozsoki, Z.; Uhde-Stone, C.; Tu, Z.J.; Allan, D.; et al. An RNA-Seq transcriptome analysis of orthophosphate-deficient white lupin reveals novel insights into phosphorus acclimation in plants. Plant Physiol. 2013, 161, 705–724. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.N.; Książkiewicz, M.; Rychel, S.; Besharat, N.; Taylor, C.M.; Wyrwa, K.; Jost, R.; Erskine, W.; Cowling, W.A.; Berger, J.D.; et al. The loss of vernalization requirement in narrow-leafed lupin is associated with a deletion in the promoter and de-repressed expression of a Flowering Locus T (FT) homologue. New Phytol. 2017, 213, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Plewiński, P.; Ćwiek-Kupczyńska, H.; Rudy, E.; Bielski, W.; Rychel-Bielska, S.; Stawiński, S.; Barzyk, P.; Krajewski, P.; Naganowska, B.; Wolko, B.; et al. Innovative transcriptome-based genotyping highlights environmentally responsive genes for phenology, growth and yield in a non-model grain legume. Plant Cell Environ. 2020, 43, 2680–2698. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Q.; Cregan, P.B.; Nelson, R.L.; Wang, X.; Wu, J.; Jiang, G.-L. Genome-wide association study for flowering time, maturity dates and plant height in early maturing soybean (Glycine max) germplasm. BMC Genom. 2015, 16, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, W.W.; Weigel, D. Structural features determining flower-promoting activity of Arabidopsis FLOWERING LOCUS T. Plant Cell 2014, 26, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Kumimoto, R.W.; Gnesutta, N.; Calogero, A.M.; Mantovani, R.; Holt, B.F., 3rd. A distal CCAAT/NUCLEAR FACTOR Y complex promotes chromatin looping at the FLOWERING LOCUS T promoter and regulates the timing of flowering in Arabidopsis. Plant Cell 2014, 26, 1009–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Adrian, J.; Pankin, A.; Hu, J.; Dong, X.; von Korff, M.; Turck, F. Induced and natural variation of promoter length modulates the photoperiodic response of FLOWERING LOCUS T. Nat. Commun. 2014, 5, 4558. [Google Scholar] [CrossRef]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [Green Version]
- Mikołajczyk, J. Genetic studies in Lupinus angustifolius. Part. III. Inheritance of the alkaloid content, seed hardness and length of the growing season in blue lupin. Genet. Pol. 1966, 7, 181–196. [Google Scholar]
- Gladstones, J.; Hill, G. Selection for economic characters in Lupinus angustifolius and L. digitatus. 2. Time of flowering. Aust. J. Exp. Agric. 1969, 9, 213–220. [Google Scholar] [CrossRef]
- Książkiewicz, M.; Rychel, S.; Nelson, M.N.; Wyrwa, K.; Naganowska, B.; Wolko, B. Expansion of the phosphatidylethanolamine binding protein family in legumes: A case study of Lupinus angustifolius L. FLOWERING LOCUS T homologs, LanFTc1 and LanFTc2. BMC Genom. 2016, 17, 820. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.M.; Kamphuis, L.G.; Zhang, W.; Garg, G.; Berger, J.D.; Mousavi-Derazmahalleh, M.; Bayer, P.E.; Edwards, D.; Singh, K.B.; Cowling, W.A.; et al. INDEL variation in the regulatory region of the major flowering time gene LanFTc1 is associated with vernalization response and flowering time in narrow-leafed lupin (Lupinus angustifolius L.). Plant Cell Environ. 2019, 42, 174–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychel-Bielska, S.; Plewiński, P.; Kozak, B.; Galek, R.; Książkiewicz, M. Photoperiod and vernalization control of flowering-related genes: A case study of the narrow-leafed lupin (Lupinus angustifolius L.). Front. Plant Sci. 2020, 11, 572135. