
Maternal Heat Stress Alters Expression of Genes Associated with Nutrient Transport Activity and Metabolism in Female Placentae from Mid-Gestating Pigs

 , , , ,
, , , , 
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Fetal and Placental Morphometry
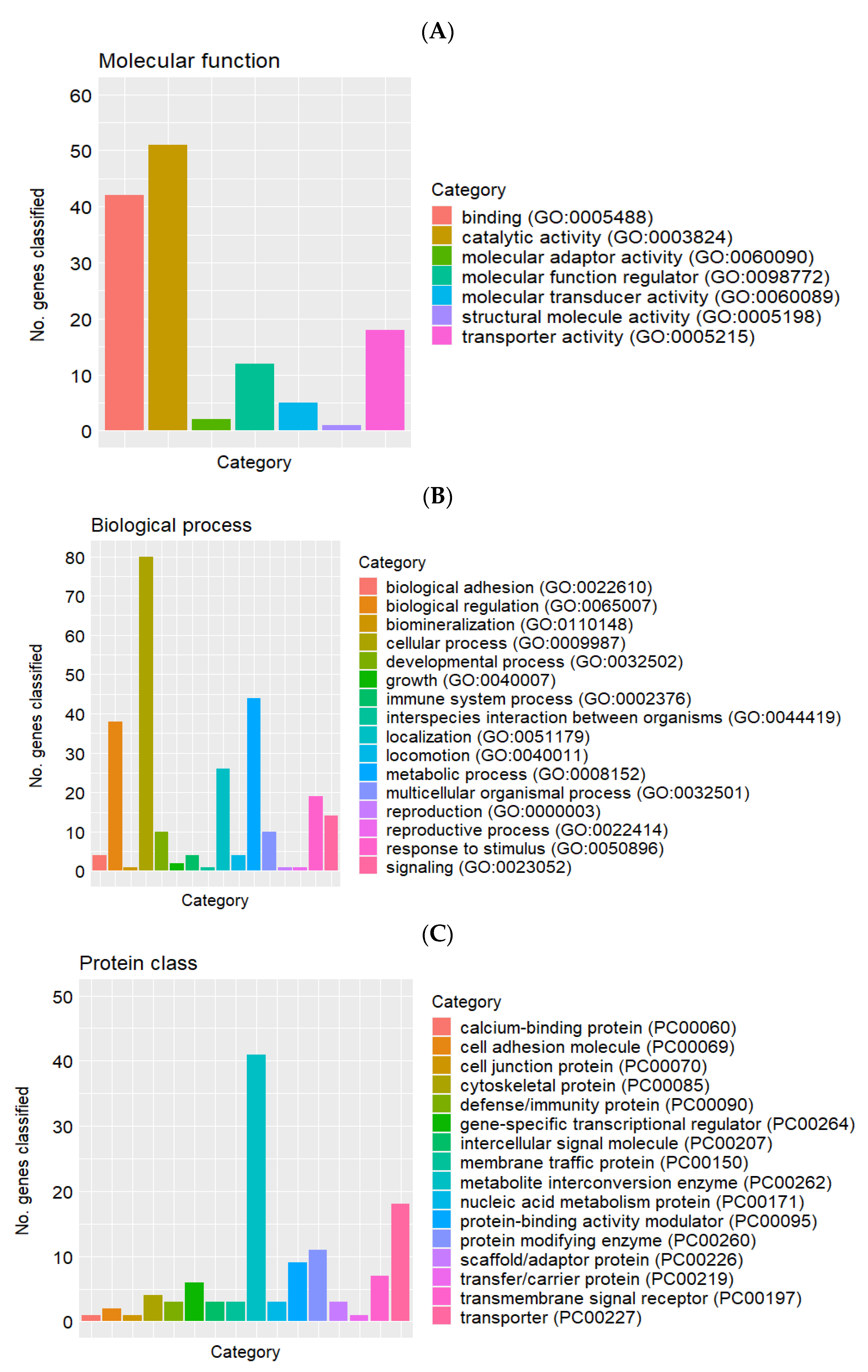
2.2. Differentially Expressed Genes, and Gene Ontology and Pathway Enrichment Analyses
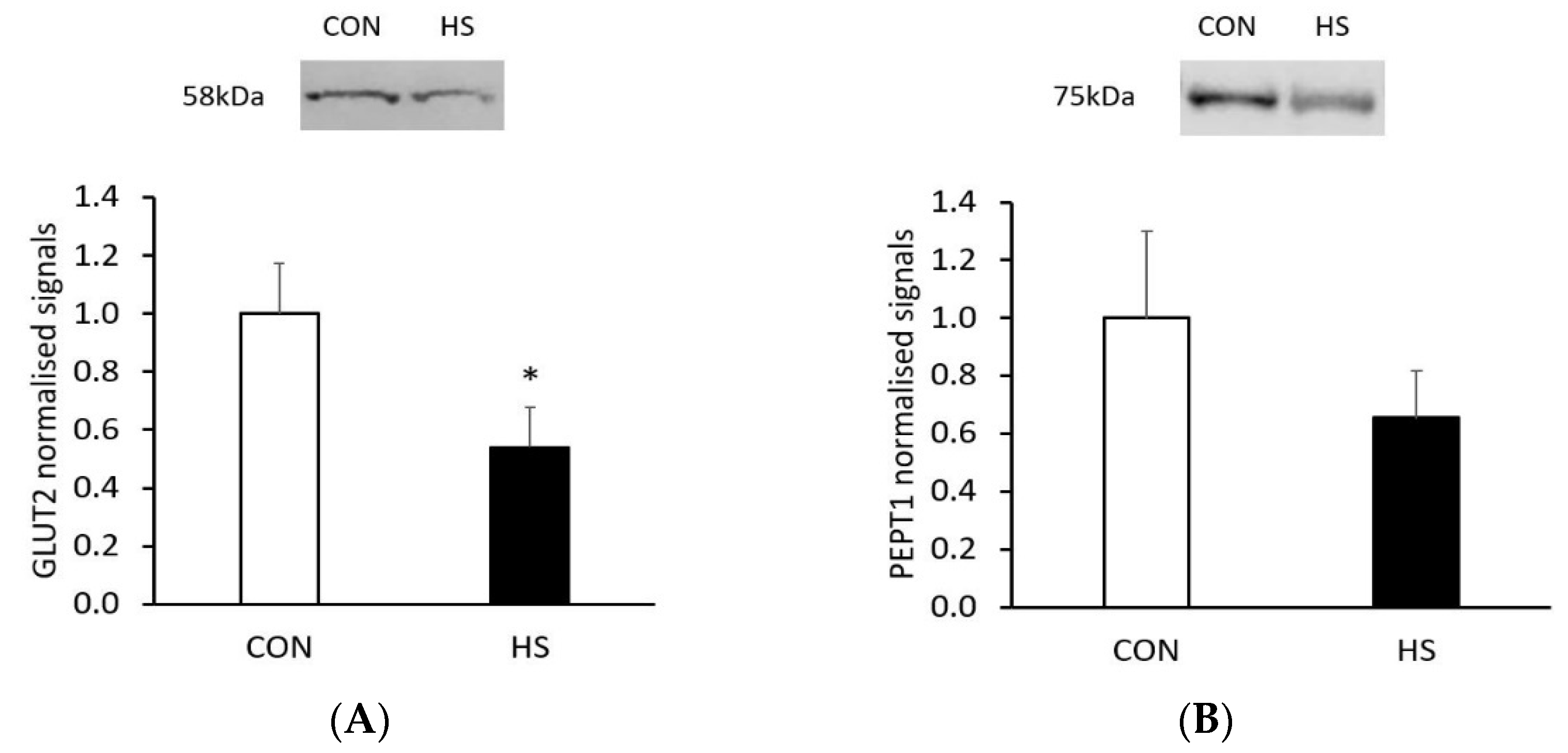
2.3. Western Blot
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethical Approval
5.2. Animals and Experimental Design
5.3. Tissue Collection and Morphometric Analysis
5.4. Total RNA Isolation, Library Preparation, and Sequencing
5.5. Sequencing Data Reprocessing
5.6. Differential Gene Expression, and Gene Ontology (Go) and Pathway Enrichment Analyses
5.7. Western Blot Analysis
5.8. Statistics for Placental and Fetal Morphology, and Western Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M.E.; Biensen, N.J.; Ford, S.P. Novel insight into the control of litter size in pigs, using placental efficiency as a selection tool. J. Anim. Sci. 1999, 77, 1654–1658. [Google Scholar] [CrossRef] [Green Version]
- Vonnahme, K.A.; Wilson, M.E.; Ford, S.P. Conceptus competition for uterine space: Different strategies exhibited by the Meishan and Yorkshire pig. J. Anim. Sci. 2002, 80, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Wilkening, R.; Meschia, G. Some aspects of placental function in chronically heat-stressed ewes. J. Dev. Physiol. 1987, 9, 17–29. [Google Scholar] [PubMed]
- Bell, A. Prenatal programming of postnatal productivity and health of livestock: A brief review. Aust. J. Exp. Agric. 2006, 46, 725–732. [Google Scholar] [CrossRef]
- Liu, F.; Ford, E.M.; Morrison, R.S.; Brewster, C.J.; Henman, D.J.; Smits, R.J.; Zhao, W.; Cottrell, J.J.; Leury, B.J.; Dunshea, F.R. The Greater Proportion of Born-Light Progeny from Sows Mated in Summer Contributes to Increased Carcass Fatness Observed in Spring. Animals 2020, 10, 2080. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Stewart, K.R.; Safranski, T.J.; Ross, J.W.; Baumgard, L.H. In utero heat stress alters postnatal phenotypes in swine. Theriogenology 2020, 154, 110–119. [Google Scholar] [CrossRef]
- Brown, L.D.; Rozance, P.J.; Thorn, S.R.; Friedman, J.E.; Hay, W.W., Jr. Acute supplementation of amino acids increases net protein accretion in IUGR fetal sheep. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E352–E364. [Google Scholar] [CrossRef] [Green Version]
