
Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency

 ,
,  ,
,  ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Pappa2 Deletion Reduces Body and Bone Length
2.2. Pappa2 Deletion Alters Hydroxyapatite Crystallinity in the Femur of Male Mice, But Not Female Mice
2.3. Pappa2 Deletion Alters Bone Composition in the Femur of Male Mice, but Not Female Mice
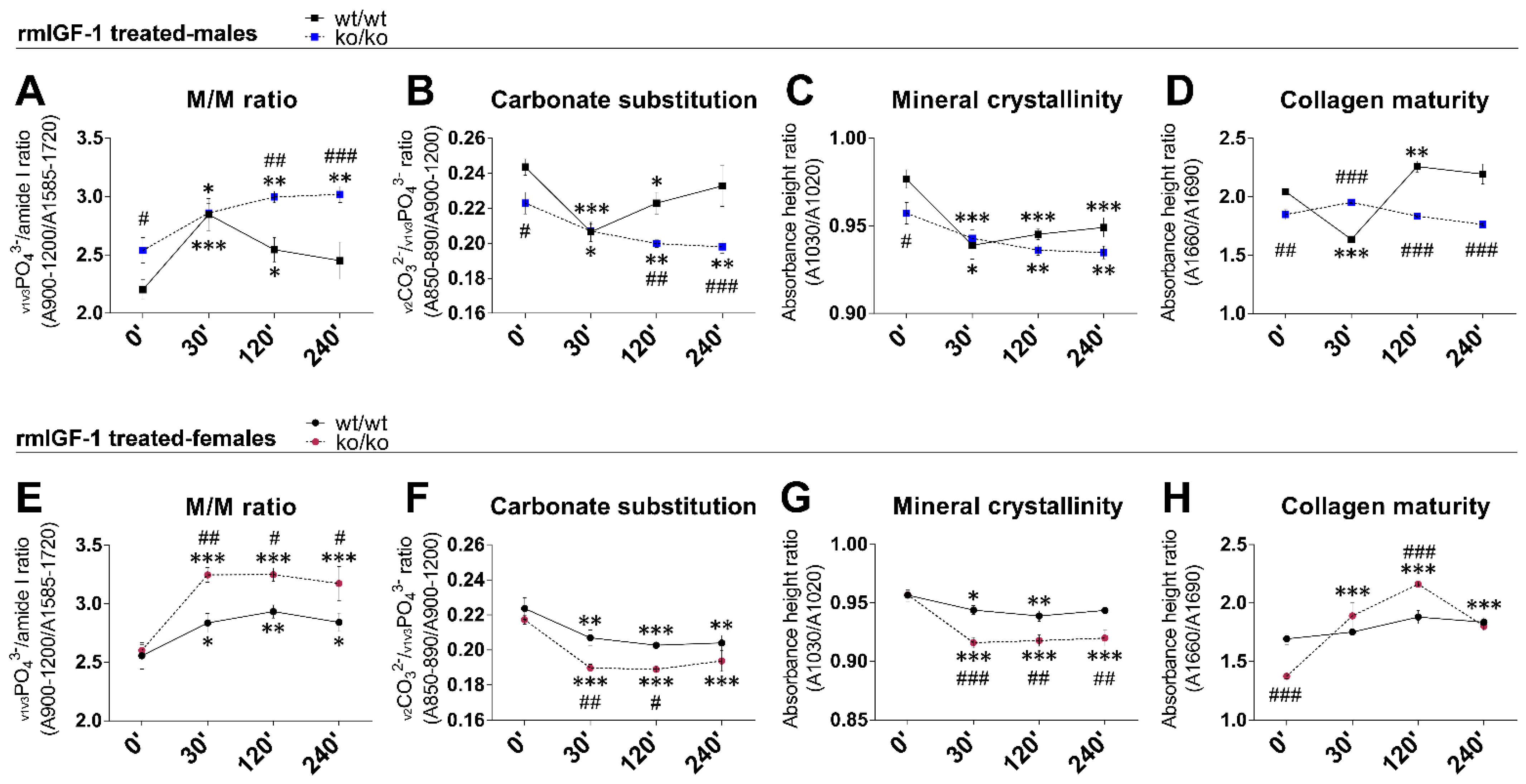
2.4. rmIGF-1 Induces Sex-Specific Changes in Bone Composition of Pappa2ko/ko Mice
2.5. Pappa2 Deletion Affects Igfbp3 Gene Expression in the Tibia
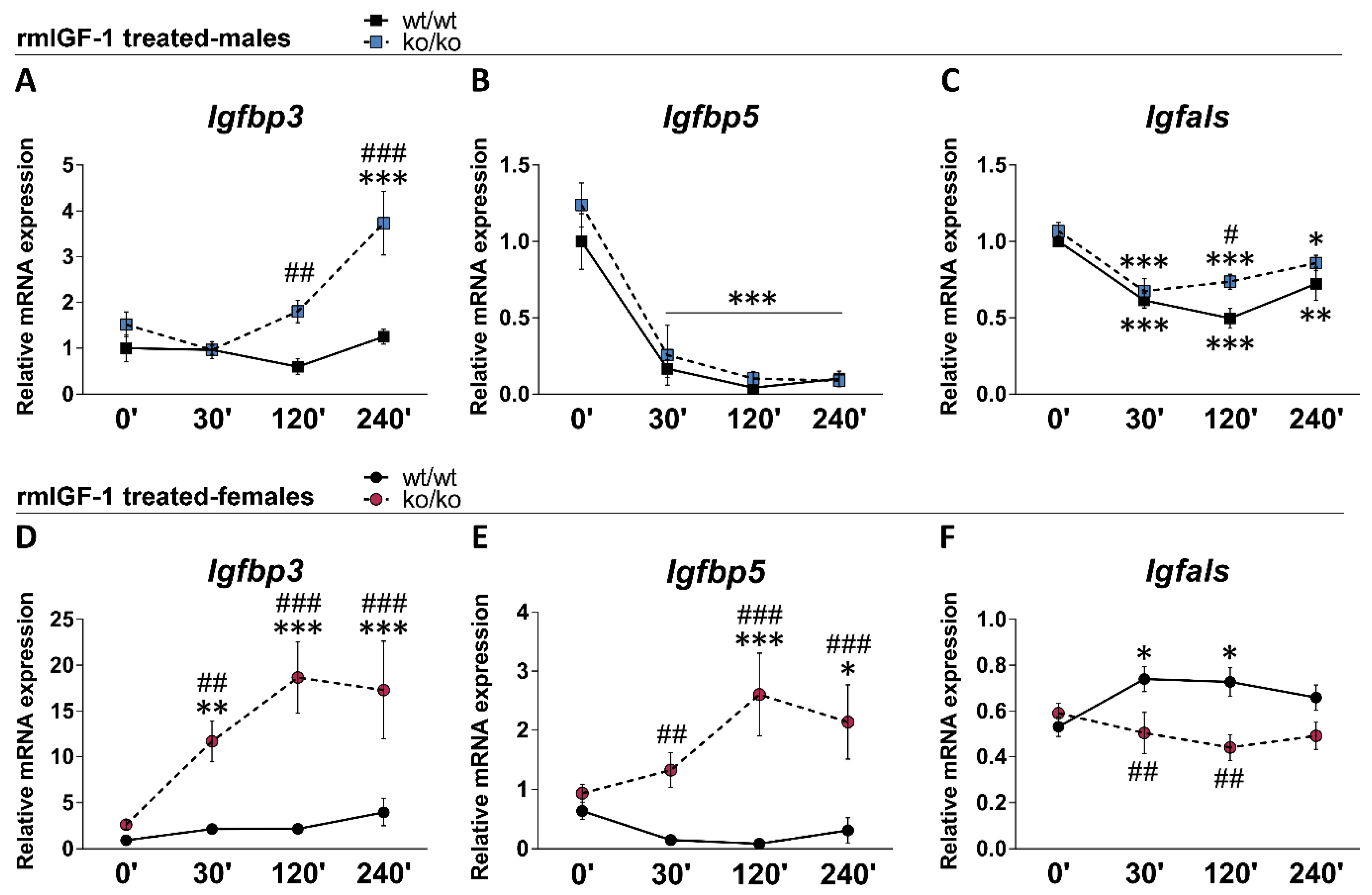
2.6. rmIGF-1 Induces Sex-Specific Changes in Local IGF-1 System of Pappa2ko/ko Mice
2.7. Pappa2 Deletion Affects the Gene Expression of Bone Remodeling Markers in the Tibia
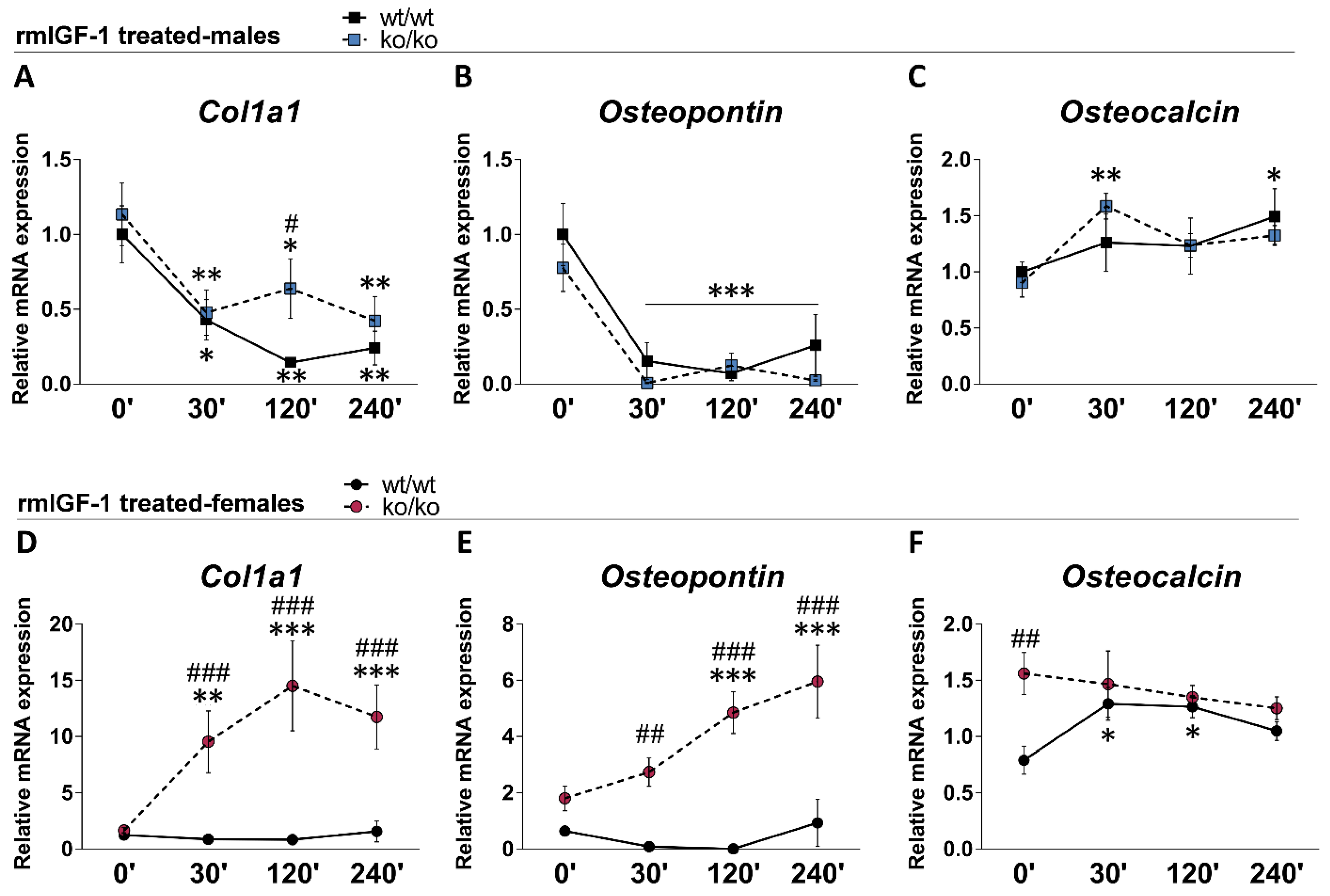
2.8. rmIGF-1 Induces Sex-Specific Changes in Bone Remodeling Markers of Pappa2ko/ko Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Sample Collection
4.4. Sample Preparation
4.5. X-ray Powder Diffraction
4.6. ATR-FTIR Spectroscopy
4.7. RNA Isolation and RT-qPCR Analysis
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohan, S.; Richman, C.; Guo, R.; Amaar, Y.; Donahue, L.R.; Wergedal, J.; Baylink, D.J. Insulin-like growth factor regulates peak bone mineral density in mice by both growth hormone-dependent and -independent mechanisms. Endocrinology 2003, 144, 929–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govoni, K.E.; Baylink, D.J.; Mohan, S. The multi-functional role of insulin-like growth factor binding proteins in bone. Pediatr. Nephrol. 2005, 20, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Baylink, D.; Lau, K.H.; Mohan, S. The role of IGF system in the rise and fall in bone density with age. J. Musculoskelet Neuronal Interact. 2007, 7, 304–305. [Google Scholar] [PubMed]
- Tritos, N.A.; Klibanski, A. Effects of Growth Hormone on Bone. Prog. Mol. Biol. Transl. Sci. 2016, 138, 193–211. [Google Scholar] [CrossRef]
- Yakar, S.; Isaksson, O. Regulation of skeletal growth and mineral acquisition by the GH/IGF-1 axis: Lessons from mouse models. Growth Horm IGF Res. 2016, 28, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Rogol, A.D. Sex steroids, growth hormone, leptin and the pubertal growth spurt. Endocr. Dev. 2010, 17, 77–85. [Google Scholar] [CrossRef]
- Liu, Z.; Mohan, S.; Yakar, S. Does the GH/IGF-1 axis contribute to skeletal sexual dimorphism? Evidence from mouse studies. Growth Horm IGF Res. 2016, 27, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Southmayd, E.A.; De Souza, M.J. A summary of the influence of exogenous estrogen administration across the lifespan on the GH/IGF-1 axis and implications for bone health. Growth Horm IGF Res. 2017, 32, 2–13. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Powell-Braxton, L.; Hollingshead, P.; Warburton, C.; Dowd, M.; Pitts-Meek, S.; Dalton, D.; Gillett, N.; Stewart, T.A. IGF-I is required for normal embryonic growth in mice. Genes Dev. 1993, 7, 2609–2617. [Google Scholar] [CrossRef]
- He, J.; Rosen, C.J.; Adams, D.J.; Kream, B.E. Postnatal growth and bone mass in mice with IGF-I haploinsufficiency. Bone 2006, 38, 826–835. [Google Scholar] [CrossRef]