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Erskine, W.; Berger, J.D.; Nelson, M.N. Phenotypic characterisation and linkage mapping of domestication syndrome traits in yellow lupin (Lupinus luteus L.). Theor. Appl. Genet. 2020, 133, 2975–2987. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.M.; Huynh, M.; Udall, J.A.; Kilian, A.; Adhikari, K.N.; Berger, J.D.; Erskine, W.; Nelson, M.N. The first genetic map for yellow lupin enables genetic dissection of adaptation traits in an orphan grain legume crop. BMC Genet. 2019, 20, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtin, N.; Salvo-Garrido, H.; Till, B.; Caligari, P.D.S.; Rupayan, A.; Westermeyer, F.; Olivos, M. Genetic and comparative mapping of Lupinus luteus L. highlight syntenic regions with major orthologous genes controlling anthracnose resistance and flowering time. Sci. Rep. 2020, 10, 19174. [Google Scholar] [CrossRef] [PubMed]
- Laurie, R.E.; Diwadkar, P.; Jaudal, M.; Zhang, L.; Hecht, V.; Wen, J.; Tadege, M.; Mysore, K.S.; Putterill, J.; Weller, J.L.; et al. The Medicago FLOWERING LOCUS T homolog, MtFTa1, is a key regulator of flowering time. Plant Physiol. 2011, 156, 2207–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, V.; Laurie, R.E.; Vander Schoor, J.K.; Ridge, S.; Knowles, C.L.; Liew, L.C.; Sussmilch, F.C.; Murfet, I.C.; Macknight, R.C.; Weller, J.L. The pea GIGAS gene is a FLOWERING LOCUS T homolog necessary for graft-transmissible specification of flowering but not for responsiveness to photoperiod. Plant Cell 2011, 23, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Thomson, G.; Taylor, J.; Putterill, J. The transcriptomic response to a short day to long day shift in leaves of the reference legume Medicago truncatula. PeerJ 2019, 7, e6626. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Yamada, T.; Tabata, S.; Kanazawa, A.; Harada, K.; et al. Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, R.; Hayashi, T.; Zhu, J.; Zhao, C.; Xu, M.; Yamaguchi, N.; Sayama, T.; Ishimoto, M.; Kong, L.; Shi, X.; et al. A soybean quantitative trait locus that promotes flowering under long days is identified as FT5a, a FLOWERING LOCUS T ortholog. J. Exp. Bot. 2016, 67, 5247–5258. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Cai, Y.; Qu, M.; Wang, L.; Sun, H.; Jiang, B.; Wu, T.; Liu, L.; Sun, S.; Wu, C.; et al. Soybean adaption to high-latitude regions is associated with natural variations of GmFT2b, an ortholog of FLOWERING LOCUS T. Plant Cell Environ. 2020, 43, 934–944. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Jiang, B.; Ma, L.; Zhang, S.; Zhai, H.; Xu, X.; Hou, W.; Xia, Z.; Wu, C.; Sun, S.; et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation. New Phytol. 2018, 217, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Ortega, R.; Hecht, V.F.G.; Freeman, J.S.; Rubio, J.; Carrasquilla-Garcia, N.; Mir, R.R.; Penmetsa, R.V.; Cook, D.R.; Millan, T.; Weller, J.L. Altered expression of an FT cluster underlies a major locus controlling domestication-related changes to chickpea phenology and growth habit. Front. Plant Sci. 2019, 10, 824. [Google Scholar] [CrossRef] [Green Version]
- Samineni, S.; Kamatam, S.; Thudi, M.; Varshney, R.K.; Gaur, P.M. Vernalization response in chickpea is controlled by a major QTL. Euphytica 2016, 207, 453–461. [Google Scholar] [CrossRef]
- Valverde, F.; Mouradov, A.; Soppe, W.; Ravenscroft, D.; Samach, A.; Coupland, G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 2004, 303, 1003–1006. [Google Scholar] [CrossRef] [Green Version]