- Yates, D.T.; Clarke, D.S.; Macko, A.R.; Anderson, M.J.; Shelton, L.A.; Nearing, M.; Allen, R.E.; Rhoads, R.P.; Limesand, S.W. Myoblasts from intrauterine growth-restricted sheep fetuses exhibit intrinsic deficiencies in proliferation that contribute to smaller semitendinosus myofibres. Am. J. Physiol. Endocrinol. Metab. 2014, 592, 3113–3125. [Google Scholar] [CrossRef]
- Dreiling, C.E.; Carman, F.S., III; Brown, D.E. Maternal endocrine and fetal metabolic responses to heat stress. J. Dairy. Sci. 1991, 74, 312–327. [Google Scholar] [CrossRef]
- Ross, J.C.; Fennessey, P.V.; Wilkening, R.B.; Battaglia, F.C.; Meschia, G. Placental transport and fetal utilization of leucine in a model of fetal growth retardation. Am. J. Physiol. Endocrinol. Metab. 1996, 270, E491–E503. [Google Scholar] [CrossRef]
- Anderson, A.H.; Fennessey, P.V.; Meschia, G.; Wilkening, R.B.; Battaglia, F.C. Placental transport of threonine and its utilization in the normal and growth-restricted fetus. Am. J. Physiol. Endocrinol. Metab. 1997, 272, E892–E900. [Google Scholar] [CrossRef] [PubMed]
- Limesand, S.W.; Regnault, T.; Hay, W.W., Jr. Characterization of glucose transporter 8 (GLUT8) in the ovine placenta of normal and growth restricted fetuses. Placenta 2004, 25, 70–77. [Google Scholar] [CrossRef]
- Vallet, J.L.; McNeel, A.K.; Miles, J.R.; Freking, B.A. Placental accommodations for transport and metabolism during intra-uterine crowding in pigs. J. Anim. Sci. Biotechnol. 2014, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Krombeen, S.K.; Bridges, W.C.; Wilson, M.E.; Wilmoth, T.A. Factors contributing to the variation in placental efficiency on days 70, 90, and 110 of gestation in gilts. J. Anim. Sci. 2019, 97, 359–373. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, F.; Bell, A.W.; Le, H.H.; Cottrell, J.J.; Leury, B.J.; Green, M.P.; Dunshea, F.R. Controlled elevated temperatures during early-mid gestation cause placental insufficiency and implications for fetal growth in pregnant pigs. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Hu, J.; Johnson, G.A.; Spencer, T.E. Polyamine synthesis from proline in the developing porcine placenta. Biol. Reprod. 2005, 72, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.; Fernández, E.; Torrents, D.; Estévez, R.; López, C.; Camps, M.; Lloberas, J.; Zorzano, A.; Palacín, M. Identification of a membrane protein, LAT-2, that co-expresses with 4F2 heavy chain, an L-type amino acid transport activity with broad specificity for small and large zwitterionic amino acids. J. Biol. Chem. 1999, 274, 19738–19744. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Aliaga, I.; Wagner, C.A. Regulation and function of the SLC38A3/SNAT3 glutamine transporter. Channels 2016, 10, 440–452. [Google Scholar] [CrossRef]