- Wang, Y.; Nishida, S.; Sakata, T.; Elalieh, H.Z.; Chang, W.; Halloran, B.P.; Doty, S.B.; Bikle, D.D. Insulin-like growth factor-I is essential for embryonic bone development. Endocrinology 2006, 147, 4753–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, K.A.; Camacho-Hübner, C.; Savage, M.O.; Clark, A.J. Intrauterine growth retardation and postnatal growth failure associated with deletion of the insulin-like growth factor I gene. N. Engl. J. Med. 1996, 335, 1363–1367. [Google Scholar] [CrossRef] [PubMed]
- Abuzzahab, M.J.; Schneider, A.; Goddard, A.; Grigorescu, F.; Lautier, C.; Keller, E.; Kiess, W.; Klammt, J.; Kratzsch, J.; Osgood, D.; et al. Intrauterine Growth Retardation (IUGR) Study Group. IGF-I receptor mutations resulting in intrauterine and postnatal growth retardation. N. Engl. J. Med. 2003, 349, 2211–2222. [Google Scholar] [CrossRef] [PubMed]
- Miyakoshi, N.; Richman, C.; Kasukawa, Y.; Linkhart, T.A.; Baylink, D.J.; Mohan, S. Evidence that IGF-binding protein-5 functions as a growth factor. J. Clin. Investig. 2001, 107, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overgaard, M.T.; Boldt, H.B.; Laursen, L.S.; Sottrup-Jensen, L.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A2 (PAPP-A2), a novel insulin-like growth factor-binding protein-5 proteinase. J. Biol. Chem. 2001, 276, 21849–21853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, S.; Baylink, D.J. IGF-binding proteins are multifunctional and act via IGF-dependent and -independent mechanisms. J. Endocrinol. 2002, 175, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunn, R.C.; Fowlkes, J.L. Insulin-like growth factor binding protein proteolysis. Trends Endocrinol. Metab. 2003, 14, 176–181. [Google Scholar] [CrossRef]
- Salih, D.A.; Mohan, S.; Kasukawa, Y.; Tripathi, G.; Lovett, F.A.; Anderson, N.F.; Carter, E.J.; Wergedal, J.E.; Baylink, D.J.; Pell, J.M. Insulin-like growth factor-binding protein-5 induces a gender-related decrease in bone mineral density in transgenic mice. Endocrinology 2005, 146, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Ranke, M.B. Insulin-like growth factor binding-protein-3 (IGFBP-3). Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Rosen, C.J.; Beamer, W.G.; Ackert-Bicknell, C.L.; Wu, Y.; Liu, J.L.; Ooi, G.T.; Setser, J.; Frystyk, J.; Boisclair, Y.R.; et al. Circulating levels of IGF-1 directly regulate bone growth and density. J. Clin. Investig. 2002, 110, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Wu, Y.; Setser, J.; Rosen, C.J. The role of circulating IGF-I: Lessons from human and animal models. Endocrine 2002, 19, 239–248. [Google Scholar] [CrossRef]
- Dauber, A.; Muñoz-Calvo, M.T.; Barrios, V.; Domené, H.M.; Kloverpris, S.; Serra-Juhé, C.; Desikan, V.; Pozo, J.; Muzumdar, R.; Martos-Moreno, G.Á.; et al. Mutations in pregnancy-associated plasma protein A2 cause short stature due to low IGF-I availability. EMBO Mol. Med. 2016, 8, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Argente, J.; Pérez-Jurado, L.A. Genetic causes of proportionate short stature. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 499–522. [Google Scholar] [CrossRef]