- Valverde, F. CONSTANS and the evolutionary origin of photoperiodic timing of flowering. J. Exp. Bot. 2011, 62, 2453–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierre, J.B.; Huguet, T.; Barre, P.; Huyghe, C.; Julier, B. Detection of QTLs for flowering date in three mapping populations of the model legume species Medicago truncatula. Theor. Appl. Genet. 2008, 117, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Pierre, J.-B.; Bogard, M.; Herrmann, D.; Huyghe, C.; Julier, B. A CONSTANS-like gene candidate that could explain most of the genetic variation for flowering date in Medicago truncatula. Mol. Breed. 2011, 28, 25–35. [Google Scholar] [CrossRef]
- Herrmann, D.; Barre, P.; Santoni, S.; Julier, B. Association of a CONSTANS-LIKE gene to flowering and height in autotetraploid alfalfa. Theor. Appl. Genet. 2010, 121, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.L.; Yang, C.H. Late-flowering genes interact with early-flowering genes to regulate flowering time in Arabidopsis thaliana. Plant Cell Physiol. 1999, 40, 702–708. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Bajaj, D.; Das, S.; Saxena, M.S.; Badoni, S.; Kumar, V.; Tripathi, S.; Gowda, C.L.; Sharma, S.; Tyagi, A.K.; et al. A genome-scale integrated approach aids in genetic dissection of complex flowering time trait in chickpea. Plant Mol. Biol. 2015, 89, 403–420. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Michaels, S.D.; Amasino, R.M. Regulation of flowering time by histone acetylation in Arabidopsis. Science 2003, 302, 1751–1754. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of flowering time. Plant Cell 2002, 14, S111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Jin, Y.; Shi, H.; Yang, W. GmFLD, a soybean homolog of the autonomous pathway gene FLOWERING LOCUS D, promotes flowering in Arabidopsis thaliana. BMC Plant Biol. 2014, 14, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johanson, U.; West, J.; Lister, C.; Michaels, S.; Amasino, R.; Dean, C. Molecular analysis of FRIGIDA, a major determinant of natural variation in Arabidopsis flowering time. Science 2000, 290, 344–347. [Google Scholar] [CrossRef]
- Shindo, C.; Aranzana, M.J.; Lister, C.; Baxter, C.; Nicholls, C.; Nordborg, M.; Dean, C. Role of FRIGIDA and FLOWERING LOCUS C in determining variation in flowering time of Arabidopsis. Plant Physiol. 2005, 138, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, D.; He, Y. FRIGIDA establishes a local chromosomal environment for FLOWERING LOCUS C mRNA production. Nat. Plants 2018, 4, 836–846. [Google Scholar] [CrossRef]
- Hecht, V.; Foucher, F.; Ferrandiz, C.; Macknight, R.; Navarro, C.; Morin, J.; Vardy, M.E.; Ellis, N.; Beltran, J.P.; Rameau, C.; et al. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 2005, 137, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Lyu, J.; Cai, Z.; Li, Y.; Suo, H.; Yi, R.; Zhang, S.; Nian, H. The floral repressor GmFLC-like is involved in regulating flowering time mediated by low temperature in soybean. Int. J. Mol. Sci. 2020, 21, 1322. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liu, Y.; Tao, Y.; Xu, C.; Li, X.; Zhang, X.; Han, Y.; Yang, X.; Sun, J.; Li, W.; et al. Identification of genetic loci and candidate genes related to soybean flowering through genome wide association study. BMC Genom. 2019, 20, 987. [Google Scholar] [CrossRef] [PubMed]
- Michaels, S.D.; Bezerra, I.C.; Amasino, R.M. FRIGIDA-related genes are required for the winter-annual habit in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3281–3285. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ransom, C.; Ludwig, P.; van Nocker, S. Genetic analysis of early flowering mutants in Arabidopsis defines a class of pleiotropic developmental regulator required for expression of the flowering-time switch FLOWERING LOCUS C. Genetics 2003, 164, 347–358. [Google Scholar]