- Wu, G.; Ott, T.L.; Knabe, D.A.; Bazer, F.W. Amino acid composition of the fetal pig. J. Nutr. 1999, 129, 1031–1038. [Google Scholar] [CrossRef]
- Daniel, H. Molecular and integrative physiology of intestinal peptide transport. Annu. Rev. Physiol. 2004, 66, 361–384. [Google Scholar] [CrossRef]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therien, A.G.; Blostein, R. Mechanisms of sodium pump regulation. Am. J. Physiol. Cell Physiol. 2000, 279, C541–C566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.; Karlsson, L.; Wennergren, M.; Jansson, T.; Powell, T.L. Activity and protein expression of Na+/K+ ATPase are reduced in microvillous syncytiotrophoblast plasma membranes isolated from pregnancies complicated by intrauterine growth restriction. J. Clin. Endocrinol. Metab. 2003, 88, 2831–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mérillat, A.-M.; Charles, R.-P.; Porret, A.; Maillard, M.; Rossier, B.; Beermann, F.; Hummler, E. Conditional gene targeting of the ENaC subunit genes Scnn1b and Scnn1g. Am. J. Physiol. Renal Physiol. 2009, 296, F249–F256. [Google Scholar] [CrossRef] [PubMed]
- Page, K.R.; Ashworth, C.; McArdle, H.; Finch, A.; Nwagwu, M. Sodium transport across the chorioallantoic membrane of porcine placenta involves the epithelial sodium channel (ENaC). J. Physiol. 2003, 547, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell. Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Gambling, L.; Danzeisen, R.; Gair, S.; Lea, R.G.; Charania, Z.; Solanky, N.; Joory, K.D.; Srai, S.K.S.; Mcardle, H.J. Effect of iron deficiency on placental transfer of iron and expression of iron transport proteins in vivo and in vitro. Biochem. J. 2001, 356, 883–889. [Google Scholar] [CrossRef]
- Richards, M.P. Zinc, copper, and iron metabolism during porcine fetal development. Biol. Trace Elem. Res. 1999, 69, 27–44. [Google Scholar] [CrossRef]
- Vallet, J.L.; Miles, J.R.; Freking, B.A. Effect of fetal size on fetal placental hyaluronan and hyaluronoglucosaminidases throughout gestation in the pig. Anim. Reprod. Sci. 2010, 118, 297–309. [Google Scholar] [CrossRef]
- Vallet, J.L.; Freking, B.A. Differences in placental structure during gestation associated with large and small pig fetuses. J. Anim. Sci. 2007, 85, 3267–3275. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Kim, J.; Ka, H.; Johnson, G.A.; Wu, G.; Song, G. Select nutrients in the uterine lumen of sheep and pigs affect conceptus development. J. Reprod. Dev. 2012, 58, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Uldry, M.; Ibberson, M.; Hosokawa, M.; Thorens, B. GLUT2 is a high affinity glucosamine transporter. FEBS Lett. 2002, 524, 199–203. [Google Scholar] [CrossRef]