- Argente, J.; Pérez-Jurado, L.A. History and clinical implications of PAPP-A2 in human growth: When reflecting on idiopathic short stature leads to a specific and new diagnosis: Understanding the concept of “low IGF-I availability”. Growth Horm. IGF Res. 2018, 40, 19. [Google Scholar] [CrossRef] [PubMed]
- Ranke, M.B.; Wit, J.M. Growth hormone—past, present and future. Nat. Rev. Endocrinol. 2018, 14, 285–300. [Google Scholar] [CrossRef]
- Muñoz-Calvo, M.T.; Barrios, V.; Pozo, J.; Chowen, J.A.; Martos-Moreno, G.Á.; Hawkins, F.; Dauber, A.; Domené, H.M.; Yakar, S.; Rosenfeld, R.G.; et al. Treatment with recombinant human insulin-like growth factor-1 improves growth in patients with PAPP-A2 deficiency. J. Clin. Endocrinol. Metab. 2016, 101, 3879–3883. [Google Scholar] [CrossRef] [Green Version]
- Hawkins-Carranza, F.G.; Muñoz-Calvo, M.T.; Martos-Moreno, G.Á.; Allo-Miguel, G.; Del Río, L.; Pozo, J.; Chowen, J.A.; Pérez-Jurado, L.A.; Argente, J. rhIGF-1 treatment increases bone mineral density and trabecular bone structure in children with PAPP-A2 deficiency. Horm Res. Paediatr. 2018, 89, 200–204. [Google Scholar] [CrossRef]
- Conover, C.A.; Boldt, H.B.; Bale, L.K.; Clifton, K.B.; Grell, J.A.; Mader, J.R.; Mason, E.J.; Powell, D.R. Pregnancy-associated plasma protein-A2 (PAPP-A2): Tissue expression and biological consequences of gene knockout in mice. Endocrinology 2011, 152, 2837–2844. [Google Scholar] [CrossRef] [Green Version]
- Christians, J.K.; de Zwaan, D.R.; Fung, S.H. Pregnancy associated plasma protein A2 (PAPP-A2) affects bone size and shape and contributes to natural variation in postnatal growth in mice. PLoS ONE 2013, 8, e56260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, N.; Christians, J.K. PAPP-A2 expression by osteoblasts is required for normal postnatal growth in mice. Growth Horm IGF Res. 2015, 25, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Christians, J.K.; Amiri, N.; Schipilow, J.D.; Zhang, S.W.; May-Rashke, K.I. Pappa2 deletion has sex- and age-specific effects on bone in mice. Growth Horm. IGF Res. 2019, 44, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Christians, J.K.; Bath, A.K.; Amiri, N. Pappa2 deletion alters IGFBPs but has little effect on glucose disposal or adiposity. Growth Horm. IGF Res. 2015, 25, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Sandstedt, J.; Törnell, J.; Norjavaara, E.; Isaksson, O.G.; Ohlsson, C. Elevated levels of growth hormone increase bone mineral content in normal young mice, but not in ovariectomized mice. Endocrinology 1996, 137, 3368–3374. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, F.; Lochmüller, E.M.; Koller, B.; Wehr, U.; Weusten, A.; Rambeck, W.; Hoeflich, A.; Wolf, E. Body composition, bone mass and microstructural analysis in GH-transgenic mice reveals that skeletal changes are specific to bone compartment and gender. Growth Horm. IGF Res. 2002, 12, 116–125. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, H.; Basta-Pljakic, J.; Cardoso, L.; Kennedy, O.D.; Jasper, H.; Domené, H.; Karabatas, L.; Guida, C.; Schaffler, M.B.; et al. Serum IGF-1 is insufficient to restore skeletal size in the total absence of the growth hormone receptor. J. Bone Miner. Res. 2013, 28, 1575–1586. [Google Scholar] [CrossRef]