- Wang, X.; Wu, F.; Xie, Q.; Wang, H.; Wang, Y.; Yue, Y.; Gahura, O.; Ma, S.; Liu, L.; Cao, Y.; et al. SKIP is a component of the spliceosome linking alternative splicing and the circadian clock in Arabidopsis. Plant Cell 2012, 24, 3278–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, J.; Shang, X.; Lv, W.; Xia, C.; Wang, C.; Feng, J.; Cao, Y.; He, H.; Li, L.; et al. SKIP regulates environmental fitness and floral transition by forming two distinct complexes in Arabidopsis. New Phytol. 2019, 224, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Tong, A.; Huo, Y.; Yan, Z.; Yang, W.; Yang, X.; Wang, X.-X. SKIP controls flowering time via the alternative splicing of SEF pre-mRNA in Arabidopsis. BMC Biol. 2017, 15, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Li, M.; Xu, C.; Yang, X.; Li, D.; Zhao, X.; Wang, K.; Li, Y.; Zhang, X.; Liu, L.; et al. Natural variation in GmGBP1 promoter affects photoperiod control of flowering time and maturity in soybean. Plant J. 2018, 96, 147–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, L.; Li, H.; Gao, Y.; Li, Y.; Wu, X.; Teng, W.; Han, Y.; Zhao, X.; Li, W. GmGBP1, a homolog of human ski interacting protein in soybean, regulates flowering and stress tolerance in Arabidopsis. BMC Plant Biol. 2013, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Brock, M.T.; Maloof, J.N.; Weinig, C. Genes underlying quantitative variation in ecologically important traits: PIF4 (phytochrome interacting factor 4) is associated with variation in internode length, flowering time, and fruit set in Arabidopsis thaliana. Mol. Ecol. 2010, 19, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.; Lucyshyn, D.; Jaeger, K.E.; Alós, E.; Alvey, E.; Harberd, N.P.; Wigge, P.A. Transcription factor PIF4 controls the thermosensory activation of flowering. Nature 2012, 484, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Arya, H.; Singh, M.B.; Bhalla, P.L. Genomic and molecular analysis of conserved and unique features of soybean PIF4. Sci. Rep. 2018, 8, 12569. [Google Scholar] [CrossRef]
- Karlgren, A.; Gyllenstrand, N.; Kallman, T.; Sundstrom, J.F.; Moore, D.; Lascoux, M.; Lagercrantz, U. Evolution of the PEBP gene family in plants: Functional diversification in seed plant evolution. Plant Physiol. 2011, 156, 1967–1977. [Google Scholar] [CrossRef] [Green Version]
- Hedman, H.; Kallman, T.; Lagercrantz, U. Early evolution of the MFT-like gene family in plants. Plant Mol. Biol. 2009, 70, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Y.; Kardailsky, I.; Lee, J.S.; Weigel, D.; Ahn, J.H. Acceleration of flowering by overexpression of MFT (MOTHER OF FT AND TFL1). Mol. Cells 2004, 17, 95–101. [Google Scholar]
- Vaistij, F.E.; Barros-Galvão, T.; Cole, A.F.; Gilday, A.D.; He, Z.; Li, Y.; Harvey, D.; Larson, T.R.; Graham, I.A. MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, 8442–8447. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.K.; Samanta Roy, D.; Chatterjee, I.; Bone, C.R.; Springer, C.J.; Kelly, J.K. Identification of a major QTL that alters flowering time at elevated [CO2] in Arabidopsis thaliana. PLoS ONE 2012, 7, e49028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.; Xing, Y.; Wang, X.; Ma, H. [Variation of CO2 concentration in solar greenhouse in Northern China]. J. Appl. Ecol. 2003, 14, 354–358. [Google Scholar]
- Caicedo, A.L.; Stinchcombe, J.R.; Olsen, K.M.; Schmitt, J.; Purugganan, M.D. Epistatic interaction between Arabidopsis FRI and FLC flowering time genes generates a latitudinal cline in a life history trait. Proc. Natl. Acad. Sci. USA 2004, 101, 15670–15675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunes-Espinoza, L.C.; Huyghe, C.; Papineau, J.; Pacault, D. Effect of genotype and environment on pod wall proportion in white lupin: Consequences to seed yield. Aust. J. Agric. Res. 1999, 50, 575–582. [Google Scholar] [CrossRef]