- Vallet, J.L.; Miles, J.R.; Freking, B.A.; Meyer, S. Glucosamine supplementation during late gestation alters placental development and increases litter size. J. Anim. Sci. BiotechnoL. 2017, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hers, H.-G. Mechanisms of blood glucose homeostasis. J. Inherit. Metab. Dis. 1990, 13, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Song, G.; Wu, G.; Bazer, F.W. Functional roles of fructose. Proc. Natl. Acad. Sci. USA 2012, 109, E1619–E1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krombeen, S.K.; Shankar, V.; Noorai, R.E.; Saski, C.A.; Sharp, J.L.; Wilson, M.E.; Wilmoth, T.A. The identification of differentially expressed genes between extremes of placental efficiency in maternal line gilts on day 95 of gestation. BMC Genom. 2019, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Burns, G.; Brooks, K.; Wildung, M.; Navakanitworakul, R.; Christenson, L.K.; Spencer, T.E. Extracellular vesicles in luminal fluid of the ovine uterus. PLoS ONE 2014, 9, e90913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidarimath, M.; Khalaj, K.; Kridli, R.T.; Kan, F.W.; Koti, M.; Tayade, C. Extracellular vesicle mediated intercellular communication at the porcine maternal-fetal interface: A new paradigm for conceptus-endometrial cross-talk. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Gebremedhn, S.; Gad, A.; Aglan, H.S.; Laurincik, J.; Prochazka, R.; Salilew-Wondim, D.; Hoelker, M.; Schellander, K.; Tesfaye, D. Extracellular vesicles shuttle protective messages against heat stress in bovine granulosa cells. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef]
- Dickinson, H.; Moss, T.; Gatford, K.L.; Moritz, K.M.; Akison, L.; Fullston, T.; Hryciw, D.H.; Maloney, C.; Morris, M.; Wooldridge, A. A review of fundamental principles for animal models of DOHaD research: An Australian perspective. J. Dev. Orig. Health Dis. 2016, 7, 449–472. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic. Acids. Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods. 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic. Acids. Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J.G. Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic. Acids. Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Treatments | p Values | |
|---|---|---|---|
| CON (n = 5) | HS (n = 5) | ||
| Placental weight (g) | 88 ± 6 | 112 ± 12 | 0.053 |
| Fetal weight (g) | 104 ± 7 | 92 ± 4 | 0.08 |
| Placental efficiency (fetal/placental weight) | 1.20 ± 0.09 | 0.87 ± 0.11 | 0.023 |
| Placental surface area (cm2) | 725 ± 59 | 824 ± 31 | 0.09 |
| Data are expressed as means with standard error of the means (SEM). | |||