- Courtland, H.W.; DeMambro, V.; Maynard, J.; Sun, H.; Elis, S.; Rosen, C.; Yakar, S. Sex-specific regulation of body size and bone slenderness by the acid labile subunit. J. Bone Miner. Res. 2010, 25, 2059–2068. [Google Scholar] [CrossRef]
- DeMambro, V.E.; Clemmons, D.R.; Horton, L.G.; Bouxsein, M.L.; Wood, T.L.; Beamer, W.G.; Canalis, E.; Rosen, C.J. Gender-specific changes in bone turnover and skeletal architecture in igfbp-2-null mice. Endocrinology 2008, 149, 2051–2061. [Google Scholar] [CrossRef]
- Maridas, D.E.; DeMambro, V.E.; Le, P.T.; Nagano, K.; Baron, R.; Mohan, S.; Rosen, C.J. IGFBP-4 regulates adult skeletal growth in a sex-specific manner. J. Endocrinol. 2017, 233, 131–144. [Google Scholar] [CrossRef]
- Tanner, S.J.; Hefferan, T.E.; Rosen, C.J.; Conover, C.A. Impact of pregnancy-associated plasma protein-a deletion on the adult murine skeleton. J. Bone Miner. Res. 2008, 23, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Olson, L.E.; Ohlsson, C.; Mohan, S. The role of GH/IGF-I-mediated mechanisms in sex differences in cortical bone size in mice. Calcif. Tissue Int. 2011, 88, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeman, E. During aging, men lose less bone than women because they gain more periosteal bone, not because they resorb less endosteal bone. Calcif. Tissue Int. 2001, 69, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.L.; Khosla, S.; Melton, L.J., 3rd. Sex steroids and the construction and conservation of the adult skeleton. Endocr. Rev. 2002, 23, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Christoforidis, A.; Maniadaki, I.; Stanhope, R. Growth hormone/insulin-like growth factor-1 axis during puberty. Pediatr. Endocrinol. Rev. 2005, 3, 5–10. [Google Scholar] [PubMed]
- Veldhuis, J.D.; Roemmich, J.N.; Richmond, E.J.; Rogol, A.D.; Lovejoy, J.C.; Sheffield-Moore, M.; Mauras, N.; Bowers, C.Y. Endocrine control of body composition in infancy, childhood, and puberty. Endocr. Rev. 2005, 26, 114–146. [Google Scholar] [CrossRef] [PubMed]
- Seeman, E. Estrogen, androgen, and the pathogenesis of bone fragility in women and men. Curr. Osteoporos Rep. 2004, 2, 90–96. [Google Scholar] [CrossRef]
- Bauss, F.; Lang, K.; Dony, C.; Kling, L. The complex of recombinant human insulin-like growth factor-I (rhIGF-I) and its binding protein-5 (IGFBP-5) induces local bone formation in murine calvariae and in rat cortical bone after local or systemic administration. Growth Horm. IGF Res. 2001, 11, 1–9. [Google Scholar] [CrossRef]
- Ning, Y.; Schuller, A.G.; Bradshaw, S.; Rotwein, P.; Ludwig, T.; Frystyk, J.; Pintar, J.E. Diminished growth and enhanced glucose metabolism in triple knockout mice containing mutations of insulin-like growth factor binding protein-3, -4, and -5. Mol. Endocrinol. 2006, 20, 2173–2186. [Google Scholar] [CrossRef]
- Mukherjee, A.; Rotwein, P. Insulin-like growth factor binding protein-5 in osteogenesis: Facilitator or inhibitor? Growth Horm. IGF Res. 2007, 17, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Rotwein, P. Insulin-like growth factor-binding protein-5 inhibits osteoblast differentiation and skeletal growth by blocking insulin-like growth factor actions. Mol. Endocrinol. 2008, 22, 1238–1250. [Google Scholar] [CrossRef] [Green Version]