- Julier, B.; Huyghe, C.; Papineau, J.; Milford, G.F.J.; Day, J.M.; Billot, C.; Mangin, P. Seed yield and yield stability of determinate and indeterminate autumn-sown white lupins (Lupinus albus) grown at different locations in France and the UK. J. Agric. Sci. 1993, 121, 177–186. [Google Scholar] [CrossRef]
- Julier, B.; Huyghe, C. Description and model of the architecture of four genotypes of determinate autumn-sown white lupin (Lupinus albus L.) as influenced by location, sowing date and density. Ann. Bot. 1993, 72, 493–501. [Google Scholar] [CrossRef]
- Alkemade, J.; Messmer, M.; Arncken, C.; Leska, A.; Annicchiarico, P.; Nazzicari, N.; Książkiewicz, M.; Voegele, R.T.; Finckh, M.; Hohmann, P. A high-throughput phenotyping tool to identify field-relevant anthracnose resistance in white lupin. Plant Dis. 2020. [Google Scholar] [CrossRef]
- Park, T.; Casella, G. The Bayesian Lasso. J. Am. Stat. Assoc. 2008, 103, 681–686. [Google Scholar] [CrossRef]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar]
- Searle, S.R.; Casella, G.; McCulloch, C.E. Variance Components; John Wiley & Sons: New York, NY, USA, 2008. [Google Scholar]
- Jiang, B.; Zhang, S.; Song, W.; Khan, M.A.A.; Sun, S.; Zhang, C.; Wu, T.; Wu, C.; Han, T. Natural variations of FT family genes in soybean varieties covering a wide range of maturity groups. BMC Genom. 2019, 20, 230. [Google Scholar] [CrossRef]
- Huyghe, C. Winter growth of autumn-sown white lupin (Lupinus albus L.) main apex growth model. Ann. Bot. 1991, 67, 429–434. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Neff, J.D.; Chory, J.; Pepper, A.E. dCAPS, a simple technique for the genetic analysis of single nucleotide polymorphisms: Experimental applications in Arabidopsis thaliana genetics. Plant J. 1998, 14, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Turk, E.; Kalishman, M. Web-based primer design for single nucleotide polymorphism analysis. Trends Genet. 2002, 18, 613–615. [Google Scholar] [CrossRef]
- Thiel, T.; Kota, R.; Grosse, I.; Stein, N.; Graner, A. SNP2CAPS: A SNP and INDEL analysis tool for CAPS marker development. Nucleic Acids Res. 2004, 32, e5. [Google Scholar] [CrossRef] [PubMed]
- Frichot, E.; François, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Frichot, E.; Mathieu, F.; Trouillon, T.; Bouchard, G.; François, O. Fast and efficient estimation of individual ancestry coefficients. Genetics 2014, 196, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Lange, K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinform. 2011, 12, 246. [Google Scholar] [CrossRef] [Green Version]
- Kendall, M.G. A New Measure of Rank Correlation. Biometrika 1938, 30, 81–93. [Google Scholar] [CrossRef]
- Miller, R.G. Normal Univariate Techniques. In Simultaneous Statistical Inference; Springer: New York, NY, USA, 1981; pp. 37–108. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | 2020 n 1 | 2018 n | 2018 v 2 | 2015 n | 2015 v |
|---|---|---|---|---|---|
| wild or primitive | 67.4 ± 8.6 3 | 54.3 ± 8.2 | 39.4 ± 2.7 | 56.6 ± 8.8 | 46.8 ± 6.2 |
| landrace | 64.7 ± 9.3 | 53.5 ± 11.3 | 38.8 ± 2.8 | 53.6 ± 9.4 | 46.0 ± 4.3 |
| mutant | 74.5 ± 8.6 | 59.5 ± 14.4 | 39.4 ± 1.5 | 65.1 ± 15.5 | 46.8 ± 2.3 |
| cultivar | 56.3 ± 6.2 | 44.5 ± 6.6 | 38.0 ± 5.5 | 49.7 ± 10.4 | 42.7 ± 2.8 |
| cross derivative | 58.5 ± 5.8 | 46.1 ± 5.1 | 38.7 ± 3.2 | 46.4 ± 3.6 | 43.3 ± 2.1 |