| Gene Symbol | Full Gene Name | Log2 Fold Change | False Discovery Rate |
|---|---|---|---|
| IGSF5 | immunoglobulin superfamily member 5 | 2.04 | 7 × 10−6 |
| ZNF691 | zinc finger protein 691 | 0.70 | 9 × 10−4 |
| SLC7A10 | solute carrier family 7 member 10 | 0.64 | 4 × 10−3 |
| ABTB1 | ankyrin repeat and BTB domain containing 1 | 0.53 | 7 × 10−3 |
| MRVI1 | murine retrovirus integration site 1 homolog | 0.45 | 7 × 10−5 |
| OSGIN1 | oxidative stress induced growth inhibitor 1 | 0.39 | 2 × 10−2 |
| SLC30A2 | solute carrier family 30 member 2 | 0.34 | 6 × 10−2 |
| SLC26A6 | solute carrier family 26 member 6 | 0.28 | 3 × 10−2 |
| SLC25A25 | solute carrier family 25 member 25 | 0.19 | 7 × 10−2 |
| GPI | glucose-6-phosphate isomerase | −0.16 | 7 × 10−2 |
| ATP1A1 | ATPase Na+/K+ transporting subunit alpha 1 | −0.19 | 8 × 10−2 |
| GPRC5C | G protein-coupled receptor class C group 5 member C | −0.22 | 5 × 10−2 |
| SLC11A2 | solute carrier family 11 member 2 | −0.24 | 9 × 10−2 |
| SLC8B1 | solute carrier family 8 member B1 | −0.24 | 8 × 10−2 |
| SCNN1G | sodium channel epithelial 1 gamma subunit | −0.26 | 6 × 10−2 |
| SLC20A1 | solute carrier family 20 member 1 | −0.28 | 4× 10−2 |
| SLC40A1 | solute carrier family 40 member 1 | −0.32 | 2 × 10−2 |
| ATP8B1 | ATPase phospholipid transporting 8B1 | −0.32 | 4 × 10−2 |
| DEFB1 | defensin beta 1 | −0.41 | 7 × 10−2 |
| IGFBP5 | insulin-like growth factor-binding protein 5 | −0.45 | 6 × 10−2 |
| ATP13A3 | ATPase 13A3 | -0.60 | 1 × 10−2 |
| ST3GAL1 | ST3 beta-galactoside alpha-2,3-sialyltransferase 1 | −0.60 | 2 × 10−3 |
| SLC2A2 | solute carrier family 2 member 2 | −0.62 | 1 × 10−2 |
| B3GNT7 | betaGal beta-1,3-N-acetylglucosaminyltransferase 7 | −0.73 | 6 × 10−7 |
| ST3GAL3 | ST3 beta-galactoside alpha-2,3-sialyltransferase 3 | −0.75 | 1 × 10−3 |
| SLC38A3 | solute carrier family 38 member 3 | −0.76 | 2 × 10−2 |
| FABP5 | fatty acid binding protein 5 | −0.79 | 8 × 10−4 |
| ANXA1 | annexin A1 | −0.81 | 8× 10−4 |
| SLC7A8 | solute carrier family 7 member 8 | −0.87 | 7 × 10−4 |
| CCN2 | cellular communication network factor 2 | −0.88 | 5 × 10−4 |
| ECM1 | extracellular matrix protein 1 | −1.00 | 2 × 10−4 |
| PTGER4 | prostaglandin E receptor 4 | −1.15 | 2 × 10−3 |
| KCNK5 | potassium two pore domain channel subfamily K member 5 | −1.20 | 1 × 10−5 |
| SLC45A3 | solute carrier family 45 member 3 | −1.37 | 6 × 10−8 |
| FBP2 | fructose-bisphosphatase 2 | −1.44 | 5 × 10−4 |
| SPINK4 | serine peptidase inhibitor, Kazal type 4 | −1.45 | 1× 10−8 |
| SLC15A1 | solute carrier family 15 member 1 | −1.46 | 1 × 10−13 |
| IRX3 | iroquois homeobox 3 | −1.50 | 3 × 10−12 |
| LHFPL4 | LHFPL tetraspan subfamily member 4 | −1.60 | 1 × 10−6 |