- Allard, J.B.; Duan, C. IGF-Binding Proteins: Why Do They Exist and Why Are There So Many? Front. Endocrinol. 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Menon, R.K.; Cohen, P.; Hwang, D.; Clemens, T.; DiGirolamo, D.J.; Kopchick, J.J.; Le Roith, D.; Trucco, M.; Sperling, M.A. Liver-specific deletion of the growth hormone receptor reveals essential role of growth hormone signaling in hepatic lipid metabolism. J. Biol. Chem. 2009, 284, 19937–19944. [Google Scholar] [CrossRef] [Green Version]
- Szulc, P. Bone turnover: Biology and assessment tools. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Ivaska, K.K.; Lenora, J.; Gerdhem, P.; Akesson, K.; Väänänen, H.K.; Obrant, K.J. Serial assessment of serum bone metabolism markers identifies women with the highest rate of bone loss and osteoporosis risk. J. Clin. Endocrinol. Metab. 2008, 93, 2622–2632. [Google Scholar] [CrossRef] [PubMed]
- Szulc, P.; Bauer, D.C. Biochemical markers of bone turnover in osteoporosis. In Osteoporosis, 4th ed.; Marcus, R., Feldman, D., Dempster, D.W., Luckey, M., Cauley, J.A., Eds.; Academic Press Elsevier: Amsterdam, The Netherlands, 2013; pp. 1573–1610. [Google Scholar]
- Boskey, A.L.; Imbert, L. Bone quality changes associated with aging and disease: A review. Ann. N. Y. Acad. Sci. 2017, 1410, 93–106. [Google Scholar] [CrossRef]
- Elliott, J.C. Calcium Phosphate Biominerals. In Phosphates: Geochemical, Geobiological and Material Importance, Reviews in Mineralogy and Geochemistry; Kohn, M.J., Rakovan, J., Hughes., J.M., Eds.; Mineralogical Society of America: Washington, DC, USA, 2002; Volume 48, pp. 427–454. [Google Scholar]
- Hughes, J.M.; Rakovan, J. The Crystal Structure of Apatite, Ca5(PO4)3(F,OH,Cl). In Phosphates: Geochemical, Geobiological and Material Importance, Reviews in Mineralogy and Geochemistry; Kohn, M.J., Rakovan, J., Hughes, J.M., Eds.; Mineralogical Society of America: Washington, DC, USA, 2002; Volume 48, pp. 1–12. [Google Scholar]
- Astala, R.; Stott, M.J. First principles investigation of mineral component of bone: CO3 substitutions in hydroxyapatite. Chem. Mater. 2005, 17, 4125–4133. [Google Scholar] [CrossRef]
- Pasteris, J.D.; Wopenka, B.; Freeman, J.J.; Rogers, K.; Valsami-Jones, E.; van der Houwen, J.A.; Silva, M.J. Lack of OH in nanocrystalline apatite as a function of degree of atomic order: Implications for bone and biomaterials. Biomaterials 2004, 25, 229–238. [Google Scholar] [CrossRef]
- Nakamura, M.; Hiratai, R.; Hentunen, T.; Salonen, J.; Yamashita, K. Hydroxyapatite with high carbonate substitutions promotes osteoclast resorption through osteocytelike cells. ACS Biomater. Sci. Eng. 2016, 2, 259–267. [Google Scholar] [CrossRef]
- Vallet-Regi, M.; Navarette, D.A. Nanoceramics in Clinical Use: From Materials to Applications, 2nd ed.; Royal Society of Chemistry: Cambridge, MA, USA, 2015. [Google Scholar]
- de Campos Vidal, B.; Mello, M.L.S. Collagen type I amide I band infrared spectroscopy. Micron 2011, 42, 283–289. [Google Scholar] [CrossRef]