| Marker Name | QTL | Primers (5′→3′) | Detection Method | Products Kiev Mutant (bp) | Products P27174 (bp) |
|---|---|---|---|---|---|
| TP56963 | 2 | TGCTCGAAATGCCCAAATCCATCA TTGATGCTCGCAGTGAAGAGATAA | CAPS, HinfI | 878, 384 | 878, 304, 80 |
| TP235608 | 2 | GTAGTCCCAAACATGAACGCAG TCATCTGCATACTTGTCATTCCT | CAPS, AflII | 217 | 179.38 |
| TP94353 | 2 | CAGCATTTATGTTGTTGGGACA GGAACCCTGCAATTTGGATAAGG | CAPS, RsaI | 60, 51 | 111 |
| TP278885 | 2 | CCATTTGAATAGCTGCAAATCGCTTCCG CCTTTGATTGTTGAAGCCTATGC | dCAPS, HpaII | 112 | 86, 26 |
| TP115697 | 2 | TGGCTCCTGTTATGTCACTCA TGAATTTGAGACAAACTCAGTGGTA | dCAPS, RsaI | 111, 24 | 135 |
| TP114357 | 2 | GCCATTCTGGATGGATAACCG TGGACCATCAGCTGACTTCAA | dCAPS, HpaII | 124 | 105, 19 |
| TP100150 | 3 | TATTGCAGCCAATCCATCACTC ACTTTCTTCATCTGATGTTGACGA | CAPS, HpyCh4V | 87, 38, 30, 5 | 117, 38, 5 |
| TP288840 | 3 | CTGCAATATATTCTTTAAGACCTGAT CTGGAGGAATCTAATATAAGTTGTT | dCAPS, MboI | 60 | 37, 23 |
| TP3177 | 3 | CGTGACAAGTGTTCCACGG ATCTGGTTGGAAGCTTGTTGTG | CAPS, SspI | 169 | 114, 55 |
| TP360542 | 3 | GAGCCAGGAATAAGGGTGGTG ACTGGATAGTAAAACCCCATAGAATTACT | dCAPS, TaqI | 113 | 82, 31 |
| TP345457 | 4 | CACAATTCACTACCACAGATCAACC GATTTCGTCCATCCAAGGATTCTTC | CAPS, BseDI | 227, 39, 12 | 143, 84, 39, 12 |
| TP11750 | 4 | AAAACCACTGAAAAGGTTCCACA CAGGCGATAATATACTCGTCCA | CAPS, AciI | 209 | 135, 74 |
| TP402859 | 4 | CTGGTGGCAAAAGAAGCAGAA AAAGCCAGGAAAGCACATTGG | CAPS, HpaII | 198 | 112, 86 |
| TP2488 | 5 | ACCTTGTTATTGATGCTAGCTTCT TGTTTGAGGGAAGGCAGGTGGAAT | dCAPS, TspEI | 48, 25, 21 | 48, 46 |
| TP86766 | 5 | CAGCATGCAAGAAAGCTG TCCTTTCTTCTCCTTCTCTTTC | CAPS, DdeI | 64 | 48, 16 |
| TP47264 | 5 | TAACATGCAGCACTCACCAAC TCTGGTTTCTGGGTAATGAGGA | CAPS, MboI | 171 | 105, 66 |
| TP30473 | 5 | CAGCACACAACCGCAATAAC ATAATTACAGGAAAATATGGTCTTG | CAPS, HpyCH4V | 28.25 | 53 |
| Cluster | Marker | Linkage Group | Position (cM) | QTL | 2015 p-Value 1 | 2018 p-Value | 2020 p-Value |
|---|---|---|---|---|---|---|---|
| V2 | TP56963 2 | ALB02 | 96.4 | 2 | 0.048 3 | 0.021 | 0.017 |
| V2 | TP235608 | ALB02 | 96.8 | 2 | 0.111 | 0.048 | 0.018 |
| V2 | PIF4-F6 | ALB04 | 7.3 | - | 0.073 | 0.044 | 0.017 |
| V4 | TP86766 | ALB16 | 2.2 | 5 | 0.046 | 2.8 × 10−4 | 4.0 × 10−5 |
| V4 | TP30473 | ALB16 | 6.7 | 5 | 0.044 | 0.019 | 0.003 |
| V5 | FTc1-F4 | ALB14 | 53.9 | - | 0.051 | 0.016 | 0.008 |
| V5 | TP86766 | ALB16 | 2.2 | 5 | 0.034 | 0.020 | 0.003 |
| V6 | MFTa1-F1 | ALB05 | 68.1 | - | 0.039 | 0.032 | 0.023 |
| V6 | CO-F1 | ALB10 | 15.1 | - | 0.027 | 0.009 | 0.012 |
| V6 | FRI31-F1 | ALB16 | 5.8 | 5 | 0.003 | 0.087 | 0.016 |
| V6 | TP2488 | ALB16 | 0.3 | 5 | 0.062 | 0.029 | 0.039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rychel-Bielska, S.; Surma, A.; Bielski, W.; Kozak, B.; Galek, R.; Książkiewicz, M. Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.). Int. J. Mol. Sci. 2021, 22, 3856. https://doi.org/10.3390/ijms22083856
Rychel-Bielska S, Surma A, Bielski W, Kozak B, Galek R, Książkiewicz M. Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.). International Journal of Molecular Sciences. 2021; 22(8):3856. https://doi.org/10.3390/ijms22083856
Chicago/Turabian StyleRychel-Bielska, Sandra, Anna Surma, Wojciech Bielski, Bartosz Kozak, Renata Galek, and Michał Książkiewicz. 2021. "Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.)" International Journal of Molecular Sciences 22, no. 8: 3856. https://doi.org/10.3390/ijms22083856
APA StyleRychel-Bielska, S., Surma, A., Bielski, W., Kozak, B., Galek, R., & Książkiewicz, M. (2021). Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.). International Journal of Molecular Sciences, 22(8), 3856. https://doi.org/10.3390/ijms22083856