| CHST8 | carbohydrate sulfotransferase 8 | −1.75 | 3 × 10−7 |
| Pathway Database | Term | Pathway ID | FDR (False Discovery Rate) | Number of Genes Enriched |
|---|---|---|---|---|
| Reactome | Lysosomal oligosaccharide catabolism | R-SSC-8853383 | 0.042 | 2 |
| * Metabolism of carbohydrates | R-SSC-71387 | 0.000 | 10 | |
| Keratan sulfate biosynthesis | R-SSC-2022854 | 0.007 | 4 | |
| * Glycosaminoglycan metabolism | R-SSC-1630316 | 0.008 | 6 | |
| Sialic acid metabolism | R-SSC-4085001 | 0.009 | 4 | |
| * Synthesis of substrates in N-glycan biosynthesis | R-SSC-446219 | 0.043 | 4 | |
| O-linked glycosylation of mucins | R-SSC-913709 | 0.043 | 4 | |
| * O-linked glycosylation | R-SSC-5173105 | 0.044 | 5 | |
| Transport of inorganic cations/anions and amino acids/oligopeptides | R-SSC-425393 | 0.039 | 5 | |
| * SLC-mediated transmembrane transport | R-SSC-425407 | 0.008 | 9 | |
| KEGG | Metabolic pathways | KEGG:01100 | 0.001 | 31 |
| Lysosome | KEGG:04142 | 0.002 | 8 | |
| Glycosaminoglycan biosynthesis—keratan sulfate | KEGG:00533 | 0.027 | 3 | |
| Glycosphingolipid biosynthesis—ganglio series | KEGG:00604 | 0.034 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Liu, F.; Marth, C.D.; Green, M.P.; Le, H.H.; Leury, B.J.; Bell, A.W.; Dunshea, F.R.; Cottrell, J.J. Maternal Heat Stress Alters Expression of Genes Associated with Nutrient Transport Activity and Metabolism in Female Placentae from Mid-Gestating Pigs. Int. J. Mol. Sci. 2021, 22, 4147. https://doi.org/10.3390/ijms22084147
Zhao W, Liu F, Marth CD, Green MP, Le HH, Leury BJ, Bell AW, Dunshea FR, Cottrell JJ. Maternal Heat Stress Alters Expression of Genes Associated with Nutrient Transport Activity and Metabolism in Female Placentae from Mid-Gestating Pigs. International Journal of Molecular Sciences. 2021; 22(8):4147. https://doi.org/10.3390/ijms22084147
Chicago/Turabian StyleZhao, Weicheng, Fan Liu, Christina D. Marth, Mark P. Green, Hieu H. Le, Brian J. Leury, Alan W. Bell, Frank R. Dunshea, and Jeremy J. Cottrell. 2021. "Maternal Heat Stress Alters Expression of Genes Associated with Nutrient Transport Activity and Metabolism in Female Placentae from Mid-Gestating Pigs" International Journal of Molecular Sciences 22, no. 8: 4147. https://doi.org/10.3390/ijms22084147
APA StyleZhao, W., Liu, F., Marth, C. D., Green, M. P., Le, H. H., Leury, B. J., Bell, A. W., Dunshea, F. R., & Cottrell, J. J. (2021). Maternal Heat Stress Alters Expression of Genes Associated with Nutrient Transport Activity and Metabolism in Female Placentae from Mid-Gestating Pigs. International Journal of Molecular Sciences, 22(8), 4147. https://doi.org/10.3390/ijms22084147