- Awonusi, A.; Morris, M.D.; Tecklenburg, M.M. Carbonate assignment and calibration in the Raman spectrum of apatite. Calcif. Tissue Int. 2007, 81, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Kamitakahara, M.; Ohtsuki, C.; Miyazaki, T. Coating of bone-like apatite for development of bioactive materials for bone reconstruction. Biomed. Mater. 2007, 2, R17. [Google Scholar] [CrossRef] [PubMed]
- Amenta, E.; King, H.E.; Petermann, H.; Uskoković, V.; Tommasini, S.M.; Macica, C.M. Vibrational spectroscopic analysis of hydroxyapatite in HYP mice and individuals with X-linked hypophosphatemia. Ther. Adv. Chronic. Dis. 2018, 9, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Gourion-Arsiquaud, S.; Faibish, D.; Myers, E.; Spevak, L.; Compston, J.; Hodsman, A.; Shane, E.; Recker, R.R.; Boskey, E.R.; Boskey, A.L. Use of FTIR spectroscopic imaging to identify parameters associated with fragility fracture. J. Bone Miner. Res. 2009, 24, 1565–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrew, M.; Liao, L.; Fujimoto, M.; Khoury, J.; Hwa, V.; Dauber, A. PAPPA2 as a Therapeutic Modulator of IGF-I Bioavailability: In Vivo and in Vitro Evidence. J. Endocr. Soc. 2018, 2, 646–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narusawa, K.; Nakamura, T.; Suzuki, K.; Matsuoka, Y.; Lee, L.J.; Tanaka, H.; Seino, Y. The effects of recombinant human insulin-like growth factor (rhIGF)-1 and rhIGF-1/IGF binding protein-3 administration on rat osteopenia induced by ovariectomy with concomitant bilateral sciatic neurectomy. J. Bone Miner. Res. 1995, 10, 1853–1864. [Google Scholar] [CrossRef]
- Seeman, E. Overview of bone microstructure, and treatment of bone fragility in chronic kidney disease. Nephrology 2017, 22 (Suppl. S2), 34–35. [Google Scholar] [CrossRef] [Green Version]
- Fowlkes, J.L.; Thrailkill, K.M.; Liu, L.; Wahl, E.C.; Bunn, R.C.; Cockrell, G.E.; Perrien, D.S.; Aronson, J.; Lumpkin, C.K., Jr. Effects of systemic and local administration of recombinant human IGF-I (rhIGF-I) on de novo bone formation in an aged mouse model. J. Bone Miner. Res. 2006, 21, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, M.; Fernando, A.; Martins, G.; Freitas, J.; Judas, F.; Figueiredo, H. Effect of the calcination temperature on the composition and microstructure of hydroxyapatite derived from human and animal bone. Ceram. Int. 2010, 36, 2383–2393. [Google Scholar] [CrossRef]
- Grynpas, M. The crystallinity of bone mineral. J. Miner. Sci. 1976, 11, 1691–1696. [Google Scholar] [CrossRef]
- Farlay, F.D.; Panczer, G.; Rey, C.; Delmas, P.; Boivin, G. Mineral maturity and crystallinity index are distinct characteristics of bone mineral. J. Bone Miner. Metab. 2010, 28, 433–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa, Y.; Guo, Y.; Feng, X.; Wang, M.; Li, P.; Gao, Y.; Yang, X.; Jiang, T. Are different crystallinity-index-calculating methods of hydroxyapatite efficient and consistent? New J. Chem. 2017, 41, 5723–5731. [Google Scholar] [CrossRef]
- Surovell, T.A.; Stiner, M.C. Standardizing infra-red measures of bone mineral crystallinity: An experimental approach. J. Arch. Sci. 2000, 28, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Dal Sasso, G.; Asscher, Y.; Angelini, I.; Nodari, L.; Artioli, G. A universal curve of apatite crystallinity for the assessment of bone integrity and preservation. Sci. Rep. 2018, 8, 12025. [Google Scholar] [CrossRef] [PubMed]
- Querido, W.; Ailavajhala, R.; Padalkar, M.; Pleshko, N. Validated Approaches for Quantification of Bone Mineral Crystallinity Using Transmission Fourier Transform Infrared (FT-IR), Attenuated Total Reflection (ATR) FT-IR, and Raman Spectroscopy. Appl. Spectrosc. 2018, 72, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Paschalis, E.P.; Mendelsohn, R.; Boskey, A.L. Infrared assessment of bone quality: A review. Clin. Orthop. Relat. Res. 2011, 469, 2170–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschalis, E.P. Fourier Transform Infrared Imaging of Bone. Methods Mol. Biol. 2019, 1914, 641–649. [Google Scholar]
- Taylor, E.A.; Lloyd, A.A.; Salazar-Lara, C.; Donnelly, E. Raman and Fourier Transform Infrared (FT-IR) Mineral to Matrix Ratios Correlate with Physical Chemical Properties of Model Compounds and Native Bone Tissue. Appl. Spectrosc. 2017, 71, 2404–2410. [Google Scholar] [CrossRef]
- Rivera, P.; Ramírez-López, M.T.; Vargas, A.; Decara, J.; Vázquez, M.; Arco, R.; Gómez de Heras, R.; Argente, J.; Rodríguez de Fonseca, F.; Chowen, J.A.; et al. Perinatal free-choice of a high-calorie low-protein diet affects leptin signaling through IRS1 and AMPK dephosphorylation in the hypothalami of female rat offspring in adulthood. Acta Physiol. 2019, 226, e13244. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| wt/wt Male | ko/ko Male | wt/wt Female | ko/ko Female | Interaction (Genotype vs. Sex) | Genotype | Sex | |
|---|---|---|---|---|---|---|---|
| Body length (cm) | 16.5 ± 0.12 | 15.8 ± 0.08 ### | 16.3 ± 0.07 | 15.2 ± 0.16 ***/&& | ns | F1.91 = 70.78 p < 0.0001 | F1.91 = 13.04 p = 0.0005 |
| Femur length (cm) | 1.61 ± 0.04 | 1.25 ± 0.03 ### | 1.41 ±0.02 ### | 1.2 ± 0.002 *** | F1.24 = 5.46 p = 0.028 | F1.24 = 87.42 p < 0.0001 | F1.24 = 17.70 p = 0.0003 |
| Tibia length (cm) | 1.07 ± 0.03 | 0.87 ± 0.01 ### | 1.08 ± 0.03 | 0.88 ± 0.02 *** | ns | F1.24 = 45.82 p < 0.0001 | ns |
| Femur weight (g) | 0.32 ± 0.02 | 0.21 ± 0.004 ### | 0.21 ± 0.01 ### | 0.18 ± 0.004 | F1.24 = 6.93 p = 0.014 | F1.24 = 25.01 p < 0.0001 | F1.24 = 20.05 p = 0.0002 |
| Tibia weight (g) | 0.21 ± 0.01 | 0.11 ± 0.008 ### | 0.14 ± 0.007 ### | 0.08 ± 0.004 *** | F1.24 = 7.07 p = 0.013 | F1.24 = 93.98 p < 0.0001 | F1.24 = 37.49 p < 0.0001 |
| Femur weight/length ratio (g/cm) | 0.2 ± 0.01 | 0.16 ± 0.006 # | 0.15 ± 0.008 ### | 0.15 ± 0.003 | ns | ns | F1.24 = 10.79 p = 0.0031 |
| Tibia weight/length ratio (g/cm) | 0.2 ± 0.015 | 0.13 ± 0.005 ### | 0.13 ± 0.008 ### | 0.1 ± 0.002 * | ns | F1.24 = 31.50 p < 0.0001 | F1.24 = 31.04 p = 0.0031 |
| Femur/body weight ratio (mg/g) | 10.12 ± 1.22 | 7.09 ± 0.18 # | 8.39 ± 0.81 | 9.22 ± 0.33 | F1.24 = 6.43 p = 0.018 | ns | ns |
| Tibia/body weight ratio (mg/g) | 6.62 ± 0.61 | 3.95 ± 0.25 ### | 5.54 ± 0.36 | 4.53 ± 0.31 | ns | F1.24 = 20.22 p = 0.0001 | ns |
| wt/wt Male | ko/ko Male | wt/wt Female | ko/ko Female | Interaction (Genotype vs. Sex) | Genotype | Sex | |
|---|---|---|---|---|---|---|---|
| Sample Crystallinity (%) | 45.40 ± 3.87 | 59.57 ± 2.02 ## | 60.17 ± 2.77 ## | 57.85 ± 2.07 | F1.24 = 8.73 p = 0.0069 | F1.24 = 4.51 p = 0.044 | F1.24 = 5.47 p = 0.027 |
| Crystallite Size (LVol-IB, nm) | 36.25 ± 5.75 | 24.86 ± 1.64 # | 27.36 ± 3.20 | 24.61 ± 2.41 | ns | F1.24 = 3.95 p = 0.05 | ns |
| wt/wt Male | ko/ko Male | wt/wt Female | ko/ko Female | Interaction (Genotype vs. Sex) | Genotype | Sex | |
|---|---|---|---|---|---|---|---|
| Mineral-to-matrix ratio 2 | 2.20 ± 0.08 | 2.53 ± 0.01 # | 2.55 ± 0.11 # | 2.60 ± 0.04 | ns | F1.24 = 4.97 p = 0.035 | F1.24 = 4.26 p = 0.049 |
| Carbonate substitution 3 | 0.243 ± 0.004 | 0.222 ± 0.006 ## | 0.223 ± 0.006 # | 0.217 ± 0.002 | ns | F1.24 = 6.30 p = 0.019 | F1.24 = 6.96 p = 0.014 |
| Mineral crystallinity 4 | 0.976 ± 0.005 | 0.957 ± 0.006 # | 0.956 ± 0.005 # | 0.956 ± 0.003 | F1.24 = 4.23 p = 0.05 | F1.24 = 4.21 p = 0.05 | ns |
| Collagen maturity 5 | 2.044 ± 0.021 | 1.848 ± 0.042 ## | 1.692 ± 0.047 ### | 1.373 ± 0.022 *** | ns | F1.24 = 52.84 p < 0.0001 | F1.24 = 136.3 p < 0.0001 |
| wt/wt Male | ko/ko Male | wt/wt Female | ko/ko Female | Interaction (Genotype vs. Sex) | Genotype | Sex | |
|---|---|---|---|---|---|---|---|
| Igfbp3 | 1.00 ± 0.29 | 1.51 ± 0.27 | 0.93 ± 0.19 | 2.63 ± 0.51 ** | ns | F1.24 = 10.54 p = 0.003 | ns |
| Igfbp5 | 1.00 ± 0.18 | 1.23 ± 0.14 | 0.63 ± 0.14 | 0.93 ± 0.15 | ns | ns | F1.24 = 4.64 p = 0.041 |
| Igfals | 1.00 ± 0.03 | 1.06 ± 0.05 | 0.53 ± 0.04 ## | 0.59 ± 0.04 && | ns | ns | F1.24 = 108.6 p < 0.0001 |
| wt/wt Male | ko/ko Male | wt/wt Female | ko/ko Female | Interaction (Genotype vs. Sex) | Genotype | Sex | |
|---|---|---|---|---|---|---|---|
| Col1a1 | 1.00 ± 0.19 | 1.13 ± 0.20 | 1.25 ± 0.20 | 1.65 ± 0.26 | ns | ns | ns |
| Osteopontin | 1.00 ± 0.20 | 0.77 ± 0.15 | 0.64 ± 0.14 | 1.80 ± 0.44 * | F1.24 = 6.72 p = 0.016 | ns | ns |
| Osteocalcin | 1.00 ± 0.09 | 0.90 ± 0.12 | 0.79 ± 0.12 | 1.56 ± 0.18 **/& | F1.24 = 10.15 p = 0.004 | F1.24 = 6.16 p = 0.02 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio, L.; Vargas, A.; Rivera, P.; López-Gambero, A.J.; Tovar, R.; Christians, J.K.; Martín-de-las-Heras, S.; Rodríguez de Fonseca, F.; Chowen, J.A.; Argente, J.; et al. Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency. Int. J. Mol. Sci. 2021, 22, 4048. https://doi.org/10.3390/ijms22084048
Rubio L, Vargas A, Rivera P, López-Gambero AJ, Tovar R, Christians JK, Martín-de-las-Heras S, Rodríguez de Fonseca F, Chowen JA, Argente J, et al. Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency. International Journal of Molecular Sciences. 2021; 22(8):4048. https://doi.org/10.3390/ijms22084048
Chicago/Turabian StyleRubio, Leticia, Antonio Vargas, Patricia Rivera, Antonio J. López-Gambero, Rubén Tovar, Julian K. Christians, Stella Martín-de-las-Heras, Fernando Rodríguez de Fonseca, Julie A. Chowen, Jesús Argente, and et al. 2021. "Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency" International Journal of Molecular Sciences 22, no. 8: 4048. https://doi.org/10.3390/ijms22084048
APA StyleRubio, L., Vargas, A., Rivera, P., López-Gambero, A. J., Tovar, R., Christians, J. K., Martín-de-las-Heras, S., Rodríguez de Fonseca, F., Chowen, J. A., Argente, J., & Suárez, J. (2021). Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency. International Journal of Molecular Sciences, 22(8), 4048. https://doi.org/10.3390/ijms22084048